
Abstract
Potato Cyst Nematodes (PCN) are a globally important and economically extremely damaging potato pest. Chemical control has limited impact and cannot clear a field of PCN, but genetic resistance in potatoes is an effective and important part of natural management of PCN. There are two closely related species of PCN; Globodera rostochiensis against which the H1 resistance gene from S. tuberosum ssp. andigena (CPC 1673) has proven effective in Europe for several decades, and G. pallida for which potato varieties with resistance are also commercially available. However, G. pallida resistant genotypes are mainly limited to processing varieties and most varieties grown are still susceptible to the species. The limited number of suitable varieties with resistance against G. pallida has led to its spread throughout the UK and now infestations in Scottish seed land are threatening the future of potato seed production. Novel potato genotypes with resistance against G. pallida sometimes combine PCN resistances from more than one parental source. Multiplication of a panel of G. pallida populations which differ in virulence against host species were compared on recently developed breeding lines with single or stacked resistance genes. Potato genotypes with multiple resistance genes show reduced multiplication in G. pallida pathotypes that were able to overcome a single resistance gene.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The potato cyst nematodes (PCN) Globodera rostochiensis and G. pallida are sedentary endoparasites of one of the world’s most important staple food crops, potatoes. Originating in South America, PCN can now be found in many potato growing regions across the globe. Current management costs and yield losses associated with PCN are estimated to be around £31 million in the UK alone (Price et al., 2021).
Control of PCN most often relies on integrated pest management (IPM) practices. This includes a combination of cultural practices such as use of certified seed stock, chemical controls, and use of resistant potato varieties. No chemicals exist specifically for control of PCN. Instead, generic nematicides are used which, due to their modes of action and environmental concerns, are becoming increasingly restricted in most markets. Host resistance remains the best tool for managing PCN. The H1 resistance gene from Solanum tuberosum ssp. andigena (CPC 1673) has proven to be durable for more than 50 years against G. rostochiensis. However, widespread deployment of this resistance has led to an increase in prevalence of G. pallida in the UK (Minnis et al., 2002). It has been more difficult to achieve high levels of resistance against G. pallida and it is expected that resistance to G. pallida may not be as durable as H1 due to increased variability in virulence and genetic diversity within G. pallida populations present in the UK (Hockland et al., 2012; Phillips & Trudgill, 1998).
The majority of PCN movement is through passive transport of cysts with mobile juveniles not expected to move more than 1 m in soil (EPPO, 2018). Populations that remain localised to one area and exposed to one source of resistance pose a threat to resistance. Repeat exposure to the same resistance allows selection of the few individuals that may be able to develop into females on resistant varieties. Breakdown of resistance in the cultivar Innovator, which contains resistance from S. vernei, by G. pallida populations has been reported in Germany (Mwangi et al., 2019; Niere et al., 2014). Further resistance breakdown is evident in the Netherlands with both occurrences resulting from a single adaptation event (Grenier et al., 2020). Increased genetic diversity in G. pallida populations can be represented by differences in pathotype and virulence. Selection for increased virulence was demonstrated after multiple rounds of multiplication on resistances from S. vernei (source of GpaV resistance) and S. tuberosum ssp. andigena CPC 2802 (source of H3 resistance) (Phillips & Blok, 2008). More recently, Varypatakis et al., (2019, 2020) demonstrated that selection for virulence was specific for the source against which the population was originally selected. This work also identified regions of the G. pallida genome that were impacted by this selection pressure. Poor stewardship of resistant varieties may therefore lead to increased incidence of more virulent populations that break resistance. Methods of reducing the selection for virulent populations are therefore needed.
