
Abstract
Fecal sterols are traditionally ascribed as important biomarkers for animal excrement, and have been widely used to identify the source of organic matter and to reconstruct paleoecological changes in Antarctic terrestrial, aquatic, and marine ecosystems. However, the in situ microbial hydrogenation of cholesterol to coprostanol could have significance as a proxy to infer paleoenvironmental studies in Antarctica, particularly in anoxic sediment. Here, we report that abundant coprostanol, which was traditionally deemed as a biomarker for human sewage contamination, was found in three anoxic sediment profiles (AC2, BI, and CH1), which were strongly influenced by animal excrement at North Victoria Land, western Ross Sea, Antarctica. Our results suggest that the high concentrations of coprostanol in these three sediment profiles were not due to animal excrement, since coprostanol is not present in penguin guano and is a minor component in seal excrement. Coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) ratios suggest that coprostanol in the sediment cores of AC2 and BI was primarily derived from bacterial hydrogenation of cholesterol introduced by penguin guano. Coprostanol in CH1 sediments is related to human sewage due to intensive research activities from 1968 to 2006 in this region. However, the low abundance of coprostanol and the ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) in a relatively oxidizing sediment core (IIL1) infer that coprostanol was likely contributed by seal settlement. Together with high-throughput sequencing of the 16S rRNA gene, the conversion of cholesterol to coprostanol by anaerobic bacteria (e.g., Eubacterium coprostanoligenes) could occur in anoxic aquatic systems. Our results suggest that the presence of coprostanol in Antarctic lacustrine sediment with anoxic conditions does not necessarily indicate seal activity and human waste as the microbial hydrogenation of cholesterol to coprostanol should also be considered.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Lipid biomarkers are ubiquitously distributed and excellently preserved in soils and aquatic sediment, and have been regarded as a suitable proxy for paleoclimate and paleoecological studies at different time scales (Castañeda and Schouten 2011; Huang and Meyers 2019; Summons et al. 2021; Chen et al. 2022). Among these compounds, fecal sterols are important biomarkers for mammal excrement and have been widely used to indicate sewage contamination in industrial and population centers over the past several decades (Schönning et al. 2002; Martins et al. 2018; Cabral et al. 2019). With increasing scientific expeditions and tourism in polar regions, fecal sterols are also used to assess possible impacts of human activities on regional environments (Venkatesan and Mirsadeghi 1992; Martins et al. 2002, 2014; Leeming et al. 2015; Hargan et al. 2018) as well as the migration history of early human and seabirds (D'Anjou et al. 2012; Cheng et al. 2016; Vachula et al. 2019).
Mammals and birds exhibit different distribution characterizations of fecal sterols (Bull et al. 2002). For example, cholesterol comprises over 98% of total sterols in fresh penguin guano (Martins et al. 2002; Leeming et al. 2015) and generally is the most abundant sterol in lacustrine sediment influenced by animal excrement (Ceschim et al. 2016). Cholestanol is regarded as the product of microbial hydrogenation of cholesterol by aerobic bacteria (Bull et al. 2002). The variation of cholesterol and cholestanol contents in ornithogenic sediment profiles has been successfully used to reconstruct historical changes of penguin populations in Antarctica (Wang et al. 2007; Huang et al. 2010; Hu et al. 2013; Zheng et al. 2022). Coprostanol and epi-coprostanol are predominantly distributed in human waste accounting for 50% of total sterols, while campestanol and stigmasterol are high in mammal excrement (e.g., cows and sheep; D'Anjou et al., 2012). Therefore, the ratio of coprostanol/stigmasterol is deemed as an indicator for activities of humans and animals in polar region (D'Anjou et al. 2012; Vachula et al. 2019). In addition, fecal sterols (e.g., coprostanol) were also present in marine mammal (e.g., whales, seals; Leeming et al. 2015) feces and have been used as a proxy for delineating historical seal population changes in Antarctica (Huang et al. 2011). Considering the importance of fecal sterols in evaluating human activities and paleoecological studies in polar regions, it is particularly important to accurately identify their sources and biogeochemical transformation processes in sediment.
According to a previous study, there are three possible sources for fecal sterols (coprostanol) in Antarctic aquatic sediment including human waste, marine mammal excrement, and microbial hydrogenation of sterols by bacteria under anoxic sedimentary conditions (Leeming et al. 2015). For example, human activities such as the discharge of sewage are the predominant source of fecal sterols in the vicinity of the research stations in Antarctica (Montone et al. 2010; Martins et al. 2014, 2018). Besides human activities, marine mammals (e.g., seals) also can produce significant quantities of coprostanol (Venkatesan and Santiago 1989; Leeming et al. 2015). Beside the two above-mentioned sources, in situ microbial hydrogenation of sterols by bacteria under anoxic conditions can contribute significantly to the coprostanol in aquatic sediment. As reviewed by Kenny et al. (2020), cholesterol can be converted to coprostanol by human gut microbiota under anoxic condition. For example, many anaerobic bacteria (genus Eubacterium, Bifidobacterium, Clostridium, and Lactobacillus) can convert cholesterol to coprostanol (Gérard et al. 2007; Kriaa et al. 2019; Kenny et al. 2020; Juste and Gérard 2021). Actually, the conversion of cholesterol to coprostanol through the hydrogenation process was observed in anoxic conditions (Gaskell and Eglinton 1975, 1976; Nishimura 1982; Jen and Han 1996; Speranza et al. 2018). For example, high concentration of coprostanol was found in anaerobic sludge digestion due to the diagenetic transformation of cholesterol (Ahmad and Eskicioglu 2019). Nonetheless, no research combined with sequencing data and redox proxies has been conducted to confirm whether cholesterol can be converted to coprostanol by microorganisms in Antarctic lacustrine sediment. Furthermore, the factors that control the conversion of cholesterol to coprostanol remains elusive.
To answer the questions on whether such conversion can occur and how in situ sediment conditions determine the conversion of cholesterol, we chose four sediment profiles (AC2, BI, CH1, and IIL1) collected at North Victoria Land, Ross Sea, Antarctica. We explored the fecal sterols in these sediment profiles, and analyzed different environmental samples including fresh penguin guano and seal excrement, ornithogenic soils, microbial mats, terrestrial moss and soils for comparative purposes. We also compared fecal sterol distributions with the redox proxy such as Oxygen Index, active iron Fe(III)/Fe(II) and Fe/Mn ratios and bacterial diversity obtained by the high-throughput sequencing of 16S rRNA gene. This study aimed to accurately identify the source of fecal sterols in Antarctic lacustrine sediment including human and marine mammal wastes and microbial transformation, and to evaluate whether the conversion of cholesterol to coprostanol by bacteria occurs under anoxic conditions. Our study helps understand the biochemical transformation of fecal sterols in Antarctic lacustrine sediment, and lays the foundation for further reconstructing paleo-environmental and paleo-ecological changes in Antarctica.
Material and methods
Setting and sampling
The Ross Sea is a deep embayment in Antarctica with an average depth of 500 m. The weather in this region is extremely severe due to the presence of three different air masses from the Ross Sea, the Ross Ice Shelf, and Victoria Land (Monaghan et al. 2005). The annual mean air temperature is -18 °C, and the temperature can reach -50 °C and 8 °C in austral winter and summer time, respectively (Nie et al. 2016).
