
Abstract
In the face of the ongoing biodiversity crisis and limited conservation funding, surrogate approaches have become a valuable tool to represent biodiversity. Management surrogates are those that indirectly benefit an ecological system or species by representing the management requirements of co-occurring biodiversity. Recent findings highlight the cost-effective potential of surrogate species in managing threatened species, however, evaluating higher levels of biodiversity as management surrogates remains unexplored. Here, we sought to maximize conservation outcomes for threatened species and threatened ecological communities (TECs) by prioritizing management based on overlapping distributions, threats, and costs. We describe a prioritization framework for identifying TECs that could serve as cost-effective surrogates, and compare it with prioritizing threatened species only or both species and TECs. We show that when the objective is to maximize benefits for threatened species, a community approach performs poorly due to limited geographic overlap and high costs, while prioritizing species returned 7.5 times more benefits delivered to species under the same budget. Yet, if the objective is to maximize benefits across species and TECs simultaneously, a combined approach including both as surrogates delivers the greatest benefit for the same costs as a species-only approach. Range sizes and taxonomic groups significantly influenced the priority list, with threatened invertebrates and TECs of smaller ranges more likely to be selected as surrogates. Overall, this study emphasizes the importance of incorporating accurate data on factors such as threats and costs for identifying effective management surrogates, and highlights the potential benefits of prioritizing across multiple biodiversity features.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In ecological theory, surrogacy is the extent to which an ecological process or feature (e.g. species, ecosystem or abiotic factor—a surrogate) effectively represents another aspect of an ecological system (Rodrigues and Brooks 2007; Hunter et al. 2016). With ongoing declines of species, degradation of ecosystems (Ceballos et al. 2015), and limited funding available for conservation (Waldron et al. 2017), it is imperative to prioritize management so that it efficiently achieves conservation outcomes (Rodrigues and Brooks 2007; Wilson et al. 2009b). Deciding how and where to allocate resources across multiple species and ecosystems is a challenging task. Therefore, the use of surrogates can be a tool for maximizing benefits for biodiversity when it is unfeasible to manage all features of conservation concern (Wiens et al. 2008; Hunter et al. 2016). Surrogates can be used for different purposes, either for providing information about an aspect of an ecological system by measuring one component of an ecosystem to represent another (indicator surrogates; Lindenmayer and Likens 2011; Hunter et al. 2016) or by representing an aspect of that system that is the main focus of management (management surrogates; Hunter et al. 2016). Management surrogates can facilitate conservation decisions for a particular ecological system, where the primary goal is to manage an entity to achieve a broader benefit beyond the surrogate itself (Wilson et al. 2009a; Hunter et al. 2016).
To date, most of the surrogacy literature has centered around the concept of representation, by using different taxonomic groups, environmental units, functional groups, land classes or habitat types to represent target species as the focus of conservation (e.g., Lombard et al. 2003; Carmel and Stoller-Cavari 2006; Grantham et al. 2010; Lindenmayer et al. 2014; Ware et al. 2018; Falconer and Ford 2020). The term umbrella is commonly used to describe species that act as surrogates because they have broad distributions and habitat requirements, therefore, protecting a sufficiently large area of habitat for these species should protect the species that share that distribution (Noss 1990; Caro 2003). However, umbrella species can also serve as management surrogates, when management actions taken for these species benefit many co-occurring species facing similar threats (Roberge and Angelstam 2004; Hunter and Gibbs 2006). A study by Ward et al. (2020) used the concept of umbrella species to develop a prioritization framework for choosing species that would benefit the largest number of other co-occurring species with similar management requirements (i.e., shared threats), which increased management efficiencies and was more cost-effective than the approach used by the Australian Federal Government.
Management surrogates can differ by the type of biodiversity feature used: species-level (e.g., fine-filters) and ecosystems or ecological communities (e.g., coarse-filters) (Moilanen et al. 2009). Bioregions and ecoregions have been widely applied as landscape-level surrogates in conservation planning to guide reserve design (e.g., Chauvenet et al. 2020; Olson and Dinerstein 2002; Wilson et al. 2007), and represent biodiversity at the species and genetic level (Noss 1987). At a finer geographic scale, ecosystem or communities can serve as surrogates by representing co-occurring species assemblages (Noss 1996; Wiens et al. 2008). However, we argue that they could also be used as a management tool to prioritize actions benefiting associated biodiversity by managing ecosystems or communities, though this approach remains underexplored. Some strengths of managing at the community- or ecosystem-level is the potential protection offered to lesser known species that might be overlooked under species-level management, the protection of ecological patterns and processes, and ensuring continued provision of ecosystem services (Ferrier et al. 2009; Nicholson et al. 2009). Here, we focus on ecological communities, an assemblage of species that co-occur in space and time (Begon et al. 2006). When formally described, communities can take many forms, such as mapped vegetation types, land-classes or abiotic environmental classifications (Ferrier et al. 2009).
