
Abstract
We examined variability in otolith chemistry of wild caught fish in relation to in situ temperature and salinity within the California Current System. Barium, magnesium, and iron from the most recent growth zone in otoliths differentiated pelagic juvenile shortbelly rockfish (Sebastes jordani) residing in water masses with distinct temperature and salinity properties from central and southern California spanning nearly 500 km of coastline. The 3-element signature also discriminated fish that resided in different water masses that were associated with mesoscale cyclonic eddy circulation in the Santa Barbara Channel. Variability in otolith chemistry reflected the spatial patterns of both horizontal gradients and vertical gradients in water mass properties related to circulation. Although we found that the concentrations of particular elements in otoliths were correlated to ambient temperature or salinity, we suggest that these parameters are more useful as an identifying signature of distinct water masses associated with unique otolith signatures rather than as factors directly affecting otolith chemistry. Other factors varying among the water masses or among the fish populations residing in the water masses may also affect otolith chemistry. We discuss how oceanographic phenomena associated with the 1997–1998 El Niño and the persistent, recirculating eddy in the Channel may have affected coastal ocean conditions and variation in otolith chemistry of fish in the study area.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Elemental and isotopic assays of fish otoliths have been directed at questions concerning the tracking and mixing of marine and freshwater populations. Inferences about fish homing behavior (Secor et al. 1995; Thorrold et al. 2001), connectivity estimates (Miller and Shanks 2004; Standish et al. 2008), and larval dispersal histories (Swearer et al. 1999; FitzGerald et al. 2004; Hamilton et al. 2008) have been based on otolith chemistry as geographic tags. These applications of otolith chemistry do not necessarily require the determination of how element concentrations, salinity, and temperature in the environment are related to element concentrations in the otolith; what is required is that the chemical assays of fish otoliths discriminate individuals from different locations. However, at least two important limitations in utilizing otolith chemistry to assess the movement of individuals and the mixing of populations among locations stem from not knowing how environmental variability relates to the spatial and temporal variability in otolith chemistry.
The first limitation is the reliability of site-specific otolith elemental signatures over time. Sufficient consistency in the spatial variability of otolith elemental signatures allows discrimination among habitat sites within a year; however, many studies that have compared otolith chemical composition among locations over two or more years have found significant interannual variability (Milton et al. 1997; Dove and Kingsford 1998; Patterson et al. 1999; Campana et al. 2000; Gillanders 2002; Hamer et al. 2003; Miller and Shanks 2004; Brown 2006). This temporal inconsistency is especially problematic if the goal, for example, is to examine a population over time to estimate connectivity via migration or larval dispersal among spatially distinct habitats.
The second limitation is that individuals of unknown residency can only be assigned to the set of sampled locations that define the spatial variability of the signatures. There is an unknown degree of error in assigning fish of unknown residency to a specific site of a set of sampled locations, because fish residing in unsampled locations might share the same elemental signature as fish from the set of sampled locations. This can occur even if the spatial variability of the otolith elemental signatures defined by a set of sampled locations is consistent over time and the elemental signatures are good discriminators of the sites sampled.
Presently, inferences are limited to the geographic scale of sampling, the localities sampled, and the time of sampling within individual studies, because of the uncertainty of changing environmental conditions that affect the variability in otolith chemistry over time and space. We turn our attention from the utility of using otolith chemistry as a location-based discriminator, and focus on the prospect of utilizing otolith chemistry to identify water mass residency. A motivation for this study is that if unique elemental signatures are associated with distinct water masses, then a reference or atlas identifying these relationships might be developed. Otolith chemistry and oceanography might prove useful for reconstructing the transport histories of individuals and estimating the connectivity of populations across broad regions and over time.
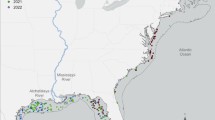
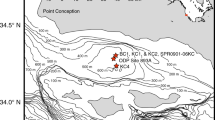
The central goal of this study was to test the hypothesis that variability among otolith trace element signatures of wild-caught fish was associated with in situ water masses. To identify distinct water masses, we examined temperature and salinity measured during pelagic juvenile fish sampling off central California in May 1998 (Sakuma et al. 2000) and off the southern California coast in June 1998 (Nishimoto and Washburn 2002) spanning a distance of nearly 500 km (Fig. 1). The otolith signatures of shortbelly rockfish (Sebastes jordani) from these surveys were defined by a suite of elements assayed from the outer otolith material grown in the recent days preceding capture. Otolith sampling included fish found concentrated in a persistent cyclonic eddy in the western Santa Barbara Channel (Nishimoto and Washburn 2002; Figs. 1b and 2). We examined the variability in otolith chemistry in relation to temperature and salinity across the study area and the three-dimensional circulation of the eddy.

The study area. Midwater trawling and oceanographic surveys in late spring 1998 were conducted off (a) central California and (b) southern California. Shortbelly rockfish specimens and temperature-salinity profiles were collected in central California from stations numbered 1–6, and in southern California from stations 7–24. The eddy (shaded gray area) in the western Santa Barbara Channel was defined by station dynamic height ϕ < 0.37 (5 dbar/200 dbar) (Nishimoto and Washburn 2002). The closed circles mark stations where fish were absent or no otoliths were sampled

East-west vertical sections of: (a) potential temperature, T (°C); and (b) salinity, S (psu), along the section indicated by the line in Fig. 1. Vertical dotted lines in (a) indicate T-S profile locations. The cyclonic flow of the eddy produced the large uplift of isotherms and isohalines. Two horizontal dashed lines bound the 14-m trawling stratum. The average headrope depth for all hauls in the Santa Barbara Channel survey was 20 m (from Nishimoto and Washburn (2002))
Materials and methods
Water mass sampling and otolith chemistry
Similar methods were used in the central and southern California surveys (detailed in Wyllie Echeverria et al. 1990; Nishimoto and Washburn 2002; Sakuma et al. 2000) to collect the otoliths and oceanographic data for this study. Fish were collected at night with a modified Cobb mid-water trawl with a 9 mm codend towed at depth for 15 min at ~5 km h−1 covering ~1.5 km (Nishimoto and Washburn 2002). The opening of the net used in both surveys was approximately 10 m wide and 14 m high when trawling at a headrope depth of 20 m. The depth interval for each haul was estimated as the distance from the mean depth, d0, of the headrope (rope across the top of the net opening) to 14 m below d0 (Table 1). Vertical profiles of potential temperature T and salinity S (averaged into 1-m depth bins) were obtained either immediately before or after each haul to at least 200 m or a few meters above shallower bottom depths. A water mass where fish were sampled was defined by the profile within the depth interval fished.