One strategy to improve durability of resistance of potato against G. pallida is to combine more than one source of resistance. Rigney et al. (2017) found there was an additive effect on host resistance when two quantitative trait loci (QTLs) from S. vernei (GpaV) and S. tuberosum ssp. andigena (GpaIVsadg) were combined, or stacked, which was consistent when tested with several G. pallida populations. This approach requires molecular markers for each resistance source to demonstrate successful introgression in a breeding programme. However, while markers provide a powerful tool for genotype analysis, resistance from sources such as S. vernei can be polygenic (Dale & Phillips, 1982) and therefore phenotype analysis continues to be essential for progressing potato breeding material to market. PCN are not the only pathogens infecting potatoes and production of new resistant material must also consider numerous other pathogenic challenges. Incorporation of commercially required traits further complicates development of new PCN resistant potato genotypes. While a broad spectrum of resistances is useful in all varieties it is also important to note that different combinations of tuber shape, size and dry matter will make different genotypes preferentially suited to consumer or processing markets. Therefore, breeding companies benefit from having a variety of parental material suited to these different markets when planning genotype crossing programmes.
Here we consider the effect of stacking 2 key G. pallida resistance genes from different sources to combat infection by G. pallida populations differing in virulence. Specifically, whether the presence of more resistance genes confers added protection against PCN populations that could otherwise overcome a single source of host resistance. Additionally, we highlight whether stacking resistance genes that provide resistance to G. rostochiensis has any detrimental effects on the resistance against G. pallida. The efficacy of some resistance genes varied between nematode populations and where they are effective, stacking genes increases resistance. Unlike the study from Rigney et al. (2017), G. pallida populations were screened against potato genotypes from more than one segregating population to determine whether the same stacked resistance genes resulting from different parental material provide the same additive control against PCN.
Materials and methods
Globodera pallida populations
Globodera pallida populations were taken from the James Hutton Institute population collection. Population Chavornay from Switzerland is the European standard specified in EU Directive 2007/33/EC. Luffness originates from Scotland and has high virulence to the differential 62.33.3 used in the Kort et al. (1977) pathotype scheme (Phillips & Trudgill, 1998) while population Lindley is a “standard” UK population that has been used for many years to assess resistance in new breeding material by James Hutton Limited (JHL). Populations preceded by “N” were produced following multiple rounds of multiplication of the population Newton on either Solanum vernei (N8906 and N11305) or S. tuberosum ssp. andigena CPC 2802 (N11415 and N12674) as described in Phillips and Blok (2008). Population virulence is directly associated with host resistance scored against the cultivar Morag as the least resistant tested cultivar from the S. vernei group (Phillips & Trudgill, 1998). In assays virulence scores per population are, Luffness (87.4%); Chavornay (26.9%); Lindley (12.5%); N8906 (33.0%); N11305 (33.0%); N11415 (33.0%); N12674 (33.0%). Newton subpopulations are assumed to be less than or equal to the main Newton population scored by Phillips and Trudgill (1998).
Potato genotypes
Genotypes were produced from targeted crossing between 2009–2018. Plants were grown from true botanical seed after the initial crossing and were subsequently clonally propagated. PCN resistance gene markers were used to select genotypes that had inherited the required genes. Genomic DNA was extracted using a modified cetyltrimethylammonium bromide (CTAB) procedure from Doyle and Doyle (1987). The presence of resistance genes was inferred through molecular markers, using primers and protocols as published in Finkers-Tomczak et al., (2011) for 57R representing the H1 gene; Sattarzadeh et al., (2006) for HC representing resistance from GpaV; and an unpublished marker ‘H3’, representing the H3 resistance gene. Marker presence/absence in different potato genotypes and parental material for each of the crosses can be seen in Online Resource 3. The majority of G. pallida resistance used here originates from commercial varieties already available in the UK, for example, Innovator, Vales Everest and Ambassador. It is assumed that the presence of H1 will have no effect on G. pallida.
Potato genotypes were grown to renew tuber stock annually. Tubers were stored at 4 °C until use.
G lobodera pallida multiplication assays
Assays were performed twice in 2019, once in 2020 and once in 2021. Deep Rootrainers (Haxnicks) were filled 2/3 with peat-based compost, 20 ± 2 cysts added and then the remaining 1/3 of the Rootrainer filled with compost. A single sprout was removed from a tuber with a melon scoop and placed sprout down on the compost (Rigney et al., 2017). Each population of G. pallida was treated separately in a completely randomised design with 4 blocks. Rootrainers were arranged in trays which accommodated 32 plants and, depending on the number of genotypes tested, each genotype was present in 3 or 4 trays. These were arranged on a glasshouse bench with 16 h daytime temperature of 20 °C (± 4 °C), 8 h of 16 °C (± 4 °C) night-time, with additional lighting as required. The plants were watered and fertilized as appropriate. After 8–9 weeks the Rootrainers were opened and the number of G. pallida females visible on the root surface counted as an indicator for population multiplication.