Despite the large body of research exploring cost-effective approaches to prioritize threat management for protecting species (e.g., Joseph et al. 2009; Carwardine et al. 2012; Auerbach et al. 2014; Cattarino et al. 2015; Ward et al. 2020), there are no studies comparing whether species or ecological communities, or a combination of both, deliver the greatest biodiversity benefits as management surrogates at a continental scale. Importantly, recent literature suggests a need to shift towards integrative approaches to selecting surrogates (Lundberg and Arponen 2022), and efforts to assess how to combine coarse and fine filter approaches (i.e. ecosystems or land classes and species) have been made, yet still with a focus on species representation (e.g., Lombard et al. 2003; Di Minin and Moilanen 2014). To date, no study has answered the questions: Would using threatened ecological communities (TEC) as management surrogates provide benefits to threatened species? Can a structured prioritization framework help identify TECs that act as management surrogates to cost-effectively maximize biodiversity outcomes? Would a combination of species and ecological communities deliver greater benefits to both biodiversity features?
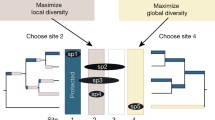
In this study, we aim to evaluate the potential benefits for biodiversity under two prioritization scenarios that consider threatened species and/or TECs as management surrogates (Fig. 1). The first objective is to determine if managing threats to TECs as surrogates would cost-effectively maximize the number of co-occurring threatened species that receive a conservation benefit (Scenario 1a), compared to only managing species (Scenario 1b; Fig. 1). A second objective is to maximize benefits to both species and TECs across the region, by comparing outcomes from prioritizing across species and TECs (Scenario 2a) versus species alone as surrogates (Scenario 2b; Fig. 1). This is important if the remit is to ensure that important biodiversity features are not overlooked. We expect that explicitly prioritizing both species and TECs will result in more threatened biodiversity being cost-effectively managed, given the potential overlaps in threats and distribution. Testing these two scenarios can elucidate how to efficiently prioritize management efforts by clearly identifying objectives (i.e. which features as the conservation target), and which surrogate strategies perform best. These hypotheses have not yet been tested, and the Australian continent offers an opportunity to achieve this.

Scenarios used to compare our prioritization framework. The objective of Scenario 1 was to maximize the benefit to all threatened species, whereas Scenario 2 considered benefits for both species and threatened ecological communities (TECs). All scenarios worked under the assumption that to maximize benefit, the surrogate (e.g., umbrella species) and the benefitting feature (e.g., any other threatened species) must overlap in threats (represented by fire icon) and geographic distributions (represented by overlapping grey circles). Green circles denote the benefits for threatened species, while orange circles represent the benefits for both threatened species and TECs. Scenario 1a: Only considered how TECs perform as surrogates to benefit all threatened species; Scenario 1b: Only considered how species perform as surrogates to benefit other threatened species; Scenario 2a: considered both species and TECS as surrogates to benefit all threatened species and TECs; Scenario 2b: Only considered species as surrogates to benefit all threatened species and TECs
Methods
Spatial data
We focused on Australian species and ecological communities listed as threatened (i.e., vulnerable, endangered or critically endangered) under the national environmental legislation. We used the Australian Government’s 0.1-degree gridded spatial data of 1393 threatened species (hereafter, species) for which there was data available on continental Australia, and 83 threatened ecological communities (hereafter, TECs) from the Species and Communities of National Environmental Significance Database, extracted on 15 December 2020. All data was processed using ArcGIS Pro 2.7.1 and R 3.6.3 (R Core Team 2020), using the packages ‘sf’ (Pebesma 2018) and ‘tidyverse’ (Wickham et al. 2019); clipped to the extent of the Australian landmass, and projected to the Geocentric Datum of Australia 1994. For both datasets, we limited our analysis to the ‘known’ and ‘likely to occur’ parts of the distribution (following Ives et al. 2016 and Renwick et al. 2017), to more accurately represent areas of suitable or preferred habitat, as ‘may-occur’ habitat represents mostly the outer envelope of their range. We excluded migratory and exclusively marine species (n = 134). We applied two raster masks at 100 × 100 m gridded cell resolution, which increases the accuracy of occurrences and reduces false positive errors, following the methods used in Ward et al. (2019). These masks refined the species and TECs distributions to areas of native vegetation, removing areas where native vegetation has been cleared or converted to other land uses, based on the National Vegetation Information System (NVIS V5.0; Appendix 1) (DAWE 2020a); and the Catchment Scale Land Use of Australia dataset (ABARES 2020). For the TECs distribution layer we applied both the NVIS and land use masks, but retained only natural land-use categories (Fig. 2, Step 1a). For the species distribution layer, we applied the land-use mask by creating a natural land-use layer for known and likely occurrences, plus a non-natural land-use layer for known occurrences, which were then combined into a single layer (Fig. 2, Step 1b). This was because species can still persist in disturbed and converted environments, unlike TECs. We resolved spatial inconsistencies and overlaps between 17 TECs to ensure that the ranges for each TEC were spatially exclusive (Appendix 1). When the overlap between two TECs was greater than 500 km2 or 50% of a TEC’s range, the area of overlap was assigned to the more narrowly distributed TEC, otherwise, it was left unmodified. We then intersected the species and TECs layers to obtain the area and proportion of overlap between the distribution of each species and community (Fig. 2, Step 1c).

