
Abstract
The impacts of mine contaminants on ecological connectivity in rivers and streams are poorly documented globally. We used acoustic telemetry to evaluate and refine conceptual models of fish movement in Magela Creek, a stream in the wet-dry tropics of Australia. This creek receives wastewater discharge from a nearby uranium mine, and a secondary objective was to describe behavioural responses of fish to one such discharge event. Of 55 fish (black bream Hephaestus fuliginosus, saratoga Scleropages jardinii, sharp-nose grunter Syncomistes butleri) tagged in dry season refuge pools 18 km upstream of the mine lease area (RPA [Ranger Project Area]), 16 (29%) moved downstream after the first wet season flows, using the RPA as habitat for 3–5 months before moving upstream to their previous locations as flows receded. Of 39 fish (spangled perch Leiopotherapon unicolor, barred grunter Amniataba percoides, black catfish Neosilurus ater) tagged ~ 8–12 km downstream of the RPA in the late wet season, only two were subsequently detected in the RPA. Direct and camera-based observations of 12 species of upstream-migrating fish during mine-water discharge in the late wet season showed no evidence of mine-water avoidance. Our results demonstrate that Magela Creek provides wet season habitat for fish within the RPA and acts as a migration pathway that connects lowland reaches and floodplains to upstream dry season refuges. Use by fish of waterbodies within the RPA highlights the need to manage the site to ensure that future contaminant egress and water quality do not adversely affect fish migration and habitat suitability.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Connectivity plays a critical role in facilitating ecological processes across the full range of global ecosystem typologies (Crooks and Sanjayan 2006). In freshwater ecosystems, a wide range of human activities can adversely impact ecological connectivity, including dams and other barriers, consumptive water use, habitat destruction, and pollution from a wide variety of sources (see Crook et al. 2015). It is well documented that pollutants from mining have caused significant impacts on freshwater ecosystems globally (Wright et al. 2017). However, most research on the effects of mining contaminants on freshwater biota has focussed on bioaccumulation and the physiological (and to a lesser extent behavioural) effects of contaminants, acidification, and sedimentation (Byrne et al. 2012; Hogsden and Harding 2012; Affandi and Ishak 2019). The impacts of mining on the broad-scale movements of fish and other biota—for example, by contaminating critical migration pathways—and the implications for population and ecosystem processes remain poorly documented.
The Ranger uranium mine commenced operations in 1981 and was one of Australia’s largest uranium producers over the 40-year period of ore processing. The mine site is located within the Alligator Rivers Region, Northern Territory, Australia, and is surrounded by the World Heritage–listed, Kakadu National Park. Rehabilitation works at the mine in preparation for closure are due to be completed by 2026, and the rehabilitated landform is predicted to become a source of both contaminated surface water runoff, but particularly exfiltrating groundwater with elevated electrical conductivity (EC) mainly associated with magnesium sulfate (MgSO4). This solute is derived from the Mg‐based minerals (such as chlorite schist) present in the waste rock used to construct the landform and to cap the pits. Solute egress modelling predicts that within 10 years of the mine closure, the reach of Magela Creek on the mine lease area (hereafter, RPA [Ranger Project Area]) and Coonjimba Billabong will have Mg concentrations greater than the site-specific guideline value for surface waters (2.9 mg/L Mg) and that, without mitigation, concentrations above this limit may remain for extensive periods of time (RMCP 2020, Supervising Scientist 2021). The guideline value was derived from laboratory, field, and semi-field experiments, and is a concentration that theoretically protects the majority (i.e. 99%) of aquatic species from chronic sublethal toxicity, if not exceeded.
Saline (MgSO4) plumes associated with surface and groundwater egress to Magela Creek have been identified as potential threats to ecological connectivity and fish migration in Magela Creek via direct toxicity and avoidance behaviour (Bishop et al. 1995; Humphrey et al. 2016). However, while there is some existing information on lethal and sublethal toxicity of MgSO4 for native fish of the region (van Dam et al. 2010; Pease et al. 2021), concentrations of MgSO4 that may lead to avoidance behaviour under field conditions are currently unknown. While there are no statutory environmental requirements that site-specific guideline values must be met within the RPA during either mine operations or after rehabilitation, concentrations of contaminants must also not reach levels that would disrupt ecological connectivity, including fish movement, through and past the lease boundaries. Contaminant effects upon macroinvertebrate communities in the creek channels within the RPA (i.e. trophic groups potentially sustaining migrating fishes) have been the subject of a separate study (Mooney et al. 2023).
Detailed studies of fish movements in the sand channel reaches of Magela Creek near the Ranger mine were conducted in the 1980s and 1990s using visual observations, netting and trapping (summarised in Bishop and Forbes 1991; Bishop et al. 1995; see Figures S1, S2, and S3 in Supplementary Material 1 for conceptual representations of fish movement patterns in Magela Creek). The small size of the shallow creek channels, contrasting sandy stream bed, and high water clarity lend themselves to visual approaches to fish observation (human or camera). Bishop and Forbes (1991) and Bishop et al. (1995) reported upstream migrations along the creek of up to 27 fish species towards and at the end of the wet season, numerically dominated by two small-bodied species, chequered rainbowfish Melanotaenia splendida inornata and northwest glassfish Ambassis sp. (peaking at > 100,000 fish/hour). Based on declining numbers from downstream to upstream sites, Bishop et al. (1995) inferred that the likely source of these fish was the extensive floodplain system downstream of Mudginberri Billabong, and that in addition to dry season floodplain refuges (non-flowing pools known in Australia as ‘billabongs’), they were moving large distances along the creek channel to occupy other refuges below and within the RPA, and in upstream reaches below the Arnhem Land Plateau escarpment (see Fig. 1 for described locations).

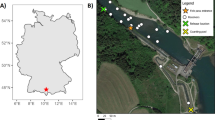
Top: map of the study region showing location of Ranger Uranium Mine. Bottom: aerial photograph showing the locations of the mine-water discharge point MG001 and the acoustic receivers (yellow dots) installed during the study. Magela Crossing is located immediately downstream (north) of Mudginberri Billabong
Larger-bodied terapontid species, including black bream Hephaestus fuliginosus juveniles, spangled perch Leiopotherapon unicolor, and barred grunter Amniataba percoides, were also observed moving in the sand channels upstream in the late wet season (Bishop and Forbes 1991; Bishop et al. 1995). Based on observations of increases in the number of terapontids migrating in the sand channels from lowland to upstream sites near the escarpment, Bishop et al. (1995) concluded that these species occupy the lowland reaches of Magela Creek (i.e. sand channels and adjacent backflow billabongs upstream of Mudginberri Billabong) during the wet season, before moving upstream to refuge billabongs below the escarpment where they spend the dry season. These observations of large-scale fish migration within and through the RPA suggest that future egress of saline groundwater to surface waters from the rehabilitated mine site has the potential to reduce connectivity between upstream refugia and lowland floodplains, thus, interrupting a key process that underpins the functioning and productivity of the river-floodplain ecosystem (see Pettit et al. 2017).
