
Abstract
Stock assessors commonly use models that incorporate biological data such as fish length and age to determine the status of fish stocks and how different management scenarios affect stock size. Ages used in assessment models need to be accurate and precise because ageing error can affect many model inputs and potentially result in stock mismanagement. Despite the requirement for sound age data, few studies have investigated accuracy of ages for groundfish captured in coastal nearshore waters off Oregon, USA. In this study, we validate otolith ages for Black Rockfish (Sebastes melanops), Copper Rockfish (Sebastes caurinus) and Cabezon (Scorpaenichthys marmoratus), three species with recreational and commercial importance to Oregon. Ages obtained by traditional break-and-burn methods were validated using secondary ion mass spectrometry (SIMS) to examine otolith stable oxygen isotope ratios (δ18O) over a fish’s lifetime. This technique relies on the inverse relationship that exists between otolith δ18O and ambient water temperature, and independent counts of δ18O maxima should be comparable to ages obtained by visual counts of otolith growth marks laid down during cold water periods. Locations of δ18O maxima in otolith chronologies matched well with locations of visual growth marks in otoliths of all three species, maxima counts were strongly positively correlated with age, and variation in otolith δ18O decreased with age. However, significant variability in the δ18O chronologies caused by variability in intra-seasonal upwelling and resulting water temperature variations made maxima counts difficult in several samples. Correct interpretation of chronologies required knowledge of location of the first annulus, the compression of growth zones with age, and an assumption of the seasonal amplitude of the ambient water δ18O.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Stock assessments are valuable tools that enable fisheries managers to determine past and current status of fish stocks and allow for predictions of how current and future management scenarios affect population dynamics. These data-intensive, statistically robust assessment models often incorporate three general types of data: (1) catch and effort data—the number of fish removed from a stock by fishing; (2) abundance data—a measure of the number of fish in a stock; and (3) sampled biological data—which include fish lengths and weights, ages, reproductive rates, genetics and information on fish movements (Hilborn and Walters 1992). Age-structured stock assessment models are the most commonly used; therefore, a stock’s age composition and the modeled relationship between fish length and age are the most valuable biological metrics for the models (Wallace and Fletcher 1997). Generally, annual ages are obtained by counting growth marks on calcified structures from a fish, such as scales, fin rays and most commonly, otoliths (see Panfili et al. 2002 for a comprehensive description of hard parts used in age estimation, as well as data use and application). Errors in the estimation of age from these structures are especially problematic because they can affect all model input data including catch and stock weights-at-age, length and maturity-at-age, growth rates, and any age-structured CPUE indices (Reeves 2003); therefore, ages estimated for stock assessments need to be accurate and precise.
Otoliths are calcareous structures found in the inner ear systems of teleost fishes that are formed extracellularly by the precipitation of calcium carbonate (usually as aragonite in sagittae) over a protein matrix (Campana 1999). Otoliths accrete by deposition of concentric layers of these materials around a core, producing periodic growth increments with differing optical densities that researchers count to estimate fish age (Wright et al. 2002). In addition to providing an indication of age, microchemical examination of an otolith’s structure can lend insights into the physiological and nutritional status of a fish, as well as past environmental conditions that the fish experienced (Sturrock et al. 2014). Because an otolith is acellular, metabolically inert, and grows throughout the life of the fish, any elements or compounds accreted onto its surface are permanently retained (Campana and Neilson 1985; Campana 1999). Otoliths therefore contain a complete record of the temperature and chemical composition of the ambient water the fish experienced over its lifespan (Campana 1999). The literature is replete with examples of researchers using otolith chemical composition in ecological investigations, including but not limited to determining migration patterns and habitat usage of anadromous fishes (Secor et al. 1995; Swanson et al. 2010), identifying individual spawning stocks of fishes during mixing events (Campana et al. 1999; Geffen et al. 2011), reconstruction of environmental histories (Matta et al. 2013), and determination of natal origin (Carlson et al. 2016; Stewart et al. 2021).
Otolith microchemical analyses have also been used to validate fish age and include radiochemical dating of calcified structures (see reviews in Campana 1999, 2001) and age estimation based on seasonal patterns in otolith trace element incorporation (Hüssy et al. 2015; Heimbrand et al. 2020). Another validation method relates changes in an otolith’s oxygen isotopic composition (δ18O value) over a fish’s lifespan to oxygen isotopic differences associated with seasonal variation in water temperature. The oxygen isotopic composition of aragonitic fish otoliths exists in equilibrium with ambient seawater (Iacumin et al. 1992) and the fractionation of oxygen isotopes in otoliths is inversely related to temperature (Høie et al. 2004) and is unaffected by either otolith precipitation or fish growth rates (Thorrold et al. 1997). High-resolution sequential measurements of otolith δ18O values over the lifespan of the fish can provide a proxy for annual temperature cycles that the fish experienced. Generally, seasonal temperature cycles generate a pair of maximum (cold water) and minimum (warm water) δ18O values each year, which can be counted to determine the age of the fish. The locations of peak δ18O values can be associated with visible growth marks on the otolith to validate ages made from visible growth zone counts. This validation technique has been performed successfully in studies on Atlantic Cod (Weidman and Millner 2000; Høie and Folkvord 2006) and Pacific Cod (Kastelle et al. 2017; Helser et al. 2018), among others.
Counting growth zones on sagittae has been shown to be the most suitable method to age various rockfish species (Six and Horton 1977); as such, this is the primary method to generate ages for the majority of assessed commercially and recreationally important groundfish in the northeast Pacific (C.A.R.E. 2006). Abundant nearshore groundfish that have commercial and/or recreational importance to Oregon include Black Rockfish (Sebastes melanops Girard, 1856), Copper Rockfish (Sebastes caurinus Richardson, 1844), and Cabezon (Scorpaenichthys marmoratus Ayres, 1854). Adult Black Rockfish occur from Huntington Beach in southern California to Amchitka Island in Western Alaska but are most common from northern California to southeast Alaska (Love 2011). They are a semi-pelagic nearshore and shallow shelf species occurring at depths up to 366 m (Love 2011) but are most common in water shallower than 73 m (Love 2011). Off Oregon, females reach 50% maturity at 7.5 years and 39.5 cm (Bobko and Berkely 2004). Black Rockfish (as with other members of the genus Sebastes) have internal fertilization and populations off Oregon exhibit group synchronous oocyte development and extrude only one batch of larvae per year during winter (Wyllie-Echeverria 1987; Bobko and Berkeley 2004). Larvae and small juveniles are pelagic for approximately 6 months (Boehlert and Yoklavich 1983). Settlement occurs between May and July in estuaries, tide pools, and in nearshore kelp beds and rocky reefs at depths less than 20 m (Stein and Hassler 1989; Ottmann et al. 2018). Black Rockfish tend to inhabit deeper water as they grow and are associated with areas of high relief (Love 2011). In Oregon, they are an important component of the commercial nearshore fishery. They are the primary focus of the recreational fishery, becoming increasingly important in Oregon with the decline of salmon fishing opportunities in the late 1970s and early 1980s (Cope et al. 2016).