Methodological steps to obtain required data for prioritization framework: (1) Spatial data preparation for a threatened ecological communities (TECs), b threatened species and c intersection between both layers across Australia; (2) threats matrix for all TECs and species; (3) costs of threat abatement strategies. All of these components are needed for each equation of the prioritization framework (Step 4). In Step 1 a, known and likely occurrences of TECs were masked to a broad vegetation class layer (National Vegetation Information System) and a natural land-use layer. In b, known and likely occurrences of species were masked to a natural land-use layer, which was combined with the known occurrences masked to a non-natural land-use layer, forming the final species distribution. In c, overlaps between species and TECs across Australia were obtained, and areas and proportion of overlap were calculated
Threats data
We extracted data on the threats to all TECs from government documents used to support the legislative threat status and appropriate management strategies (e.g., listing advices, conservation advices and recovery plans found in the Australian Government’s online Species Profile and Threats Database; DAWE 2020b). Data on threats impacting Australia’s species were obtained from the most up-to-date national-scale dataset (Ward et al. 2021). We converted these datasets into a matrix of threats by TEC and species, to identify the overlaps between the known and perceived threats to both (Fig. 2, Step 2). Following Murdoch et al. (2007), Evans et al. (2011), Auerbach et al. (2015) and Ward et al. (2020), we assumed that the spatial extent of each threat was equivalent to the geographic range of the species or TEC affected by that specific threat. Though we acknowledge this may overestimate the actual area threatened, and thus potentially overestimate management costs and/or the benefit delivered to co-occurring species or TECs.
Costs of threat abatement strategies
We used the most comprehensive costs models for threatened species management in Australia developed by Yong et al. (2023). These models include 18 threat abatement strategies with detailed estimates of the costs (including labor, transport, equipment and consumables) per square kilometer per year (AUD) for the actions needed to abate 40 underlying sub-category threats (see Fig. 2, Step 3 for example data). Thus, we based our framework on threat abatement strategies rather than specific threats, as multiple threats might be mitigated by a same management action. We assumed that TECs with the same threat would benefit from the same actions prescribed for species. To account for uncertainty in costs, we used three cost estimated as presented in Yong et al. (2023): minimum, best or ‘most common across Australia’ and maximum costs. For hydrology management, we converted the cost per instream structure to costs per km−2 year−1 (see Appendix 2 for details).
Prioritization framework and scenarios
We developed a prioritization framework to identify a priority list of surrogate features that, by managing their threats, would cost-effectively maximize the number of features that benefit due to overlaps in geographic distribution and threats (Fig. 2, Step 4). The prioritization framework followed a validated prioritization formula, adapted from Ward et al. (2020), by modifying the objectives, benefit and cost functions (Appendix 3). For each of the two objectives, we evaluated the cost-effectiveness of two different surrogate approaches for management (Fig. 1). When the objective was to maximize the benefit to all threatened species (Scenario 1), we compared a community approach (Scenario 1a), where only TECs were selected as potential management surrogates versus a species approach, where only threatened species were prioritized as surrogates (Scenario 1b, Fig. 1). When the objective was to maximize the benefit to both threatened species and TECs (Objective 2), we compared a combined approach where both species and TECS were prioritized as surrogates (Scenario 2a) versus a species only approach (Scenario 2b, same as above, Fig. 1). We did not evaluate a scenario using only TECs as surrogates for this second objective because we assumed that TECs do not co-occur, thus the results would be equivalent to the community approach for objective 1 (Scenario 1a). Further, to test how well our prioritization framework compared with selecting species or communities at random, we performed a scenario with 1000 iterations, where any species, TEC, or both could be included in the priority list regardless of its cost-effectiveness. Figure 2 illustrates a schematic workflow with all required components and datasets to run the prioritization algorithm.
We assumed that all threats to each surrogate feature are managed across its entire range; that strategies are totally effective in abating the relevant threat and provide a uniform benefit; and that the same threat abatement strategy would abate a co-occurring threat for any benefiting features. We did not account for interactions between actions or threats (see Auerbach et al. 2015), and did not consider whether areas are already receiving management. The formula used for each scenario are provided in supporting information (Appendix 3). Below, we describe the framework for the community approach (Scenario 1a), however, the same rules and assumptions apply to each scenario. Each scenario produces a priority list of surrogate features (TECs, species or both), benefiting features and additional features that, if managed, would result in the most cost-effective solution, following Ward et al. (2020). A benefiting feature is any species or TEC receiving management indirectly via the management of threats of a surrogate feature, due to overlaps in their distribution and management requirements (i.e., shared habitat and threats), regardless of the amount of overlap. Additional features are those that do not offer benefit to other features through their management, but are cost-effective to manage, thus, are included in the priority list until the budget is met.
Objective function
The objective is to maximize the total number of threatened species that could benefit simultaneously from implementing management actions that abate threats for TECs, within a budget constraint. This is a maximum gains decision problem as used by Murdoch et al. (2007), Wilson et al. (2007), Auerbach et al. (2014) and Ward et al. (2020). The prioritization algorithm produces a list of TECs in order of cost-effectiveness, based on the formula, Ea,
where Ba is the benefit of a certain TEC a to all other overlapping species j (i.e. benefiting species) (Eq. 2); Ca is the cost of implementing threat abatement strategies to manage all threats for TEC a (Eq. 3).