While the previous work on fish migration in Magela Creek described the general seasonal movements of fish populations in Magela Creek, the reliance on visual observations and trapping data at set locations limits the inferences that can be made regarding ecological connectivity and the potential future impacts of mine-related contaminants. For example, data on fish moving past specific locations (see Bishop et al. 1995) do not provide information on the distances travelled or areas visited by individual fish or, importantly, the significance of movement behaviour at the population level: i.e. do the majority of individuals migrate large distances or just a small proportion? This more detailed information on fish movement is critical to improving our understanding of the population-level consequences of any impediments to connectivity caused by mine contaminant egress. This would increase our capacity to assess potential legacy impacts of the mine on aquatic biota, develop future mitigation and monitoring activities, and enhance knowledge of intraspecific fish movement patterns at landscape scales.
The current study used acoustic telemetry to examine the individual-level movements of six native fish species in Magela Creek (black bream, spangled perch, barred grunter, saratoga Scleropages jardinii, sharp-nose grunter Syncomistes butleri, and black catfish Neosilurus ater). To our knowledge, this is the first telemetric study of the movements of these species, although a recent radio-tracking study in the Katherine River, Northern Territory, examined the short-term (10 days) dry season habitat preferences of black bream (Crook et al. 2021a, b). There has also been other recent telemetry-based research examining the movement patterns of barramundi Lates calcarifer, forktail catfish Neoarius leptaspis, and diamond mullet Planiliza ordensis in the Alligator Rivers Region (Crook et al. 2016, 2020; Saboret et al. 2021).
Fish were tagged during the dry season in a refuge billabong upstream of the mine site and at the end of the wet season in the creek channel downstream of the mine, and their movements tracked over two dry–wet season cycles. We also undertook direct visual and camera-based observations of the behavioural responses of fish to a controlled mine-water discharge in Magela Creek. The primary objective of the study was to examine the seasonal movements of individual fish in the Magela Creek system to evaluate and refine key components of the system’s existing model of fish movement (Bishop and Forbes 1991; Bishop et al. 1995). Specifically, we aimed to use data on individual fish movements to confirm the inferences from Bishop and Forbes (1991) (1) that fish from upstream refugia undertake distinct downstream migrations into the lowland reaches of Magela Creek during the early wet season, followed by return upstream migrations in the mid-late wet season, and (2) that fish from upstream refugia temporarily colonise off-channel, backflow billabongs during the wet season before moving back to the main channel and dry season refugia. (Other components of the system’s existing model of fish movement, including the large-scale migrations of small-bodied fish species (especially chequered rainbowfish and northwest glassfish) from floodplain recruitment zones and upstream past the Ranger mine site, were not specifically examined.) A secondary objective of the study was to describe the behavioural responses of fish to mine-water discharge in Magela Creek and to determine the extent of wet season use of seasonally inundated water bodies within the RPA that are subject to mine-water contamination. Based on the findings of the study, we further refine the existing conceptual model of fish movement for this system and identify key periods and flow conditions when fish are residing within the RPA and may be exposed to mine-related contaminants.
Materials and methods
Study site
The study was conducted between November 2018 and June 2020 in the mid‐upper reaches of Magela Creek, a major tributary of the East Alligator River in the Northern Territory, Australia. The study region experiences a wet-dry tropical climate characterised by periods of heavy and prolonged rainfall and high stream discharge from December to April, and generally dry conditions for the remainder of the year. Magela Creek is an unregulated, seasonally flowing sand‐bed stream which has its headwaters in the Arnhem Land Plateau and flows through both the Ranger Uranium Mine lease area (RPA) and Kakadu National Park (Fig. 1). Wet season rainfall was well below the long-term average of 1554 mm in both years of the study (2018/19 = 1237 mm, 2019/20 = 1073 mm, Jabiru Airport weather station) resulting in lower-than-average stream discharge in Magela Creek (Fig. 2).

Magela Creek hydrology over the study period. Date format is day/month/year
The surface waters of streams adjacent to the Ranger mine (including Magela Creek) have naturally low pH (~ 5.5–6.5), ionic strength (Na+ < 2 mgL; Mg2+ < 1 mg/L, and SO42– < 0.5 mg/L, ~ 5–20 µS/cm) and hardness (~ 5 mg/L as CaCO3), and relatively high water temperatures throughout the year (~ 26–31° C seasonal range; Harford et al. 2015; Mooney et al. 2019). Resident biota can be highly susceptible to changes in the ionic environment given that they occur in a naturally low ionic strength environment (van Dam et al. 2010; Humphrey and Chandler 2018; Mooney et al. 2019).
Bowerbird Billabong and Mudginberri Billabong were selected to represent key in-channel refuge habitats upstream and downstream of the RPA respectively (Fig. 1). Between these reaches of the creek, there are no other permanent surface waters in the main channel, although a series of shallow off-channel, or ‘backflow’, billabongs (e.g. Georgetown, Coonjimba billabongs; see Fig. 1) generally retain some water throughout the dry season near and downstream of the RPA. Bowerbird Billabong is ~ 29 km upstream of Mudginberri Billabong and is an escarpment/rockpool billabong ~ 40 m wide and ~ 1 km long. The billabong has steep vegetated banks, with coarse sandy or rock substrate. Forming in the upper catchment and commencing below Djurrubu Falls (a large waterfall that acts as a complete barrier to fish movement), Bowerbird Billabong is part of a 10 km sand and rock pool-riffle sequence that extends through sandstone escarpments and outcrops before meeting the sand channels of Magela Creek (Fig. 1). There is no vehicle access to Bowerbird Billabong, so all access to the field site was undertaken using helicopters. Mudginberri Billabong is a naturally enlarged section of the main channel of Magela Creek, located at the downstream terminus of the sand channel below the RPA and the upstream extent of the extensive Magela floodplain. Mudginberri Billabong has steep vegetated banks, a coarse sandy or sandy-detritus substrate and is approximately 1 km long with a maximum width of ~ 90 m.
Acoustic telemetry
Acoustic telemetry was used to examine the movements of fish collected and tagged from Bowerbird Billabong and Mudginberri Billabong. The first batch of fish tagged were large-bodied species (saratoga, black bream, sharp-nose grunter) collected from Bowerbird Billabong in November 2018 using hook-and-line fishing. Acoustic transmitters (V9-2x-180 kHz, Vemco, Nova Scotia) were surgically implanted into the peritoneal cavities of 24 saratoga, 26 black bream, and five sharp-nose grunter (Table 1). The V9 transmitters used for these fish had an estimated battery life of ~ 700 days, allowing the movements of tagged fish to be tracked over two wet–dry season cycles (November 2018 to June 2020).