Statistical analyses
Each G. pallida population was treated as a separate experiment as they were set up in blocks of single populations in the glass house. A generalised linear mixed model approach with a Poisson distribution and log link was used to model the data. The significance of fixed effects was assessed using Wald Chi-square tests to allow for the Poisson distributed residuals. Diagnostic plots were examined for outliers and patterns within the residuals. Each G. pallida population was analysed separately, and random effects fitted for, assay, block within assay and Rootrainer within block to take account of positional variation which may have led to varying performance of strains between years. Expected mean counts of cysts for each potato genotype were assessed by fitting genotype as a fixed effect. A model including fixed effects for the presence of each resistance gene including two-way interactions was then fitted. No variety was included with only the H1 resistance gene and only one genotype having all three resistance genes was present so there are insufficient degrees of freedom to include the three-way interaction. The main effects allow for the assessment of the efficacy of each gene independently. No significant interaction terms would imply that the genes operate additively and independently, a significant positive interaction would imply a multiplicative effect and significant negative interaction would imply that the resistance conferred is not independent and that there is some redundancy in the protection conferred. Analyses were carried out in R 4.2.2 (R Core Team, 2022) using packages lme4 1.1–21 (Bates et al., 2015) and car 3.1–1 (Fox & Weisberg, 2019). The data and code for the analyses are available in Online Resource 1 and Online Resource 2 respectively.
Results
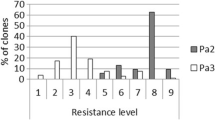
Desiree and Gemson contain no known G. pallida resistance genes, and these varieties allowed G. pallida to multiply the most. Although both Ambassador and Innovator contain GpaV from the same genetic pedigree, Ambassador was more susceptible to all populations of G. pallida. Comparing a panel of G. pallida populations against a range of commercial lines showed the difference in virulence between populations. Newton subpopulations N11415 and N12674 were created following repeated exposure to H3 resistance. These populations therefore had higher reproduction on both Vales Everest and Royal. Although N8906 and N11305 were raised on exposure to GpaV, they had relatively little multiplication on these lines here. Oddly, N12674 proved to be particularly virulent multiplying well on the most resistant commercial variety present, Innovator, representing a source of resistance that this population had not previously been exposed to (Fig. 1). Figure 1 represents data from 4 repeat experiments across 3 years with 4 replicates per variety in each experiment. Each G. pallida population was treated as a separate observation due to experimental design using blocks of single populations in the glasshouse. Female count data was pooled across all years of the trial and expected mean counts of females for each potato genotype were evaluated.

Multiplication of G. pallida populations differing in virulence against commercially available potato genotypes. Female counts represent all assays across all years of the experiment. Resistance score (RS) against G. pallida pa2/3, 1 as available on the Potato Variety Database (https://potatoes.agricrops.org/) and resistance genes (RG) have been included in the table. The error bars represent the standard error of the expected mean number of cysts, large asymmetric bars at high expected counts are a feature of count data. There is a clear difference between population multiplication in genotypes with no RG, H3 and GpaV
Assessing the response of G. pallida populations in the presence of different sources of resistance highlights population virulence against certain resistance genes (Fig. 2). Resistance provided by H3 is extremely variable in the ability to provide control against different G. pallida populations. Luffness and N8906 were controlled well by this resistance. However, H3 had less of an effect on Chavornay, N11415 and N12674 which mostly maintained or increased their population sizes. Resistance from GpaV was generally stronger than resistance from H3 with fewer populations being able to maintain initial population sizes in the presence of GpaV. Genotype 15.JHL.131.A 1 was not positive for the presence of any resistance markers but did not allow multiplication of G. pallida populations (Online Resource 1), suggesting that other minor resistance genes may have been inherited or that there is a discrepancy between marker presence and resistance.