Copper Rockfish occur from the western Gulf of Alaska to central Baja California and are most common from southeastern Alaska to northern Baja California. They are a demersal nearshore species; young-of-the-year are found in barely subtidal waters while adults can be found to 183 m (Love 2011). Off Oregon, females reach 50% maturity at 5.6 years and 34.8 cm, parturition peaks in February but may extend through summer (Hannah 2014), and like Black Rockfish, larvae are exuded in a single batch. Copper Rockfish have a two-to-three-month pelagic duration, but Oregon populations exhibit limited dispersal compared to Black Rockfish (Johansson et al. 2008). Juveniles settle in shallow waters along sand/rock interfaces and in areas with benthic macrophytes (Love 2011), but the use of Oregon estuaries as juvenile habitat is sporadic (Dauble et al. 2012; Miller and Shanks 2004b). Adults are typically found hovering above or perched upon boulder fields and other rocky substrata (Love et al. 2002). In Oregon, the commercial nearshore fishery targets them with hook and line and bottom longline gears. Similar to Black Rockfish, recreational removals have increased over time since the first recorded landings occurring in 1979, though the species is not nearly as important to Oregon as Black Rockfish (Wetzel et al. 2021).
Cabezon are a demersal cottid that range from central Baja California to Sitka, Alaska where they occur from the intertidal to a depth of 110 m (Love et al. 2005). Females reach 50% maturity by 3–4 years and 43.8 cm, are batch spawners and exhibit a protracted spawning period that ranges from February through November for Oregon populations, with peak spawning occurring in March–April (Hannah et al. 2009). Spawning occurs in recesses of natural and manmade objects in depths down to at least 20 m (Love 2011). Sticky egg masses are deposited on hard, exposed surfaces and males guard the nests (Lauth 1988). In Puget Sound, Cabezon eggs incubate an average of 34 days before hatch, but little is known regarding latitudinal variation in incubation time (Cope et al. 2019). After hatch, Cabezon remain in the plankton for 3–4 months before settling to tide pools, subtidal areas, rock cobbles, and associated drift algae and eelgrass (Love 2011; Cope et al. 2019); off Oregon, settlement occurs from April through September but peaks between late April and late June (Ottmann et al. 2018). Cabezon use these shallow water areas as nursery grounds until moving offshore to deeper nearshore reefs (Cope et al. 2019). Off Oregon, Cabezon are harvested commercially with hook and line gear, commercial bottom longline and pot gear. They are also a focal target of the recreational fishery. Within the last two decades, however, Cabezon has become a major component of the Oregon commercial nearshore live-fish fishery (Cope et al. 2019). Historically, the majority of recreational Cabezon landings in Oregon have been from the recreational ocean boat fishery with a limited amount from recreational shore fishing. The interest in the fishery has been increasing resulting in protracted fishing seasons.
Although visually derived otolith ages have been used in stock assessment models for decades, accuracy of the ageing method has not been determined for the majority of groundfish species in the northeast Pacific. The main objectives of this study were to develop high resolution δ18O chronologies for these nearshore species of interest and to validate otolith annulus formation by comparing the number and relative location of otolith δ18O maxima to translucent growth bands deposited during water temperature minima. Potentially complicating matters is the oceanography of the northeast Pacific Ocean. The California Current system forms the eastern boundary of the North Pacific Subtropical Gyre and is a monsoonal upwelling system driven by the change in location of the North Pacific High (Huyer 1983). Off Oregon, seasonal wind-driven coastal upwelling begins in late spring and continues through early fall and results in surface water temperature minima (and δ18O maxima) occurring from late spring through mid-summer, while winds during the fall and winter favor downwelling (Huyer 1977). However, the dominant variability in coastal upwelling off Oregon occurs at several-day time scales, and upwelling conditions along the coast often reverse for days at a time (Hickey and Banas 2003). The resulting shifts from colder, more saline upwelling water to warmer, less saline downwelling water would likely affect inter-seasonal variability in seawater δ18O values.
Methods
Initial age estimation via counts of visible growth marks
The Oregon Department of Fish and Wildlife’s Marine Resources Program annually samples commercial and sport landings to obtain fisheries-dependent data for stock assessments. We haphazardly selected 25 pairs of dry-stored sagittae each from Black Rockfish, Copper Rockfish, and Cabezon captured throughout the year in 2019 and representing Oregon’s north, central, and south coasts for ageing and validation (Fig. 1). Initially, the left sagitta from all fish was briefly submerged in tap water in a black-backed petri dish and the surface was examined under reflected light using a binocular dissecting microscope paired with a fiber optic light source (Fig. 2a, c, e). Analyzing the otolith surface under such conditions enhanced the dark translucent zones and allowed for a better understanding of otolith topography and growth mark location prior to subsequent sample analyses. The widely accepted method for production ageing of rockfishes is the break-and-burn method (MacLellan 1997; C.A.R.E. 2006). For Black and Copper rockfishes, the right sagitta was broken through the core in the transverse plane using a scalpel, lightly burned over an alcohol lamp, and coated in mineral oil to accentuate growth marks (Fig. 2b, d). Presumed annual growth rings were counted and age was determined based on methodology developed for rockfishes (C.A.R.E. 2006). Growth marks were initially interpreted along a path from the nucleus to the proximal margin on the dorsal lobe of each otolith, and a second interpretation was performed immediately after along a similar path on the ventral lobe before an age determination was made. For Cabezon, the right sagitta was soaked in ethanol for a week to improve clarity following methods developed by Rasmuson et al. (2019) before being sectioned as described above (Fig. 2f). Initial age readings for Cabezon were made from the nucleus to the proximal edge near the sulcal groove on the ventral lobe due to better clarity of the growth rings, with a second interpretation either on the same lobe but more towards the ventral edge or on the dorsal lobe towards the proximal edge depending on growth mark clarity. Ages were assigned without knowledge of fish length, sex, or port of landing but capture date was known to determine timing of growth mark formation. All otoliths were aged in this fashion twice by the lead author, with several weeks between reads. If the two counts were not equal, a third read was made to resolve the difference between the first two reads before a final age was assigned. Within-reader precision was estimated in terms of absolute percent error (APE) as outlined in Beamish and Fournier (1981) (Table 1). The purpose of these age estimates was threefold; they allowed us to express the relative difficulty in ageing otoliths of each species, they informed which samples would be most appropriate for validation, and because these ages will be used in future stock assessments, obtaining multiple estimates follows current C.A.R.E. protocols (C.A.R.E. 2006).

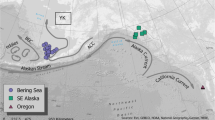
Map showing Oregon ports utilized by commercial and recreational groundfish fishery fleets

Whole otoliths and their break-and-burn preparations for Black Rockfish (a,b), Copper Rockfish (c,d) and Cabezon (e,f). These otoliths were chosen from five-year-old fish for comparative purposes and to illustrate growth marks. Red dots indicate annuli. “p” = proximal, “d” = distal, “vl” = ventral lobe, “dl” = dorsal lobe
Secondary ion mass spectrometry (SIMS)
The left sagitta of each otolith pair was prepared for SIMS oxygen isotope analysis at the Canadian Centre for Isotopic Microanalysis (CCIM), University of Alberta. Individual otoliths were placed on acrylic plates and embedded in water-soluble thermosetting adhesive. One-mm thick transverse otolith sections were made adjacent to the nucleus using a slow speed saw, and the halves were removed from the adhesive using warm water. One of the halves of each otolith were arrayed on adhesive tape and cast together in epoxy along with a calcite reference material into 25-mm diameter mounts. The mounts were ground and polished with diamond compounds to achieve a uniformly flat surface, and polarized incident light microscope images were acquired for each otolith. Prior to SIMS analysis, mounts were cleaned with soap and de-ionized water, and coated with gold. We used the polarized incident light microscope images to determine which otoliths had the clearest growth marks and to determine the area for sampling spot transects.