The second batch of fish tagged were smaller-bodied fish species (spangled perch, barred grunter, black catfish) collected in April and May 2019 using cast nets and hook-and-line fishing. These fish were collected from Magela Crossing ~ 100 m downstream of Mudginberri Billabong, from the upstream end of Mudginberri Billabong, and from the main channel of Magela Creek up to 4 km upstream of Mudginberri Billabong. The objective of this part of the study was to examine individual-level upstream movement by smaller-bodied fishes during the recessional flow period at the end of the wet season, as previously reported by Bishop et al. (1995). Acoustic transmitters (V5-1x-180 kHz, Vemco, Nova Scotia) were surgically implanted into the peritoneal cavities of 17 spangled perch, 13 barred grunter, and 9 black catfish (Table 2). The V5 transmitters used for these fish had an estimated battery life of 91 days, allowing the movements of tagged fish to be tracked over a single recessional flow period (April to August 2019). Data were screened and all false and single detections were scrutinised and removed where appropriate prior to analysis.
Fish tagged with acoustic transmitters were first anaesthetised with Aqui-S (0.03 ml/L), weighed (g) and measured (total length and fork length to the nearest mm). Fish were then placed in a V-shaped foam holding cradle lined with a wetted absorbent cloth. The transmitters were disinfected (Hibitane®) and rinsed with sterile saline prior to implantation. Where necessary, several scales were removed from the ventral surface anterior to the anal vent and slightly offset from the midline, and the area swabbed with an antiseptic solution (Betadine®). An incision was made in an anterior–posterior orientation into the peritoneal cavity using a sterile scalpel. The transmitters were inserted into the peritoneal cavity through the incision, with the incision then closed using a single layer closure using 2–3 interrupted sutures placed into the musculature 2–3 mm beneath the skin (2.0 metric, absorbable monofilament, 26 mm swaged needle, Ethicon). Antiseptic liquid spray (Betadine®) was then applied to the area. The gills were irrigated with fresh stream water throughout the procedure (2–4 min). After the procedure, fish were held in fresh stream water until they resumed normal posture and gill movement and were then released at the site of collection.
An array of 10 acoustic receivers (VR2W-180 kH, Vemco, Nova Scotia) was deployed from the downstream end of Mudginberri Billabong to the upper end of Bowerbird Billabong (Fig. 1). Receivers were attached to riparian trees using plastic-coated wire. Each receiver was weighed down with a heavy-gauge metal chain and a float was attached to ensure that the acoustic hydrophone remained in an upright position. We did not assess the transmitter detection range at the study sites; however, the range of the transmitters used is usually > 150 m (Innovasea 2020) which far exceeds the typical stream width of the system. The mid-reaches of the study area in Magela Creek consist of several braided channels, and receivers were generally placed along the western bank of the western channel, as previous studies (e.g. Bishop et al. 1995; ARRRI 1985) had demonstrated this was the preferred migration pathway for fish in the creek system. Notwithstanding, it is possible that some movements were not detected if fish used alternative channels to the receiver locations as migration pathways. Data were downloaded from the receivers in December 2018, March 2019, May 2019, August 2019, November 2019, September 2020, and November 2020.
To examine the likelihood of tagged fish being present in the RPA under different stream discharge conditions, each fish’s daily location was assigned a zero (outside of RPA) or one (inside of RPA). We then used generalised additive mixed models (GAMMs) fitted with a binomial distribution to model, for each species separately, the probability of individual saratoga or black bream being within the RPA as a function of river discharge. There were insufficient data to run similar analyses for the other species. A random intercept was included for each fish to account for the repeated measures nature of the data and a cubic regression smoother was fitted to stream discharge. 46.4% of days had zero discharge and data for the remaining discharges were heavily skewed towards low-flow days. We therefore applied a 4th root transformation to discharge data to reduce the influence of the skewing, improve model fit, and aid model interpretation. Model comparison was conducted using Akaike Information Criterion corrected for small sample size (AICc).
Fish behavioural responses to mine wastewaters in Magela Creek
A field-based examination of fish behaviour associated with an operational mine-water release from the mine discharge point MG001 on Magela Creek (Fig. 1) was undertaken on 3 March 2021. Release-quality, mine pond waters are discharged to Magela Creek at this location by pumping the mine waters from the mine site into a concrete bund, where they pass through release ports and spill a short distance down the bank into the creek. ‘Release-quality’ refers to contaminant concentrations in pond waters that may exceed the guideline value at the point of discharge, but which meet the guideline value at the compliance point, ~ 5 km downstream of the discharge point, after mixing. Visual observations were made from the bank and using underwater videography. Visual observations from the bank concentrated on identifying any clear avoidance or attraction by fish to the mine-water discharge (i.e. directed swimming away from or towards/into the mine discharge water). In addition, one underwater camera (GoPro 8 s) was deployed on the western bank and one on the eastern bank of the channel at this site, with cameras mounted on submersible stands and left to record for ~ 70 min. Mine waters enter the creek on the western bank, along the preferential migration path for small- and mid-bodied fish species (authors pers. obs., Bishop et al. 1995). Video observations focused on determining species moving past the discharge point and observing any behavioural avoidance of the discharge water. Video time that was analysed was standardised across all videos at 60 min, with the first 5 min as settling time (no data recorded). Abundances were measured using MaxN, the maximum number of fish for a given species present in one frame for the 60-min analysis period following methods described by Ebner and Morgan (2013) and King et al. (2018). Videos were processed by trained staff and identification was taken to species level where possible.
Water chemistry samples were collected prior to fish observations and video recordings from inside the release bund, Magela Creek eastern bank, and Magela Creek western bank where mine water mixed with Magela Creek water. Water samples and blank and procedural blank samples were filtered (0.45 µm) and analysed for a metal and major ion suite (Al, Co, Cr, Cu, Fe, Mn, Ni, Pb, U, Zn, Ar, Bo, Ba, Br, I, Li, Ca, K, Mg, and Na). Water quality parameters (pH, DO, EC, turbidity, and temperature) were measured in situ (EXO1 multiparameter sonde, Xylem Analytics Australia) at the same location as the water chemistry samples, as well as immediately upstream of the mine discharge. The association between tagged fish entering on-site backflow billabongs, Coonjimba and Georgetown, during the wet season, and water quality of the billabongs was examined using matching water chemistry data acquired from Energy Resources of Australia for the visitation periods (Supplementary Material 1).
Results
Seasonal movements of tagged fishes in the stream system
Downstream movement from upstream refugia
Fifty-four of the 55 fish tagged in Bowerbird Billabong were detected during the study, with a single sharp-nose grunter not detected after tagging (Table 1). Transmitters were detected for up to 570 days, and the average tracking duration of detected fish was as follows: 415 days (n = 24, range: 17–570 days) for saratoga; 406 days (n = 26, range: 9–570 days) for black bream; and 415 days (n = 4, range: 32–552 days) for sharp-nose grunter. More than 5.6 million detections were recorded during the study, with 42 fish detected more than 10,000 times and 16 fish detected more than 100,000 times (Table 1). The fate of fish detected for only short periods is unknown, but could be attributable to transmitter failure, tag expulsion, mortality, or long-term occupation of areas outside the range of the receivers.
Ten (42%) saratoga, five (19%) black bream, and one (25%) sharp-nose grunter were detected in the RPA during the study, with the remaining fish detected only on the receivers deployed near the tagging site at Bowerbird Billabong (Table 1, Figs. 3, 4). Fish that were detected only in Bowerbird Billabong (i.e. upstream of the RPA) were recorded very frequently during the dry season but had much lower detection rates during wet season flow events. This behaviour is suggestive of a wider range of movements in the reaches above the RPA during the wet season for these three species (Fig. 5).

