
Abstract
We examined the effect of concentration on nitrogen uptake patterns for a suburban stream in Maryland and addressed the question: How does NO3 − uptake change as a function of concentration and how do uptake patterns compare with those found for NH4 +? We applied a longitudinal (stream channel corridor) approach in a forested stream section and conducted short-term nutrient addition experiments in late summer 2004. In the downstream direction, NO3 − concentrations decreased because of residential development in headwaters and downstream dilution; NH4 + concentrations slightly increased. The uptake patterns for NO3 − were very different from NH4 +. While NH4 + had a typical negative relationship between first-order uptake rate constant (K c ) and stream size, NO3 − had a reverse pattern. We found differences for other metrics, including uptake velocity (V f ) and areal uptake rate (U). We attributed these differences to a stream size effect, a concentration effect and a biological uptake capacity effect. For NO3 − these combined effects produced a downstream increase in K c , V f and U; for NH4 + they produced a downstream decrease in K c and V f , and a not well defined pattern for U. We attributed a downstream increase in NO3 − uptake capacity to an increase in hyporheic exchange and a likely increase in carbon availability. We also found that K c and V f were indirectly related with concentration. Similar evidence of ‘nutrient saturation’ has been reported in other recent studies. Our results suggest that higher-order uptake models might be warranted when scaling NO3 − uptake across watersheds that are subject to increased nitrogen loading.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In-stream processing is one factor influencing nitrogen export from suburbanizing watersheds. Many factors important in nitrogen processing are affected by stream size (Vannote et al. 1980) and therefore stream size often plays a role in the downstream fate of nitrogen (Alexander et al. 2000). Along the stream size spectrum, headwater streams are generally considered the most important control on watershed nitrogen export (Peterson et al. 2001; Alexander et al. 2007). Smaller streams typically have high uptake relative to transport, because of larger surface area to volume ratios and lower velocities. However, recent modeling studies suggest that larger rivers are perhaps equally important (Wollheim et al. 2006; Ensign and Doyle 2006), mainly because of long travel times, hence more opportunity for nitrogen removal.
Whole-stream studies of nutrient processing are often based on the concept of nutrient spiraling (Webster and Patten 1979; Newbold et al. 1981; Elwood et al. 1983) and associated field experiments typically involve isotopic tracer or nutrient additions. These studies have shown that uptake is strongly controlled by stream size or discharge (e.g., Peterson et al. 2001; Wollheim et al. 2001). Generalizations drawn from these and other studies (e.g., mass balance studies) have led to the development of simulation models of nutrient uptake and export. These models range from empirical to process-based. Of the latter, a commonly used approach is to assume a first-order process conceptualization, in which uptake rate is linearly related to concentration. Uptake is then estimated using a first-order uptake rate coefficient. Typically, this coefficient changes as a function of stream size (e.g., SPARROW model by Smith et al. 1997; Alexander et al. 2000). Relationships between this coefficient or other derived uptake metrics and stream size have been used to scale across stream networks (Wollheim et al. 2006; Ensign and Doyle 2006).
Concentration is a key variable in nitrogen uptake. In suburbanizing watersheds, nitrate (NO3 −) is generally the main nitrogen species of concern. NO3 − concentrations in these watersheds can exhibit large spatial variability, mainly governed by the location of high NO3 − sources, subsequent dilution with low NO3 − waters, and uptake. As NO3 − concentrations progressively increase, uptake kinetics could change such that uptake rate is no longer linearly related to concentration. Recent field studies have shown evidence of this nutrient saturation effect (Mulholland et al. 2008; Claessens et al. 2009b). This could have implications for modeling and quantifying NO3 − loss in these watersheds.
We conducted several studies to examine the role of small streams in controlling nitrogen export from a suburbanizing watershed in Maryland. In Claessens et al. (2009a) we examined seasonal variation in longitudinal ammonium (NH4 +) uptake patterns and found that organic matter distribution played an important role. We discussed that current practices in scaling nutrient cycling across stream networks do not necessarily scale the causal factors (e.g., organic matter dynamics), but rather base the scaling on derived uptake rates or other spiraling metrics. In this study we examined how uptake metrics are affected by concentration and addressed the following question. How does NO3 − uptake change as a function of concentration and how do uptake patterns compare with those found for NH4 +? We applied a longitudinal (stream channel corridor) approach in a forested stream section and conducted short-term nutrient addition experiments. We compared uptake patterns for NO3 − and NH4 + for experiments conducted in late summer 2004.