Oxygen isotope ratios (18O/16O) were determined using the IMS-1280 multi-collector ion microprobe at CCIM. In order to determine the best methodology for SIMS analysis, we performed experimental transects on Black Rockfish and Cabezon sagittae using 15-µm diameter spots with varying distances between centers (~ 20–30 µm). These experimental transects ran from the core to the ventral edge of the ventral lobe, which provided the greatest spacing between growth marks; however, growth marks along this transect were often faded and could be difficult to see after SIMS preparation, and also tended to bunch up and become indistinguishable at the edge of the otolith in older fish, making interpretation difficult. Final transects were then chosen from an area near the core of each otolith in a straight line to the proximal edge near the sulcal groove on the otolith’s ventral lobe, an area that consistently exhibited the clearest growth marks that were more easily identified. We selected eleven sagittae from Black Rockfish, twelve from Copper Rockfish, and eight from Cabezon that exhibited the clearest growth marks from core to edge and were relatively free of major cracks or surface imperfections in the transect area for SIMS analysis (Table 2, Fig. 3). It should be noted that while these final transects began near the core area for Copper Rockfish and Cabezon, this was not the case for all Black Rockfish; in several instances, the final transects began just prior to the first visual growth mark and were informed by experimental transect data. Polarized incident light microscope images of all sagittae showing SIMS transect paths can be found in Online Resource 1.

Images of thin-sectioned Black Rockfish, Copper Rockfish, and Cabezon sagittae. The black line on each otolith indicates the probe path for SIMS analysis, and white numbers along the SIMS path are reference markers required to determine spot locations along the transect. Black spots on the Black Rockfish image illustrates the experimental transect from core to ventral edge
Final primary beam conditions included the use of 20 keV 133Cs+ ions focused to form a probe of 10-µm diameter spots with approximately 12 µm between centers and beam current ~ 2.0 nA. The primary beam was rastered for 30 s prior to analysis to clean the surface of Au and contaminants and implant Cs. The normal incidence electron gun was utilized for charge compensation. Negative secondary ions were extracted through 10 kV potential into the secondary column (Transfer section). Conditions for the Transfer section included an entrance slit width of 122 µm, field aperture of 5 × 5 mm, and a field aperture-to-sample magnification of 100 × . Automated tuning of the secondary ions in the Transfer section preceded each analysis. The energy slit was fully open. Both 16O− and 18O− were analyzed simultaneously in Faraday cups (L’2 using 1010 Ω amplifier, and H’2 with 1011 Ω) at mass resolutions of 2000 and 2275, respectively. Mean count rates for 16O− and 18O− were typically 2.0 × 109 and 4 × 106 counts/s, respectively, determined over a 60 s counting interval. Total spot-to-spot time was 200 s. Typical spot depths after completion were < 1 µm. The analytical sequence interspersed analyses of unknowns with in-house end-member calcite reference material (CCIM sample S0161 with δ18OVSMOW = + 25.33 ‰, δ18OVPDB = − 5.42 ‰; R. Stern, unpubl. data) in a 4:1 ratio. Instrumental mass fractionation (IMF) for 18O−/16O− was determined for each analytical session from utilizing all the replicate analyses of S0161 calcite. The standard deviation of multiple measurements of 18O−/16O− ratios in S0161 was ± 0.10 – 0.13 ‰, after small corrections for systematic within-session IMF drift. The calcite-calibrated data for unknowns were corrected by − 0.95 ‰ to account for the difference in IMF between calcite and the aragonite matrix of the otoliths, determined by separate experiments within CCIM (R. Stern, unpubl. data). Individual spot uncertainty in δ18O for unknowns was ± 0.25 ‰ (2σ). Individual analyses were reported in standard notation (δ18O‰) relative to Vienna Standard Mean Ocean Water (VSMOW) and Vienna Pee Dee Belemnite (VPDB). Conversion between these two standards is expressed by the formula: \(VSMOW=\frac{VPDB+29.99}{0.97001}\).
Data analysis
In his review of age validation methods, Campana (2001) recommended that two procedures be followed when it was not possible to determine a fish’s absolute age: determination of age at first increment formation and verification of increment periodicity across the entire age range of interest. To validate growth mark periodicity, post-probe polarized incident light microscope images of each otolith taken by CCIM showed the SIMS beam path and individual spot identifiers. These photographs were used to measure the distance from the beginning of each transect to the leading edge of each translucent growth mark, from the area near the otolith core to its proximal edge using AmScope (2021) camera software. SIMS output provided the planar X–Y coordinates for each spot that were used to calculate the cumulative distance of each spot from the start of the transect. δ18O values were plotted by distance from the transect start to illustrate peaks and troughs in the data, then overlaid the translucent growth mark distances to determine if observable growth marks lined up with peaks in the δ18O signal. To determine the extent of δ18O variation with fish age, we calculated the standard deviation of δ18O between growth marks for each fish and averaged those values to determine mean variation by age for each species. To test data reproducibility, δ18O values were compared between the experimental and the final transects made on the Black Rockfish and Cabezon sagittae. Because beam conditions differed between transects, we averaged the δ18O values between data peaks on both transects and evaluated data pairs using linear regression (see Helser et al. 2018). Due to growth mark crowding on the edge of the experimental transects, we restricted the analysis to the first seven peaks for Black Rockfish and the first five peaks for Cabezon.
To evaluate the use of maxima in the δ18O chronology as a proxy for true age, peaks were identified and counted in each chronology by three trained readers without knowledge of age estimates obtained by the break-and-burn method; confidence intervals (2σ) of each data point were used to determine if the values were sufficiently analytically distinct to warrant classification as maxima and the relative spacing of each maxima along the transect was used to aid in determinations. Previous studies have shown an approximate 1‰ change in the otolith δ18O composition over a 5° C change in water temperature (Thorrold et al. 1997; Weidman and Millner 2000; Høie et al. 2004; Kastelle et al. 2017), and we assumed this relationship held when evaluating seasonal cycles vs. intra-seasonal variability. We used the Coastal Upwelling Transport Index (CUTI) (mjacox.com/upwelling-indices/), a measure of upwelling strength and a proxy for water temperatures experienced by the fish over their lifespans because no synoptic measurements of water temperature exist off Oregon. The index more accurately measures upwelling strength compared to previous indices and has been validated extensively (Jacox et al. 2018).
The age at first increment formation was difficult to determine for fishes in this study. Production agers at state and federal facilities rely on fisheries-dependent catches of the species to be assessed, and due to existing size and/or seasonal take limits, ageing structures are typically unavailable from small, young fish so other sources of young fish must be used to obtain otolith samples. To validate age of first increment formation in Black Rockfish and Cabezon, we relied on data collected through a collaborative project run by Oregon State University and the Oregon Department of Fish and Wildlife’s Marine Reserves program. For Black Rockfish, we obtained radial otolith measurements from the core to the post-rostral edge from 24 young-of-the-year captured in late October 2016 (H.W. Fennie, K. Grorud-Colvert, S. Sponaugle, Oregon State University, unpubl. data) and assumed that otolith radii in these young-of-the-year fishes approximated that of 1-year-old fish. We compared these radii to similar measurements made on the distal surface of 24 age-5 Black Rockfish captured by the commercial fleet in 2020. Otoliths from these older fish were submerged in a clear petri dish of tap water and examined under transmitted light, which accentuated the otolith core and subsequent growth marks. Amscope software (2021) was used to produce digital images of the samples, and the distance from the core to the inner edge of the first visible growth mark was measured. Unfortunately, we were unable to obtain otoliths from young-of-the-year Copper Rockfish but assumed that the first growth mark identified by surface examination was the first annulus. For Cabezon, newly settled juveniles were collected from April to September over the course of several years (2013, 2015–2016, and 2018–2019), aged, and otolith radii were measured from core to dorsal edge. The average age and radius of these fish was 99 days and 350.7 µm, respectively (M. Wilson, K. Grorud-Colvert, S. Sponaugle, Oregon State University, unpubl. data). We used this mean radial measurement as a benchmark to determine location of the first annulus.