Predicted mean G. pallida final population (pf) following growth of populations differing in virulence against different potato genotypes. Genotypes that also include the H1 gene for G. rostochiensis resistance are shown with a red cross. Black triangles indicate genotypes without H1. Female counts represent all assays across all years of the experiment. Stacking resistance from both GpaV and H3 reduced all populations regardless of virulence
Critically, all populations were prevented from multiplying on any potato genotypes stacking resistance from H3 and GpaV. As expected, addition of the H1 gene for resistance against G. rostochiensis to either H3 or GpaV did not confer additional G. pallida resistance.
Using the data from Fig. 2, a model was developed to better understand the impact of individual and combined resistance sources on PCN reproduction (Fig. 3).

Poisson model showing predicted population fold change after exposure to different sources of resistance. A fold change of 1 is expected where resistance genes have no effect on reducing the PCN population. Newton populations selected on either GpaV or H3 are marked with triangles
When the number of resistance genes was considered, without distinguishing between which genes those were, the best model used a linear response as opposed to treating the number of R-genes as a factor. Though not always significant, the estimated effect of more resistance genes was always positive.
When considering the resistance genes separately, the cause of the differences in populations becomes clear. As seen before, GpaV provided the strongest protection against all PCN populations. H3 provided protection against the Lindley, Luffness, N8906 and N11305 populations, but not against Chavornay, Newton 11,415 or Newton 12,674 populations. As expected, H1 provided no protection against any G. pallida populations, the use of this independent factor helps validate the model. The effects of stacking genes were additive, with no interaction between the genes. This is consistent with the result seen above, for populations where H3 is effective, stacking effects were evident and H3 is acting independently of GpaV. The effect is masked in N8906 because genotypes with the H1 gene showed a highly variable response to N8906. Though not significant, there was a mean increase in cyst counts. Importantly, there is no detrimental effect seen when stacking H1 resistance with G. pallida resistance genes. Fold change in the expected mean cyst counts associated with the presence of genes GpaV, H3 and H1 are available in Online Resource 4.
Discussion
In the UK, and much of Europe, PCN is often used to describe either G. rostochiensis or G. pallida at the species level. Testing a panel of G. pallida populations against different sources of resistance highlights that for effective control of PCN through use of host resistance, improved population diagnostics are necessary.
The panel of G. pallida populations used to assess resistance of different genotypes consisted of 7 populations that had previously been shown to differ in their virulence. When tested against many potato genotypes it is clear that virulence and pathotypes are not something that can be scored and applied across a broad range of hosts. However, there are clear differences in the ability of separate populations to reproduce on different hosts.
Host resistance against PCN is currently the best way of reducing PCN populations. Breakdown of resistance is therefore a major concern. Long crop rotations remove availability of a host for several years, reducing generational turnover. However, breakdown of resistance by G. pallida populations has been reported in Germany (Mwangi et al., 2019) and the Netherlands (Grenier et al., 2020). In both cases the ability to overcome resistance was due to adaptation and not a novel introduction to Europe. Intensive use of resistant varieties coupled with equipment sharing between Germany and the Netherlands likely caused and spread these populations (Grenier et al., 2020). To replicate increased virulence against a source of G. pallida resistance, populations that were raised in the presence of certain resistance genes were used. These populations are proven to have become genetically adapted to overcome resistance, replicating the process observed in the virulent German and Dutch populations. Here, Newton subpopulations raised against H3 (N11415 and N12674) were able to overcome H3 resistance. These populations were less virulent against GpaV. Stacking both H3 and GpaV resistances reduced multiplication of this H3-breaking population. Newton subpopulations N8906 and N11305 were both created by selection on GpaV. As shown by the model, GpaV proved to be a stronger source of resistance compared to H3. Consequently, these populations, which were bred to overcome GpaV, struggled to maintain their population size in the presence of this resistance. Again, genotypes stacking GpaV and H3 were nonetheless capable of reducing multiplication of these populations. In an applied setting, this data importantly confirms that GpaV and H3 work independently of each other. Therefore, where resistance-breaking PCN populations do occur, potato genotypes with stacked resistance genes can be used for an additive effect on population reduction.