Study area
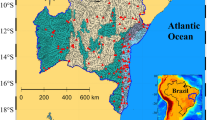
This study was conducted on the main stem of Baisman Run (BARN) and an 80 m reach of Pond Branch (POBR), which is a small tributary of BARN, located in Baltimore County in Maryland, about 15 km north of the city of Baltimore (Fig. 1). The 3.8 km2 BARN watershed and the 0.4 km2 POBR watershed are two of several watersheds monitored as part of the Baltimore Ecosystem Study Long Term Ecological Research program (BES-LTER). Land-use in the BARN watershed is characterized by low-density residential in the upper portion and forested in the lower portion (Fig. 1). POBR is entirely forested and serves as the forest reference watershed for the BES-LTER. BARN is particularly suited for investigating in-stream processing, as it has a step change in nitrate loading, with high loadings from the upper, developed portion and low loadings from the lower, forested portion. The BARN watershed has been the focus of several studies to examine nitrogen fluxes (Groffman et al. 2004) and the role of small streams in controlling nitrogen export from suburban land-use (Claessens et al. 2009a, b).

Baisman Run watershed with stream network (false color image using EMERGE digital aerial imagery). Lighter colored areas in the headwaters indicate development. Numbered sections correspond to location of nutrient addition experiments. POBR is Pond Branch reach
Methods
The nutrient addition experiments reported here were conducted in late summer 2004. We did two sets of short-term additions (NO3 − and NH4 +) in the main stem of BARN and one short-term NO3 − addition in POBR. The NH4 + additions in BARN are described in Claessens et al. (2009a), for which we used OTIS-MM (Claessens and Tague 2009), a transport-based nutrient addition approach adapted to account for the effect of nutrient saturation. The NO3 − addition in POBR is described in Claessens and Tague (2009), for which we also used OTIS-MM. The methodology presented here refers to the BARN NO3 − additions only, for which we used a standard first-order uptake approach (e.g., Stream Solute Workshop 1990). We did not use OTIS-MM because of high NO3 − background concentrations. We used short-term additions of a conservative solute (bromide) to estimate transport and transient storage (TS) characteristics, using the one-dimensional transport with inflow and storage (OTIS) solute transfer model (Runkel 1998). We used TS as a surrogate for the hyporheic zone, while acknowledging that TS includes both hyporheic exchange and surface storage.
We used a section (sequence of 3–5 reaches covered in a single addition; Fig. 1) as the defining spatial unit, instead of individual reaches. We did this because of analytical accuracy constraints associated with the high NO3 − background concentrations in BARN. For the BARN NH4 + additions (Claessens et al. 2009a) we used a stream reach as the defining spatial unit. In this paper we re-worked these NH4 + data to correspond with the sections used for the NO3 − additions.
Nutrient addition experiments
The NO3 − additions in BARN were conducted ~2 weeks after the NH4 + additions, for the same reaches under similar flow conditions. Samples were collected at the same station locations (i.e., reach boundaries), at background and plateau only. We conducted the experiments using an upstream progression, starting from the lower end of BARN, and all four sections were covered in 2 days of experiments. The NO3 − addition in the most upstream section (Section 1) failed because of a thunderstorm during the latter part of the experiment.
The injectate concentration was aimed at raising Br− background by 2–3 mg L−1 (background was 0.003–0.12 mg L−1); and NO3 − background by 0.14 mg N L−1 (background was 1.6–4.2 mg N L−1). For the injection duration we aimed for at least 2½ h plateau duration for the most downstream reach.