Otoliths from 68 specimens of shortbelly rockfish sampled from the central and southern California collections were assayed using laser ablation inductively coupled plasma mass spectrometry (LA-ICPMS) (Fig. 1, Table 1). The fish ranged from 12.8 mm to 47.2 mm SL. Shortbelly rockfish were the most abundant of the pelagic juvenile rockfishes collected in the survey. Adults of this active, schooling species range from southern British Columbia to southern Baja California, and larvae are found as much as 400 km from shore (Love et al. 2002). Fifty-seven specimens represent all areas defined by Nishimoto and Washburn (2002) where shortbelly rockfish were collected during 3–15 June 1998 in southern California (Fig. 1b, Table 1). Eleven otolith samples from central California were collected during 11–27 May 1998 from the Farallon Islands west of San Francisco to Cypress Point, Monterey (stations 1–6 in Fig. 1a and Table 1). The otoliths of one to four specimens per station were assayed (Table 1).
The otolith samples from the two surveys were handled somewhat differently. Specimens from the 1998 southern California survey were subsampled in 2000 from an ETOH-preserved archive, measured, and the sagittae extracted. The fish had been frozen at sea, then thawed and archived within several months after collection. The otoliths were rinsed in deionized water, air dried and stored in plastic bags for several months until the time when each otolith was affixed to a plastic slide using epoxy resin (Epo-Thin, Buehler). The fish collected in the central California survey were frozen at sea. Within several months, the specimens were thawed and measured, and the otoliths extracted, rinsed, dried and affixed to glass slides using a clear nail polish epoxy (Sally Hansen’s Hard-as-Nails brand).
All otoliths were polished down to about 15 μm above the center of otolith nucleus using a lapping wheel and 9, 3, and 1 μm 3 M diamond polishing films. After polishing, decontamination steps described by Ruttenberg et al. (2005) and Warner et al. (2005) were performed in a clean laboratory on individually isolated otoliths.
Otolith material produced within 1 week preceding capture was analyzed on a Finnigan MAT Element 2-sector field ICPMS as described in Warner et al. (2005) and Ruttenberg et al. (2005). A VG UV microprobe Nd:YAG (Neodymium-doped Yttrium Aluminum Garnet) 266 nm laser ablation system was outfitted with a helium aerosol carrier system to transfer the sample from the enclosed sample chamber to the ICPMS as described in Zacherl et al. (2003). The laser was set at 0.1 mJ at 3 Hz. The laser was used to ablate three sample spots in the same growth zone near the edge of the anterior rostrum of each otolith with care taken to avoid epoxy that overlaid one or two outermost increments. For the southern California otoliths, run in May 2002, the laser aperture was set to 1 μm which emitted a beam that ablated a pit diameter of about 20 μm. For the central California otoliths, run in September 2003, the laser aperture was set to 2 μm which ablated a pit diameter of about 30 μm.
Two samples were collected at each of the three spots to assay a suite of isotopes: Calcium 48 (48Ca), Strontium 86 (86Sr) Barium 138 (138Ba) and Lead 208 (208Pb) were determined by using low resolution mode (R = 300); Magnesium 24 (24 Mg), Calcium 48 (48Ca), Manganese 55 (55Mn), Iron 56 (56Fe), and Zinc 64 or Zn 66 (64Zn for SBC samples and 66Zn for CC samples) were determined in medium resolution mode (R = 4,000). Eight laser pulses were emitted on a targeted spot to collect each sample. Each targeted spot was pre-ablated with two laser pulses as a precautionary cleaning measure. The isotope intensities of each sample were blank-corrected by subtracting isotope intensities of a 1% nitric acid (HNO3) instrument blank preceding the sample sequence. The plasma conditions were kept similar for both ablated material and solution-based standards by aspirating a 1% nitric acid solution during the ablation process and keeping the gas flows constant. The abundance of an element was expressed as a ratio relative to the amount of calcium to control for the amount of material analyzed per sample spot. A solution standard of known element/Ca ratios was used to correct for instrument bias and convert to molar elemental ratios. Solution-based standards were used in an attempt to match the high calcium matrix of the otoliths. The currently available solid standard materials, such as NIST 612 glass, are very different in composition from the otolith (Pearce et al. 1997). Solid calcium carbonate pellets have been prepared to solve this issue, but there are problems associated with heterogeneity.
Average detection limits for each element per sequence (n = 20 sequences), calculated as 3 × SD of the intensity of 1% HNO3 blanks and expressed as ratios of the isotope intensity and mean otolith Ca48 intensity, were 24 μmol·mol−1 for Mg/Ca, 0.95 μmol·mol−1 for Mn/Ca, 12.74 μmol·mol−1 for Fe/Ca, 6.77 μmol·mol−1 for Zn/Ca, 59 μmol·mol−1 for Sr/Ca, 0.06 μmol·mol−1 for Ba/Ca, 0.06 μmol·mol−1 for Pb/Ca.
We analyzed solid glass standard reference material (NIST 612) at the beginning and end of each workday to check overall analytical precision or repeatability for the entire study. Average percent relative standard deviations (n = 10 workdays) were 21% for Mg/Ca, 13% for Mn/Ca, 26% for Fe/Ca, 37% for Zn/Ca, 39% for Sr/Ca, 10% for Ba/Ca, 10% for Pb/Ca. The NIST 612 was ablated using the same method parameters as that used for the otolith samples. The constraint of using small 20 μm and 30 μm spot sizes resulted in low signal to noise for Sr/Ca estimates in the NIST 612 standard. An alternative reproducibility estimate for the study was determined for Sr/Ca by ablating material (three spots) along a randomly selected band of growth increments between the core and the edge of an otolith at the end of each sequence. The average percent relative standard deviation was 8% for Sr/Ca (n = 19 sequences).