In Europe, the genetic diversity in populations of G. rostochiensis is significantly reduced compared to that of G. pallida (reviewed: Price et al., 2021). Therefore, strong resistance against G. rostochiensis is provided by the H1 gene. This gene has remained durable for over 70 years in the UK. However, widespread use of H1 selected for populations of G. pallida in much of the UK making it the now predominant species. Importantly, our model highlights that there is no detrimental effect on the G. pallida resistance conveyed by GpaV or H3 when stacked with H1. This model provides reassurance that if dual species PCN resistance is stacked within the same potato genotype then there will be no selection for either species.
Genotypes showing stacked resistance against G. pallida and with low G. pallida multiplication values offer excellent parental material for future crosses with many of the genotypes in this study showing high levels of resistance to all nematode populations in the panel. A significant drawback of relying solely on molecular markers for identification of the presence of resistance genes is the discrepancy seen between marker and resistance phenotype. In the data presented here, the usually polygenic GpaV resistance from S. vernei was generally indicated well by the HC marker. However, the irregularity between marker and resistance phenotype was visible in line 18.WC.6.A.36. Genotypes that molecular marker results indicate have stacked G. pallida resistance from both H3 and GpaV (12.Z.134.b.25, 13.P.7.A.4, 15.JHL.137.A.4, 15.JHL.137.A.5, 15.JHL.140.A.1 and 15.JHL.140.A.5) all reduced PCN population sizes, regardless of the population virulence against one specific source of resistance. Importantly, these crosses result from different parental material, therefore as long as resistance genes are present, PCN control remains constant. Practically, where growers are relying on commercial varieties with PCN resistance, this work signifies the importance of either rotating the source of resistance used, or preferentially, using genotypes with stacked PCN resistance.
Data Availability
All data generated or analysed during this study are included in this published article [and its supplementary information files].
References
Bates, D., Maechler, M., Bolker, B., and Walker, S., (2015) Fitting linear mixed-effects models using lme4. J Stat Soft, 67(1) 1–48. https://doi.org/10.18637/jss.v067.i01
Dale, M., & Phillips, M. (1982). An investigation of resistance to the white potato cyst-nematode. Journal of Agricultural Science, 99(2), 325–328. https://doi.org/10.1017/S0021859600030094
Doyle, J. J., & Doyle, J. L. (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin, 19(1), 11–15.
EPPO. (2018). PM 9/26 (1) National regulatory control system for Globodera pallida and Globodera rostochiensis. OEPP/EPPO Bulletin, 48(3), 516–532. https://doi.org/10.1111/epp.12510
Finkers-Tomczak, A., Bakker, E., de Boer, J., van der Vossen, E., Achenback, U., Goals, T., et al. (2011). Comparative sequence analysis of the potato cyst nematode resistance locus H1 reveals a major lack of co-linearity between three haplotypes in potato (Solanum tuberosum ssp.). TAG. Theoretical and Applied Genetics., 122, 595–608. https://doi.org/10.1007/s00122-010-1472-9
Fox, J. and Weisberg, S. (2019) An R companion to applied regression. Third edition. Thousand Oaks CA: Sage. https://socialsciences.mcmaster.ca/jfox/Books/Companion/
Grenier, E., Kiewnick, S., Smant, G., Fournet, S., Montarry, J., Holterman, M., et al., (2020) Monitoring and tackling genetic selection in the potato cyst nematode Globodera pallida. EFSA. 17(6). https://doi.org/10.2903/sp.efsa.2020.EN-1874
Hockland, S., Niere, B., Grenier, E., Blok, V., Phillips, M., den Nijs, L., et al. (2012). An evaluation of the implications of virulence in non-European populations of Globodera pallida and G. rostochiensis for potato cultivation in Europe. Nematology, 14, 1–13. https://doi.org/10.1163/138855411X587112
Kort, J., Ross, H., Rumpenhorst, H. J., & Stone, A. R. (1977). International scheme for identifying and classifying pathotypes of potato cyst nematodes Globodera rostochiensis and G. pallida. Nematologica, 23, 333–339. https://doi.org/10.1163/187529277x00057
Minnis, S. T., Haydock, P. P. J., Ibrahim, S. K., Grove, I. G., Evans, K., & Russell, M. D. (2002). Potato cyst nematodes in England and Wales – occurrence and distribution. The Annals of Applied Biology, 140, 187–195. https://doi.org/10.1111/j.1744-7348.2002.tb00172.x
Mwangi, J.M., Niere, B., Finckh, M.R., Krussel, S. & Kiewnick, S. (2019) Reproduction and life history traits of a resistance breaking Globodera pallida population. Nematology, 52, 1–13. https://doi.org/10.21307/jofnem-2019-079
Niere, B., Krüssel, S., & Osmers, K. (2014). Auftreten einer außergewöhnlich virulenten Population der Kartoffelzystennematoden. J. Kulturpflanzen, 66, 426–427.