Benefit function
The benefit per TEC a can be calculated following Eq. 2, adapted from Ward et al. (2020). The benefit function accounts for the spatial overlap between TEC a and each species j, and the proportion of threats experienced by species j that would be abated if all the threats to TEC a were managed (i.e. common threats), so the benefit received by the benefiting species would be proportional to the amount of overlap. For example, if species j had three threats, two of which were shared by TEC a, and TEC a occurred in 50% of species j range, the benefit to j from a would be 0.33. Importantly, the remaining required management for species j (i.e., proportion of overlap or actions not included under TEC a), could be covered by the next most cost-effective TEC (or any following TEC prioritized) if they share geographic range and threats. Therefore, any surrogate TEC to be managed would have a base benefit of 1 (i.e., benefit to self), plus any extra benefit from co-occurring species. The benefit, Ba, of implementing all required threat abatement strategies for TEC a is:
where n is the number of species, Oaj is the proportion of overlap between the distributions of TEC a and species j (i.e., proportion of species j distribution captured within TEC a distribution); Wa is the weight, which is 1 for all TECs in this study, but could be varied depending on factors like threat status or value of ecosystem services; m is the total number of threat abatement strategies (across TEC a and species j); k is a specific threat abatement strategy; Tak and Tjk are binary variables that indicate if threat abatement strategy k is a strategy required for TEC a and species j, respectively.
Cost function
The total cost of managing all threats for TEC a, Ca, is represented by the cost of a specific threat abatement strategy, k, and the geographic range of TEC a:
where pk is the cost (AUD km−2 year −1) for implementing a threat abatement strategy k; Ga is the area over which the strategy is required for TEC a (represented by the spatial distribution of TEC a, in km2); s is the total number of threat abatement strategies required by the surrogate TEC.
Budget constraint
A budget constraint was used to determine which species or TECs could be cost-effective management surrogates under each scenario. We used AU$122 M year−1, based on the estimate of how much the Australian government and state governments invested on targeted threatened species recovery in 2018–2019 (Wintle et al. 2019). We accounted for a 3% inflation rate over a 2-financial-year period, which provided a yearly budget of $125,660,000.
Prioritization algorithm
The solution to this prioritization problem was determined using an iterative greedy algorithm that selected a priority list of management surrogate features in order of cost-effectiveness, which aimed to benefit the largest number of threatened species and/or TECs (depending on the objective) for any given budget. We calculated the return-on-investment based on the benefit gained per dollar spent as the budget increased. The iterative searching for the next most cost-effective surrogate feature stopped when the budget was met.
where Ba(z) is the benefit of managing all TECs that are in vector z, which represents the list of surrogate TECs prioritized based on cost-effectiveness (Eq. 1); h is the total number of TECs; Ca is the cost of implementing all threat abatement strategies for surrogate TEC a, subject to (s.t.) a budget.
Importantly, not all species and TECs can be identified as cost-effective surrogates and fit within the budget. For example, some species or TECs do not overlap in their distributions with any other species or community, or do not share any threats. Further, some species or TECs might provide high benefits but be widely distributed and face several threats, presenting high management costs and low cost-effectiveness. Once a feature is prioritized, it is removed from the iteration process so its benefits are not double counted. We evaluated the performance of each scenario by comparing the overall benefit, number of surrogates, additional and benefiting features obtained under each surrogate approach. The analysis was performed in R 3.6.3 (R Core Team 2020).
Generalized linear model
We modeled the likelihood of a species or TEC to be selected as a priority feature (i.e. surrogate or additional) for the combined approach (Scenario 2a), to determine which factors influenced selection. We fitted a generalized linear model with a binomial error distribution and a logit link function. Cost was excluded as an explanatory variable because it was confounded with the response variable, and Variance Inflation Factor (VIF) was used to test collinearity between the other covariates. The model included geographic range of species and TEC, taxonomic group, and their interaction, to assess whether priority features are influenced by these variables. We compared five models: with and without the interaction between the predictors, each predictor separately, and a null model (no variables and an intercept at 1). We selected the model with the lowest Akaike Information Criterion (AIC) value as the best-fitting model (Burnham and Anderson 2002).
Results
Maximizing benefit for threatened species
Under the assumed budget of $125 million year−1, prioritizing species alone (Scenario 1b) performed better, maximizing benefits for a larger number of threatened species than if threatened ecological communities were used as management surrogates (Fig. 3a, Table 1). By focusing on TECs (Scenario 1a), we found that 22 TECs (27%) across Australia could convey benefit to other species and act as cost-effective surrogates, and 19 TECs (23%) were included to the priority list as additional features until the budget was met (Table 1). This means that a community approach benefits considerably fewer species (n = 13 or 1% of species) relative to a species-only approach where 30% of all species would receive some benefit (n = 98 benefiting species (7% of species), plus 56 surrogate and 264 additional species) (Table 1, Fig. 3a). Our results also indicate that a species-only approach delivers higher total (i.e. cumulative) benefits for threatened species per dollar invested relative to a community approach under any given budget (Fig. 3b, Table 1). Both scenarios, however, performed overwhelmingly better than the random scenario, where species and communities were selected randomly over 1000 iterations. The maximum benefit obtained for the species and community scenarios were 230 and 15, respectively, with a median number of seven surrogate species for the species approach and 0 for the community approach, as the first randomly selected feature exceeded the budget. Priority species and TECs selected when using lower and upper costs estimates followed a similar pattern to the best estimate costs, with higher overall benefits, cost-effectiveness and more surrogate features selected under the lower cost scenario, and the upper cost scenario presenting the poorest performance (Appendix 4, Fig. a1 and a2).