We excluded negative blank-corrected intensity values from the sample spot dataset. We also excluded outliers from the sample spot dataset. Outliers were identified by examining frequency distribution and normal probability plots of the log-transformed element/Ca sample spot data. The outlier values were at least 1.5 times the interquartile range from the median of the log-transformed data (SPSS, Inc 2002). The exclusions reduced the original dataset of three samples of seven isotopes per otolith to 0–3 samples of each isotope per otolith. The abundance of each element from the otolith edge was estimated from either a single sample spot or the mean of two or three sample spots after the molar ratio dataset was log-transformed (i.e., log(μmol element/mol Ca)). A specimen lacked an abundance estimate for a given element if the readings from all three sample spots at the otolith edge were excluded.
Relating otolith chemistry to in situ water masses
We examined the T-S distribution within the trawling depth ranges to resolve water mass groups. Principal component analysis (PCA) of T (15 1-m bin averaged variables, Td = 0, 1, 2, …, 14) and S (15 variables, Sd = 0, 1, 2, ..., 14) profile data over the depth interval of each haul was used to confirm the water mass groups. T and S values from each profile were normalized by subtracting the mean of all profiles and dividing by the standard deviation of all profiles.
Otoliths were assigned to the water mass groups by pairing hauls with vertical T-S profiles. All analyses were performed on log-transformed otolith element ratio data. Prior to the analyses, we assessed the normality and homoscedacity of the otolith data among water mass groups (SPSS, Inc 2002). We evaluated whether fish size was a confounding factor influencing the effect of water mass type on otolith element concentration. Rather than use log(element/Ca) in our analyses when the relationship between an element and fish size was significant (Pearson correlation coefficient r, α = 0.01), we used the residual values from the least squares regression of log(element/Ca) as a function of log(SL) to remove the fish size effect on the concentration of an element in otoliths (Systat Software Inc 2006).
We performed one-way analysis of variance (ANOVA) to test for the effect of the water mass group on individual log-transformed element ratios in otoliths (α = 0.01) (SPSS, Inc 2002). Although we would have preferred to use a nested ANOVA to account for within-group variability in individual elemental abundances due to station (i.e., water mass defined by the CTD cast data at an individual station) differences, the dataset was unsuitable because 10 of the 24 stations were represented by only one otolith, including all but one of the stations from central California. The Tukey’s honestly significant difference (HSD) test was used to detect a posteriori differences among means (α = 0.01).
We used PCA to examine whether variability among the 7-element signatures of otoliths showed natural groupings by water mass. Otoliths with excluded data from all three sample spots for at least one of the seven elements were not included in the PCA.
To test whether the elemental signature of otoliths identified water mass membership, we used canonical discriminant function analysis (DFA) (SPSS Inc 2002). The DFA included only the elements that showed significant differences among water masses as determined in the ANOVA. In comparison to the PCA, the DFA was run on an expanded subset of otoliths that had readings of the elements which showed a water mass grouping effect regardless of whether abundances of the other elements could be estimated.
The classification accuracy of each DFA was evaluated by leave-one-out cross-validation, also called jack-knife reclassification (SPSS Inc 2002; White and Ruttenberg 2007). The prior probabilities of group membership were assumed to be uniform. We used the randomization test of White and Ruttenberg (2007) to estimate the probability (p-value) that the observed jack-knife reclassification success rate was drawn from a null distribution of jack-knife values (n = 5,000 randomizations) given no difference among groups of samples.
We examined relationships between element abundances in the otolith and relationships between otolith elemental properties and T and S. We used the Pearson correlation coefficient, r, to determine whether otolith element abundances that were identified as the best discriminators of the water mass groupings co-varied (SPSS, Inc 2002). We estimated the correlation between the average otolith element abundance at a station and the average T or S. If the correlation was significant, we used least squares regression to model the relationship between T or S, the environmental predictor, and otolith log[element/Ca], the dependent variable, by fitting a linear equation to the observed data (Systat Software Inc 2006).
Results
Water masses
Based on the T-S observations, the assayed fish resided in a range of water masses across the study area (Fig. 3). We resolved four water mass groups: central California (CC), southern California (SC), uplifted deep water in the eddy (UE), and stratified water in the eddy (SE) (Fig. 3 and Table 1). The CC samples (blue T-S curves) were cooler (T < 11.7°C) than the SC water masses which were sampled outside of the eddy (green T-S curves). Both CC and SC water mass groups were less saline (S < 33.70 psu) than the three water mass samples (station 7, black curve; stations 9 and 14, red curves) within the eddy in the Santa Barbara Channel defined by dynamic height ϕ < 0.37 (Fig. 1b; Nishimoto and Washburn 2002) and one station at the southwestern boundary of the eddy (station 8, black curve).

Relationship of T-S profile data from the 14-m trawling stratum at stations where otoliths were sampled. The water mass samples were assigned to four groups: central California, CC, blue; uplifted deep water in the eddy, UE, black; stratified water mass in the eddy, SE, red; southern California, SC, green. The water mass groups are separated by grey horizontal and vertical reference lines at 11.7°C and 33.7 psu. Each T-S plot is labeled with a station number (see Fig. 1). Markers, P and S, in the plot are reference T-S of San Francisco Bay Plume water and subarctic water, respectively
The two water mass samples in the UE group (Fig. 3, black T-S curves, stations 7 and 8) have T and S properties within the trawling range of 19 m to 33 m that are consistent with deep water uplifted from 100 m within the eddy described by Nishimoto and Washburn 2002 (Fig. 2, T < 11.7°C and S > 33.7 psu). The water mass samples in the SE group (Fig. 3, red T-S curves, stations 9 and 14), share T-S properties of uplifted deep water (UE group) and the most saline SC samples found outside of the eddy within the Santa Barbara Channel (Figs. 1b and 3, stations 10, 15, 19, 20).
The T-S properties of the central California samples are bounded by the characteristic T-S of two distinct end-members: San Francisco Bay Plume water (12.6, 32.85 psu, defined by Hurst and Bruland 2008) and subarctic water (mean temperature 10.06°C, mean salinity 33.36 psu, defined by Warn-Varnas et al. 2007) transported by the equatorward California Current (Fig. 3). The intermediate T-S properties of our central California samples from waters 50 m and shallower (stations 2, 3, 5 and 6; d0 ≤ 36 m) indicate advection and mixing of the two relatively fresh sources along the Central California coast. The two deeper CC samples (stations 1 and 4; d0 = 75 and 77 m, respectively) and UE group are different “deep” water masses; although similar in temperature, the two CC samples are less saline than the UE group.