Movements of the 10 saratoga (SAR) that entered the RPA during the study. Blue symbols represent tag detections, and a blue line connects consecutive detections. The orange-shaded area shows the approximate locations of the RPA and the broken line shows the discharge location (MG001). The orange line shows creek discharge. Tag codes, standard lengths, and weights are shown for each fish. Date format is day/month/year

Movements of the five black bream (SGR) and one sharp-nose grunter (SNG) that entered the RPA during the study. Blue symbols represent tag detections, and a blue line connects consecutive detections. The orange-shaded area shows the approximate locations of the RPA and the broken line shows the discharge location (MG001). The orange line shows creek discharge. Tag codes, standard lengths, and weights are shown for each fish. Date format is day/month/year

Plot showing detections of all tagged fish (all species) at the downstream Bowerbird Billabong acoustic receiver over the study period. Each row of data points shows the detections for an individual fish and the solid line shows creek discharge. This figure demonstrates the reduction in detections in Bowerbird Billabong during periods of high flow, which is attributed to migration into the RUM lease and localised movements upstream of the lease. Date format is day/month/year
Downstream movement into the RPA for all three species was initiated soon after the first wet season flows and was strongly related to stream discharge, with the likelihood of tagged saratoga (ΔAICc to intercept only model: 2190.1) and black bream (ΔAICc to intercept only model: 638.1) being present in the RPA increasing substantially at flows greater than ~ 50 ML/day (Fig. 6a, b). Three of the five black bream that moved into the RPA moved as far downstream as the discharge point at MG001, but none were detected any further downstream in either year of the study (Fig. 4a–e). Of the 10 saratoga that moved into the RPA, seven reached the MG001 discharge point and six of these subsequently moved further downstream (Fig. 3). One of these six fish (Fig. 3b) moved downstream beyond the RPA to Mudginberri Billabong, where it was detected on the most downstream receiver on 6 February 2019 and again on 24 March 2019. It appears likely that this fish had moved further downstream into the lower floodplain reaches of the Magela Creek system during this time, before moving back upstream to Bowerbird Billabong by 5 April 2019. The single sharp-nose grunter that moved into the RPA reached the discharge point in the first year (Fig. 4f). In the second year, this fish moved through the RPA and was last detected on the most downstream receiver in Mudginberri Billabong, potentially moving further downstream from Mudginberri Billabong.

Results of generalised additive mixed modelling showing the probability of tagged saratoga (a) and black bream (b) from Bowerbird Billabong occurring in the RPA relative to creek discharge. The vertical line and solid circle highlight the minimum flow upon which fish were detected in the RPA. Note the x axis is presented on the 4th root scale to facilitate ready interpretation of each species’ response to changes in discharge given the highly skewed nature of discharge data. Smaller jittered points are individual observations of fish position inside (Pr = 1) or outside (Pr = 0) of the RPA
Return upstream movement by large-bodied fishes
A striking feature of the data was the accurate upstream homing behaviour exhibited towards the end of the wet season. All five of the black bream that moved into the RPA made return movements back to the same ~ 1 km reach of Bowerbird Billabong in the first year of the study, with three fish also exhibiting similar behaviour the following year (the transmitters on the other two fish ceased being detected during the second year). Most black bream made multiple (up to five) return movements between Bowerbird Billabong and the RPA in the same year (Fig. 4). Six saratoga (including the fish that reached Mudginberri Billabong) also timed their return to Bowerbird Billabong as wet season flows decreased in the first year (Fig. 3). Return movement between Bowerbird Billabong and the RPA was less frequent for saratoga than black bream, with only two of the ten saratoga undertaking two return movements during the wet season (Fig. 3b, c). Another two saratoga were detected in the upstream region of the RPA as flows receded in the first year (but not in Bowerbird Billabong) in the 2019 dry season, suggesting that these fish may have moved into the area of permanent water just downstream of the Bowerbird Billabong receivers prior to the 2019 dry season (Fig. 3f, g). Two saratoga completed return movements in the second year, while one that had moved downstream the previous year was not detected in the RPA in the second year. Transmitters for the remaining saratoga that moved into the RPA in the first year ceased being detected at various stages during the second year. The single sharp-nose grunter that entered the RPA made three return movements in the first year before moving downstream out of the study area during high flows in the second year (Fig. 4f).
Upstream movement by smaller-bodied fishes
Of the nine black catfish tagged at Magela Crossing, immediately downstream of Mudginberri Billabong in late April and early May 2019, eight were subsequently detected in Mudginberri Billabong between 12 and 108 days after tagging (Table 2). One of these fish (tagged on 26 April 2019) was detected on 7 May 2019 within the RPA ~ 8 km upstream of Mudginberri Billabong (MC6) and on 8 May 2019 (MC1) (Fig. 7a). Flows during this upstream movement ranged from ~ 150 to 250 ML/day (Fig. 7a). This fish was not detected on the main channel receivers between MC1 and MC6 (MC3, MC4). No other black catfish were detected in the RPA during the study.

Movements of the one black catfish (BCF) and one barred grunter (BGR) that moved upstream into the RPA on recessional flows at the end of the wet season. Blue symbols represent tag detections and the blue line connects consecutive detections. The orange-shaded area shows the approximate location of the Ranger RPA and the broken line shows the discharge location (MG001). The orange line shows creek discharge. Tag codes, standard lengths, and weights are shown for each fish. Date format is day/month/year
Of the eight barred grunter tagged at Magela Crossing, five were subsequently detected in Mudginberri Billabong for 17–106 days after tagging (Table 1). The four fish tagged at the upstream end of Mudginberri Billabong were all subsequently detected in Mudginberri Billabong for 5–96 days after tagging. None of these fish were detected upstream of Mudginberri Billabong. The single barred grunter tagged upstream (4 km) of Mudginberri Billabong was detected three times on 14 May 2019 in the RPA at MC6 and 18 times on 25 May 2019 at MC1 (Fig. 7b). Flow in Magela Creek was 29 ML/day when this fish was detected at MC6 and 0.2 ML/day when the fish reached MC1 at the upstream end of the RPA (Fig. 7b). Like the black catfish that moved upstream, this fish was not detected on the main-channel receivers between MC1 and MC6 (MC3, MC4).
Of the six spangled perch tagged at Magela Crossing, only one was detected in Mudginberri Billabong (Table 2). The remaining fish presumably spent the recessional flow and subsequent dry season in permanent pools downstream of Mudginberri Billabong. As a relatively small-bodied terapontid, it is also possible that the tagged spangled perch experienced high rates of predation downstream of Mudginberri Billabong. The single spangled perch that moved into the main body of Mudginberri Billabong was detected 14 times on 6 July 2019 on the downstream receiver, 72 days after it was tagged. Of the 11 spangled perch tagged up to 4 km upstream of Mudginberri Billabong, none were detected by the acoustic array.