Results
Break-and-burn ages and identification of the first annulus
The 25 Black Rockfish aged by counting growth zones on break-and-burned otoliths ranged from 5 to 25 years old. When viewed under reflected light, burned sections exhibited relatively wide opaque zones separated by narrow dark brown or black translucent zones (hereafter referred to as annuli) that were easily counted. True annuli were of high contrast, were continuous, and could be followed from the dorsal lobe to the ventral lobe by examining the distal surface of the otolith (Fig. 2b). Abrupt optical discontinuities (checks) were noted within the first five opaque zones, but they were irregularly spaced, relatively thin compared to annuli, and merged into true annuli in the distal area of the section which made them relatively easy to distinguish. Subsequent growth zones were relatively narrow and regularly spaced, with no checks observed. The greatest difficulty in ageing this species was counting annuli near the proximal edge in older fish due to very narrow spacing between growth marks. We identified the first continuous bold mark surrounding the core as the first annulus, which was relatively wide and distinct (mean maximum diameter of 2560 µm, range 2200–3100 µm), and annuli appeared on the proximal edge beginning in early March. Double blind reads on these samples resulted in an APE of 1.08%, indicating high within-reader precision for these samples (see Campana 2001); twenty-one of the reads matched, 3 differed by 1 year, and one difficult sample differed by 5 years (Table 1). When we compared otolith radii between newly settled young-of-the-year and older Black Rockfish, we found that the mean radial measurement from the core to the post rostral edge in the young-of-the-year fish (2326.3 µm) was similar to the mean measurement made to the inner edge of the first growth mark in older Black Rockfish (2352.2 µm). Differences were not significant (t test; df = 48, t = − 0.72, p = 0.48), corroborating our determination of the first annulus.
The 25 Copper Rockfish aged in this study ranged from 4 to 11 years old and were more difficult to age than Black Rockfish. It was often difficult to determine the leading edge of the first annulus due to multiple checks that occurred in the core area, especially on the dorsal lobe, and we had to follow the zonation from the burnt face over the distal surface and determine the boldest mark before delimiting the extent of the first annulus. Using this method, we determined a mean maximum diameter of the first annulus of 2100 µm (range 1700–2500 µm). The following 4 annuli were bold, widely spaced, and better defined, but subsequent annuli were relatively faint and often difficult to see on the edge (Fig. 2d). Double blind reads on these samples resulted in an APE of 1.65%, only slightly less precise than for Black Rockfish. Despite considerable fading between first and second reads, we matched on 20 reads, with the remaining 5 differing by a single year (Table 1). Annuli were noticed on the otolith margin by early May. Although no age-0 Copper Rockfish were available to validate the age of first increment formation, we followed C.A.R.E. (2006) guidelines and assumed that the first continuous dark zone formed outside the nucleus that could be followed along the distal surface of the otolith was the first annulus. In samples with multiple checks in this area, we looked for where these discontinuities merged near or at the edge of the sulcal groove.
The 25 Cabezon aged in this study ranged from 2 to 12 years and were relatively difficult to handle and age based on their small otolith size (only approximately 5000 µm in length but thickening with age). The first two annuli were relatively prominent but contained numerous checks and split rings that added to ageing difficulty. Rasmuson et al. (2019) examined both break-and-burn and thin sectioned otoliths and reported mean annulus maximum diameters of between 1200 and 1500 µm for the first and 1900 µm for the second annulus in this species. The mean otolith radius of newly settled Cabezon was used as a benchmark and aided in the identification of the first annulus, which was the first bold, thick translucent growth zone outside of this mean radial measurement. These measurements were used to aid interpretation of growth marks in this study, and mean diameters of the first and second annulus were measured at 1410 µm (range 1240–1600 µm) and 1830 µm (range 1700–1980 µm), respectively. Annuli were distinct and easy to identify along the ventral lobe towards the proximal surface (Fig. 2f). Double blind reads on these samples resulted in an APE of 2.23%, with 18 age matches and 7 off by 1 year (Table 1). Annuli were noted on the otolith edge from mid-May through mid-July.
Secondary ion mass spectrometry
Preparation of otolith samples for SIMS analysis was a complex, time-consuming endeavor that included embedding, sectioning and polishing sections to a flat plane free of cracks and surface imperfections for isotopic ratio quantification. In many cases, especially for Black Rockfish and Copper Rockfish (species with relatively large otoliths) (Fig. 3) samples needed to be slightly over-polished to obtain a clear probe path from core to edge. This over-polishing resulted in several otolith samples exhibiting edge erosion or that had annuli that were difficult to see in polarized incident light microscope images. Over-polishing was not necessary for the Cabezon otoliths, but cracks in the sample were more prevalent in these smaller otoliths. Although these sample preparation artifacts reduced the number of samples analyzed by SIMS, we were still able to analyze multiple samples from each species caught off multiple Oregon ports.
SIMS proved to be effective for producing detailed otolith δ18O chronologies with adequate temporal resolution for identification of seasonal trends in water temperatures. In situ analysis of oxygen isotope ratios was possible using small diameter (10 µm) probe spots with minimal spacing and allowed for multiple spot analyses per visible growth zone on the otoliths. We averaged 90, 99, and 67 probe spots per sample for Black Rockfish, Copper Rockfish, and Cabezon, respectively (Table 2). Locations of annuli counted during break-and-burn ageing generally corresponded to maxima in the δ18O chronologies, inferring formation during periods when water temperatures were relatively cold. Generally, chronologies were characterized as having one strong signal peak and trough between annuli; however, the δ18O chronologies could be highly irregular and chaotic between maxima. Chronologies were especially irregular between the otolith core and the first annulus. For brevity, we show three δ18O chronologies, one per species, in Fig. 4; chronologies of the remaining otoliths can be viewed in Online Resource 2. The variation in δ18O values between the otolith core and the first annulus meant that the use of maxima counts as a proxy for age could not occur until the location of the first annulus along each chronology was established; further, interpretation of the δ18O signal from first annulus to the otolith margin required an understanding of the general trend of decreasing otolith increment width with age. This made the counting of maxima difficult and rather subjective; however, blind peak counts made by three independent analysts showed general agreement between break-and-burn ages and δ18O maxima (R = + 0.79 for Black Rockfish and + 0.92 for both Copper Rockfish and Cabezon) (Fig. 5). Of 33 comparisons, Black Rockfish break-and-burn ages agreed with peak counts 13 times and underaged compared to peak counts an additional 13 times, with greater deviation seen in older (> 8 years) specimens. Ages between methods matched on 24 of 36 Copper Rockfish, with all other comparisons off by one year. Cabezon ages matched on 9 of 24 attempts, in 11 instances the maxima count exceeded the break-and-burn age, and deviation tended to be larger for older fish (> 7 years). The annual variation in otolith δ18O was highest during the first year of life and decreased with age. This downward trend was slight for Black Rockfish, intermediate for Copper Rockfish, and relatively pronounced for Cabezon (Fig. 6).