Sodium bromide and sodium nitrate were dissolved in DI water in the lab, transported in carboys to the field and injected using a peristaltic pump (Wheaton) powered by a marine battery. The pump was calibrated before each injection and checked post-injection (flow rate deviations were less than 1%). The solute was injected at a natural or temporary artificially constructed flow constriction, followed by a 20–25 m mixing reach. Sampling stations were located at upstream and downstream reach boundaries (six stations per section). Water samples were collected as grab samples in the thalweg, always in the same spot. At each station two samples were collected, at background and at plateau. The grab samples were collected using 250 ml plastic sample bottles that had been acid-washed and rinsed with stream water, and were immediately filtered (0.7 μm Whatman GF/F) using syringes and filter holders into 20 ml plastic scintillation vials that had been rinsed with filtered sample water. The samples were placed on ice and upon return from the field were kept either refrigerated (bromide) or frozen (nutrients).
The experiments were conducted during baseflow conditions. The additions started early in the morning and sampling was completed by mid-afternoon. Diurnal fluctuations in flow (because of evapotranspiration) were substantial (~10% decrease during the experiment) and were accounted for during the calibration of the OTIS model parameters. Wetted stream widths were measured every 10 m for a representative cross-section.
Bromide samples were analyzed on an ion chromatograph (Dionex DX-120) at the MBL Ecosystems Center in Woods Hole, MA. Nutrient samples were analyzed on a Lachat autoanalyzer at the Cary Institute of Ecosystem Studies in Millbrook, NY. Samples were run out of sequence to reduce analytical artifacts introduced by instrument drift and changes in standards and reagents.
Data analysis
Transport and transient storage
Transport and TS parameters for each section were estimated from the Br− concentrations using the OTIS solute transport model (Runkel 1998), which is based on the Bencala and Walters (1983) TS model. The following parameters were estimated: stream channel cross-sectional area (A), storage zone cross-sectional area (A s ), dispersion coefficient (D) and storage zone exchange coefficient (α). Parameters were estimated using a semi-automatic, nonlinear least square algorithm as part of OTIS-P. After model calibration we calculated the relative size of the TS zone (A s /(A + A s )). We also calculated the fraction of median reach travel time due to TS (F med), a transport-based TS metric introduced by Runkel (2002). We calculated F med by adopting a standardized median reach travel time of 18 min (F T18med ).
Uptake
NO3 − uptake parameters for each section were estimated using OTIS. Using the OTIS first-order uptake model we estimated the NO3 − first-order uptake rate constant, K c . NH4 + uptake parameters for each section were estimated using OTIS-MM. See Claessens and Tague (2009) for governing equations and method details. The following OTIS-MM NH4 + uptake parameters were estimated: lateral inflow reactive solute concentration (C L ), maximum uptake rate (U max) and half-saturation constant (K s ). Parameters were estimated using a semi-automatic, nonlinear least square algorithm built into OTIS-P, as well as through manual calibration. For each experiment, iterative calibrations were performed until convergence was achieved for all parameters.
Uptake metrics
We calculated the following interrelated uptake metrics (Stream Solute Workshop 1990):
where: K c is first-order uptake rate constant [T−1], which describes uptake on a volumetric basis; S w is uptake length [L], which is the average travel distance of nutrients before removal; U is uptake rate [M L−2 T−1], which describes uptake rate per unit area of stream bottom; V f is uptake velocity [L T−1], which describes the vertical velocity of nutrients towards the benthos; C is concentration [M L−3]; u is velocity [L T−1]; and h is water depth [L]. Fundamental units are mass [M], length [L] and time [T]. These basic uptake metrics are based on first-order kinetics and are not transport-based. To calculate these metrics, we used a simulation approach using either OTIS (NO3 −) or OTIS-MM (NH4 +). First, we estimated median velocity (u) from OTIS. Next, we calculated uptake lengths (S w ) for a sequence of OTIS (NO3 −) or OTIS-MM (NH4 +) simulations. Similar to empirical methods (e.g. Webster and Ehrman 1996), we determined S w by regressing plateau concentrations against distance, after correcting for background and dilution. A total of six simulations were performed with increasing levels of solute addition, from which we estimated S w at background concentration through extrapolation (similar to the approach suggested by Dodds et al. 2002). Uptake rate constant (K c ), uptake rate (U) and uptake velocity (V f ) were calculated subsequently using Eq. 1.