Use of backflow billabongs
Four individuals of the large-bodied fish were detected in backflow billabongs in the RPA during the study: two saratoga and one black bream in Georgetown Billabong, and one saratoga in Coonjimba Billabong (Figs. 3, 4). Like the observed homing of fish to dry season refugia in Bowerbird Billabong, these fish exhibited remarkable fidelity to specific backflow billabongs across the 2 years of the study, presumably utilising them as wet season feeding grounds. Given that only a small proportion of the Bowerbird Billabong fish population was tagged (and allowing for some possible mortality or transmitter failure post-tagging), these observations suggest that significant numbers of fish may use Georgetown and Coonjimba backflow billabongs as temporary habitat during the wet season.
Saratoga 46780 (Fig. 3c) moved downstream from Bowerbird Billabong into the RPA on 27 January 2019, before moving into Georgetown Billabong 11 h later. It was detected in Georgetown Billabong until early April and made at least two excursions back to the main channel during this time, including one return movement to Bowerbird Billabong, where it was detected from 30 March to 6 April 2019. It was again detected in Georgetown Billabong from 7 to 11 April and returned to Bowerbird Billabong by 17 April 2019, where it spent the 2019 dry season. The estimated residence time in Georgetown Billabong in 2019 was approx. 68 days. The following wet season, this fish moved back downstream again to Georgetown Billabong where it was detected from 27 January until its final detection there on 12 April 2020, an estimated residence time of approx. 77 days.
Saratoga 46646 (Fig. 3g) moved downstream from Bowerbird Billabong and was detected in Georgetown Billabong from 29 January to 8 April 2019, a 70-day period. It was detected on multiple occasions moving within the main channel (MC1 to MC6) during this time. After moving upstream of the RPA over the 2019 dry season (but not detected at Bowerbird Billabong), this fish was again detected back in Georgetown Billabong from 24 January to 8 February 2020, a 16-day period. It then moved back into the main channel where it was last detected at MC6 on 14 April 2020.
The single black bream (46762, Fig. 4c) that used a backflow billabong moved downstream from Bowerbird Billabong into the RPA on 3 January 2019 before moving into Georgetown Billabong on 13 January. It was detected in Georgetown Billabong until 6 March 2019 and then made an excursion back to Bowerbird Billabong where it was detected from 10–21 March 2019. It was again detected in Georgetown Billabong on 26 March 2019 before moving back upstream to Bowerbird Billabong, where it spent the 2019 dry season. The estimated residence time in Georgetown Billabong in 2019 was approx. 53 days. This fish returned to the RPA the following wet season on 10 January 2020 and was last detected in the main channel at MC1 on 19 January 2020.
The only fish that was detected in Coonjimba Billabong during the study was a saratoga (46307, Fig. 3e) that entered the RPA and moved into Coonjimba Billabong on 20 February 2019, where it was detected four times over a 2-min period before returning to the main channel. Thereafter, this fish was detected infrequently in the main channel until 9 April, before moving upstream to Bowerbird Billabong by 15 May 2019 where it spent the 2019 dry season. The following wet season, this fish was sporadically detected in Coonjimba Billabong (628 detections from 31 January to 3 March 2020, a 33-day period). During this time, the fish was detected in the main channel (MC3) on 24 February and 1 and 2 March. It was then detected at MC3 and MC4 from 5 to 10 March, with the last detection on 10 March 2020.
Water quality conditions in the backflow billabongs during the periods of residence by tagged fish are described below.
Fish behavioural responses to mine wastewaters
Responses to mine discharge into Magela Creek
Schools of chequered rainbowfish were observed (visual and camera) swimming through mine-water plumes at the mine discharge site and preferentially swimming up mine-water channels to the base on the bund wall on the western side of the creek. Chequered rainbowfish were also observed inside the bund which contained 100% Retention Pond 1 (RP1) water. The camera on the western bank was placed near the main discharge and mixing channel. Twelve species of fish were recorded at this location (Table 3), with the most abundant fish species recorded being chequered rainbowfish, glassfish, and fly-specked hardyhead (Craterocephalus stercusmuscarum). None of the fish observed at this location showed any signs of avoidance behaviour when moving upstream through the mine discharge waters. The camera on the eastern bank recorded five species and overall fish abundance (MaxN) was much lower than that recorded on the western bank (Table 3). Magela Creek has naturally low EC as represented by the upstream and eastern bank EC measurements (Table 4). Water contained within the release bund had an EC of 130 µS/cm and elevated concentrations of Mn, U, Mg, Na, K, and Ca compared to concentrations in Magela Creek. When mixed with Magela Creek water, mine waters were diluted, reducing the EC and concentrations of mining-related contaminants (Tables 4 and 5).
Fish residence in billabongs subject to elevated mine-related contaminants
During the periods in which the two tagged saratoga and one black bream were present in Georgetown Billabong, water quality sampling in 2019 showed a mean Mg of 3.8 mg/L (maximum = 5.75 mg/L), mean Mn of 7.4 µg /L (maximum = 20.5 µg/L), and mean DO of 4.5 mg/L (minimum = 0.33 mg/L) (Supplementary Material 1). In 2020, when these fish were resident in Georgetown Billabong, mean Mg was 2.4 mg/L (maximum = 3.7 mg/L), mean Mn was 7.0 µg /L (maximum = 13.8 µg/L), and mean DO was 3.8 mg/L (minimum = 2.5 mg/L). In the case of the single saratoga that was detected in Coonjimba Billabong, for the period of its very brief visit on 20 February 2019, Mg was estimated to be 8.3 mg/L, Mn 20 µg/L, and DO 2.7 mg/L (from measurements made on 18 and 25 February 2019). During the extended period of visitation to Coonjimba Billabong by this same fish in 2020, mean Mg was 9.3 mg/L (maximum = 11.20 mg/L), mean Mn was 41.4 µg/L (maximum = 107 µg/L), and mean DO was 2.7 mg/L (minimum = 0.76 mg/L).
Discussion
Seasonal movements
The current study provides new information on the movements of the six study species, including their patterns of spatial and temporal residency in the Magela Creek channel and backflow billabongs in the RPA (see Figures S1, S2, and S3 in Supplementary Material 1 for refined conceptual representations of fish movements in Magela Creek). Our findings broadly corroborate the inferences of Bishop and Forbes (1991) and Bishop et al. (1995), based on visual observations, (1) that large-bodied fish move into the lowland reaches of Magela Creek (i.e. the RPA) during the early wet season before moving upstream to dry season refuge billabongs in the mid-late wet season, and (2) that a proportion of fish which migrate downstream from upstream refugia during the wet season temporarily colonise off-channel, backflow billabongs before moving back to their dry season refugia in the mid-late wet season.