a Number of surrogate features, benefiting species and additional species or threatened ecological communities (TECs) included in the solution under each management approach for Scenario 1. ‘Surrogate features’ are TECs (from a potential n = 83 in Scenario 1a) or species (from a potential n = 1425 in Scenario 1b) that were selected to provide cost-effective management to other co-occurring threatened species with similar threats. In Scenario 1, ‘Benefiting features’ are species that receive benefit (i.e., protection or management) from the threat abatement strategies implemented for the surrogate features. ‘Additional features’ are species or TECs added to the list in order of cost-effectiveness until the budget is reached, however they do not contribute to another species’ benefit. b Shows the number of threatened species that would receive benefit per additional dollar invested under each surrogate approach. All costs presented are the best cost estimates of threat abatement strategies, and cumulative costs are presented in millions of Australian dollars (AUD) per year
Maximizing benefit for species and ecological communities
When the aim is to prioritize surrogate features that maximize benefits for both threatened species and TECs, the two scenarios produced very similar results (Table 1, Fig. 4). Under the budget constraint of $125 million year−1, both species and combined approach (i.e., prioritizing across species and TECs as management surrogates) presented almost the same overall cost-effectiveness (E = 0.035 vs 0.036, respectively), and the same number of benefiting features (n = 45), yet differed by one more surrogate and 11 extra additional features under a combined approach (Fig. 4a, Table 1). Both approaches presented a similar trend in their cumulative benefits, however, the combined approach showed slightly higher benefits for similar costs relative to the species-only approach (B = 396 for a combined-approach versus B = 386 for species-only; Fig. 4b, Table 1). As per Scenario 1, the random scenario showed an overwhelmingly poor performance compared with both approaches. The maximum benefit obtained after 1,000 iterations were 32 and 31 for the combined and species-only approach, respectively, with a median number of surrogate features of one selected per iteration in both scenarios. Similar to Scenario 1, cumulative benefits under lower and upper costs estimates followed similar patterns to the best-estimate costs, with higher number of features selected under lower costs (Appendix 4, Fig. a3 and a4).

a Number of surrogate features, benefiting species and additional species or threatened ecological communities (TECs) included in the solution under each management approach for Scenario 2. ‘Surrogate features’ are TECs (from a potential n = 83 in Scenario 2a) or species (from a potential n = 1425 in Scenarios 2a and 2b) that were selected to provide cost-effective management to other co-occurring threatened species or TECs with similar threats. In Scenario 1, ‘Benefiting features’ are species and TECs that receive benefit (i.e., protection or management) from the threat abatement strategies implemented for the surrogate features. ‘Additional features’ are species or TECs added to the list in order of cost-effectiveness until the budget is reached, however they do not contribute to another species’ benefit. b Shows the number of threatened species and TECs that would receive benefit per additional dollar invested. Cumulative benefits increase when adding the next most cost-effective species (green line) or the next most cost-effective species and TEC (orange line). All costs presented are the best cost estimates of threat abatement strategies, and cumulative costs are presented in millions of Australian dollars (AUD) per year
Characteristics of priority species and TECs
Plants were the most common taxon selected both as surrogates and additional features for the species-only approaches (Scenarios 1b and 2b), as 79% of species prioritized were plants, on average across scenarios. This was followed by invertebrates (7%), and mammals (4%). Moreover, the average number of threat abatement strategies that would be managed per priority species across scenarios is 1.6, ranging from one to three strategies. Yet, for ecological communities (Scenario 1a) these range from three to ten threat abatement strategies, with an average of six strategies per TEC, explaining the higher costs of management for TECs. The best-fitted model for the combined approach was the one including the interaction term between taxonomic group and geographic range (see Appendix 5 for AIC values for all models). We found that both variables had a strong effect on the species and TECs prioritized, both being important predictors to describe which types of features were more likely to be selected (Scenario 2a). This relationship was largely driven by plants, invertebrates, and TECs (considered as another taxonomic group for this purpose), being more likely to be selected relative to other taxonomic groups. The range of all threatened species and TECs used for the analyses varied widely from very narrowly distributed (< 1 km2) to widespread species and TECs (> 1000 km2). However, in the combined approach, the prioritization algorithm selected priority features with smaller ranges than those not prioritized (Fig. 5), pattern strongest for TECs and invertebrates (Appendix 5).

Natural log of the geographic range (km2) of threatened species per taxonomic group, categorized into those that were prioritized (orange boxes) and those not prioritized (green boxes) under the combined-approach scenario (Scenario 2a). Threatened ecological communities (TECs) were considered as an additional taxonomic group for this purpose
Discussion
With the current rates of biodiversity loss (Ceballos et al. 2015), and scarce resources allocated towards conservation, there is a need to implement structured and transparent prioritization frameworks that efficiently maximize benefits to biodiversity (Carwardine et al. 2008; Joseph et al. 2009). Here, we show that we can maximize the benefits to threatened biodiversity by prioritizing the management of surrogates that represent cost-effective management of threatened species and threatened ecological communities for any given budget. While using Australia as a case study, the novelty of this study centers on assessing whether managing higher order biodiversity features, such as TECs as surrogates, would be a cost-effective strategy for threatened species management. Further, we explored what constitutes a suitable management surrogate, and whether combining approaches could maximize biodiversity outcomes.