Measurements of δ18O values (‰ VSMOW) from a representative Black Rockfish, Copper Rockfish, and Cabezon. Error bars represent ± 2σ. Vertical lines represent location of growth marks along the SIMS transect, with corresponding calendar year of formation above each line. The probe transect stopped prior to the edge for this Cabezon sample, where a growth mark was present. Growth marks are found on the otolith edge in late spring of each year

Age bias plots showing numbers of individuals by each break-and-burn age and δ18O maxima count combination for (a) Black Rockfish, (b) Copper Rockfish, and (c) Cabezon. Numbers along the dashed line indicate numbers of samples where the two ageing methods agree

Standard deviation of δ18O values (‰ VSMOW) between annuli on otoliths for (a) Black Rockfish, (b) Copper Rockfish, and (c) Cabezon. Trendline is a loess smoother fit to the data; shaded area represents the 95% confidence interval band
Comparison of average annual δ18O values between the experimental and final transects showed a significant relationship for both Black Rockfish and Cabezon (Fig. 7). For Black Rockfish, the regression of final on experimental transect δ18O values was statistically significant (r2 = 0.70, p < 0.001) with an average offset of 0.54‰ between transects. The regression for Cabezon was also statistically significant (r2 = 0.91, p < 0.001), with an average offset of 0.33‰ between transects. In both cases, average δ18O values were higher in the final transect compared to the experimental transect. We also noted a general trend of increasing average δ18O values with distance from the otolith core in sagittae from both species.

Average δ18O values (‰ VSMOW) calculated between each SIMS peak along experimental transects (X1) and final transects (XN) for (a) Black Rockfish and (b) Cabezon. The solid line represents a linear fit to all points in the regression, and the dashed line represents the 1:1 relationship. The maximum number of peaks analyzed for the analysis was seven for Black Rockfish and five for Cabezon and reflects the age at which annuli crowding on the edge prevented accurate δ18O measurements. All peak-to-peak comparisons were made for Cabezon; for Black Rockfish, there were eight instances where the final transect was truncated in the area prior to peak 1 compared to the experimental transect, so those comparisons were removed from the analysis. Black Rockfish #OR1974564-22 was also not analyzed due to difficulties in identifying growth marks along the experimental transect
Discussion
This study represents the first attempt at using SIMS to validate otolith increment counts in commercially and recreationally important northeast Pacific nearshore groundfish species off Oregon, USA. Since acknowledgement of the dearth of validation studies for most northeast Pacific groundfish made over 20 years ago (Munk 2001) and repeated calls for such studies by stock assessment authors (Cope et al. 2016), we found few validation studies conducted over the past two decades on Oregon groundfish in the published literature. Such studies have included age validation of demersal slope Aurora Rockfish (Sebastes aurora Gilbert, 1890) using cross-dating techniques (Thompson and Hannah 2010) and validation of shelf Canary Rockfish (Sebastes pinniger Gill, 1864) using bomb radiocarbon validation (Piner et al. 2005a). Piner et al. (2005b) also attempted to use oxygen stable isotopes to validate ages in Black Rockfish, but their study used micromilling to obtain an average age-specific δ18O value per year to relate to warm water produced by the 1983 El Niño. At the time of their study, the use of ion microprobe for otolith δ18O analysis was limited due to relatively large spot diameters and lower analytical precision (Weidel et al. 2007); therefore, their study lacked the temporal resolution necessary to delineate intra-seasonal variations in water temperature. This current study serves as an important contribution to the list of northeast Pacific species with validated ages, including Black Rockfish, the most recreationally caught and arguably the most important nearshore rockfish species to Oregon.
The seasonal (and intra-seasonal) variation in otolith δ18O was influenced by coastal circulation dynamics. Currents off Oregon are dominated by the California Current, an eastern boundary current that flows north to south along the west coast of North America and Baja California. During spring and summer, predominately northerly winds drive surface waters offshore via Ekman transport resulting in seasonal upwelling of cold, salty nutrient-rich water from depth up on the continental shelf. The seasonal variation in the physical oceanography of the continental shelf off Oregon can be large, with surface and bottom water temperature minima and salinity maxima typically occurring in late spring through summer (Huyer 1977). Upwelling events during the summer are typified by periods of upwelling followed by periods of relaxation, resulting in changes in temperature and salinity at the scale of days to weeks (Huyer 1983). Locally, upwelling circulation is modified by interactions with tidal and wind-driven currents as well as bathymetry (Gan et al. 2005). In the fall, currents off Oregon become characteristically downwelling favorable and are dominated by large winter storms. Alongshore flow on the shelf is northwards advecting warm waters into nearshore habitats. These circulation dynamics are obvious in the CUTI, an estimate of vertical transport near the west coast of North America (Fig. 8). A negative relationship exists between CUTI and water temperature (Fig. 9); off central Oregon, a decrease in water temperature of approximately 2.75° C corresponds to an increase in the CUTI by 1.0. Salinity of the ambient water has been shown to interact positively with water temperature to significantly influence otolith δ18O (Campana 1999; Elsdon and Gillanders 2002) and varies seasonally due to local runoff, riverine inputs, and coastal upwelling and alongshore advection (Huyer 1977). Production ageing of otoliths from these species has shown that annulus formation typically occurs in mid- to late spring each year (Mark Terwilliger, ODFW, unpubl. data) and the fish examined in this study followed that trend: timing of annulus formation corresponded to an increase in the seasonal upwelling index off Oregon and resulting colder water temperatures. This correlation between the CUTI upwelling index and temperature observations suggests there is a linkage between seasonal circulation dynamics and δ18O.

Mean monthly Coastal Upwelling Transport Index (mjacox.com/upwelling-indices/) at 44°N latitude, 1988–2021. Positive values represent timing and magnitude of upwelling events, while negative values represent the same for downwelling periods

Relationship between the Coastal Upwelling Transport Index at 44° N latitude and monthly mean water temperatures obtained along the Oregon Line of the Ocean Observatories Initiative’s (OOI) Coastal Endurance Array, located off Newport, OR
The impetus behind age validation studies is the subjectivity and uncertainty of visual interpretations of growth marks used to estimate age (Campana 2001; Katayama 2018). Visible growth marks on the otoliths of these species were identifiable and easily enumerated. For these species, annuli appeared relatively dark and wide under reflected light until fish reached maturity, after which annuli appeared as thin, dark, regularly spaced narrow bands (Types A and B zones, respectively; see Katayama 2018). The primary difficulties were identifying annuli occurring on the otolith margin and differentiating checks from annuli. The relative ease of ageing these samples was reflected in APE values far lower than Campana’s (2001) reference point value of 3.65% for species with moderate longevity and reading complexity, inferring high single-ager precision, and ages estimated in this study fall within expected length-at-age ranges published in stock assessments (Black Rockfish: Cope et al. 2016; Copper Rockfish: Wetzel et al. 2021; Cabezon: Cope et al. 2019). Despite the variability in the δ18O chronologies, we demonstrated that ages estimated by standard break-and-burn methods were highly correlated with δ18O maxima counts, with general agreement between the two methods. These maxima coincided with the seasonal increase in the CUTI and coincident decrease in water temperatures off Oregon. Despite the general agreement, the analysis required knowledge of first annulus location, general otolith increment width spacing and potential seasonal amplitude of the δ18O values. With this information in hand, counting δ18O peaks as a proxy for age was relatively straightforward for several of the samples. However, there were instances, especially for Black Rockfish, when the seasonal δ18O signal was overwhelmed by intra-seasonal variation in δ18O that made it difficult to distinguish peaks without reference to the visible growth marks (Online Resource 2). Decreased variability in the δ18O values with age typically had a dampening effect which made peaks at older ages more pronounced and easier to identify. While the variability in δ18O did decrease slightly with age for Black Rockfish (Fig. 6), intra-seasonal peaks occurred more frequently than in Copper Rockfish and Cabezon, leading to a greater percentage of overestimated ages using peak counts.