We found that adults of both terapontid species (black bream and sharp-nose grunter) tagged at Bowerbird Billabong during the dry season, as well as saratoga, undertook large-scale downstream movements between upstream refugia and the lowland reaches of Magela Creek during the wet season. These movements were initiated on a rising hydrograph and there was a high probability (> 85%) that some of the tagged individuals from Bowerbird Billabong were present in the RPA once flows exceeded 50 ML/d. Similar initiation of large-scale movement by fish in response to increasing stream discharge has been widely reported in other systems (e.g. Marshall et al. 2016). Nonetheless, the individual-level information available from telemetry in our study showed that only a minority of the adult population undertook downstream movements into the RPA: most tagged fish remained upstream of the RPA throughout the wet season, with 81% (21/26) of black bream, 58% (14/24) of saratoga, and 66% (2/3) of sharp-nose grunter only detected upstream of the RPA during the study. While this may be associated with a propensity to migrate shorter distances in wet seasons of lower rainfall (such as the two years of the study), the finding is an important refinement to the fish movement model of Bishop and Forbes (1991), which does not account for such interannual variations, nor the variation in the movement propensity of individual fish. For the wet seasons like those experienced during the study, our results suggest that the majority of black bream and saratoga that reside in upstream refuges during the dry season are unlikely to be exposed to mine contaminant egress during the wet season. Even if more extensive downstream movement occurred in higher rainfall wet seasons, dilution of mine contaminants would likely confer greater protection for fishes in the RPA. In the case of diverse spatial migration habits, the risk among fish is spread (sensu the ‘portfolio effect”; Schindler et al. 2015), with individual variation in movement behaviour conferring population-level resilience to any transient effects of contaminant egress on connectivity between upstream refugia and the lowland reaches of the Magela Creek system.
Many of the fish that utilised the channel or backflow billabongs within the RPA during the wet season subsequently returned to Bowerbird Billabong towards the end of the wet season in the first year, and several of these fish repeated the same pattern of movement in the second year of the study. These findings demonstrate accurate homing behaviour by black bream, saratoga, and sharp-nose grunter, and are similar to previous acoustic and radio-telemetry studies of barramundi and forktail catfish in the nearby South Alligator River (Crook et al. 2016; 2020), which reported large-scale migrations onto the inundated floodplain during the wet season and accurate homing to previous locations (refuge billabongs) as the floodwaters receded. High site fidelity by saratoga—not just to dry season refuge areas but to wet season billabong feeding grounds—was also observed in our study, suggesting that these fish exhibit fidelity to specific feeding areas over multiple years. These observations of homing behaviour are another important refinement to the fish movement model of Bishop and Forbes (1991), as they demonstrate that the dispersal of fish—and consequently fish assemblage composition across the landscape—is strongly non-random and is influenced by the habitat preference and sensory capabilities of the resident fish species.
Homing behaviour of this type is likely to have important implications for energetic subsidies across aquatic-terrestrial ecotones in the system by allowing fish to utilise highly productive feeding grounds on inundated, lowland floodplains during the wet season (Bishop and Forbes 1991; Jardine et al. 2012), while maintaining their ability to relocate to suitable dry season refuges, thereby avoiding stranding and desiccation as floodwaters recede (Crook et al. 2021a, b). Telemetry studies in temperate systems have similarly shown that accurate homing to previous locations after large-scale movement associated with high flows is common across a range of taxonomic groups (Cyprinidae, e.g. Lucas et al. 2000; Percicthyidae, e.g. Koehn et al. 2008; Galaxiidae, e.g. David and Closs 2002). While the recent telemetry studies conducted in the Alligator Rivers Region have documented homing behaviour in several species, few other long-term telemetry studies have been conducted in tropical rivers, and the prevalence and importance of site fidelity and homing behaviour by fishes remains an important gap in our knowledge of tropical river ecosystems.
Of the 37 fish tagged in the Mudginberri Billabong area, only one black catfish and one barred grunter moved upstream into the RPA. Observations made while in the field suggested that there were low numbers of upstream-migrating fish on recessional flows in the low-rainfall 2018–19 wet season compared to observations in previous years with higher rainfall and stream discharge (Bishop et al. 1995; Supervising Scientist Branch unpubl. data). The low numbers of fish and rapidly falling stream discharge in the post-wet period in 2019 made it very challenging to collect fish on recessional flows as planned. While the observations of two upstream migrants demonstrate that some migration occurred through the RPA on recessional flows as low as 0.2 ML/d, it is possible that the low flows in the late wet season resulted in a lack of rheotactic response to stimulate the mass upstream migration by fish in Magela Creek. There may have also been a lack of recruitment of small species available to migrate due to a lack of prolonged floodplain inundation in that year. Nonetheless, historical observations of fish movement towards the end of the wet season (Bishop and Forbes 1991; Bishop et al. 1995; Supervising Scientist Branch unpubl. data) provide strong evidence of extensive upstream migration by fish through the RPA at the end of the wet season and suggest that the lowland regions of Magela Creek and the downstream floodplains are important sources of fish into the sand channels and upstream refuges.
Fish behavioural responses to mine wastewaters in Magela Creek
A secondary (albeit limited) objective of the study was to describe the behavioural responses of fishes encountering contaminated mine waters associated with the Ranger mine. Our direct observations and underwater video recording of fish behaviour at the mine discharge site in March 2021 aimed to determine whether a contaminant plume in the Magela sand channels might act as a barrier that inhibits fish migration and connectivity in Magela Creek. We did not find evidence that any of the 12 species observed were deterred by, or avoided, mine wastewater released at the discharge site. In fact, chequered rainbowfish appeared to preferentially swim towards and into the mine discharge channels, and showed no adverse responses to EC of up to 130 µS/cm and Mg concentrations of 11 mg/L.
These observations differ from historical observations of fish avoidance of mine water in the field and laboratory. Visual observations at the MG001 site in March 1985 reported strong avoidance behaviour by upstream-migrating chequered rainbowfish associated with releases of wastewater from Ranger Retention Pond 4 (RP4) (ARRRI 1985). This was inferred from the observation of the lateral movement of fish away from the preferred migration pathway along the western bank of the creek receiving the mine-water plume, and an overall increase in fish moving along the eastern bank of the creek. Subsequent laboratory tests on this species with RP4 water also demonstrated a highly significant avoidance reaction, confirming the field observations. During the same RP4 water releases in March 1985, suppression of reproduction in freshwater mussels downstream of the wastewater plume was observed (ARRRI 1985), while subsequent ecotoxicological testing conducted on RP4 water also showed relatively high toxicity to the freshwater Cladocera, Moinodaphnia macleayi, (ARRRI 1991). These historical biological observations indicate that RP4 water had a higher toxicity than expected based on its metal and major ion concentrations (Table 5, Supplementary Material 1). This toxicity and the historical field effects (fish avoidance and freshwater mussel reproduction suppression) were considered to be associated with an unknown polar organic substance (ARRRI 1991). Although we observed no evidence of avoidance behaviour by fish at mine discharge concentrations of Mg up to 11 mg/L, it should be noted that our observations are based on small numbers of fish and lack temporal replication. Additional monitoring would be required to confirm our preliminary observations. This additional work, through field observations and videography, would also need to assess the implications of fish preferentially swimming towards saline mine waters. For example, if fish were inadvertently ‘lured’ to the contaminated waters of a shallow backflow billabong only to be subjected to poor water quality and/or stranding from water subsidence over the ensuing dry season, this would represent an undesirable outcome.