Results
Physical and hydrologic characteristics
The average (~100 m) channel gradient for the sections decreased downstream and ranged between 1.9 and 1.5% (Table 1). The channel roughness was highest in Section 2 and decreased downstream. Because flow and flow fluctuations were similar for the NO3 − and NH4 + addition experiments, we collected physical and hydrologic characteristics for the NH4 + addition experiments only. The sections ranged in discharge from 6 to 26 L s−1. Cross-sectional areas (solved by OTIS) ranged from 0.11 to 0.23 m2. Median velocities (determined from OTIS simulation) ranged from 0.05 to 0.10 m s−1 and were only minimally influenced by slower transport in the TS zone. Measured wetted stream widths ranged from 1.5 to 3.2 m and water depths (obtained by dividing cross section area by stream widths) were 7 cm for all sections. The relative size of the TS zone (A s /(A + A s )) was similar for all sections, ranging from 13 to 17%. The relative time that water spent in TS (F T18med ) was slightly more variable, ranging from 3 to 7%. Based on reach level data we found that F T18med generally increased downstream (results not shown).
Chemical characteristics
Background NO3 − concentrations were elevated because of residential development (Table 1; 30-fold difference between BARN and POBR) and decreased downstream (Fig. 2a). Although tributaries can shift the baseline, there is a consistent downstream decrease in concentration. This decrease is primarily due to dilution with low NO3 − water from tributaries and groundwater in the forested middle and lower catchment (Claessens et al. 2009b). Background NH4 + concentrations were low (Table 1; mean = 11 μg N L−1), similar to non-impacted systems, and increased downstream (Fig. 2b). Dissolved organic nitrogen (DON) concentrations generally increased downstream (Fig. 2c).

a Background NO3− concentrations for detailed synoptic (40 m) (September 6, 2004); b background NH4+ concentrations for sampling locations (September 1–4, 2004); c DON concentrations for detailed synoptic (~100 m) (September 6, 2004)
Comparison of uptake metrics against other studies
We compared the NO3 − and NH4 + uptake metrics against three published studies (Table 2). Hall et al. (2002) conducted NH4 + additions (n = 37) and Bernhardt et al. (2002) conducted NO3 − additions (n = 19) in the Hubbard Brook Experimental Forest (HBEF). Ensign and Doyle (2006) compiled an extensive data set of published nutrient addition results (n = 404; including both NO3 − and NH4 +). All three data sets largely contain measurements for relatively unimpacted systems. Average discharge in the two HBEF studies and median discharge in Ensign and Doyle (2006) were similar to our study. Hence, differences due to a possible stream size effect were minimized.
For NO3 − in Baisman Run, K c was a factor 4–9 smaller than these studies; V f was a factor 3–14 smaller; and U was a factor 15–44 larger. Apart from having elevated NO3 − concentration, these forested sections of BARN represent a fairly unimpacted stream. The large difference between BARN and these studies suggests that NO3 − uptake was affected by elevated NO3 − concentrations. For NO3 − in Pond Branch, K c was a factor 1–3 smaller; V f was a factor 2–8 smaller; and U was a factor 1–3 larger. This illustrates that NO3 − uptake in Pond Branch was similar to other unimpacted systems. For NH4 + in Baisman Run, K c was a factor 1–2 smaller; V f was a factor 1–2 smaller; and U was similar. This illustrates that NH4 + uptake in Baisman Run was similar to unimpacted systems. Even though NO3 − concentrations in BARN are elevated, this seems to have little effect on NH4 + uptake.
Spatial patterns of uptake metrics
Spatial patterns of uptake metrics were determined (Fig. 3). For BARN NO3 −, both K c and V f increased with discharge; and the patterns for U and S w were not well defined. For BARN NH4 +, both K c and V f decreased with discharge; the pattern for U was not well defined; and S w increased with discharge. POND values are shown for reference only.