PCA of the T-S profile data was consistent with the distribution of water masses (i.e., T-S curves in Fig 3). The first two principal components, PC1 and PC2, explained 97% of the variance among the water mass samples (data not shown). PC1 separated CC samples from the southern California samples (SC, UE, and SE). PC2 separated the eddy samples, UE and SE, from SC. PC 3, which explained 2% of the variance, separated SE from UE, SC, and CC.
Relationships between otolith signatures and water masses
Otolith concentrations of Mg, Ba, and Fe significantly varied among the four water masses (Fig. 4, Table 2). Mg/Ca was highest in otoliths from UE and significantly differed among the other three water masses (Tukey HSD test, P < 0.001). Ba concentration in the otoliths from the CC water mass was significantly higher than in otoliths from each of the water masses from the Santa Barbara Channel region (Tukey HSD test, P < 0.001). Fe concentration in otoliths from central California was lower than those from the UE (Tukey HSD test, P = 0.009), SE (P = 0.024, marginally significant), and SC water masses (P < 0.001). The estimated Fe concentrations of 30 otoliths from 18 stations, including all otoliths from central California, were below our calculated detection limit. Neither Ba nor Fe differed between water masses in the Santa Barbara Channel. The effect of water mass on Pb was marginally significant (ANOVA, P = 0.024); Pb in otoliths was higher in CC than in SE (Tukey HSD test, p = 0.013). Pb concentrations of 28 otoliths from 18 stations were below our calculated detection limit. The effect of water mass on the concentration of Mn, Zn, or Sr in otoliths was not significant.

Average log concentration of Mg, Mn, Fe, Zn, Sr, Ba, and Pb (log[μmol element/mol Ca] ± 95% CI) in otoliths (N) from the four water masses: CC, UE, SE, and SC. The detection limit for an element/Ca is demarcated as a horizontal line if within the range of otolith element concentrations
Fish were significantly larger in UE than in CC (Tukey HSD test, P = 0.047) and SC (P = 0.001) (ANOVA, n = 60, F = 4.665, d. f. = 3, P = 0.006). Sr was the only element in fish otoliths that was related to fish size (Fig. 5; log[μmol Sr/mol Ca] = 3.63–0.25 log(mm SL), n = 66, r 2 = −0.18, p = 0.0003). We removed the confounding effect of fish size on otolith Sr by using the residual values from the regression of log(Sr/Ca) on log(SL) in the ANOVA and in the analyses reported herein.

Otolith Sr v body length. Log(mm SL) was significantly but weakly correlated with Log [Sr/Ca]. Marker color indicates the water mass groups: CC, blue; UE, black; SE, red; SC, green
Natural groupings of otolith signatures reflected the distinctiveness of the water masses where the fish resided. The first two principal components of PCA run on a subset of 49 otoliths with all seven elements explained 64% of the variance in elemental signatures among otoliths (Fig. 6). Inspection of the plot suggested that fish residing in central California had otolith signatures that differed from that of fish in southern California, and that fish residing in the UE water mass were distinct from fish from the SE and SC water masses. Separation of otolith signatures from CC and UE along PC1 are consistent with the lack of overlap in T and S properties of the water masses (Fig. 3). The lack of separation of otoliths from SE and SC (Fig. 4) is consistent with the intersection of the T-S properties of the two water masses for T > 11.6o C (Fig. 3). The separation of otoliths elemental signatures into water mass groupings in the PCA corresponds with the ANOVA findings of a water mass effect on the elemental concentrations of Mg, Ba, and Fe.

Principal components analysis for a subset of 49 otoliths using Mg, Mn, Fe, Zn, Sr, Ba, and Pb. Colored symbols indicate the water mass groups from where otoliths were sampled as in Fig. 3
We found that the otolith signature of three elements, Mg, Ba, and Fe was a good predictor of water mass residency. DFA was run on a subset of 60 otoliths that had readings of Mg, Ba, Fe, three elements that differed among water masses, regardless of whether the other four elements could be estimated (Fig. 7). Cross-validation showed that 70% of the otoliths correctly grouped into one of the four assigned water mass types (Table 3), significantly better than 25.3% (SD = 8.2% upper 95% quartile of the null distribution = 38.3% expected by chance (p = 0.0002)). The inclusion of Pb in the DFA (55 otoliths with Mg, Ba, Fe, and Pb) had an insignificant effect on the classification accuracy (plot of the DFA results not shown); cross-validation showed that 70.9% of the 4-element signatures correctly grouped into the assigned water masses.

Canonical Discriminant Analysis (CDA) of 60 otoliths using Mg, Ba, and Fe. Colored symbols indicate the water mass groups from where otoliths were sampled as in Fig. 6
The 3-element (Mg, Ba, and Fe) discriminant function correctly grouped 100% of the otoliths collected in CC, and 100% of the otoliths collected in UE corresponding with the distinct T-S properties of the two water masses (Table 3). The signature poorly discriminated the residency of fish collected in SC and SE; however, none of the otoliths from these two groups were misclassified as CC otoliths. Fifty-eight percent of the otoliths from SE were incorrectly reassigned to UE (3 of 12 otoliths) and SC (4 of 12 otoliths). The lack of a distinctive SE otolith signature and the similarity of otolith signatures of fish collected from SE to otolith signatures from UE and SC (Figs. 4, 6 and 7) are consistent with the mixed water mass properties of SE (Fig. 3).
The strength in discriminating otoliths from the different water masses was associated with trends of decreasing otolith Ba and increasing otolith Mg with increasing S. Ba/Ca in the otoliths was negatively correlated with S and the regression explained 62% of the variance in Ba/Ca (Fig. 6d). Mg concentration was correlated with S although the regression explained only 25% of the variance in Mg/Ca (Fig. 8).