The two saratoga and one black bream detected in Georgetown Billabong appear to have been exposed to Mg concentrations of up to 5.75 mg/L in 2019 and 3.73 mg/L in 2020. The single saratoga that occupied Coonjimba Billabong was exposed to Mg concentrations estimated at ~ 8.3 mg/L in 2019 and up to 11.2 mg/L in 2020. These levels are below or approximately equal to the Mg concentration of 11 mg/L to which fish showed no behavioural response in the field observations at the mine discharge site. Field observations and EC spot measurements in Coonjimba Billabong (Supplementary Material 1) suggest that connectivity of Coonjimba Billabong to the main Magela Creek occurred in the early wet season of 2019 between 21st and 29th January—the period when large-bodied fish species from Magela Creek could potentially enter Coonjimba Billabong. Both the ~ 3-week delay between billabong inundation/connectivity and detection of the saratoga in the billabong (i.e. 20 February) and lack of any ensuing residence of this fish in the billabong could suggest unfavourable conditions for fish residence between late January and 20th February. In this period, mean Mg in the billabong was 8.1 mg/L (maximum = 8.7 mg/L), mean Mn was 28 µg/L (maximum = 40 µg/L), and mean dissolved oxygen (DO) was 2.4 mg/L (minimum = 1.9 mg/L). While Mg and Mn concentrations in this January–February 2019 period in Coonjimba were no greater than exposure concentrations that occurred during the residence of tagged fish in the billabong in 2020 and in Georgetown Billabong in 2019 and 2020, DO concentrations were notably lower in Coonjimba in 2019 than periods supporting fish residence at other times and locations (2.2 versus 3.2–4.4 mg/L respectively, at the time of initial visitation). These observations could suggest that low DO, or other unmeasured water quality conditions associated with low DO, periodically limit the use of backflow billabongs by fish during the wet season.
Toxicity estimates for Mg and Mn are not currently available for saratoga, black bream, and sharp-nose grunter; however, estimates of sublethal (growth) effects for the sympatric northern purple-spotted gudgeon Mogurnda mogurnda are above the levels observed in the backflow billabongs during the study (7-day growth rate 10% effect concentration [EC10] = 50 mg/L Mg, 80 mg/L Mn; Pease et al. 2021). Furthermore, an annual monitoring program conducted since 1988 has not recorded significant change in the fish assemblage composition in Coonjimba or Georgetown billabongs compared to reference sites (Supervising Scientist 2020). These observations suggest that mine-related contaminants are unlikely to have had major impacts on fish assemblages in the backflow billabongs over recent decades. However, further research is required to determine whether fish avoid backflow billabongs during periods of elevated Mg and co-contaminants, and/or when DO levels are low.
Conclusions
Our study broadly supports the key tenets of the model of fish movement in the Magela Creek system originally proposed by Bishop and Forbes (1991) and adds important refinements, including observations of individual variation in migration propensity and multi-year site fidelity. Our findings further confirm the importance of the sand channel reaches of Magela Creek within the RPA, acting as a migration pathway for fish that connects the lowland reaches and floodplains of the system to the escarpment refuge billabongs. We also show that the sand channels and backflow billabongs are important temporary habitats for fish in the region, with a proportion of tagged fish taking up extended residency in these habitats during the wet season. Previous observational data demonstrate the biodiversity value of the sand channels and backflow billabongs within the RPA, with > 30 species of native fish recorded from the sand channels (Bishop et al. 1995; Supervising Scientist Branch unpubl. data). Based on these observations, we hypothesise that there may be significant consequences to the health of fish assemblages in Magela Creek if future contaminant concentrations reach levels that adversely affect connectivity or habitat quality.
The timing of fish movement into the RPA suggests that the key periods when fish may be at risk from mine-derived solutes are during the wet season, when fish are actively using the sand channels and backflow billabongs as habitat, and the recessional flow period at the end of the wet season, when fish are actively migrating through the sand channels to reach dry season refuges. In the wet season, we found that considerable numbers of large-bodied fish from the escarpment billabongs move downstream to take up residence in the sand channels during periods of elevated flow. Such flows have a significant dilution effect on mine-related contaminants in the creek, which lowers the potential risks of wastewater contamination during higher flow periods. Our observations of tagged fish moving repeatedly past the mine discharge site, and a lack of avoidance behaviour at the mine discharge site, provide no evidence that fish migration in Magela Creek was adversely affected by mine contaminant egress during the study. Nonetheless, the extensive use of channels and wetlands within the RPA by fish emphasizes the importance of managing legacy mine sites to ensure that contaminant egress and impacts on water quality do not adversely affect biotic connectivity and the ecological processes it supports.
Data availability statement
Data will be made available upon reasonable request.
Change history
27 February 2023
Missing Open Access funding information has been added in the Funding Note.
References
Affandi FA, Ishak MY (2019) Impacts of suspended sediment and metal pollution from mining activities on riverine fish population - a review. Environ Sci Pollut Res 26:16939–16951
ARRRI (1985) Alligator rivers region research institute annual research summary. Supervising scientist for the alligator rivers region. Australian Government Publishing Service, Canberra. 155 pp
ARRRI (1991) Alligator rivers region research institute annual research summary, 1988–89. Supervising scientist for the alligator rivers region. Australian Government Publishing Service, Canberra. 156 pp
Bishop KA, Forbes MA (1991) The freshwater fishes of northern Australia. In: Haynes CD, Ridpath MG, Williams MAJ (eds) Monsoonal Australia, Landscape, Ecology and Man in the Northern Lowlands. Balkema, Rotterdam, pp 79–107
Bishop KA, Pidgeon RWJ, Walden DJ (1995) Studies on fish movement dynamics in a tropical floodplain river: prerequisites for a procedure to monitor the impacts of mining. Aust J Ecol 20:81–107
Byrne P, Wood PJ, Reid I (2012) The impairment of river systems by metal mine contamination: a review including remediation options. Crit Rev Environ Sci Technol 42:2017–2077
Crook DA, Lowe WH, Allendorf FW, Erős T, Finn DS, Gillanders BM, Hadwen WL, Harrod C, Hermoso V, Jennings S, Kilada RW (2015) Human effects on ecological connectivity in aquatic ecosystems: integrating scientific approaches to support management and mitigation. Sci Total Environ 534:52–64
Crook DA, Buckle DJ, Allsop Q, Baldwin W, Saunders TM, Kyne PM, Woodhead JD, Maas R, Roberts B, Douglas MM (2016) Use of otolith chemistry and acoustic telemetry to elucidate migratory contingents in barramundi Lates calcarifer. Mar Freshw Res 68:1554–1566