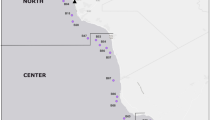
All these four sediment cores (AC2, BI, CH1 and IIL1) were collected using PVC tubes (about 12 cm diameter) that were hammered into the sediment layer. After transport back to the laboratory, all these sediment cores were sectioned at 0.5 cm. AC2 sediment profile was sampled from a small pond at Adélie Cove (74°46′05.00″S, 164°00′57.7″E), Ross Sea in February 2016 (Fig. 1; Supplementary Table S1). There is no research station in this region. More than 11,438 Adélie penguin breeding pairs inhabit Adélie Cove (Humphries et al. 2017). The detailed description of the sampling site has been reported by Povero et al. (2002). Specifically, Adélie Cove is a small deep V-shaped bay located in coastal Terra Nova Bay. The climate is extremely severe because of strong katabatic winds with low air temperature and precipitation. Specifically, the core (16.5 cm in length) was generally dark gray with a foul smell, and penguin eggshells and feathers were found throughout the sediment profile.

Study area and sampling site on the North Victoria Land, western Ross Sea, Antarctica. (a) Ross Sea. (b) Beaufort Island. (c) Inexpressible Island
The Beaufort Island (BI) sediment core (166°58′23.6″E, 76°58′23.6″S) was collected from a small catchment near an active Adélie penguin colony in January 2010 (Fig. 1). Beaufort Island (166°58′ E, 76°58′ S) is 5 km long and about 2.5 km wide. Most of the island is covered by snow and ice and the maximum elevation is 771 m above sea level (masl). No research stations exist on this island. More than 60,000 breeding pairs of Adélie penguins inhabit Beaufort Island, most on the southwest end, with a small Emperor penguin colony at the north end as well (Seppelt et al. 1999; Lynch and LaRue 2014). There are some small lakes or ponds near the Adélie penguin colony at the southwest end of the island that receive input from penguin guano. Our previous studies indicated that the source of organic matter in the BI sediment profile mainly derived from penguin guano and aquatic algae (Chen et al. 2013; Liu et al. 2013; Nie et al. 2014; Chen et al. 2020). The detailed description of the sampling site and sectioning of the sediment profile used in this study have been described by Nie et al. (2012). Briefly, the core BI (20.5 cm in length) gradually transforms from brown clay into black fine-grained ornithogenic soils from the bottom to top. The chronology of the BI sediment profile was determined by AMS14C and 210Pb-137Cs dating, with the results reported in our previous study (Chen et al. 2015), and the inferred age of bottom section from the BI profile was ~ 1640 AD.
CH1 sediment core was collected from a small pond located at Cape Hallett (72°19′S; 170°16′E), Ross Sea in January 2016 (Fig. 1). Cape Hallett peninsula is about 32 km long and 8 km wide including four parts based on landscape including Seabee Hook, Flat, Summit and Scree slope (Green et al. 2015). Seabee Hook is almost completely occupied by an Adélie penguin colony and a large number of abandoned penguin colonies can be found in the Flat area. More than 60,000 breeding pairs occupy on the Seabee Hook (Gordon 2003). There formerly was a research station built by American and New Zealand personnel in 1968, but it was abandoned in 2006. The climate is extremely cold with the highest monthly average air temperatures at -1.35 °C (January). Core CH1 (15 cm in length) is also largely influenced by penguin guano with a generally dark gray color and foul smell. Abundant penguin remains (eggshells, feathers and bones) can be found in the sediment profile. Additionally, core CH1 is near (about 500 m) the abandoned research station.
The IIL1 sediment core was collected from small pond on Inexpressible Island, Terra Nova Bay in January 2016. A detailed description of the sampling site is provided by Wei et al. (2016) and Chen et al. (2019). Inexpressible Island has an area of approximately 50 km2 (Fig. 1). A large number of abandoned Adélie penguin colonies are located there (Baroni and Orombelli 1994; Emslie and Patterson 2007), and an active colony of more than 20,000 breeding pairs occupy ice-free terrain surrounding Seaview Bay (He et al. 2017). The detailed sectioning of the IIL1 sediment profile has been reported by Wei et al. (2016). Core IIL1 (54 cm in length) is largely influenced by penguin guano with a dark color and foul smell below 26 cm. From 26 to 20 cm depth, the influence of penguin guano gradually decreased. The top 20 cm of the IIL1 sediment core is mainly microbial mat deposition with a gray-yellow color. Eggshells, bones and feathers can be found in the sediment profile. The age-depth model of the IIL1 sediment profile has been published in a previous study (Jin et al. 2021), and the bottom (oldest) age is dated to ~ 1659 year BP.
For comparison, two fresh Adélie penguin guano samples were collected from Cape Crozier, Ross Island (Table S1); two fresh seal excrements were collected from East Antarctica (Zhongshan station) and the Antarctic Peninsula (Great Wall Station, King George Island); three ornithogenic soils and two soils under moss samples were sampled at North Victoria Land. Terrestrial moss samples were sampled from Fildes Peninsula (King George Island, Antarctic Peninsula) and Campo Icarus, and Cape Irizar in the Ross Sea (Table S1). All samples were kept at - 20 °C prior to analysis.
Analysis of physicochemical properties
The Oxygen Index, Fe(III)/Fe(II), and Fe/Mn data are cited from our previous studies (Lou et al. 2016; Zheng et al. 2022). Oxygen Index represents the oxygen content of the organic matter, and can be used to reflect the redox nature of sediment (Meyers and Lallier-Vergès, 1999; Jacob et al. 2004). The OI value is calculated by the sum of CO (S3CO) and CO2 (S3CO2) released during pyrolysis, normalized to total organic carbon (TOC). The TOC was determined by potassium dichromate oxidation-capacity (Liu et al. 2013). Rock–Eval analysis followed the procedure described in Sanei and Goodarzi (2006), and was measured at the State Key Laboratory of Organic Geochemistry, Guangzhou Institute of Geochemistry, Chinese Academy of Sciences (RockEval-6 Standard, Vinci Technologies). Active iron is extremely sensitive to the variability of sedimentary environment and Fe(III)/Fe(II) values reflect the redox conditions in aquatic sediment (Haese et al. 1997). The analysis of active iron in the BI sediment profile was followed by the method of Haese et al. (1997). The Fe/Mn ratios are good indicator of redox conditions in lacustrine sediment with a higher value indicating a reducing environment (García-Rodríguez et al. 2021). The composition of Fe and Mn was measured by inductively coupled plasma optical emission spectrometry (ICP-OES) after microwave digestion (Perkin Elmer 2100DV; Zheng et al. 2022).
The contents of total organic matter in the BI and IIL1 sediment profiles are cited from Liu et al. (2013) and Wei et al. (2016), respectively. The δ13C values of total organic matter (δ13CTOC) in the BI and IIL1 sediment profiles were cited from our previous studies (Liu et al. 2013; Wei et al. 2016; Jin et al. 2021). The methods of carbon stable isotope analysis using the sealed tube combustion method were described in detail by Wei et al (2016) and Liu et al. (2013).