Initial experimentation with transect location allowed us to make inferences regarding the reproducibility of δ18O data for Black Rockfish and Cabezon. We saw a change in the otolith growth plane at approximately the age at maturity in these species that caused a compression of otolith growth marks on the leading edge of the experimental transect (Online Resource 1); however, we were able to compare average δ18O up until that point (age seven for Black Rockfish and age five for Cabezon). Up to those ages, we were able to identify similar numbers of seasonal cycles along both transects, δ18O values were generally lower along the experimental transect compared to the final transect, and the relationship between average δ18O values along the two transects was linear and positive (Fig. 7). The offset seen for both species may be related to a matrix effect due to the change in beam size and sample density between transects or perhaps is due to a difference in organic content between transects as proteins are known to contain lower δ18O values (Shiao et al. 2014; Helser et al. 2018). The general increase in average otolith δ18O and general decrease in δ18O variability with age infers possible ontogenetic movements from shallower to colder, deeper water with age for these species, or perhaps may be due to reduced organics incorporated into the otoliths with age. Shiao et al. (2014) noted reduced δ18O values in the core and larval growth areas of deep-sea fishes associated with protein-induced fractionation effects. Regardless, our goal in this study was to use seasonal variation in otolith δ18O values to validate annual marks in otoliths, so only relative changes in the δ18O signal through time were of interest.
In his study comparing analytical techniques to measure oxygen isotopes in fish otoliths, Helser et al. (2018) noted that the high-resolution in situ sampling of SIMS provided much more detail in describing variability in δ18O values than did conventional micromilling and isotope ratio mass spectrometry, which rely on the conglomeration of bulk powder samples that often represent timescales that differ from the ones in question (Valley and Kita 2009). This increased detail, coupled with seasonal upwelling that varies regionally and the life history strategies of these species, complicated interpretation of the δ18O chronologies. For example, variability in δ18O values prior to age-1 that prevented us from identifying an initial maximum may be due to the extended pelagic larval and juvenile duration exhibited by these species as well as their recruitment into shallow habitats with dynamic temperature and salinity regimes. All rockfish are live-bearers and have a pelagic larval and early juvenile stage that lasts from 2 to 6 months and returns to nearshore habitats to recruit (Moser & Boehlert 1991). After parturition, Black Rockfish larvae and juveniles remain offshore for approximately three months, and although previous studies have shown a dominant alongshore dispersal distance of < 120 km, the distance can be greater for a large percentage of individuals (Miller and Shanks 2004a). They then recruit to shallow water and near-coastal habitats, where they remain for several months before migrating to slightly deeper coastal waters (S. Bobko, Oregon State University, unpubl. data). Copper Rockfish have a two-to-three-month pelagic duration, and compared to Black Rockfish, Oregon populations exhibit restricted dispersal due to habitat barriers (Johansson et al. 2008). Cabezon are not live-bearers but the young of the year spend 3–4 months as pelagic larvae and juveniles with an extensive dispersal (O’Connell 1953). Inter-seasonal short-term variation in the CUTI and the ecology of these species also influenced interpretation of δ18O between the first annulus and the otolith margin. Unlike Copper Rockfish and Cabezon, both which are demersal with relatively small home ranges, Black Rockfish are semi-pelagic, are known to move more than other demersal rockfishes, and have a better chance of encountering source waters with differing δ18O values. Black Rockfish are known to make vertical movements from the bottom to the midwaters, and large schools have been reported at the surface when chasing prey (Love 2011). As discussed earlier, although the circulation off Oregon changes seasonally, strong local variability results in alongshore changes in oxygen isotopic ratios. As such, due to their location in the water column and their propensity for alongshore movement and vertical migrations they likely experience water masses with greater variability in water temperature, salinity and δ18O at the scales of days, weeks or months than their demersal counterparts. We hypothesize this is why there were a greater number of intra-annual peaks in δ18O and a relatively slight decrease in otolith δ18O seasonal variability for Black Rockfish.
Our study found that otolith δ18O was highly correlated with break-and-burn ages in these three species of nearshore groundfish off Oregon. The utility of SIMS to validate annuli in these species was complicated by intra-seasonal variation in the upwelling strength off the coast of Oregon. We found that intra-reader precision was high when ageing these species by traditional break-and-burn methods, and that δ18O maxima counts can be used to validate ages up to 12 years for Black Rockfish, 9 years for Copper Rockfish, and 11 years for Cabezon. For this study, a continuous δ18O signal was sampled in a transect across the otolith cross section in evenly spaced spots from the core area to the proximal edge using a finite-width probe. Because growth zones become depressed with age, especially in long-lived rockfish species, it may be necessary to selectively choose areas on the otolith to probe to ensure visible growth zones are cleanly sampled in order to validate older ages in these species.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
AmScope (2021) The microscope camera software help manual, p 136
Beamish RJ, Fournier DA (1981) A method for comparing the precision of a set of age determinations. Can J Fish Aquat Sci 38:982–983. https://doi.org/10.1139/f81-132
Bobko SJ, Berkely SA (2004) Maturity, ovarian cycle, fecundity, and age-specific parturition of Black Rockfish (Sebastes melanops). Fish Bull 102:418–429
Boehlert GW, Yoklavich MM (1983) Effects of temperature, ration, and fish size on growth of juvenile Black Rockfish, Sebastes Melanops. Environ Biol Fish 8(1):17–28. https://doi.org/10.1007/BF00004942
C.A.R.E. (Committee of Age Reading Experts) (2006) Manual on generalized age determination procedures for groundfish. Prepared for the Technical Subcommittee of The Canada/U.S. Groundfish Committee, p 52
Campana SE (1999) Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Mar Ecol Prog Ser 188:263–297. https://doi.org/10.3354/meps188263
Campana SE (2001) Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol 59:197–242. https://doi.org/10.1111/j.1095-8649.2001.tb00127.x
Campana SE, Neilson JD (1985) Microstructure of fish otoliths. Can J Fish Aquat Sci 42:1014–1032. https://doi.org/10.1139/f85-127
Campana SE, Chouinard GA, Hanson JM, Fréchet A (1999) Mixing and migration of overwintering cod stocks near the mouth of the Gulf of St. Lawrence. Can J Fish Aquat Sci 56:1873–1881. https://doi.org/10.1139/f99-083