Crook DA, Buckle DJ, Morrongiello JR, Allsop QA, Baldwin W, Saunders TM, Douglas MM (2020) Tracking the resource pulse: movement responses of fish to dynamic floodplain habitat in a tropical river. J Anim Ecol 89:795–807
Crook DA, Keller K, Adair BJ, Luiz OJ, Waugh PS, Schult J, Dostine PL, Townsend SA, King AJ (2021a) Use of radiotelemetry to quantify diel habitat preferences and minimum environmental flow requirements of a tropical riverine fish (Black bream Hephaestus fuliginosus). Ecohydrology 14:e2290
Crooks KR, Sanjayan M (2006) Connectivity conservation. Cambridge University Press, Cambridge, UK
David BO, Closs GP (2002) Behavior of a stream-dwelling fish before, during, and after high-discharge events. Trans Am Fish Soc 131:762–771
Ebner BC, Morgan DL (2013) Using remote underwater video to estimate freshwater fish species richness. J Fish Biol 82:1592–1612
Harford A, Mooney T, Trenfield M, van Dam R (2015) Manganese toxicity to tropical freshwater species in low hardness water. Environ Toxicol Chem 34:2856–2863
Hogsden KL, Harding JS (2012) Consequences of acid mine drainage for the structure and function of benthic stream communities: a review. Freshw Sci 31:108–120
Humphrey CL, Bishop KA, Dostine PL (2016) Vulnerability of fish and macroinvertebrates to key threats in streams of the Kakadu region, northern Australia: assemblage dynamics, existing assessments and knowledge needs. Mar Freshw Res 69:1092–1109
Humphrey CL, Chandler L (2018) Use of field-effects information to inform surface water guideline values for magnesium sulfate in Magela Creek. Supervising Scientist Report 213, Supervising Scientist, Commonwealth of Australia, pp. 220
Innovasea (2020) V5 180 kHz Coded transmitters datasheet. https://www.innovasea.com/fish-tracking/products/acoustic-transmitters/. Accessed 17 Jan 2023
Jardine TD, Pusey BJ, Hamilton SK, Pettit NE, Davies PM, Douglas MM, Sinnamon V, Halliday IA, Bunn SE (2012) Fish mediate high food web connectivity in the lower reaches of a tropical floodplain river. Oecologia 168:829–838
King AJ, George A, Buckle DJ, Novak PA, Fulton CJ (2018) Efficacy of remote underwater video cameras for monitoring tropical wetland fishes. Hydrobiologia 807(1):145–164
Koehn JD, Nicol SJ, McKenzie JA, Lieschke JA, Lyon JP, Pomorin K (2008) Spatial ecology of an endangered native Australian Percichthyid fish, the trout cod Maccullochella macquariensis. Endanger Species Res 4:219–225
Lucas MC, Mercer T, Peirson G, Frear PA (2000) Seasonal movements of coarse fish in lowland rivers and their relevance to fisheries management. In: Cowx IG (ed) Management and Ecology of River Fisheries. Blackwell, London, pp 87–100
Marshall JC, Menke N, Crook DA, Lobegeiger JS, Balcombe SR, Huey JA, Fawcett JH, Bond NR, Starkey AH, Sternberg D, Linke S (2016) Go with the flow: the movement behaviour of fish from isolated waterhole refugia during connecting flow events in an intermittent dryland river. Freshw Biol 61:1242–1258
Mooney TJ, Pease CJ, Hogan AC, Trenfield M, Kleinhenz LS, Humphrey C, van Dam RA, Harford AJ (2019) Freshwater chronic ammonia toxicity: a tropical-to-temperate comparison. Environ Toxicol Chem 38:177–189
Mooney TJ, Harford AJ, Hanley J, Walker S, Sandgren S, Jansen A, Humphrey C (2023) Seasonal responses of macroinvertebrate assemblages to magnesium in a seasonally flowing stream. Env Pollut 316:120586
Pease CJ, Trenfield MA, Mooney TJ, van Dam RA, Walker S, Tanneberger C, Harford AJ (2021) Development of a sub-lethal chronic toxicity test for the northern trout gudgeon, Mogurnda mogurnda, and application to uranium, magnesium, and manganese. Environ Toxicol Chem 40:1596–1605
Pettit NE, Naiman RJ, Warfe DM, Jardine TD, Douglas MM, Bunn SE, Davies PM (2017) Productivity and connectivity in tropical riverscapes of northern Australia: ecological insights for management. Ecosystems 20:492–514
RMCP (2020) Ranger mine closure plan. Energy Resources of Australia. Closure Plan - Energy Resources of Australia. https://www.energyres.com.au/ranger-rehabilitation/mine-closure-plan/. Accessed 3 Feb 2022
Saboret G, Buckle DJ, King AJ, Douglas MM, Crook DA (2021) Partial migration in diadromous fishes drives the allocation of subsidies across the freshwater-marine ecotone. Anim Migr 8:40–55
Schindler DE, Armstrong JB, Reed TE (2015) The portfolio concept in ecology and evolution. Front Ecol Environ 13:257–263
Supervising Scientist (2020) Annual technical report. Supervising Scientist Branch, Commonwealth of Australia, pp 226
Supervising Scientist (2021) Magnesium in surface water — rehabilitation standard for the Ranger uranium mine (version 2). Supervising Scientist Branch, Commonwealth of Australia, pp 13
van Dam RA, Hogan AC, McCullough CD, Houston MA, Humphrey CL, Harford AJ (2010) Aquatic toxicity of magnesium sulfate, and the influence of calcium, in very low ionic concentration water. Environ Toxicol Chem 29:410–421
Wright IA, Belmer N, Davies PJ (2017) Coal mine water pollution and ecological impairment of one of Australia’s most ‘protected’ high conservation-value rivers. Water Air Soil Pollut 228:90
Acknowledgements
We pay our respects to all Traditional Owners of Kakadu National Park and the Darwin region where we conduct research and monitoring, and acknowledge their Elders past, present, and emerging. The authors are grateful to the Djurrubu Rangers and Gundjeihmi Aboriginal Corporation for their assistance with project logistics and fieldwork and acknowledge Energy Resources of Australia Ltd. for their co-operation and permissions for site access, and provision of water quality data. We thank Northern Australia Helicopters for transporting the team and equipment safely to Bowerbird Billabong. We gratefully acknowledge the extensive foundational science conducted by Dr. Keith Bishop and other staff of the Supervising Scientist Branch (SSB) over many years which formed the basis for the ideas underpinning our study. This study was funded through the National Environmental Science Program Northern Australia Environmental Resources Hub. Thanks also to the staff at the NESP Northern Australia Environmental Resources Hub, particularly Jane Thomas, Clare Taylor, and Patch Clapp, for their support over the life of the project. The Supervising Scientist Branch (Australian Department of Climate Change, Energy, the Environment and Water) also provided in-kind support for this project. We thank Andrew Jansen (SSB) for preparing Fig. 1 and the conceptual figures of seasonal fish migration and residency (Supplementary Material 1). We gratefully acknowledge the constructive input from two anonymous reviewers.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics approval
This research was conducted under Charles Darwin University Animal Ethics Committee approval A18030, Northern Land Council permits 79102 and 82708, and NT Fisheries Act Special permit S17/3454.
Conflict of interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Crook, D.A., Wedd, D., Adair, B.J. et al. Evaluation and refinement of a fish movement model for a tropical Australian stream subject to mine contaminant egress. Environ Biol Fish 106, 469–490 (2023). https://doi.org/10.1007/s10641-022-01381-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-022-01381-y