Lipid extraction and quantification
Organic geochemical analysis of AC2 and BI sediment cores and environmental samples (seal excrement, penguin guano, microbial mats, soils, terrestrial moss, and ornithogenic soils) in this study followed the general procedure described in Vachula et al. (2019). Briefly, the freeze-dried samples (~ 1 g) were homogenized and extracted with dichloromethane (DCM): methanol (9:1 by volume) using an accelerated solvent extractor (Dionex ASE 200) at Brown University, Providence, RI. The total lipid extract was dried under N2 and separated into acid (4% acetic acid in ether) and neutral (DCM: isopropanol) fractions using aminopropylsilyl gel columns. The neutral fraction was further separated into N1 (Hexane), N2 (DCM), N3 (Hexane: Ethyl acetate; 3:1 by volume), N4 (MeOH) by flash column chromatography using silica gel (40–63 μm, 60 Å). The N3 fraction was silylated with N,O-bis(trimethylsilyl) trifluoroacetamide (BSTFA) prior to quantification.
Analytical methods for CH1 and IIL1 sediment followed the general procedure from Cheng et al. (2016). Briefly, about 0.1 g freeze-dried sediment samples were sonicated with 10 mL dichloromethane (DCM) for 10 min with activated copper, repeated three times. The total lipid extract was concentrated to ~ 0.5 mL under N2 at room temperature and was cleaned using LC-Si SPE column eluted with 15 mL DCM. The eluted fraction including sterols and stanols was dried under a gentle flow of N2, CH1 and IIL1 sediment samples were analyzed using high temperature Gas Chromatography & Q Exactive Orbitrap Mass Sepctrometer.
Previous studies have compared the extraction efficiency between ASE and ultrasonication (Heemken et al. 1997), which showed that the recoveries of these two methods are equal; thus, our methods do not affect our interpretation in this study. The samples were quantified using an Agilent 6890N GC-FID system with an Agilent DB-1 column (30 m × 320 μm × 0.17 μm). The GC test method followed Longo et al. (2013). The initial temperature was 50 °C, followed by heating at 20 °C min−1 to 255 °C, then heating at 3 °C min−1 to 300 °C, and finally heating at 10 °C min−1 to 320 °C where it was held for 15 min. The lipid concentrations of the samples were determined using the peak areas obtained from the GC-FID results. The same GC column and oven temperature program were used for GC–MS analyses of selected samples for compound identification, which were performed on an Agilent 6890 GC interfaced to a 5973N quadruple mass spectrometer. Compounds were identified by comparison of GC retention times and mass spectra with published literature (Vachula et al. 2019). Compounds were quantified using an internal standard 5α-cholestane and by the proportional relation of the peak areas for each biomarker (supplementary materials, Tables S2–S6 and Fig. S1). The concentration of cholesterol and cholestanol in BI sediment profile was published by Zheng et al. (2022).
DNA extraction, PCR, sequencing and data analysis
DNA was extracted from 0.5 g of frozen sediment (AC2, CH1, and IIL1) using a FastDNA SPIN Kit for Soil (MP Biomedicals, Solon, OH, USA) following the standard protocols as previously described (Zhao et al. 2018; Chen et al. 2021). BI sediment profile was not analyzed due to lack of material. The concentration of each DNA extract was checked using a NanoDrop ND-2000 spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE, USA). The 16S rRNA gene (V4 region) was amplified by primers 515F/806R using the Illumina MiSeq platforms (Bates et al. 2011). Target PCR products were then sequenced on an Illumina HiSeq 2500 sequencer (PE 250) at the Magigene Biotechnology Co Ltd (Guangzhou, China).
Sequencing data were processed using the software QIIME2 (version: 2021.2) (Bolyen et al. 2019). Specifically, sequences were denoised and clustered to amplicon sequence variants (ASVs) by DADA2 (Callahan et al. 2016), the dada2 denoise-single method with the parameters –p-trim-left 56, –p-trunc-len 315 was applied during the analysis. The generated ASVs were assigned taxonomy by the classify-sklearn method of q2-feature-classifier plugin (Bokulich et al. 2018) against SILVA reference alignment release 138 (Quast et al. 2013). The ASVs identified to be the "Eubacterium_coprostanoligenes_group" in the Silva database were selected to build the phylogenetic tree, and the full length 16S rRNA references sequences were downloaded from Silva database (https://www.arb-silva.de/). The sequences were aligned using MAFFT (Katoh and Standley 2013) and then trimmed by trimAL (Capella-Gutiérrez et al. 2009) with -automated1 parameter. The phylogenetic tree was built using IQ-TREE 2 (Minh et al. 2020) with the model MFP, 1000 ultrafast bootstraps and visualized by tvBOT (Xie et al. 2023).
Results
Fecal sterols distribution in environmental samples
The abundance of cholesterol in fresh seal excrement and penguin guano was very high, with the ratio of cholesterol/total sterols over 98% (Table 1). Most of seal excrement contained less than 1% coprostanol, and coprostanol and epi-coprostanol were not detected in penguin guano (Table 1), while cholesterol accounted for over 90% of the total sterols. Cholesterol was also predominant in microbial mats, soils, terrestrial moss, and ornithogenic soils sampled from modern penguin colony, with a relative abundance over 95% (Table 1), but coprostanol was not detected in all these environmental samples.
Fecal sterols distribution in lacustrine sediment profiles
Four C27 fecal sterols (cholesterol, cholestanol, coprostanol, and epi-coprostanol), were detected in the AC2, BI, CH1, and IIL1 sediment profiles (Table 2). The most abundant sterols were cholesterol in the AC2, BI, and IIL1 sediments, but coprostanol for CH1 sediment samples (Supplementary Figs. S2 and S3). Epi-coprostanol was not detected in the CH1 and IIL1 sediment.
The ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) can be used as organic proxies to trace the sources of fecal sterols in aquatic sediment (Leeming et al. 2015). In this study, the coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) ratios exhibited obviously different values in these sediment with higher values in CH1 and lower in IIL1, but AC2 and BI had similar values (Table 2 and Fig. 2). Specifically, the ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) varied from 0.1 to 1.4 and 0.4 to 0.8 in the AC2 sediment core, with an average of 0.5 and 0.6, respectively. These two ratios ranged from 0.5 to 1.0 and 0.4 to 0.7 in the BI sediment core, with averages of 0.8 and 0.6, respectively. In the CH1 sediments, coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) displayed large variations from 3.1 to 10.8 and 0.9 to 1.0, with averages of 6.6 and 0.9, respectively. The IIL1 sediment core was characterized by small ranges with an average of 0.1 and 0.3 for these two ratios, respectively.

The ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) in the AC2, BI, CH1 and IIL1 sediment collected in Ross Sea, Antarctica. The error bars are the 1σ standard deviations of all measured samples in the sediment profiles. The red area represents the ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) in seal excrement in Antarctica
The diversity of bacteria
The taxonomic compositions of bacteria in the AC2, CH1 and IIL1 sediment profiles are given in the supplementary materials (Figs. S4–S6). The dominant bacteria are different in these three explored sediment core. Specifically, Actinobacteriota (relative abundance varying from 6.1 to 15.6%), Bacteroidota (13.5 to 23.1%), Chloroflexi (6.1 to 19.5%), Firmicutes (12.5 to 38.8%), Proteobacteria (12.7 to 33.8%) dominated (relative abundance > 5%) in the AC2 sediment core (Supplementary Figure S4). In the CH1 sediment core, Bacteroidota (6.7 to 19.4%), Firmicutes (37.3 to 81.7%), Proteobacteria (3.5 to 28.3%) were the most abundant bacteria (Figure S5). Actinobacteriota (4.9 to 29.3%), Bacteroidota (2.9 to 35.2%) and Proteobacteria (45.2 to 86.9%) were predominant in the IIL1 sediment core (Fig. S6). We subsequently selected the sequences of ASVs identified to be the "Eubacterium_coprostanoligenes_group" in all the sequenced samples to build the phylogenetic tree (Fig. S7), and it indicates that the selected ASVs clustered together with the reference sequences of Eubacterium coprostanoligenes. Among these three explored sediment profiles, Eubacterium coprostanoligenes can be detected in the AC2 and CH1 sediment profiles, with relative abundance to total sequences ranging from 0 to 1.1%, and 0 to 1.2%, respectively. However, Eubacterium coprostanoligenes cannot be found in the IIL1 sediment profile.