Carlson AK, Finchel MJ, Graeb BDS (2016) Otolith microchemistry reveals natal origins of Walleyes in Missouri River reservoirs. N Am J Fish Manage 36(2):341–350. https://doi.org/10.1080/02755947.2015.1135214
Cope JM, Sampson D, Stephens A, Key M, Mirick PP, Stachura M, Tsou T-S, Weyland P, Berger A, Buell T, Councill E, Dick EJ, Fenske KH, Monk M, Rodomsky BT (2016) Assessments of California, Oregon and Washington Stocks of Black Rockfish (Sebastes melanops) in 2015. Pacific Fishery Management Council, Portland, OR, p 402
Cope JM, Berger AM, Whitman AD, Budrick JE, Bosley KM, Tsou T, Niles CB, Privitera-Johnson K, Hillier LK, Hinton KE, Wilson MN (2019) Assessing Cabezon (Scorpaenichthys marmoratus) stocks in waters off of California and Oregon, with catch limit estimation for Washington State. Pacific Fishery Management Council, Portland, OR. p 439
Dauble AJ, Heppell SA, Johansson ML (2012) Settlement patterns of young-of-the-year rockfish among six Oregon estuaries experiencing different levels of human development. Mar Ecol Prog Ser 448:143–154. https://doi.org/10.3354/meps09504
Elsdon TS, Gillanders BM (2002) Interactive effects of temperature and salinity on otolith chemistry: challenges for determining environmental histories of fish. Can J Fish Aquat Sci 59:1796–1808. https://doi.org/10.1139/f02-154
Gan JP, Allen JS, Samelson R (2005) On open boundary conditions for a limited-area coastal model off Oregon. Part 2: response to wind forcing from regional mesoscale atmospheric model. Ocean Model 8:155–173. https://doi.org/10.1016/j.ocemod.2003.12.007
Geffen AJ, Nash RDM, Dickey-Collas M (2011) Characterization of herring populations west of the British Isles: an investigation of mixing based on otolith microchemistry. ICES J Mar Sci 68(7):1447–1458. https://doi.org/10.1093/icesjms/fsr051
Hannah RW (2014) Length and age at maturity of female Copper Rockfish (Sebastes caurinus) from Oregon waters based on histological evaluation of ovaries. ODFW Informational Report #2014–04, Oregon Department of Fish and Wildlife, p 18
Hannah RW, Blume MTO, Thompson JE (2009) Length and age at maturity of female Yelloweye Rockfish (Sebastes rubberimus) and Cabezon (Scorpaenichthys marmoratus) from Oregon waters based on histological evaluation of maturity. ODFW Informational Report #2009–04, Oregon Department of Fish and Wildlife, p 32
Heimbrand Y, Limburg KE, Hüssy K, Casini M, Sjöberg R, Palmén Bratt A-M, Levinsky S-E, Karpushevskaia A, Radke K, Öhlund J (2020) Seeking the true time: exploring otolith chemistry as an age-determination tool. J Fish Biol 97:552–565. https://doi.org/10.1111/jfb.14422
Helser TE, Kastelle CR, McKay JL, Orland IJ, Kozdon R, Valley JW (2018) Evaluation of micromilling/conventional isotope ratio mass spectrometry and secondary ion mass spectrometry of δ18O values in fish otoliths for schlerochronology. Rapid Commun Mass Spectrom 32:1781–1790. https://doi.org/10.1002/rcm.8231
Hickey BM, Banas NS (2003) Oceanography of the U.S. Pacific Northwest coastal ocean and estuaries with application to coastal ecology. Estuaries 26(4B):1010–1031. https://doi.org/10.1007/BF02803360
Hilborn R, Walters CJ (1992) Qualitative fisheries stock assessment: choice, dynamics, and uncertainty. Routledge, Chapman & Hall Inc, p 575
Høie H, Folkvord A (2006) Estimating the timing of growth rings in Atlantic Cod otoliths using stable oxygen isotopes. J Fish Biol 68(3):826–837. https://doi.org/10.1111/j.0022-1112.2006.00957.x
Høie H, Otterlei E, Folkvord A (2004) Temperature-dependent fractionation of stable oxygen isotopes in otoliths of juvenile cod (Gadus morhua L.). ICES J Mar Sci 61:243–251. https://doi.org/10.1016/j.icesjms.2003.11.006
Hüssy K, Gröger J, Heidemann F, Hinrichsen H-H, Marohn L (2015) Slave to the rhythm: seasonal signals in otolith microchemistry reveal age of eastern Baltic Cod (Gadus morhua). ICES J Mar Sci 73:1019–1032. https://doi.org/10.1093/icesjms/fsv247
Huyer A (1977) Seasonal variation in temperature, salinity, and density over the continental shelf off Oregon. Limnol Oceanogr 22(3):442–453. https://doi.org/10.4319/lo.1977.22.3.0442
Huyer A (1983) Coastal upwelling in the California current system. Prog Oceanogr 12:259–284. https://doi.org/10.1016/0079-6611(83)90010-1
Iacumin P, Bianucci G, Longinelli A (1992) Oxygen and carbon isotopic composition of fish otoliths. Mar Biol 113:537–542. https://doi.org/10.1007/BF00349696
Jacox MG, Edwards CA, Hazen EL, Bograd SJ (2018) Coastal upwelling revisited: Ekman, Bakun, and improved upwelling indices for the U.S. West Coast. J Geophys Res Oceans 123:1–19. https://doi.org/10.1029/2018JC014187
Johansson ML, Banks MA, Glunt KD, Hassel-Finnegan HM, Buonaccorsi VP (2008) Influence of habitat discontinuity, geographical distance, and oceanography on fine-scale population genetic structure of Copper Rockfish (Sebastes caurinus). Mol Ecol 17(13):3051–3061. https://doi.org/10.1111/j.1365-294x.2008.03814.x
Kastelle CR, Helser TE, McKay JL, Johnston CG, Anderl DM, Matta ME, Nichol DG (2017) Age validation of Pacific Cod (Gadus macrocephalus) using high-resolution stable oxygen isotope (δ18O) chronologies in otoliths. Fish Res 185:43–53. https://doi.org/10.1016/j.fishres.2016.09.024
Katayama S (2018) A description of four types of otolith opaque zone. Fish Sci 84:735–745. https://doi.org/10.1007/s12562-018-1228-z
Lauth RR (1988) Seasonal spawning cycle, spawning frequency, and batch fecundity of the Cabezon, Scorpaenichthys marmoratus, in Puget Sound, Washington. Fish Bull 87:145–154
Love M (2011) Certainly more than you want to know about the fishes of the Pacific Coast. Really Big Press, Santa Barbara, California, p 672
Love MS, Yoklavich M, Thorsteinson L (2002) The rockfishes of the northeast Pacific. University of California Press, Berkeley, p 404
Love MS, Mecklenburg CW, Mecklenburg TA, Thorsteinson LK (2005) Resource inventory of marine and estuarine fishes of the West Coast and Alaska: a checklist of North Pacific and Arctic Ocean species from Baja California to the Alaska - Yukon border (USGS Unnumbered Series). U.S Geological Survey, Seattle, p 276
MacLellan SE (1997) How to age rockfish (Sebastes) using S. alutus as an example-the otolith burnt section technique. Can Tech Rep Fish Aquat Sci 2146:39
Matta MA, Orland IJ, Ushikubo T, Helser TE, Black BA, Valley JW (2013) Otolith oxygen isotopes measured by high-precision secondary ion mass spectrometry reflect life history of a Yellowfin Sole (Limanda aspera). Rapid Commun Mass Spectrom 27:691–699. https://doi.org/10.1002/rcm.6502
Miller JA, Shanks AL (2004a) Evidence for limited larval dispersal in Black Rockfish (Sebastes melanops): implications for population structure and marine-reserve design. Can J Fish Aquat Sci 61:1723–1735. https://doi.org/10.1139/F04-111
Miller JA, Shanks AL (2004b) Ocean-estuary coupling in the Oregon upwelling region: abundance and transport of juvenile fish and of crab megalopae. Mar Ecol Prog Ser 271:267–279. https://doi.org/10.3354/meps271267
Moser HG, Boehlert GW (1991) Ecology of pelagic larvae and juveniles of the genus Sebastes. Environ Biol Fish 20:203–224. https://doi.org/10.1007/978-94-011-3792-8_20
Munk KM (2001) Maximum ages of groundfishes in waters off Alaska and British Columbia and considerations of age determination. Alsk Fish Res Bull 8(1):12–21
O’Connell CP (1953) The life history of the Cabezon Scorpaenichthys marmoratus (Ayres). Fish Bull 93:1–76