Our findings suggest that, when the objective is to benefit threatened species (Scenario 1; Fig. 1), resources should be invested in species management, since TECs in Australia offered lower benefits to species when treated as surrogates due to their high management costs and low co-occurrence (Fig. 3). Yet, if the aim is to conserve both TECs alongside species (Scenario 2; Fig. 1), then using both features as surrogates would deliver the greatest overall benefits (albeit marginal in this case study) under the same budget (Fig. 4), while ensuring that both features are explicitly represented. Our study further contributes to the management surrogacy literature by providing an integrated framework to assess surrogates that may be useful for conservation management, and should be tested in other contexts. Incorporating ecosystems and ecological communities in conservation planning and prioritization decisions is essential, as reflected in international commitments such as the Convention on Biological Diversity post-2020 Global Biodiversity Framework (Nicholson et al. 2021; CBD 2022), and legislative mandates in many countries, including Australia (DAWE 2020b). However, government policies still tend focus predominantly on species (e.g. Australian Threatened Species Action Plan; DCCEEW 2022).
The poor performance of the community approach is likely driven by three factors. First, ecological communities across Australia face several complex threats, hence their recovery is resource intensive. Restoring habitat is one of the primary methods to manage ecological communities and ecosystems (Ferrer-Paris et al. 2019), being critical for preventing further degradation and loss. Habitat restoration was the most expensive strategy in our case study, and has been found to be the most expensive and least cost-effective strategy in other studies (Martin et al. 2018). Yet, it is required by 95% of TECs, explaining why the budget was quickly reached under the community approach. Second, TECs have faced long-term declines due to clearing and degradation, occurring in mostly fragmented and small patches (Tulloch et al. 2016). This resulted in small geographic overlaps with species, relative to large co-occurring distributions among species. Given the level of fragmentation and lack of connectivity of TECs in Australia, they do not adhere to the widespread distributions proposed by the umbrella concept (Wilcox 1984). This might be a common factor of threatened ecosystems listed due to a decline or restricted distribution (Bland et al. 2017), yet, this hypothesis needs to be further examined in other case studies. Third, there are only 83 TECs in Australia compared with 1393 threatened species used in this analysis. This large difference between the number of potential surrogates per feature could also explain why species perform overwhelmingly better.
Despite the calls to move from a species-focused approach towards one that focuses management on ecological communities or ecosystems (Noss 1996; Keith 2015), we show that for the Australian context, using threatened ecological communities as management surrogates may not be a beneficial strategy on its own. Yet, our analyses helped identify factors that influence suitable management surrogates, including those that are both cost-effective and provide significant benefits to other biodiversity features. Based on our data, taxonomic groups and geographic distribution significantly influenced our prioritization solution (Fig. 5). Specifically, both TECs and invertebrate species with smaller geographic ranges were more likely to be selected as management surrogate because of the lower management costs. This information can be valuable for future conservation planning and prioritization efforts in the Australian context, and should be tested in a wide range of contexts to explore their generality.
Management implications
Despite the large body of literature on the use of surrogates, such as indicators, umbrellas, or flagship species (Roberge and Angelstam 2004; Wiens et al. 2008; Caro 2010; Lundberg and Arponen 2022), and habitat-based surrogates (e.g., Lombard et al. 2003; Di Minin and Moilanen 2014; Lindenmayer et al. 2014), no study has compared the effectiveness of threatened ecological communities as management surrogates, and how to prioritize across both species and ecosystems to improve conservation management. We highlight the importance of implementing spatial prioritization approaches that account for threat abatement actions and costs to identify management surrogates, as suggested by Cattarino et al. (2015), Ward et al. (2020) and Salgado-Rojas et al. (2023), rather than solely accounting for spatial representation. The latter may not fully address all the necessary biodiversity requirements to effectively mitigate threats and ensure their persistence. Our findings are supported by Di Minin and Moilanen (2014), who concluded that habitat types are a critical part of successful surrogate strategies to account for poorly known taxonomic groups, aligning with our combined approach between species-level and above species-level management. Similarly, Lombard et al. (2003) found that land classes (i.e., broad habitat units derived from vegetation layers and environmental variables) performed well as surrogates for known plant distributions, but specific-species that fall through this coarse filter (e.g. fauna) would benefit from targeting both land classes and species data simultaneously.
We suggest that the framework presented here can provide information on the type of cost-effective surrogates and inform finer-scale decisions about which management strategies to implement. When data on threats and distribution of threatened species and ecological communities (or ecosystems) are available, practitioners and decision-makers can follow this process to determine optimal conservation strategies. Yet, in situations where there is limited capacity or data availability, incorporating threatened ecological communities or ecosystems alongside species in management and planning may lead to improved conservation outcomes and ensure comprehensive management at both levels of biodiversity.
Assumptions and limitations
Our study relied on two main assumptions. First, we assumed that threats to a species or community occurred across their entire distribution, which might have influenced the selection of management surrogates with smaller geographic distributions. This may lead to overestimation of the extent of overlapping threats between features, which would increase their cost-effectiveness. Ideally, the specific location and spatial variation in the intensity of threats affecting species and TECs should be accounted for (Wilson et al. 2009b; Evans et al. 2011), potential through integrating factors like land form, soil type, patch area, and human population density (Maron et al. 2013). Threats can also interact when their effects co-occur spatially and temporally (Côté et al. 2016). However, these data are rarely, if ever, available at the continental scale, and were unavailable at the time of analysis. Given the importance of the quality of the spatial, threats and cost data in our methodology, it is noteworthy to recognize that the effectiveness of our prioritization framework is contingent upon the availability and reliability of such data.