Discussion
Distributions and sources of fecal sterols in sediments
Previous studies showed that BI and IIL1 sediment cores are significantly influenced by penguin guano according to inorganic elements associated with penguin guano (e.g., phosphorus), C/N ratios, and carbon isotope ratios of total organic matter (Liu et al. 2013; Wei et al. 2016; Jin et al. 2021; Zheng et al. 2022). For example, higher concentration of P, and lower δ13CTOC and C/N values indicate the input of animal guano-derived organic matter accounted for a larger proportion in the BI and IIL1 sediment profiles (Liu et al. 2013; Jin et al. 2021). Cholesterol is the most abundant sterol, comprising over 98% of the total sterols in fresh penguin guano (Leeming et al. 2015), consistent with our analysis of fresh Adélie penguin guano where cholesterol was higher than 99% (Table 1). According to comparative study, the concentrations of cholesterol in these four sediment profiles (AC2, BI, CH1, and IIL1) were similar with the fresh animal excrement and ornithogenic soil samples, while much higher than that of microbial mats, terrestrial moss, and soils (Supplementary Tables S2–S6). The above analyses are consistent with our field investigations that active colonies were observed at these four study area during our field investigations. Cholestanol in penguin guano is usually minor because penguins have no metabolic mechanism that can convert cholesterol to cholestanol (Martin et al. 1973). However, high abundance of cholestanol was also found in these four explored sediment profiles and likely originated from the microbial-mediated conversion of cholesterol in ornithogenic soil under aerobic conditions (Bull et al. 2002; Wang et al. 2007; Wisnieski et al. 2014). We assumed that, therefore, cholesterol in these four sediment profiles was mainly derived from animal excrement.
Coprostanol and epi-coprostanol were also found in the AC2, BI, CH1 and IIL1 sediment profiles (Table 2, Supplementary Figs. S2 and S3). According to many previous studies (Martins et al. 2002, 2014; Leeming et al. 2015), all three potential sources including human wastes, seal activities, and microbial hydrogenation of cholesterol to coprostanol by bacteria in anoxic conditions should be considered in Antarctic lacustrine sediment. The ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) can be used to distinguish the sources of fecal sterols in sediment (Fig. 2; Fattore et al. 1996; Leeming et al. 1996; Martins et al. 2014; Araújo et al. 2021). For example, the coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) values with the range of 0 to 1 and 0 to 0.5, respectively are inferred to be biogenic sources, while the ratio values more than 1 and 0.5 are strongly related to sewage input. The values of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) vary from 0 to 1 and 0.5 to 1, respectively, may indicate a microbial hydrogenation contribution. In this study, the ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) displayed large ranges in the four sediment profiles (Figs. 2, 3), indicating coprostanol may have different sources. We discuss the potential sources separately based on the ratios of fecal sterols.

The average values of coprostanol abundance (%), coprostanol/cholesterol ratio, and OI in four sediment profiles. The limit values of coprostanol/cholesterol on the Y-axis are different in the AC2, BI, CH1, and IIL1 sediment profiles. The error bars are the 1σ standard deviations of all measured samples in the sediment profiles
Sources of coprostanol in the AC2 and BI sediment profiles
High abundances of coprostanol and epi-coprostanol in the AC2 and BI sediment profiles indicate that they may be strongly influenced by human sewage because these compounds are generally associated with human fecal matter (Bull et al. 2002). However, Beaufort Island and Adélie Cove are protected reserves and no human occupations have been occasionally established there except temporary camps, and these locations are also distant from research stations. Furthermore, the ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) in AC2 and BI sediment cores were obviously lower than the values of sewage contamination (Fig. 2), indicating that coprostanol cannot be attributed to anthropogenic sources (Martins et al. 2014). Importantly, the bottom age of the BI sediment profile is ~ 1640 AD, suggesting no human activities at that time (Chen et al. 2015). Given these constraints, therefore, all results indicate that the high abundances of coprostanol and epi-coprostanol in the AC2 and BI sediment cores could not be attributed to anthropogenic sources.
The other likely source of the coprostanol would be derived from either marine mammals (seals) hauling out on the beach for breeding, resting, or molting, or microbial hydrogenation of sterols by bacteria in anoxic sediment. According to our previous studies, the extremely low δ13C values of organic matter in the BI sediment profile suggest that the total organic matter in the sediments mainly originated from animal excrement transported by melt-water erosion and runoff (Liu et al. 2013). However, seal hairs were not found throughout the AC2 and BI sediment profiles, though small numbers of Weddell and Elephant seals have occasionally been observed resting or molting on the beach (Hall et al. 2006; Mezgec et al. 2017; Koch et al. 2019). The AC2 sediment profile was collected in a small pond located on a highland that seals cannot access. Thus, we contemplated that the high levels of coprostanol and epi-coprostanol in the AC2 and BI sediment profiles were not caused by a large input of seal excrement, also supported by the distribution of sterols in fresh seal excrement. Leeming et al. (2015) reported that Elephant seal excrement had high cholesterol levels that accounted for more than 99% of the total sterols, with coprostanol and epi-coprostanol below 1%. This finding is similar to our results that the relative abundance of cholesterol is higher than 96% in seal excrement (Table 1). However, the average abundance of coprostanol is ~ 30% in the AC2 and BI sediment, and obviously higher than that of seal excrement. The different ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) between sediment and seal excrement infer that such high abundance of coprostanol may not be directly derived from seal excrement (Fig. 2). Additionally, some active Adélie penguin colonies were found at AC2 and BI sampling sites during our field investigations. On the contrary, the microbial-mediated bio-hydrogenation of cholesterol to coprostanol in deposited penguin guano should be considered.
At present, two major pathways for the conversion of cholesterol to coprostanol by gut microbiota include direct stereospecific reduction and an indirect pathway involving the intermediate formation of 4-cholelesten-3-one and coprostanone (Kriaa et al. 2019; Juste and Gérard 2021). For example, Juste and Gérard (2021) systematically summarized the associated active strains isolated from human and animal feces, and a hog sewage lagoon. Researchers have isolated a few bacteria belonging to the genus Eubacterium, Bifidobacterium, Clostridium, and Lactobacillus, which can convert cholesterol to coprostanol (Ren et al. 1996; Kriaa et al. 2019; Mukherjee et al. 2020). In this study, the Eubacterium coprostanoligenes can also be found throughout the AC2 sediment core (Supplementary Fig. S7). Additionally, the relative abundance of the Eubacterium coprostanoligenes has similar change pattern with the concentration of coprostanol in the AC2 sediment profile (Fig. 4). These results support our inference that cholesterol can be converted to coprostanol by specific bacteria in sediment.