Ottmann D, Grorud-Colvert K, Huntington B, Sponaugle S (2018) Interannual and regional variability in settlement of groundfishes to protected and fished nearshore waters of Oregon, USA. Mar Ecol Prog Ser 598:131–145. https://doi.org/10.3354/meps12616
Panfili J, Pontual H, Troadec H, Wright PJ (eds) (2002) Manual of fish sclerochronology. Ifremer-IRD co-edition, Brest, France, p 464
Piner KR, Hamel OS, Menkel JL, Wallace JR, Hutchinson CE (2005a) Age validation of Canary Rockfish (Sebastes pinniger) from off the Oregon coast (USA) using the bomb radiocarbon method. Can J Fish Aquat Sci 62:1060–1066. https://doi.org/10.1139/f05-082
Piner KR, Haltuch MA, Wallace JR (2005b) Preliminary use of oxygen stable isotopes and the 1983 El Niño to assess the accuracy of aging Black Rockfish (Sebastes melanops). Fish Bull 103:553–558. 1834/26218
Rasmuson LK, Kautzi LA, Aylesworth L, Wilson MN, Grorud-Colvert K (2019) Age reading of Cabezon (Scorpaenichthys marmoratus): 1) comparison of thin-section and break-and-burn methods and 2) comparison of growth curve fits. ODFW Information Report #2019–04, Oregon Department of Fish and Wildlife, p 49
Reeves SA (2003) A simulation study of the implications of age-reading errors for stock assessment and management advice. ICES J Mar Sci 60:314–328. https://doi.org/10.1016/S1054-3139(03)00011-0
Secor DH, Henderson-Arzapalo A, Piccoli PM (1995) Can otolith microchemistry chart patterns of migration and habitat utilization in anadromous fishes? J Exp Mar Biol Ecol 192(1):15–33. https://doi.org/10.1016/0022-0981(95)00054-U
Shiao J-C, Itoh S, Yurimoto H, Iizuka Y (2014) Liao Y-C (2014) Oxygen isotopic distribution along the otolith growth axis by secondary ion mass spectrometry: applications for studying ontogenetic change in the depth inhabited by deep-sea fishes. Deep-Sea Res I(84):50–58. https://doi.org/10.1016/j.dsr.2013.10.006
Six LD, Horton HF (1977) Analysis of age determination methods for Yellowtail Rockfish, Canary Rockfish, and Black Rockfish off Oregon. Fish Bull 75(2):405–414
Stein D, Hassler TJ (1989) Species profiles: life histories and environmental requirements of coastal fishes and invertebrates (Pacific southwest): Brown Rockfish, Copper Rockfish, Black Rockfish. U.S. Fish and Wildlife Service Biological Report 82: (11.113)
Stewart KP, McMahon TE, Koel TM, Humston R (2021) Use of otolith microchemistry to identify subbasin natal origin and use by invasive Lake Trout in Yellowstone Lake. Hydrobiol 848(10):2473–2481. https://doi.org/10.1007/s10750-021-04568-z
Sturrock AM, Trueman CN, Milton JA, Waring CP, Cooper MJ, Hunter E (2014) Physiological influences can outweigh environmental signals in otolith microchemistry research. Mar Ecol Prog Ser 500:245–264. https://doi.org/10.3354/meps10699
Swanson HK, Kidd KA, Babaluk JA, Wastle RJ, Yang PP, Halden NM, Reist JD (2010) Anadromy in Arctic populations of Lake Trout (Salvelinus namaycush): otolith microchemistry, stable isotopes, and comparisons with Arctic Char (Salvelinus alpinus). Can J Fish Aquat Sci 67(5):842–853. https://doi.org/10.1139/F10-022
Thompson JE, Hannah RW (2010) Using cross-dating techniques to validate ages of Aurora Rockfish (Sebastes aurora): estimates of age, growth, and female maturity. Environ Biol Fish 88(4):377–388. https://doi.org/10.1007/s10641-010-9653-6
Thorrold SR, Campana SE, Jones CM, Swart PK (1997) Factors determining δ13C and δ18O fractionation in aragonitic otoliths of marine fish. Geochim Cosmochim Acta 61(14):2909–2919. https://doi.org/10.1016/S0016-7037(97)00141-5
Valley JW, Kita NT (2009) In situ oxygen isotope geochemistry by ion microprobe. In: Fayak M (ed) Secondary ion mass spectrometry in the earth sciences. Mineralogical Association of Canada Short Course (41), Toronto, Ontario, pp 19-63
Wallace RK, Fletcher KM (1997) Understanding fisheries management: a manual for understanding the federal fisheries management process, including analysis of the 1996 Sustainable Fisheries Act, 2nd ed., Mississippi-Alabama Sea Grant Consortium publication 00-05, p 62
Weidel BC, Ushikubo T, Carpenter SR, Kita NT, Cole JJ, Kitchell JF, Pace ML, Valley JW (2007) Diary of a Bluegill (Lepomis macrochirus): daily δ13C and δ18O records in otoliths by ion microprobe. Can J Fish Aquat Sci 64:1641–1645. https://doi.org/10.1139/f07-157
Weidman CR, Millner R (2000) High-resolution stable isotope records from North Atlantic Cod. Fish Res 46:327–342. https://doi.org/10.1016/S0165-7836(00)00157-0
Wetzel CR, Langseth BJ, Cope JM, Whitman AD (2021) The status of Copper Rockfish (Sebastes caurinus) in U.S. waters off the coast of Oregon in 2021 using catch and length data. Pacific Fisheries Management Council, Portland, Oregon, p 134
Wright PJ, Panfili J, Morales-Nin B, Geffen AJ (2002) Otoliths. In: Panfili J, Pontual H (de), Troadec H, Wright PJ (eds) Manual of fish sclerochronology. Brest, France: Ifremer-lRD coedition, pp 31–57
Wyllie-Echeverria T (1987) Thirty-four species of California rockfishes: maturity and seasonality of reproduction. Fish Bull 85(2):229–250
Acknowledgements
We are grateful to Will Fennie, Megan Wilson, Kirsten Grorud-Colvert and Su Sponaugle, Oregon State University, for providing age and otolith measurement data for young-of-the-year Black Rockfish and Cabezon. Patrick McDonald and Betty Kamikawa, Pacific States Marine Fisheries Commission, read otolith chronologies and provided peak counts. Ryan Fields (Oregon Department of Fish and Wildlife) assisted in figure preparation. We thank Dave Fox and Jessica Watson (Oregon Department of Fish and Wildlife) and Jessica Miller (Oregon State University) for their helpful reviews which improved the manuscript.
Funding
This work was funded through the Interjurisdictional Fisheries Act, a financial assistance program administered by the National Oceanic and Atmospheric Administration’s (NOAA) National Marine Fisheries Service (NMFS); NOAA Award No. NA 19NMF4070280, Subcontract: 20-35G / Job 1151G.20.
Author information
Authors and Affiliations
Contributions
The study conception was developed by Leif Rasmuson, and all authors contributed to study design. Funding acquisition was by Leif Rasmuson. Material preparation and data collection from SIMS analysis were performed by Richard Stern. All other material preparation and data collection, and all data analyses were performed by Mark Terwilliger. The first draft of the manuscript was written by Mark Terwilliger, Leif Rasmuson and Richard Stern, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
This research did not include the use of live animals. All fish used in this study were sampled as part of ongoing commercial and recreational fisheries sampling. This sampling project collected data to monitor commercial and recreational fisheries so as to promote sustainable fisheries management. Because no fish were harvested intentionally for this project, no approval of research ethics committees was required to accomplish the goals of this study.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Terwilliger, M.R., Rasmuson, L.K. & Stern, R.A. Age validation of Black Rockfish, Copper Rockfish, and Cabezon using secondary ion mass spectrometry (SIMS) to elucidate seasonal patterns in otolith stable oxygen isotopes. Environ Biol Fish 106, 553–573 (2023). https://doi.org/10.1007/s10641-023-01392-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-023-01392-3