NO3 − and NH4 + uptake terms vs. discharge. Squares are NO3 −; open squares are NO3 − for Pond Branch; triangles are NH4 +. Left axis is NO3 −; right axis is NH4 +
Concentration and uptake metrics
Nutrient uptake metrics were compared against their respective ambient nutrient concentrations (Fig. 4). For NO3 −, both K c and V f had a negative relationship with concentration (Fig. 4a, b). For comparison, in Fig. 4c, d we present values for NO3 − loss metrics obtained from a long-term mass balance study (Claessens et al. 2009b); similarly, both K c and V f for NO3 − loss had a negative relationship with concentration. Also for NH4 +, both K c and V f had a negative relationship with concentration (Fig. 4e, f). For NH4 + we present the reach-level data reported in Claessens et al. (2009a), because they span a larger range of background concentrations.

NO3 − and NH4 + uptake terms vs. their respective background concentrations. (a, b) NO3 − uptake (open squares are Pond Branch); (c, d) NO3 − loss, based on long-term mass balance study (Claessens et al. 2009b); (e, f) NH4 + uptake
Discussion
Spatial patterns of NO3 − and NH4 + uptake are controlled by physical/hydrologic, chemical and biological factors. In our discussion below we attribute distinct differences in uptake patterns to these factors, through a stream size effect, a concentration effect and an uptake capacity effect. In broad terms, the stream size effect corresponds to physical/hydrologic factors that affect surface area to volume ratios and residence time. The concentration effect corresponds to chemical factors that affect uptake kinetics (e.g., nutrient saturation). The uptake capacity effect corresponds to biological factors that affect nutrient demand (size and composition of stream biotic population). These factors are inter-related and their relative importance varies across stream size (Vannote et al. 1980). Hence, attributing any uptake pattern to one single effect or combination thereof is complicated.
Spatial patterns of uptake
For NH4 + there was a typical negative relationship between K c and stream size: as stream size increased, K c decreased. Similar relationships between K c and stream size are commonly used in simulation models of nitrogen uptake (Smith et al. 1997; Alexander et al. 2000). This stream size effect results from smaller streams having relatively larger surface area to volume ratios, hence more contact between water column and streambed, therefore larger K c . This stream size effect is a general pattern; at finer spatial scales NH4 + K c patterns do not necessarily follow this trend (e.g., see Claessens et al. 2009a).
Interestingly, NO3 − had a reverse pattern: as stream size increased, K c increased as well. This can partially be explained by the longitudinal pattern of NO3 − concentrations and its direct effect on K c . In BARN, NO3 − concentrations were highest in the headwaters and decreased downstream, mainly because of dilution with low NO3 − lateral inputs from forested areas. K c is based on a first-order process conceptualization, which describes a linear increase in uptake rate with an increase in concentration. Thus, for a given uptake capacity, a lower concentration produces a higher K c ; this is the concentration effect. Hence, for NO3 − the stream size effect was overwhelmed by factors that lead to an increase in K c , including the concentration effect. The increase in K c can only partially be attributed to the concentration effect, because the increase in K c (226%) was larger than the decrease in concentration (68%). Therefore, our results suggest that the increase in K c also reflects a downstream increase in NO3 − uptake capacity. This increase in uptake capacity was particularly evident in the lower portion.
The S w pattern for NO3 − was very different from NH4 +. S w patterns respond to both an uptake component (negative relationship with K c ) and an advection component (positive relationship with velocity) (see Eq. 1). In the case of NH4 + these two components acted complementary (decrease in K c and increase in velocity), resulting in a downstream increase in S w . In the case of NO3 − the two components acted in opposite direction (increase in K c and increase in velocity), resulting in a S w pattern that was not well defined. The fact that NO3 − S w strongly decreased in the lower portion clearly indicates that the uptake component dominated. This increase in the relative importance of the uptake component is due to the uptake capacity effect and the concentration effect, as discussed above.
Hence, distinct uptake patterns emerged in our study, which we attributed to a stream size effect, a concentration effect and an uptake capacity effect. For NO3 − the three effects acted as follows moving downstream: (1) the stream size effect reduced K c (reduced surface area to volume ratios); (2) the concentration effect increased K c (lower concentrations because of dilution); and (3) the uptake capacity effect increased K c (more organic carbon supply; see next section). The overall effect on NO3 − uptake was a downstream increase in K c , V f and U. In the case of NH4 + the three effects acted as follows moving downstream: (1) the stream size effect reduced K c (reduced surface area to volume ratios); (2) the concentration effect slightly reduced K c (higher concentrations because of mineralization); and (3) the uptake capacity effect slightly increased K c (more hyporheic exchange). The overall effect on NH4 + uptake was a downstream decrease in K c and V f and a not well defined pattern for U.