Least squares regression, \( {\hbox{y}} = \left( {{\hbox{a}} \pm 95\% \,{\hbox{CI}}} \right){\hbox{x}} + \left( {{\hbox{b}}\pm 95\% \,{\hbox{CI}}} \right) \), fit to the mean log otolith concentration of Ba, Mg, and Fe v mean T (black Xs in plots on left panel) and mean S (black Xs in plots on right panel) at each station (n = 23). Dashed lines are 95% confidence bands. Log[element/Ca] per otolith are plotted (colored symbols indicate the water masses from where otoliths were sampled as in Fig. 6). The detection limit for Fe/Ca, the dotted horizontal line in the plots, was within the range of otolith Fe concentrations
Ba and Mg concentrations in otoliths from UE and SE differed from those in otoliths from CC despite similar T between 10 and 12 C (Fig. 8). T was a weak predictor of Fe explaining only 19% of the variance (Fig. 8).
Discussion
Otolith element/Ca ratios varied among the sampled water masses in the California Current System. Examining the variability in otolith chemistry in relation to temperature and salinity allowed us to not only distinguish fish from geographically distant regions with contrasting oceanographic conditions but also to discriminate fish that resided in different water mass environments that were associated with a local, physical phenomenon —cyclonic eddy circulation in the Santa Barbara Channel. We found that Ba, Mg, and Fe varied significantly among the distinguishable water masses that we sampled. Fish from central California had elevated otolith Ba/Ca compared with fish from southern California. In contrast, otolith Fe/Ca was elevated in fish from southern California compared to fish from central California. Otolith Mg/Ca was highest in fish collected from deep water uplifted to shallow depths by cyclonic eddy circulation in the Santa Barbara Channel and lowest in fish from central California. The 3-element signature correctly predicted the residency of 100% of the fish collected in the cold core of the eddy in the Santa Barbara Channel and separated 100% of the fish collected in the Channel region from fish collected from Central California.
Two of the three elements, Ba/Ca and Mg/Ca, have been useful for distinguishing fish from different coastal and oceanic locations of other studies. Fe/Ca is typically excluded from otolith microchemistry analyses, because uncertainty in element abundance estimates is assumed to be high for signals below detection limits (Campana 1999). However, Ben-Tzvi et al. (2007) found that LA-ICPMS element/Ca measurements that would be discarded as noise using conventional DL standards (e.g., 3 × SD) often formed sequences that were symmetrical about the otolith core (palindrome). Considering their argument that these sub-DL signals can contain valuable information, we included sub-DL signals of Fe/Ca and other elements in our analyses.
We found that otolith Ba/Ca was negatively correlated with salinity of the water mass occupied by the wild-caught fish (Fig. 8). We did not measure elemental concentrations in water mass samples; however, we surmise that ambient Ba/Ca was negatively associated with salinity and that otolith Ba/Ca reflected the availability of Ba/Ca. Laboratory and field studies have determined that otolith Ba/Ca correlates with ambient Ba/Ca (Bath et al. 2000; Elsdon and Gillanders 2003, 2005b). Compared to the influence of ambient Ba/Ca, the effect of ambient temperature and salinity on otolith Ba/Ca has been shown to be minimal (Bath et al. 2000; Elsdon and Gillanders 2004, 2005b). Elsdon and Gillanders (2005a) attributed the inverse relationship between otolith Ba/Ca of black bream and ambient salinity in estuarine and coastal environments of southern Australia to the input of Ba from fluvial sediments in freshwater runoff. In the Southern California Bight, ambient Ba/Ca in estuarine waters (about 6 μmol·mol−1) were higher than coastal waters (about 3 to 5 μmol·mol−1) among several studies that were not conducted during our survey year (Forrester 2005; Warner et al. 2005; Lloyd et al. 2008). In a three-year study off central California, Brown (2006) found that whole otolith Ba/Ca of two flatfish species along central California was highest in 1998 (the year of our collections) and lowest in 1999, in correspondence with runoff levels.
River runoff is the major source of Ba to coastal waters (Chan et al. 1977) and may have contributed substantially to the Ba load off Central California during our study. Increased precipitation in California and coastal runoff are frequently associated with strong El Niño events, and the winter of 1997–1998, when an exceptionally strong El Niño was fully developed, was one of the wettest seasons on record. The impact of sediment and freshwater transport in nearshore waters from coastal watersheds was spatially extensive in satellite imagery during the winter and spring of 1998 (Mertes and Warrick 2001). Oceanographic surveying with the collection of the central California specimens indicate that outflow from the San Francisco Bay, draining the largest watershed of California, generated a markedly low salinity signal that extended 100 km down the central California coast to Monterey Bay (Sakuma et al. 2000). High turbidity and elevated nutrient levels were correlated with low salinity in Monterey Bay providing evidence that low salinity in the upper 15–20 m was the result of local and remote runoff and river outflow in 1998 (Friederich et al. 2002). The concentrations of some trace elements, Ba was not among the elements measured, have been found to be higher in the San Francisco Bay plume than in coastal and upwelled waters (Hurst and Bruland 2008).
Elevated Ba/Ca in otoliths and invertebrate hard parts has been associated with upwelling (Lea et al. 1989; Patterson et al. 1999; Zacherl 2005; Brown 2006) and deep water (Zumholz et al. 2007). Dissolved Ba in oceanic waters shows a nutrient-like distribution with very low concentrations in surface waters and higher values in deep water (Chan et al. 1977). In the Santa Barbara Channel, upwelling within the cyclonic eddy is a mechanism that can draw up nutrient-rich waters from below the thermocline into shallower depths where the juvenile fish were sampled; however, otolith Ba/Ca within the cyclonic eddy was not significantly higher than that outside of the eddy (Fig. 7). Bograd and Lynn (2001) regarded the depth of the 12°C isotherm a reliable proxy for the nutricline depth in the Southern California Bight and Santa Barbara Channel. Figure 3 suggests that the nutricline was uplifted within the core of the eddy; however, it is plausible that variation in ambient Ba did not significantly affect otolith Ba among the three water masses in southern California. Warner et al. (2005) found no correspondence between the concentrations of Ba in seawater and Ba in otoliths of adult or embryonic larval kelp rockfish (Sebastes atrovirens) from nearshore reefs within our southern California study region.