Second, we assumed that the same threat abatement strategies used for managing species (Yong et al. 2023) would be effective in managing TECs facing the same threat. However, ecological communities might have specific requirements that cannot be addressed solely through species-focused management actions, such as changes in composition, structure and disruption of ecological processes (Nicholson et al. 2009). Further, we did not account for species interactions and dependencies within community assemblages, which are key properties of ecological communities (Keith 2009). Addressing these interactions, and properly incorporating them into our benefit function would provide a more realistic and positive outcome of TEC’s performance as management surrogates. Nevertheless, these assumptions about threats and actions are commonly required by large-scale conservation analyses where detailed, site-based information is lacking (e.g. Wilson et al. 2007; Auerbach et al. 2015; Cattarino et al. 2015; Ward et al. 2020). Explicitly addressing these assumptions would likely improve the effectiveness and reliability of decision-making processes (Maron et al. 2013).
While our prioritization framework aims to maximize conservation benefits received across threatened species and ecological communities, it is important to note that our methods may produce disparities in the degree of benefit conferred to certain groups of threatened species. For instance, some species relying on both terrestrial and marine habitats may require additional management to fully address marine-related threats and ensure their persistence, which was outside the scope of this study. Other prioritization approaches, such as those presented in Cattarino et al. (2015) and Salgado-Rojas et al. (2023), have defined their targets as species persistence in their conservation allocation problem. These studies developed complementarity-based multi-action planning approaches to identify an optimal combination of management actions and their spatial allocation within a site that achieves the conservation target (either at the minimum cost, or by maximizing recovery across all features–subject to a budget). Approaches such as these might address the caveats on potential uneven benefits in our study by incorporating species-specific persistence targets and associated management requirements into the prioritization process.
Suggestions for future research
The findings presented here may be influenced by the specifics of the Australian context, the methods used to define and list threatened ecological communities, as well as their limited geographic overlaps with species. Therefore, it is important to test our methods and scenarios in a wide range of other contexts, particularly countries with comprehensive lists of threatened ecosystems (e.g., Colombia, South Africa), or under an experimental approach with independent datasets, to evaluate the generalizability of our findings. Additionally, future studies could also investigate the use of other levels of biodiversity as management surrogates, such as functional groups or biomes (as per Keith et al. 2022), as well as different threatening processes and socio-economic factors that might afftect the costs of implementing management. Categories of management costs, such as labor, consumables and travel to sites (Iacona et al. 2018; Yong et al. 2023), might differ considerably across different regions and countries, and should be accounted for when implementing conservation strategies.
Future prioritization exercises could add complexity and maximize management efficiencies by incorporating complementarity of actions across sites (Chadès et al. 2014, 2019; Cattarino et al. 2015), interactions between actions (Auerbach et al. 2015), and/or interactions and co-occurrence between threats if data exist (Geary et al. 2019; Legge et al. 2019). A sensible next step in this research involves incorporating the recently developed spatial management costs for Australia (Yong et al. 2023). Moreover, we suggest that integrating approaches by managing across species and ecosystems could have conservation benefits for lesser-known species (Noss 1987; Ferrier et al. 2009), including those that are yet to be described, or with undetermined threat status. Managing above the species-level also offers the potential to protect ecological processes and ecosystem services (Nicholson et al. 2009), which were not captured in our analysis. Future studies could examine these considerations by modifying the benefit function. TECs could be assigned greater weights compared to species, considering their potential to conserve lesser-known species. Additionally, if the objective, such as restoring ecosystem function, is to be considered, the weighting variable could account for the number of ecosystem services and ecological functions provided by each TEC or species (Akçakaya et al. 2020). However, this is only possible if data on such processes and services exists, which was unavailable for our case study.
This study compared the cost-effectiveness of managing threatened species, threatened ecological communities, or a combination of both to protect threatened biodiversity, addressing a crucial knowledge gap on which surrogate strategies deliver greater biodiversity benefits. Our findings demonstrate that allocating conservation resources across multiple levels of biodiversity, incorporating both threatened species and TECs, is slightly more efficient than using a species-only or community-only approach. The prioritization framework presented in this study highlights the importance of using accurate data on threats and management costs to improve efficiency (Bode et al. 2008; Carwardine et al. 2012). Conservation practitioners and managers can adapt this framework to their specific context, integrating different conservation features and adjusting the spatial scale of prioritization as necessary. Transparent and strategic spatial conservation prioritization analyses, such as the one presented here, can help optimize the use of scarce resources for protecting threatened biodiversity.