The changes of cholesterol and coprostanol concentrations, coprostanol/cholesterol, Fe/Mn, OI values, and relative abundance of Eubacterium coprostanoligenes in the AC2 sediment profile
Sources of coprostanol in the CH1 sediment profile
For the CH1 sediment profile, the most abundant fecal sterol is coprostanol with an average abundance of ~ 80% (Table 2), obviously higher than in animal excrement and the other three explored sediment profiles (AC2, BI and IIL1). The ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) in the CH1 sediment profile are in the ranges of human fecal contamination (Fig. 2; Martins et al. 2014), indicating human sewage may be an important source of coprostanol. Wastewater from intensive human activities at the former research station at this location may have been discharged into ponds or catchments, thus accounting for the large amount of fecal sterols observed in the CH1 sediments. Many previous studies reported that abundant coprostanol was detected in Antarctic marine sediments near research stations, and relatively high ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) indicated it was primarily derived from sewage discharge (Martins et al. 2002, 2014; Leeming et al. 2015). The redox proxies (e.g., OI and Fe/Mn ratio) all indicate more suboxic sedimentary conditions in the CH1 sediment profile. Moreover, similar with AC2 sediment, sequences assigned to Eubacterium coprostanoligenes were also detected in the CH1 sediment core (Supplementary Figs. S7 and S8), suggesting microbial hydrogenation of cholesterol to coprostanol may also occur. Unfortunately, it is very difficult to distinguish the sources from sewage discharge and the microbial hydrogenation of cholesterol to coprostanol.
Sources of coprostanol in the IIL1 sediment profile
In the IIL1 sediment, the average abundance of coprostanol is lower than 10%, while cholesterol is the most abundant with an average abundance ~ 80% (Table 2). The IIL1 sediment was collected from a small pond, near many abandoned colonies of Adélie penguins on Inexpressible Island (Wei et al. 2016). Most importantly, many penguin remains including eggshells, bones, and feathers were found throughout the IIL1 sediment profile, indicating a strong influence from occupation by this species (Jin et al. 2021). Thus, it is reasonable to predict that high abundance of cholesterol is mainly derived from penguin guano. The ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) in the IIL1 sediment profile fall between the ranges of seal excrement and penguin guano (Fig. 2), suggesting the small portion of coprostanol may be contributed by seal activities in this area. As reported by previous studies, the data of ancient DNA and 14C dating of elephant seal remains indicate this seal was present on Inexpressible Island from ~ 3270 to 380 BP, but likely only as occasional molting animals (Hall et al. 2006; Koch et al. 2019; Jin et al. 2021). We conclude that relative low abundance of coprostanol in the IIL1 sediment profile may have been contributed by seal use of the beaches for molting. These results are supportive of our sequencing data that the associated bacteria (e.g., Eubacterium coprostanoligenes) were not found in the IIL1 sediment profile. Furthermore, coprostanol is minor in the IIL1 sediment core, indicating that microbial hydrogenation of cholesterol to coprostanol did not occur to a noticeable extent.
The transformation of cholesterol to coprostanol in Antarctic lacustrine sediments
A number of laboratory experiments have demonstrated that cholesterol can be converted to coprostanol by gut microbiota in higher mammals (Kenny et al. 2020; Juste and Gérard 2021). In natural environments, the conversion of cholesterol to coprostanol was also reported in aquatic sediment under anoxic conditions (Gaskell and Eglinton 1975; Nishimura 1982; Jeng and Han 1996). For example, Nishumura (1982) reported that cholesterol could be transformed to coprostanol in anoxic aquatic sediment, which was controlled by the relative contribution of autochthonous organic matter. In Antarctic lakes influenced by animal excrements, higher organic matter inputs and degradations can consume oxygen, and form more suboxic sedimentary conditions (Lou et al. 2016). More suboxic conditions are necessary for this conversion to occur in lacustrine sediment (Nishimura 1982; Bull et al. 2002; Leeming et al. 2015). The Oxygen Index (OI), active iron Fe(III)/Fe(II) and Fe/Mn ratios are sensitive to the variability of redox sedimentary conditions in aquatic sediment (Haese et al. 1997; Jacob et al. 2004; Williams and Scherer 2004). For example, lower OI value, and higher Fe/Mn and Fe(II)/Fe(III) ratios usually indicate more suboxic sedimentary conditions. In this study, all these proxies in the AC2, BI, and CH1 sediment profiles indicated that these sediment were deposited under more suboxic conditions compared with IIL1 sediment profile (Fig. 3; Lou et al. 2016), which provided a potential sedimentary environment favorable for microbial hydrogenation (Gaskell and Eglinton 1975; Nishimura 1982). The concentrations of coprostanol actually show similar patterns of change with redox proxies in the AC2 and BI sediment profiles (Figs. 4 and 5), indicating more suboxic sedimentary conditions are suitable for microbial hydrogenation. By contrast, low abundance of coprostanol and low ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) in the IIL1 sediment profile with more oxic sedimentary condition are not suitable for microbial hydrogenation. Additionally, Hu et al. (2013) analyzed fecal sterols in a sediment profile (MB6) with less suboxic condition near penguin colonies at Cape Bird, Ross Sea. Abundant cholesterol and cholestanol were found in the top 32 cm, but coprostanol and epi-coprostanol were absent (Hu et al. 2013; Lou et al. 2016). Given these estimates, we conclude that high abundance of coprostanol was finally formed through microbial hydrogenation of cholesterol in the AC2 and BI sediment profiles under more suboxic conditions.

The changes of cholesterol and coprostanol concentrations, δ13CTOC, coprostanol/cholesterol, Fe(III)/Fe(II) and OI values in the BI sediment profile. The data of δ13CTOC are cited from Liu et al. (2013). The data of OI and Fe(III)/Fe(II) was cited from Lou et al. (2016). The concentration of cholesterol was cited from Zheng et al. (2022)
The concentrations of coprostanol exhibit large ranges versus depth in the AC2 and BI sediment profiles (Figs. 4 and 5), suggesting different microbial hydrogenation strength of cholesterol to coprostanol. We found that coprostanol concentrations showed similar change patterns with guano input strength and redox sedimentary conditions (Figs. 4 and 5). High input intensity of guano can carry more organic matter and nutrition into ponds, resulting in an anoxic sedimentary condition in the AC2 and BI sediment profiles (Lou et al. 2016). These favorable conditions can cause the occurrence of bio-conversion of cholesterol to coprostanol in lacustrine sediment. Our results suggest that the higher concentrations of coprostanol are in line with more suboxic conditions and stronger guano input in the AC2 and BI sediment profiles, indicting more favorable sedimentary conditions for this conversion. However, isotope labeling cultures should be performed to further verify the transformation of cholesterol to coprostanol in natural sediment.
Conclusion
Abundant C27 fecal sterols (cholesterol, cholestanol, coprostanol, and epi-coprostanol) were found in four lacustrine sediment profiles (AC2, BI, CH1, and IIL1) that were strongly influenced by animal excrement. Cholesterol is the most abundant sterol in the AC2, BI, and IIL1 sediment profiles, while coprostanol is only predominant in CH1. Based on the ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol), the cholesterol in the IIL1 sediment profile is mainly derived from penguin guano, while a small part of coprostanol is contributed by seal feces at Inexpressible Island. However, abundant coprostanol is found in the AC2 and BI sediment profiles, which were strongly influenced by penguin guano. The ratios of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) and lack of seal hairs strongly suggest that coprostanol in the AC2 and BI sediment profiles was mainly derived from the microbial hydrogenation of cholesterol from penguin guano. However, in the CH1 sediment, the high values of coprostanol/cholesterol and coprostanol/(coprostanol + cholestanol) indicate that coprostanol is likely related to human sewage due to research activities from 1968 to 2006 at Hallett Station.