An alternate, plausible mechanism that would limit otolith Ba/Ca in the fish from the core of the eddy is localized Ba depletion from rapid plankton production in the euphotic zone where the fish were residing. Lochte and Pfannkuche (1987) observed nutrient depletion in the upper 50 m within the center of a cyclonic eddy. They proposed that the physical isolation of eddy surface waters due to the formation of a shallow thermocline lead to rapid utilization of nutrients. Off the Baja California coast, the waning stages of an upwelling-induced phytoplankton bloom were associated with Ba depletion in surface waters (Esser and Volpe 2002). The relationship between Ba removal from surface seawater, uptake by different organisms, and biological productivity are poorly understood and beyond the scope of this discussion.
In contrast to otolith Ba/Ca, Mg/Ca was relatively low in otoliths of fish from central California, where the waters were freshest in our study area, and highest in otoliths of fish residing in the core of the eddy, where waters were the most saline (Figs. 4 and 8). Unlike Ba which has a nutrient-like vertical distribution in the ocean, Mg, like Ca, has generally a conservative distribution and is present at relatively high concentration levels in constant proportion to salinity (Nozaki 1997). Ratios of conservative elements, Mg/Ca for example, are not expected to vary significantly in seawater (Lea et al. 1999). Rivers contribute Mg to coastal areas (Tipper et al. 2006); however, it is likely that the input of Mg from freshwater runoff would be swamped out by the concentration of Mg in seawater, which is about five times higher than Ca in seawater. Substantial differences in ambient Mg/Ca among the water masses we sampled seem unlikely, so variation in otolith Mg/Ca probably is due to other factors.
A large body of studies shows that Mg/Ca is useful for discriminating fish from different open coastal and oceanic locations exposed to water masses. For example, in nearshore coastal waters along Washington and Oregon, Miller and Shanks (2004) found that Mg/Ca was the most important discriminator among a suite of elements for assigning juvenile rockfish to their collection locations. Mg is one of the most important elements in otoliths separating fish from deepwater areas. Swan et al. (2006) found that Mg along with Pb were important for differentiating deep water fish stocks of the European hake, Merluccius merluccius, in the northeastern Atlantic. The otolith chemistry results were consistent with the treatment of a northern stock west of Norway and a southern stock off Portugal that were separated by an oceanographic and bathymetric feature, the Bay of Biscay. Ashford et al. (2005) used Mg along with Ba, Mn, and Sr in otoliths of deep-water Patagonian toothfish to successfully distinguish fish collected off South America from fish collected south of the subantarctic front in the Atlantic, Indian, and Pacific regions of the Southern Ocean.
The effects of ambient Mg/Ca on otolith Mg/Ca and the effects of salinity and temperature on Mg uptake are unclear. We found that otolith Mg/Ca was weakly correlated with S and not related to T. Most of the experimental studies have found no effect of salinity on otolith Mg/Ca (Fowler et al. 1995b; Hoff and Fuiman 1995; Martin and Thorrold 2005). Fowler et al. (1995a) found that Mg and Ca were more abundant in the whole otoliths of fish in low-temperature treatments. In contrast, in experimental manipulations of salinity and temperature, Martin and Thorrold (2005) observed that the uptake rate of Mg in otoliths increased with increasing temperature (although the relationship was not statistically significant) while ambient Mg/Ca did not vary among the treatments. With exception of Martin and Thorrold (2005), the limitation of most of the experimental studies was that dilution of a single natural water mass or artificial seawater was used to manipulate salinity, and neither ambient Mg/Ca nor the effect of dilution on ambient Mg/Ca were measured in the majority of studies. The effect of ambient Mg/Ca on otolith Mg/Ca and possible interactions with ambient temperature and salinity is yet to be rigorously tested.
It is arguable whether ambient Mg/Ca actually has an effect on otolith Mg/Ca, because Mg is under strict osmoregulatory control (Campana 1999). Elsdon and Gillanders (2002) found that Mg/Ca showed little or no effects due to temperature or salinity and varied considerably among individuals within temperature and salinity treatments. Endogenous controls would vary among individuals due to genetics, ontogeny, condition, metabolism, and somatic growth. Mg/Ca was highest in otoliths from UE where fish were larger than fish from CC or SC; however, Mg/Ca tended to be relatively low in otoliths from CC where fish were not the smallest among water masses. Ontogenetic differences may account for some of the variance in Mg/Ca observed among water masses. Fowler et al. (1995a, b) found that Mg in whole otoliths varied with ontogeny. Martin and Thorrold (2005) showed that uptake rate of Mg in otoliths and otolith Mg/Ca was related to otolith precipitation and somatic growth, which is often highly correlated with temperature in the field. If endogenous control of otolith Mg/Ca is not measurably mediated by ambient Mg, temperature, and/or salinity from the environment, then otolith Mg/Ca may not be a reliable indicator of water mass residency, but rather a useful discriminator for the separation of populations that have unique physiological traits meditated by genetics, growth, and condition.
It is tempting to speculate that otolith Mg/Ca was a good discriminator of the fish residing in the different water masses that we sampled, because the population of young-of-year fish within each water mass possessed unique intrinsic characteristics. Oceanographic processes that determined circulation patterns along with behaviors such as swimming capability, temperature or depth preferences, and schooling may have contributed to the isolation of these pelagic juvenile fish populations.
In conclusion, we found that otolith chemistry was a good predictor of water mass residency. Variability in otolith chemistry reflected the spatial patterns of both horizontal gradients and vertical gradients in water mass properties related to circulation. Neither ambient temperature nor salinity must necessarily relate to otolith chemistry in order to be useful for discriminating fish from different water masses; rather, we suggest that temperature and salinity are useful as identifiers of distinct water masses that are associated with unique otolith elemental signature in the residing fish. We found it sufficient to use only temperature and salinity to discriminate among water masses; however, utilizing other parameters such as 02 and nutrients (often measured in oceanographic surveys) will likely improve our ability to identify water masses and refine future investigations of this kind. If shown to be reliable (i.e., temporally consistent), then the development of a reference or atlas of water mass-specific otolith elemental signatures is promising. Potentially, the use of an atlas of water mass-specific otolith signatures with water mass monitoring and current mapping could greatly reduce the uncertainty surrounding reconstructions of larval and adult movements and connectivity estimates among locations such as spawning and nursery areas and marine reserves.