Phillips, M. S., & Blok, V. C. (2008). Selection for reproductive ability in Globodera pallida populations in relation to quantitative resistance from Solanum vernei and S. tuberosum ssp. andigena CPC2802. Plant Pathology, 57, 573–580. https://doi.org/10.1111/j.1365-3059.2007.01771.x
Phillips, M. S., & Trudgill, D. L. (1998). Variation of Virulence, in Terms of Quantitative Reproduction of Globodera pallida Populations, From Europe and South America, in Relation to Resistance from Solanum vernei and S. tuberosum ssp. andigena CPC 2802. Nematologica, 44, 409–423.
Price, J.A., Coyne, D., Blok, V.C. & Jones, J.T. (2021) Potato cyst nematodes Globodera rostochiensis and G. pallida. Mol. Plant Pathol., 22(5), 495–507. https://doi.org/10.1111/mpp.13047
R Core Team (2022) R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. https://www.r-project.org
Rigney, B., Blok, V., Griffin, D., Dalton, E., & Milbourne, D. (2017). Consistent action of two partially effective loci conferring resistance to Globodera pallida Pa2/3 across multiple nematode field populations. Plant Pathology, 66, 1031–1040. https://doi.org/10.1111/ppa.12640
Sattarzadeh, A., Achenbach, U., Lubeck, J., Strahwald, J., Tacke, E., Hofferbert, H.-R., et al. (2006). Single nucleotide polymorphism (SNP) genotyping as basis for developing a PCR-based marker highly diagnostic for potato varieties with high resistance to Globodera pallida pathotype Pa2/3. Molecular Breeding, 18, 301–312. https://doi.org/10.1007/s11032-006-9026-1
Varypatakis, K., Jones, J. T., & Blok, V. C. (2019). Screening of populations of Globodera pallida selected for increased virulence on several potato varieties. Nematology, 21, 995–998. https://doi.org/10.1163/15685411-00003283
Varypatakis, K., Véronneau, P.-Y., Thorpe, P., Cock, P. J. A., Lim, J.T.-Y., Armstrong, M. R., Janakowski, S., Sobczak, M., Hein, I., Mimee, B., Jones, J. T., & Blok, V. C. (2020). The Genomic Impact of Selection for Virulence against Resistance in the Potato Cyst Nematode. Globodera Pallida. Genes., 11(12), 1429. https://doi.org/10.3390/genes11121429
Acknowledgements
This work was funded by The Rural and Environmental Science and Analytical Services (RESAS) division of the Scottish Government. The funders had no role in study design, data collection and analysis, decision to publish or manuscript preparation. The authors would like to thank Xinwei Chen, Jane Yildiz, Linden Forster, Anne Holt, glasshouse and field staff for their help across genotype production, maintenance and PCN population rejuvenation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Vivian C. Blok is Deceased.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Price, J., Preedy, K., Young, V. et al. Stacking host resistance genes to control Globodera pallida populations with different virulence. Eur J Plant Pathol 168, 373–381 (2024). https://doi.org/10.1007/s10658-023-02761-5
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10658-023-02761-5