Compared with the isolated cholesterol-reducing bacterium, we found the cholesterol-reducing bacterium (Eubacterium coprostanoligenes) present throughout the two more suboxic sediment profiles (AC2 and CH1), but absent in the sediment profile (IIL1) with minor amounts of coprostanol. Our data indicated that cholesterol was likely converted to coprostanol and epi-coprostanol via microbial hydrogenation processes by bacteria under anoxic aquatic systems in Antarctica. Therefore, caution should be taken when using coprostanol to infer seal activities and human waste in Antarctic anoxic ponds or lakes.
Data availability
All data were present in the Supplementary Materials.
Data availability
Data will be made available on reasonable request.
References
Ahmad M, Eskicioglu C (2019) Fate of sterols, polycyclic aromatic hydrocarbons, pharmaceuticals, ammonia and solids in single-stage anaerobic and sequential anaerobic/aerobic/anoxic sludge digestion. Waste Manag 93:72–82
Araújo MP, Hamacher C, de Oliveira Farias C, Soares MLG (2021) Fecal sterols as sewage contamination indicators in Brazilian mangroves. Mar Pollut Bull 165:112149
Baroni C, Orombelli G (1994) Abandoned penguin rookeries as Holocene paleoclimatic indicators in Antarctica. Geology 22:23–26
Bates ST, Berg-Lyons D, Caporaso JG, Walters WA, Knight R, Fierer N (2011) Examining the global distribution of dominant archaeal populations in soil. ISME J 5:908–917
Bokulich NA, Kaehler BD, Rideout JR, Dillon M, Bolyen E, Knight R, Huttley GA, Gregory Caporaso J (2018) Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 6:1–17
Bolyen E, Rideout JR, Dillon MR, Bokulich NA, Abnet CC, Al-Ghalith GA, Alexander H, Alm EJ, Arumugam M, Asnicar F, Bai Y, Bisanz JE, Bittinger K, Brejnrod A, Brislawn CJ, Brown CT, Callahan BJ, Caraballo-Rodriguez AM, Chase J, Cope EK, Da Silva R, Diener C, Dorrestein PC, Douglas GM, Durall DM, Duvallet C, Edwardson CF, Ernst M, Estaki M, Fouquier J, Gauglitz JM, Gibbons SM, Gibson DL, Gonzalez A, Gorlick K, Guo J, Hillmann B, Holmes S, Holste H, Huttenhower C, Huttley GA, Janssen S, Jarmusch AK, Jiang L, Kaehler BD, Kang KB, Keefe CR, Keim P, Kelley ST, Knights D, Koester I, Kosciolek T, Kreps J, Langille MGI, Lee J, Ley R, Liu YX, Loftfield E, Lozupone C, Maher M, Marotz C, Martin BD, McDonald D, McIver LJ, Melnik AV, Metcalf JL, Morgan SC, Morton JT, Naimey AT, Navas-Molina JA, Nothias LF, Orchanian SB, Pearson T, Peoples SL, Petras D, Preuss ML, Pruesse E, Rasmussen LB, Rivers A, Robeson MS II, Rosenthal P, Segata N, Shaffer M, Shiffer A, Sinha R, Song SJ, Spear JR, Swafford AD, Thompson LR, Torres PJ, Trinh P, Tripathi A, Turnbaugh PJ, Ul-Hasan S, van der Hooft JJJ, Vargas F, Vazquez-Baeza Y, Vogtmann E, von Hippel M, Walters W, Wan Y, Wang M, Warren J, Weber KC, Williamson CHD, Willis AD, Xu ZZ, Zaneveld JR, Zhang Y, Zhu Q, Knight R, Caporaso JG (2019) Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol 37:852–857
Bull ID, Lockheart MJ, Elhmmali MM, Roberts DJ, Evershed RP (2002) The origin of faeces by means of biomarker detection. Environ Int 27:647–654
Cabral AC, Wilhelm MM, Figueira RC, Martins CC (2019) Tracking the historical sewage input in South American subtropical estuarine systems based on faecal sterols and bulk organic matter stable isotopes (δ13C and δ15N). Sci Total Environ 655:855–864
Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJ, Holmes SP (2016) DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods 13:581–583
Capella-Gutiérrez S, Silla-Martínez JM, Gabaldón T (2009) trimAl: a tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 25:1972–1973
Castañeda IS, Schouten S (2011) A review of molecular organic proxies for examining modern and ancient lacustrine environments. Quat Sci Rev 30:2851–2891
Ceschim LM, Dauner AL, Montone RC, Figueira RC, Martins CC (2016) Depositional history of sedimentary sterols around Penguin Island. Antarctica Antarct Sci 28:443–454
Chen Q, Nie Y, Liu X, Xu L, Emslie SD (2015) An 800-year ultraviolet radiation record inferred from sedimentary pigments in the Ross Sea area, East Antarctica. Boreas 44:693–705
Chen X, Dong L, Zhao W, Jian H, Wang J, Wang F (2022) The effects of metabolism and temperature on carbon isotope composition of lipids in marine bacterium Shewanella piezotolerans WP3. Chem Geol 606:120963
Chen X, Liu X, Jia H, Jin J, Kong W, Huang Y (2021) Inverse hydrogen isotope fractionation indicates heterotrophic microbial production of long-chain n-alkyl lipids in desolate Antarctic ponds. Geobiology 19:394–404
Chen X, Liu X, Wei Y, Huang Y (2019) Production of long-chain n-alkyl lipids by heterotrophic microbes: new evidence from Antarctic lakes. Org Geochem 138:103909
Chen X, Wei Y, Nie Y, Wang J, Emslie SD, Liu X (2020) Carbon isotopes of n-alkanoic acids in Antarctic ornithogenic sediments as indicators of sedimentary lipid sources and paleocological change. Sci Total Environ 709:135926
Cheng W, Sun L, Kimpe LE, Mallory ML, Smol JP, Gallant LR, Li J, Blais JM (2016) Sterols and stanols preserved in pond sediments track seabird biovectors in a High Arctic environment. Environ Sci Technol 50:9351–9360
D’Anjou RM, Bradley RS, Balascio NL, Finkelstein DB (2012) Climate impacts on human settlement and agricultural activities in northern Norway revealed through sediment biogeochemistry. Proc Natl Acad Sci USA 109:20332–20337
Emslie SD, Patterson WP (2007) Abrupt recent shift in δ13C and δ15N values in Adélie penguin eggshell in Antarctica. Proc Natl Acad Sci USA 104:11666–11669
Fattore E, Benfenati E, Marelli R, Cools E, Fanelli R (1996) Sterols in sediment samples from Venice Lagoon, Italy. Chemosphere 33:2383–2393
García-Rodríguez F, Piccini C, Carrizo D et al (2021) Centennial glacier retreat increases sedimentation and eutrophication in Subantarctic periglacial lakes: a study case of Lake Uruguay. Sci Total Environ 754:142066
Gaskell SJ, Eglinton G (1975) Rapid hydrogenation of sterols in a contemporary lacustrine sediment. Nature 254:209–211
Gaskell SJ, Eglinton G (1976) Sterols of a contemporary lacustrine sediment. Geochim Cosmochim Acta 40:1221–1228
Gérard P, Lepercq P, Leclerc M, Gavini F, Raibaud P, Juste C (2007) Bacteroides sp. strain D8, the first cholesterol-reducing bacterium isolated from human feces. Appl Environ Microbiol 73:5742–5749
Gordon S (2003) Site description and literature review of Cape Hallett and surrounding areas. Antarctica New Zealand, Latitude Gradient Project, Internal report, pp 1–16