Effects of concentration and biological uptake capacity
The comparison between the NO3 − and NH4 + uptake patterns highlights the effects of concentration and biological uptake capacity. Both NO3 − and NH4 + had distinct concentration profiles. For NO3 − this profile reflects the location of high NO3 − sources in the headwaters and subsequent dilution downstream. Concentration has an effect on uptake kinetics, which vary across temporal scales. Over a short-term time scale (e.g., short-term nutrient addition or storm event nutrient pulse), uptake might display a saturation effect (e.g., Dodds et al. 2002). Over a longer-term time scale (e.g., long-term nutrient fertilization or suburbanization), uptake is also governed by biotic uptake capacity (microbial population) (e.g., Slavik et al. 2004). Given the concentration effect, the specific location of high NO3 − sources within the larger stream network plays an important role on uptake rates and ultimately NO3 − export from the watershed.
The stream biota seem to have different longitudinal variation in uptake capacities for NO3 − and NH4 +, with NO3 − having a larger downstream increase in uptake capacity compared to NH4 +. Both downstream increases in uptake capacities can be partially attributed to an increase in hyporheic exchange, which is consistent with general patterns in hyporheic development (Boulton et al. 1998) (i.e., least important in headwaters, peaking intermediate and decreasing in lowland streams). Also, the downstream increases in uptake capacities can be attributed to a likely increase in allochthonous organic matter (e.g., leaf litter), due to downstream transport and enhanced deposition because of downstream decrease in channel gradient. While NH4 + uptake capacity increased because of likely increased mineralization of organic matter, NO3 − uptake capacity increased because of likely higher organic carbon availability (e.g., Bernhardt and Likens 2002). Although we did not measure organic carbon, the increased availability is evidenced by the downstream increase in DON at the time of this study. Further, increased NO3 − uptake capacity suggests higher rates of denitrification. Measurements of NO3 − stable isotopes (L. Claessens, unpublished) showed clear evidence of denitrification (consistent longitudinal increase in both δ15N and δ18O, coinciding with a decrease in NO3 − concentration), both in headwaters and lower reaches. Overall, it suggests that the stream features responsible for biological uptake capacity (organic matter and hyporheic exchange) played a critical role in explaining spatial variation in NO3 − and NH4 + uptake. These spatial patterns can also display seasonal variation, as shown by Claessens et al. (2009a) for NH4 + uptake.
Implications for scaling
Uptake metrics are commonly used to scale nitrogen removal across river segments or entire river networks. Examples of river network nitrogen models that incorporate uptake metrics include SPARROW (Smith et al. 1997; Alexander et al. 2000), which applies predefined K c values to several stream size classes; and RivR-N (Seitzinger et al. 2002), which relates nitrogen removal to the inverse of travel time over depth. Wollheim et al. (2006) compared these and other models to examine river size dependence in biological activity and found that the models predicted a downstream increase in V f . This supports our empirical findings for NO3 − V f .
Our empirical results show distinct downstream patterns for NO3 − and NH4 + uptake metrics. Ambient concentration played an important role in these patterns, as illustrated in Fig. 4, which shows that K c and V f of both NO3 − and NH4 + had a negative relationship with concentration. Interestingly, for NO3 − the values for POBR fit the general trend for BARN (particularly for K c ). Although in the same biogeoclimatic setting, these two streams have large differences in NO3 − loading. The same figure also shows this concentration effect for a long-term mass balance study (Claessens et al. 2009b). That study reports net NO3 − uptake or loss (versus gross uptake in this study), and therefore the values for K c and V f were lower. Similar observations regarding NO3 − V f and its relationship with concentration have also been made by Mulholland et al. (2008), as part of a cross-site comparison of stable isotope additions (LINX2). Recall that K c was derived from a first-order uptake conceptualization, which assumes independence from concentration. Hence, our results (together with the above referenced studies) provide evidence of ‘nutrient saturation’. It suggests that higher-order models might be warranted when scaling NO3 − uptake across watersheds that are subject to increased nitrogen loading. The use of such models could especially be important when assessing the effects of land-use change over time.