References
Ashford JR, Jones CM, Hofmann E, Everson I, Moreno C, Duhamel G, Williams R (2005) Can otolith elemental signatures record the capture site of Patagonian toothfish (Dissostichus eleginoides), a fully marine fish in the Southern Ocean? Can J Fish Aquat Sci 62:2832–2840
Bath G, Thorrold SR, Jones CM, Campana SE, McLaren JW, Lam JWH (2000) Strontium and barium uptake in aragonitic otoliths of marine fish. Geochim Cosmochim Acta 64:1705–1714
Ben-Tzvi O, Abelson A, Gaines SD, Sheehy MS, Paradis GL, Kiflawi M (2007) The inclusion of sub-detection limit LAICPMS data, in the analysis of otolith microchemistry, by use of a palindrome sequence analysis (PaSA). Limnol Oceanogr Methods 5:97–105
Bograd SJ, Lynn RJ (2001) Physical-biological coupling in the California current during the 1997–99 El Niño-La Niña cycle. Geophys Res Lett 28:275–278
Brown JA (2006) Classification of juvenile flatfishes to estuarine and coastal habitats based on the elemental composition of otoliths. Estuar Coast Shelf Sci 66:594–611
Campana SE (1999) Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Mar Ecol Prog Ser 188:263–297
Campana SE, Chouinard GA, Hanson JM, Frechet A, Brattey J (2000) Otolith elemental fingerprints as biological tracers of fish stocks. Fish Res 46:343–357
Chan LH, Drummond D, Edmond JM, Grant B (1977) On the barium data from the Atlantic GEOSECS expedition. Deep Sea Res 24(7):613–649
Dove SG, Kingsford MJ (1998) Use of otoliths and eye lenses for measuring trace-metal incorporation in fishes: a biogeographic study. Mar Biol 130:377–387
Elsdon TS, Gillanders BM (2002) Interactive effects of temperature and salinity on otolith chemistry: challenges for determining environmental histories of fish. Can J Fish Aquat Sci 59:1796–1808
Elsdon TS, Gillanders BM (2003) Relationship between water and otolith elemental concentrations in juvenile black bream Acanthopagrus butcheri. Mar Ecol Prog Ser 260:263–272
Elsdon TS, Gillanders BM (2004) Fish otolith chemistry influenced by exposure to multiple environmental variables. J Exp Mar Biol Ecol 313:269–284
Elsdon TS, Gillanders BM (2005a) Alternative life-history patterns of estuarine fish: barium in otoliths elucidates freshwater residency. Can J Fish Aquat Sci 62:1143–1152
Elsdon TS, Gillanders BM (2005b) Consistency of patterns between laboratory experiments and field collected fish in otolith chemistry: an example and applications for salinity reconstructions. Mar Freshw Res 56:609–617
Esser BK, Volpe AM (2002) At-sea high-resolution chemical mapping: extreme barium depletion in North Pacific surface water. Mar Chem 79:67–79
FitzGerald JL, Thorrold SR, Bailey KM, Brown AL, Severin KP (2004) Elemental signatures in otoliths of larval walleye pollock (Theragra chalcogramma) from the northeast Pacific Ocean. Fish Bull 102:604–616
Forrester GE (2005) A field experiment testing for correspondence between trace elements in otoliths and the environment and for evidence of adaptation to prior habitats. Estuaries 28:974–981
Fowler AJ, Campana SE, Jones CM, Thorrold SR (1995a) Experimental assessment of the effect of temperature and salinity on the elemental composition of otoliths using solution-based ICPMS. Can J Fish Aquat Sci 52:1421–1430
Fowler AJ, Campana SE, Jones CM, Thorrold SR (1995b) Experimental assessment of the effect of temperature and salinity on the elemental composition of fish otoliths using laser ablation ICPMS. Can J Fish Aquat Sci 52:1431–1441
Friederich G, Walz P, Burczynski M, Chavez FP (2002) Inorganic carbon in the central California upwelling system during the 1997–1999 El Nino-La Nina event. Prog Oceanogr 54:185–203
Gillanders BM (2002) Temporal and spatial variability in elemental composition of otoliths: implications for determining stock identity and connectivity of populations. Can J Fish Aquat Sci 59(4):669–679
Hamer PA, Jenkins GP, Gillanders BM (2003) Otolith chemistry of juvenile snapper Pagrus auratus in Victorian waters: natural chemical tags and their temporal variation. Mar Ecol Prog Ser 263:261–273
Hamilton SL, Regetz J, Warner RR (2008) Postsettlement survival linked to larval life in a marine fish. Proc Nat Acad Sci USA 105:1561–1566
Hoff GR, Fuiman LA (1995) Environmentally induced variation in elemental composition of red drum (Scianops ocellatus) otoliths. Bull Mar Sci 56:578–591
Hurst MP, Bruland KW (2008) The effects of the San Francisco Bay plume on trace metal and nutrient distributions in the Gulf of the Farallones. Geochim Cosmochim Acta 72:395–411
Lea DW, Shen GT, Boyle EA (1989) Coralline barium records temporal variability in equatorial Pacific upwelling. Nature 340:373–376
Lea DW, Mashiotta TA, Spero HJ (1999) Controls on magnesium and strontium uptake in planktonic foraminifera determined by live culturing. Geochim Cosmochim Acta 63:2369–2379
Lloyd DC, Zacherl DC, Walker S, Paradis G, Sheehy M, Warner RR (2008) Egg source, temperature and culture seawater affect elemental signatures in Kelletia kelletii larval statoliths. Mar Ecol Prog Ser 353:115–130
Lochte K, Pfannkuche O (1987) Cyclonic cold-core eddy in the eastern North Atlantic. II. Nutrients, phytoplankton and bacterioplankton. Mar Ecol Prog Ser 39:153–164
Love MS, Yoklavich M, Thorsteinson L (2002) The rockfishes of the northeast Pacific. University of California Press, Berkeley, p 404
Martin GB, Thorrold SR (2005) Temperature and salinity effects on magnesium, manganese, and barium incorporation in otoliths of larval and early juvenile spot Leiostomus xanthurus. Mar Ecol Prog Ser 293:223–232
Mertes LAK, Warrick JA (2001) Measuring flood output from 110 coastal watersheds in California with field measurements and SeaWiFS. Geology 29:659–662