Green TG, Seppelt RD, Brabyn LR, Beard C, Türk R, Lange OL (2015) Flora and vegetation of Cape Hallett and vicinity, northern Victoria Land, Antarctica. Polar Biol 38:1825–1845
Haese RR, Wallmann K, Dahmke A, Kretzmann U, Müller PJ, Schulz HD (1997) Iron species determination to investigate early diagenetic reactivity in marine sediments. Geochim Cosmochim Acta 61:63–72
Hall BL, Hoelzel AR, Baroni C, Denton GH, Le Boeuf BJ, Overturf B, Topf AL (2006) Holocene elephant seal distribution implies warmer-than-present climate in the Ross Sea. Proc Natl Acad Sci USA 103:10213–10217
Hargan KE, Stewart EM, Michelutti N, Grooms C, Kimpe LE, Mallory ML, Smol JP, Blais JM (2018) Sterols and stanols as novel tracers of waterbird population dynamics in freshwater ponds. Proc R Soc B 285:20180631
He H, Cheng X, Li X, Zhu R, Hui F, Wu W, Zhao T, Kang J, Tang J (2017) Aerial photography based census of Adélie Penguin and its application in CH4 and N2O budget estimation in Victoria Land, Antarctic. Sci Rep 7:12942
Heemken OP, Theobald N, Wenclawiak BW (1997) Comparison of ASE and SFE with soxhlet, sonication, and methanolic saponification extractions for the determination of organic micropollutants in marine particulate matter. Anal Chem 69:2171–2180
Hu QH, Sun LG, Xie ZQ, Emslie SD, Liu XD (2013) Increase in penguin populations during the Little Ice Age in the Ross Sea, Antarctica. Sci Rep 3:2472
Huang J, Sun L, Huang W, Wang X, Wang Y (2010) The ecosystem evolution of penguin colonies in the past 8,500 years on Vestfold Hills, East Antarctica. Polar Biol 33:1399–1406
Huang J, Sun L, Wang X, Wang Y, Huang T (2011) Ecosystem evolution of seal colony and the influencing factors in the 20th century on Fildes Peninsula, West Antarctica. J Environ Sci 23:1431–1436
Huang X, Meyers PA (2019) Assessing paleohydrologic controls on the hydrogen isotope compositions of leaf wax n-alkanes in Chinese peat deposits. Palaeogeogr Palaeoclimatol Palaeoecol 516:354–363
Humphries GRW, Naveen R, Schwaller M, Che-Castaldo C, McDowall P, Schrimpf M, Lynch HJ (2017) Mapping application for penguin populations and projected dynamics (MAPPPD): data and tools for dynamic management and decision support. Polar Rec 53:160–166
Jacob J, Disnar JR, Boussafir M, Sifeddine A, Turcq B, Albuquerque ALS (2004) Major environmental changes recorded by lacustrine sedimentary organic matter since the last glacial maximum near the equator (Lagoa do Caçó, NE Brazil). Palaeogeogr Palaeoclimatol Palaeoecol 205:183–197
Jeng WL, Han BC (1996) Coprostanol in a sediment core from the anoxic Tan-Shui estuary, Taiwan Estuarine. Coast Shelf Sci 42:727–735
Ji M, Kong W, Jia H, Delgado-Baquerizo M, Zhou T, Liu X, Ferrari BC, Malard L, Liang C, Xue K, Makhalanyane TP, Zhu Y, Wang Y, Pearce DA, Cowan D (2022) Polar soils exhibit distinct patterns in microbial diversity and dominant phylotypes. Soil Biol Biochem 166:108550
Jin J, Chen X, Xu L, Nie Y, Wang X, Huang H, Emslie SD, Liu X (2021) Chronology and paleoclimatic implications of lacustrine sediments at Inexpressible Island, Ross Sea, Antarctica. Palaeogeogr Palaeoclimatol Palaeoecol 576:110497
Juste C, Gérard P (2021) Cholesterol-to-coprostanol conversion by the gut microbiota: what we know, suspect, and ignore. Microorg 9:1881
Katoh K, Standley DM (2013) MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 30:772–780
Kenny DJ, Plichta DR, Shungin D, Koppel N, Hall AB, Fu B, Vasan RS, Shaw SY, Vlamakis H et al (2020) Cholesterol metabolism by uncultured human gut bacteria influences host cholesterol level. Cell Host Microbe 28:245–257
Koch PL, Hall BL, de Bruyn M, Hoelzel AR, Baroni C, Salvatore MC (2019) Mummified and skeletal southern elephant seals (Mirounga leonina) from the Victoria Land Coast, Ross Sea, Antarctica. Mar Mammal Sci 35:934–956
Kriaa A, Bourgin M, Mkaouar H, Jablaoui A, Akermi N, Soussou S, Maguin E, Rhimi M (2019) Microbial reduction of cholesterol to coprostanol: an old concept and new insights. Catal 9:167
Leeming R, Ball A, Ashbolt N, Nichols P (1996) Using faecal sterols from humans and animals to distinguish faecal pollution in receiving waters. Water Res 30:2893–2900
Leeming R, Stark JS, Smith JJ (2015) Novel use of faecal sterols to assess human faecal contamination in Antarctica: a likelihood assessment matrix for environmental monitoring. Antarct Sci 27:31–43
Liu X, Nie Y, Sun L, Emslie SD (2013) Eco-environmental implications of elemental and carbon isotope distributions in ornithogenic sediments from the Ross Sea region, Antarctica. Geochim Cosmochim Acta 117:99–114
Longo WM, Dillon JT, Tarozo R, Salacup JM, Huang Y (2013) Unprecedented separation of long chain alkenones from gas chromatography with a poly (trifluoropropylmethylsiloxane) stationary phase. Org Geochem 65:94–102
Lou C, Liu X, Liu W, Wu L, Nie Y, Emslie SD (2016) Distribution patterns and possible influencing factors of As speciation in ornithogenic sediments from the Ross Sea region, East Antarctica. Sci Total Environ 553:466–473
Lynch HJ, LaRue MA (2014) First global census of the Adélie penguin. Auk 131:457–466
Martin WJ, Ravi Subbiah MT, Kottke BA, Birk CC, Naylor MC (1973) Nature of fecal sterols and intestinal bacterial flora. Lipids 8:208–215
Martins CC, Aguiar SN, Wisnieski E, Ceschim LM, Figueira RC, Montone RC (2014) Baseline concentrations of faecal sterols and assessment of sewage input into different inlets of Admiralty Bay, King George Island, Antarctica. Mar Pollut Bull 78:218–223
Martins CC, Castellanos-Iglesias S, Cabral AC, de Souza AC, Ferraz MA, Alves TP (2018) Hydrocarbon and sewage contamination near fringing reefs along the west coast of Havana, Cuba: a multiple sedimentary molecular marker approach. Mar Pollut Bull 136:38–49
Martins CC, Venkatesan MI, Montone RC (2002) Sterols and linear alkylbenzenes in marine sediments from Admiralty Bay, King George Island, South Shetland Islands. Antarct Sci 14:244–252
Meyers PA, Lallier-Vergès E (1999) Lacustrine sedimentary organic matter records of Late Quaternary paleoclimates. J Paleolimnol 21:345–372
Mezgec K, Stenni B, Crosta X, Masson-Delmotte V, Baroni C, Braida M, Ciardini V, Colizza E, Melis R, Salvatore MC, Severi M, Scarchilli C, Traversi R, Udisti R, Frezzotti M (2017) Holocene Sea ice variability driven by wind and polynya efficiency in the Ross Sea. Nat Commun 8:1–12
Minh BQ, Schmidt HA, Chernomor O, Schrempf D, Woodhams MD, Von Haeseler A, Lanfear R (2020) IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol Biol Evol 37:1530–1534