In terms of extending our findings to larger scales, we should generalize our results with some caution. The general trend of the NO3 − uptake patterns we observed was most likely influenced by specific aspects of the study stream. These include the high NO3 − loading in headwaters only, a slight downstream increase in hyporheic exchange and an apparent downstream increase in biologic uptake capacity. What our results do illustrate is that variation in stream size, concentration and biological uptake capacity exert strong control on NO3 − uptake. Scaling from reach-level studies to river networks requires accounting for these factors, highlighting the need for empirical observations.
Conclusions
Our results showed distinct longitudinal uptake patterns for NO3 − and NH4 + and important differences in how these uptake patterns were affected by concentration and biological uptake capacity. This study illustrates that a common practice in simulation modeling studies, in which uptake is scaled across stream networks using ‘static’ uptake metric relationships, needs further refinement because of the inherent dependence on both concentration and biological uptake capacity. This poses a difficult question: How do concentration and biological uptake capacity vary across space and time? Also, how do they change as a result of suburbanization and other land-use change? And what is the effect on watershed nitrogen export? Given the pervasiveness of suburban sprawl and its detrimental effect on downstream waters, there is an urgent need for detailed combined field-modeling studies that address ecological and hydrological aspects of nutrient cycling within a geographic framework, using a longitudinal or stream network approach.
References
Alexander RB, Smith RA, Schwarz GE (2000) Effect of stream channel size on the delivery of nitrogen to the Gulf of Mexico. Nature 403:758–761
Alexander RB, Boyer EW, Smith RA, Schwarz GE, Moore RB (2007) The role of headwater streams in downstream water quality. J Am Water Resour Assoc 43:41–59
Bencala KE, Walters RA (1983) Simulation of solute transport in a mountain pool-and-riffle stream: a transient storage model. Water Resour Res 19:718–724
Bernhardt ES, Likens GE (2002) Dissolved organic carbon enrichment alters nitrogen dynamics in a forest stream. Ecology 83:1689–1700
Bernhardt ES, Hall RO, Likens GE (2002) Whole-system estimates of nitrification and nitrate uptake in streams of the Hubbard Brook Experimental Forest. Ecosystems 5:419–430
Boulton AJ, Findlay S, Marmonier P, Stanley EH, Valett HM (1998) The functional significance of the hyporheic zone in streams and rivers. Annu Rev Ecol Syst 29:59–81
Claessens L, Tague CL (2009) A transport-based method for estimating in-stream nitrogen uptake at ambient concentration from nutrient addition experiments. Limnol Oceanogr Methods (in press)
Claessens L, Tague CL, Groffman PM, Melack JM (2009a) Longitudinal and seasonal variation of stream N uptake in an urbanizing watershed: effect of organic matter, stream size, transient storage and debris dams. Biogeochemistry. doi:10.1007/s10533-009-9375-z
Claessens L, Tague CL, Band LE, Groffman PM, Kenworthy ST (2009b) Hydro-ecological linkages in urbanizing watersheds: an empirical assessment of in-stream nitrate loss and evidence of saturation kinetics. J Geophys Res Biogeosci. doi:10.1029/2009JG001017
Dodds WK, Lopez AJ, Bowden WB, Gregory S, Grimm NB, Hamilton SK, Hershey AE, Martí E, McDowell WH, Meyer JL, Morrall D, Mulholland PJ, Peterson BJ, Tank JT, Valett HM, Webster JR, Wollheim W (2002) N uptake as a function of concentration in streams. J N Am Benthol Soc 21:206–220
Elwood JW, Newbold JD, O’Neill RV, VanWinkle W (1983) Resource spiraling: an operational paradigm for analyzing lotic ecosystems. In: Fontaine TD, Bartell SM (eds) Dynamics of lotic ecosystems. Ann Arbor Science, Ann Arbor, MI, pp 3–27