Miller JA, Shanks AL (2004) Evidence for limited larval dispersal in black rockfish (Sebastes melanops): implications for population structure and marine reserve design. Can J Fish Aquat Sci 61:1723–1735
Milton DA, Chenery SR, Farmer MJ, Blaber SJM (1997) Identifying the spawning estuaries of the tropical shad, terubok Tenualosa toli, using otolith microchemistry. Mar Ecol Prog Ser 153:283–291
Nishimoto MM, Washburn L (2002) Patterns of coastal eddy circulation and abundance of pelagic juvenile fish in the Santa Barbara Channel, California, USA. Mar Ecol Prog Ser 241:183–199
Nozaki Y (1997) A fresh look at element distribution in the North Pacific Ocean. EOS Trans Am Geophys Union 78(21):221
Patterson HM, Thorrold SR, Shenker JM (1999) Analysis of otolith chemistry in Nassau grouper (Epinephelus striatus) from the Bahamas and Belize using solution-based ICP-MS. Coral Reefs 18:171–178
Pearce NJG, Perkins WT, Westgate JA, Gorton MJ, Jackson SE, Neal CR, Chenery SP (1997) A compilation of new and published major and trace element data for NIST SRM 610 and NIST SRM 612 glass reference materials. Geostandard Newslett 21:115–144
Ruttenberg BI, Hamilton SL, Hickford MJH, Paradis GL et al (2005) Elevated levels of trace elements in cores of otoliths and their potential for use as natural tags. Mar Ecol Prog Ser 297:273–281
Sakuma KM, Schwing FB, Pickett MH, Roberts D, Ralston S (2000) The physical oceanography off the central California coast during May–June 1998: a summary of CTD data from pelagic juvenile rockfish surveys. US Dep Commer NOAA Tech Memo, NOAA-TM-NMFS-SWFSC 297:1–77
Secor DH, Henderson-Arzapalo A, Piccoli PM (1995) Can otolith microchemistry chart patterns of migration and habitat utilization in anadromous fishes? J Exp Mar Biol Ecol 192:15–33
SPSS Inc (2002) SPSS for Windows. Release 11.5.0
Standish JD, Sheehy M, Warner RR (2008) Use of otolith natal elemental signatures as natural tags to evaluate connectivity among open-coast fish populations. Mar Ecol Prog Ser 356:259–268
Swan SC, Geffen AJ, Morales-Nin B, Gordon JDM, Shimmield T, Sawyer T, Massutı E (2006) Otolith chemistry: an aid to stock separation of Helicolenus dactylopterus (bluemouth) and Merluccius merluccius (European hake) in the Northeast Atlantic and Mediterranean. ICES J Mar Sci 63:504–513
Swearer SE, Caselle JE, Lea DW, Warner RR (1999) Larval retention and recruitment in an island population of a coral reef fish. Nature 402:799–802
Systat Software Inc (2006) SigmaPlot for Windows Version 10.0
Thorrold SR, Latkoczy C, Swart PK, Jones CM (2001) Natal homing in a marine fish metapopulation. Science 291:297–299
Tipper ET, Galy A, Gaillardet J, Bickle MJ, Elderfield H, Carder EA (2006) The magnesium isotope budget of the modern ocean: constraints from riverine magnesium isotope ratios. Earth Planet Sci Lett 250:241–253
Warner RR, Swearer SE, Caselle JE, Sheehy M, Paradis G (2005) Natal trace-elemental signatures in the otoliths of an open-coast fish. Limnol Oceanogr 50:1529–1542
Warn-Varnas A, Gangopadhyay A, Hawkins JA (2007) Water masses in the Monterey Bay during the summer of 2000. Cont Shelf Res 27:1379–1398
White JW, Ruttenberg BI (2007) Discriminant function analysis in marine ecology: some common oversights and their solutions. Mar Ecol Prog Ser 329:301–305
Wyllie Echeverria T, Lenarz WH, Reilly CA (1990) Survey of the abundance and distribution of pelagic young-of-the-year rockfishes, Sebastes, off central California. US Dep Commer NOAA Tech Memo, NOAA-TM-NMFS-SWFSC 147:1–125
Zacherl DC (2005) Spatial and temporal variation in statolith and protoconch trace elements as natural tags to track larval dispersal. Mar Ecol Prog Ser 290:145–163
Zacherl DC, Paradis G, Lea D (2003) Ba and Sr uptake into larval protoconchs and statoliths of the marine neogastropod Kelletia kelletii. Geochim Cosmochim Acta 67:4091–4099
Zumholz K, Klugel A, Hansteen T, Piatkowski U (2007) Statolith microchemistry traces the environmental history of the boreoatlantic armhook squid Gonatus fabricii. Mar Ecol Prog Ser 333:195–204
Acknowledgements
This work was funded by NOAA Grant # NA06RG0142, California Sea Grant College Program Project # R/CZ-178, through NOAA’s National Sea Grant College Program, U.S. Dept. of Commerce. The microchemistry facility was supported in part by the Partnership for the Interdisciplinary Study of Coastal Oceans funded by the David and Lucile Packard Foundation. California Artificial Reef Enhancement Foundation provided additional matching funds. MMN received support from the U.S. Minerals Management Service. We thank D. Lea for providing ICPMS laboratory resources, and M. Sheehy for guidance on otolith microchemistry procedures. We thank K. Sakuma for otolith samples and the catch and oceanographic data from NOAA’s central California surveys (NMFS/SWFSC, Santa Cruz, CA). Funding for the southern California trawling survey was provided by the Biological Resources Division of the U.S. Geological Survey under cooperative agreement 1445-CA09-95-0836. The California Department of Fish and Game provided the midwater trawl net. E. Cheng and A. Muto, high school interns in the UCSB Research Mentorship Summer Program, assisted in the laboratory. We thank S. Ralston, M. Carr, and anonymous reviewers for their useful comments on the manuscript. The statements, findings, conclusions and recommendations are those of the authors and do not necessarily reflect the views of California Sea Grant or the U.S. Dept. of Commerce.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Nishimoto, M.M., Washburn, L., Warner, R.R. et al. Otolith elemental signatures reflect residency in coastal water masses. Environ Biol Fish 89, 341–356 (2010). https://doi.org/10.1007/s10641-010-9698-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-010-9698-6