Monaghan AJ, Bromwich DH, Powers JG, Manning KW (2005) The climate of the McMurdo, Antarctica, region as represented by one year of forecasts from the Antarctic Mesoscale Prediction System. J Clim 18:1174–1189
Montone RC, Martins CC, Bícego MC, Taniguchi S, da Silva DAM, Campos LS, Weber RR (2010) Distribution of sewage input in marine sediments around a maritime Antarctic research station indicated by molecular geochemical indicators. Sci Total Environ 408:4665–4671
Mukherjee A, Lordan C, Ross RP, Cotter PD (2020) Gut microbes from the phylogenetically diverse genus Eubacterium and their various contributions to gut health. Gut Microbes 12:1802866
Nie Y, Liu X, Emslie SD (2014) Distribution and sources of rare earth elements in ornithogenic sediments from the Ross Sea region, Antarctica. Microchem J 114:247–260
Nie Y, Liu X, Sun L, Emslie SD (2012) Effect of penguin and seal excrement on mercury distribution in sediments from the Ross Sea region, East Antarctica. Sci Total Environ 433:132–140
Nie Y, Xu L, Liu X, Emslie SD (2016) Radionuclides in ornithogenic sediments as evidence for recent warming in the Ross Sea region, Antarctica. Sci Total Environ 557:248–256
Nishimura M (1982) 5β-isomers of stanols and stanones as potential markers of sedimentary organic quality and depositional paleoenvironments. Geochim Cosmochim Acta 46:423–432
Povero P, Chiantore M, Misic C, Budillon G, Cattaneo-Vietti R (2002) Land forcing controls pelagic-benthic coupling in Adelie Cove (Terra Nova Bay, Ross Sea). Ecological studies in the Antarctic Sea Ice Zone. Springer, Berlin, pp 125–132
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner FO (2013) The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res 41:D590–D596
Ren D, Li L, Schwabacher AW, Young JW, Beitz DC (1996) Mechanism of cholesterol reduction to coprostanol by Eubacterium coprostanoligenes ATCC 51222. Steroids 61:33–40
Sanei H, Goodarzi F (2006) Relationship between organic matter and mercury in recent lake sediment: the physical–geochemical aspects. Appl Geochem 21:1900–1912
Schönning C, Leeming R, Stenström TA (2002) Faecal contamination of source-separated human urine based on the content of faecal sterols. Water Res 36:1965–1972
Seppelt RD, Green TGA, Skotnicki M (1999) Notes on the flora, vertebrate fauna and biological significance of Beaufort Island, Ross Sea, Antarctica. Polarforschung 66:53–59
Speranza ED, Colombo M, Skorupka CN, Colombo JC (2018) Early diagenetic alterations of sterol biomarkers during particle settling and burial in polluted and pristine areas of the Rio de la Plata Basin. Org Geochem 117:1–11
Summons RE, Welander PV, Gold DA (2021) Lipid biomarkers: molecular tools for illuminating the history of microbial life. Nat Rev Microbiol 20:1–12
Vachula RS, Huang Y, Longo WM, Dee SG, Daniels WC, Russell JM (2019) Evidence of Ice Age humans in eastern Beringia suggests early migration to North America. Quat Sci Rev 205:35–44
Venkatesan MI, Mirsadeghi FH (1992) Coprostanol as sewage tracer in McMurdo sound, Antarctica. Mar Pollut Bull 25:328–333
Venkatesan MI, Santiago CA (1989) Sterols in ocean sediments: novel tracers to examine habitats of cetaceans, pinnipeds, penguins and humans. Mar Biol 102:431–437
Wang J, Wang Y, Wang X, Sun L (2007) Penguins and vegetations on Ardley Island, Antarctica: evolution in the past 2,400 years. Polar Biol 30:1475–1481
Wei YY, Jin J, Nie YG, Chen X, Wu LB, Fu PQ, Liu XD, Emslie SD (2016) Sources of organic matter and paleo-environmental implications inferred from carbon isotope compositions of lacustrine sediments at Inexpressible Island, Ross Sea, Antarctica. Adv Polar Sci 27:233–244
Williams AGB, Scherer MM (2004) Spectroscopic evidence for Fe(II)-Fe(III) electron transfer at the iron oxide-water interface. Environ Sci Technol 38:4782–4790
Wisnieski E, Bícego MC, Montone RC, Figueira RC, Ceschim LM, Mahiques MM, Martins CC (2014) Characterization of sources and temporal variation in the organic matter input indicated by n-alkanols and sterols in sediment cores from Admiralty Bay, King George Island. Antarctica Polar Biol 37:483–496
Xie J, Chen Y, Cai G, Cai R, Hu Z, Wang H (2023) Tree Visualization By One Table (tvBOT): a web application for visualizing, modifying and annotating phylogenetic trees. Nucleic Acids Res 51:587
Zhao K, Kong W, Wang F, Long X-E, Guo C, Yue L, Yao H, Dong X (2018) Desert and steppe soils exhibit lower autotrophic microbial abundance but higher atmospheric CO2 fixation capacity than meadow soils. Soil Biol Biochem 127:230–238
Zheng Z, Nie Y, Chen X, Jin J, Chen Q, Liu X (2022) Historical population dynamics of the Adélie penguin in response to atmospheric-ocean circulation patterns at Beaufort Island, Ross Sea, Antarctica. Glob Planet Chang 216:103892
Acknowledgements
This work was jointly supported by a National Natural Science Foundation of China (Grant Nos. 42276240, 42206243, 41976191, and 41776188), the Strategic Priority Research Program of Chinese Academy of Sciences (Grant No. XDB40000000), the Shanghai Sailing Program (22YF1418800), the China Postdoctoral Science Foundation (2022M712038), the Shanghai Frontiers Science Center of Polar Science (SCOPS). We would like to thank the Chinese Arctic and Antarctic Administration of the State Oceanic Administration for project support. We also thank the United States Antarctic Program (USAP), Antarctic Support Contract and Italian Mario Zucchelli Station for logistical support. We thank Prof. Yongsong Huang, and Dr. Jiaju Zhao, and Hongzeng Jia for their kindly help in the experimental analyses.
Author information
Authors and Affiliations
Contributions
Xin Chen, Xiaodong Liu and Jianjun Wang proposed the topic, conceived and designed the study. Xin Chen, Jing Jin, Zhangqin Zheng, Yulu Xue, Weidong Kong conducted the experiments, and prepared the manuscript with contributions from all co-authors. All the co-authors contributed to the discussion, and edited and commented on the paper.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Responsible Editor: Christian Lønborg
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, X., Wang, J., Jin, J. et al. Microbial hydrogenation of cholesterol to coprostanol by anaerobic bacteria: evidence from Antarctic lacustrine sediment. Biogeochemistry (2024). https://doi.org/10.1007/s10533-024-01121-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10533-024-01121-7