Ensign SH, Doyle MW (2006) Nutrient spiraling in streams and river networks. J Geophys Res Biogeosci 111:G04009
Groffman PM, Law NL, Belt KT, Band LE, Fisher GT (2004) Nitrogen fluxes in urban watershed ecosystems. Ecosystems 7:393–403
Hall RO, Bernhardt ES, Likens GE (2002) Relating nutrient uptake with transient storage in forested mountain streams. Limnol Oceanogr 47:255–265
Mulholland P, Hall R, Hamilton S, Peterson B, Tank J, Ashkenas L, Cooper L, Dahm C, Dodds W, Findlay S, Gregory S, Grimm N, Johnson S, McDowell W, Meyer J, Poole G, Valett M, Webster J et al (2008) Stream denitrification across biomes and its response to anthropogenic nitrate loading. Nature 458:202–205
Newbold JD, Elwood JW, O’Neill RV, Van Winkle W (1981) Measuring nutrient spiraling in streams. Can J Fish Aquat Sci 38:860–863
Peterson BJ, Wollheim WM, Mulholland PJ, Webster JR, Meyer JL, Tank JL, Martí E, Bowden WB, Valett HM, Hershey AE, McDowell WH, Dodds WK, Hamilton SK, Gregory S, Morrall DD (2001) Control of nitrogen export from watersheds by headwater streams. Science 292:86–90
Runkel RL (1998) One dimensional transport with inflow and storage (OTIS): a solute transport model for streams and rivers. US Geological Survey Water-Resources Investigation Report 98-4018, p 73
Seitzinger SP, Styles RV, Boyer EW, Alexander RB, Billen G, Howarth RW, Mayer B, van Breemen N (2002) Nitrogen retention in rivers: model development and application to watersheds in the northeastern U.S.A. Biogeochemistry 57/58:199–237
Runkel RL (2002) A new metric for determining the importance of transient storage. J N Am Benthol Soc 21:529–543
Slavik K, Peterson BJ, Deegan LA, Bowden WB, Hershey AE, Hobbie JE (2004) Long-term responses of the Kuparuk River ecosystem to phosphorus fertilization. Ecology 85:939–954
Smith RA, Schwarz GE, Alexander RB (1997) Regional interpretation of water-quality monitoring data. Water Resour Res 33:2781–2798
Stream Solute Workshop (1990) Concepts and methods for assessing solute dynamics in stream ecosystems. J N Am Benthol Soc 9:95–119
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE (1980) River continuum concept. Can J Fish Aquat Sci 37:130–137
Webster JR, Ehrman TP (1996) Solute dynamics. In: Hauer FR, Lamberti GA (eds) Methods in stream ecology. Academic Press, San Diego, pp 145–160
Webster JR, Patten BC (1979) Effects of watershed perturbation on stream potassium and calcium dynamics. Ecol Monogr 49:51–72
Wollheim WM, Peterson BJ, Deegan LA, Hobbie JE, Hooker B, Bowden WB, Edwardson KJ, Arscott DB, Hershey AE (2001) Influence of stream size on ammonium and suspended particulate nitrogen processing. Limnol Oceanogr 46:1–13
Wollheim WM, Vörösmarty CJ, Peterson BJ, Seitzinger SP, Hopkinson CS (2006) Relationship between river size and nutrient removal. Geophys Res Lett 33:L06410. doi:10.1029/2006GL025845
Acknowledgments
We thank BES-LTER and Cary Institute of Ecosystems Studies for help with field experiments and laboratory analysis; UMD Palmer Lab for use of field equipment; and MBL Ecosystems Center for use of ion chromatograph. Comments by an anonymous reviewer helped to improve this manuscript. Funding for this research was provided by: NASA Earth System Science Fellowship; NSF Doctoral Dissertation Research Improvement grant (No. 0302703); NSF Long Term Ecological Research project (DEB-0423476); and San Diego State University Shared Visions grant.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Claessens, L., Tague, C.L., Groffman, P.M. et al. Longitudinal assessment of the effect of concentration on stream N uptake rates in an urbanizing watershed. Biogeochemistry 98, 63–74 (2010). https://doi.org/10.1007/s10533-009-9376-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-009-9376-y