Data avilability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Akçakaya HR, Rodrigues ASL, Keith DA, Milner-Gulland EJ, Sanderson EW, Hedges S, Mallon DP, Grace MK, Long B, Meijaard E, Stephenson PJ (2020) Assessing ecological function in the context of species recovery. Conserv Biol 34:561–571. https://doi.org/10.1111/cobi.13425
Auerbach NA, Tulloch AIT, Possingham HP (2014) Informed actions: where to cost effectively manage multiple threats to species to maximize return on investment. Ecol Appl 24:1357–1373. https://doi.org/10.1890/13-0711.1
Auerbach NA, Wilson KA, Tulloch AIT, Rhodes JR, Hanson JO, Possingham HP (2015) Effects of threat management interactions on conservation priorities. Conserv Biol 29:1626–1635. https://doi.org/10.1111/cobi.12551
Australian Bureau of Agricultural and Resource Economics and Sciences [ABARES]. (2020). Catchment scale land use of Australia – Update December 2020. https://www.agriculture.gov.au/abares/aclump/catchment-scale-land-use-of-australia-update-december-2020
Bland LM, Keith DA, Miller RM, Murray NJ, Rodríguez JP (2017) Guidelines for the application of IUCN Red List of ecosystems categories and criteria. IUCN, Gland
Begon M, Townsend CR, Harper JL (2006) Ecology: from individuals to ecosystems, 4th edn. Blackwell Publishing, Boston
Bode M, Wilson KA, Brooks TM, Turner WR, Mittermeier RA, McBride MF, Underwood EC, Possingham HP (2008) Cost-effective global conservation spending is robust to taxonomic group. Proc Natl Acad Sci USA 105:6498–6501. https://doi.org/10.1073/pnas.0710705105
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Carmel Y, Stoller-Cavari L (2006) Comparing environmental and biological surrogates for biodiversity at a local scale. Isr J Ecol Evol 52:11–27. https://doi.org/10.1560/IJEE.52.1.11
Caro TM (2003) Umbrella species: critique and lessons from East Africa. Anim Conserv 6(2):171–181. https://doi.org/10.1017/S1367943003003214
Caro TM (2010) Indicator umbrella keystone flagship and other surrogate species. In: Caro T, Girling S (eds) Conservation by proxy. Island Press, Washington
Carwardine J, O’Connor T, Legge S, Mackey B, Possingham HP, Martin TG (2012) Prioritizing threat management for biodiversity conservation. Conserv Lett 5:196–204. https://doi.org/10.1111/j.1755-263X.2012.00228.x
Carwardine J, Wilson KA, Watts M, Etter A, Klein CJ, Possingham HP (2008) Avoiding costly conservation mistakes: the importance of defining actions and costs in spatial priority settings. PLoS ONE 3:e2586. https://doi.org/10.1371/journal.pone.0002586
Cattarino L, Hermoso V, Carwardine J, Kennard MJ (2015) Multi-action planning for threat management: a novel approach for the spatial prioritization of conservation actions. PLoS ONE 10:1–18. https://doi.org/10.1371/journal.pone.0128027
Ceballos G, Ehrlich PR, Barnosky AD, García A, Pringle RM, Palmer TM (2015) Accelerated modern human–induced species losses: entering the sixth mass extinction. Sci Adv 1:1–5. https://doi.org/10.1126/sciadv.1400253
Chadès I, Nicol S, van Leeuwen S, Walters B, Firn J, Reeson A, Martin TG, Carwardine J (2014) Benefits of integrating complementarity into priority threat management. Conserv Biol 29:525–536. https://doi.org/10.1111/cobi.12413
Chadès I, Ponce-Reyes R, Nicol S, Pascal LV, Fletcher C, Cresswell I, Cardwardine J (2019) An integrated spatial prioritisation plan for the Saving our Species program. Report prepared for the NSW Office of Environment and Heritage. CSIRO, Brisbane
Chauvenet ALM, Watson JEM, Adams VM, Di Marco M, Venter O, Davis KJ, Mappin B, Klein CJ, Kuempel CD, Possingham HP (2020) To achieve big wins for terrestrial conservation, prioritize protection of ecoregions closest to meeting targets. One Earth 2:479–486. https://doi.org/10.1016/j.oneear.2020.04.013
Convention on Biological Diversity [CBD] (2022) Nations Adopt Four goals, 23 Targets for 2030 in Landmark UN Biodiversity Agreement: 1–9. Montreal. [WWW Document] https://www.cbd.int/article/cop15-cbd-press-release-final-19dec2022
Côté IM, Darling ES, Brown CJ (2016) Interactions among ecosystem stressors and their importance in conservation. Proc Royal Soc B: Biol Sci 283:20152592. https://doi.org/10.1098/rspb.2015.2592
Department of Climate Change, Energy, the Environment and Water [DCCEEW] (2022) Threatened Species Action Plan 2022–2032. Department of Climate Change, Energy, the Environment and Water, Canberra, September
Department of Agriculture, Water and the Environment [DAWE] (2020a) National Vegetation Information System (NVIS) data products V5.0. https://www.environment.gov.au/land/native-vegetation/national-vegetation-information-system/data-products
Department of Agriculture, Water and the Environment [DAWE] (2020b) Environment Protection Biodiversity and Conservation (EPBC) Act List of Threatened Ecological Communities. http://www.environment.gov.au/cgi-bin/sprat/public/publiclookupcommunities.pl
Di Minin E, Moilanen A (2014) Improving the surrogacy effectiveness of charismatic megafauna with well-surveyed taxonomic groups and habitat types. J Appl Ecol 51(2):281–288. https://doi.org/10.1111/1365-2664.12203
Evans MC, Watson JEM, Fuller RA, Venter O, Bennett SC, Marsack PR, Possingham HP (2011) The spatial distribution of threats to species in Australia. Bioscience 61:281–289. https://doi.org/10.1525/bio.2011.61.4.8
Falconer S, Ford AT (2020) Evaluating policy-relevant surrogate taxa for biodiversity conservation: a case study from British Columbia, Canada. Can J Zool 98:279–286. https://doi.org/10.1139/cjz-2019-0178
Ferrer-Paris JR, Zager I, Keith DA, Oliveira-Miranda MA, Rodríguez JP, Josse C, González-Gil M, Miller RM, Zambrana-Torrelio C, Barrow E (2019) An ecosystem risk assessment of temperate and tropical forests of the Americas with an outlook on future conservation strategies. Conserv Lett 12:1–10. https://doi.org/10.1111/conl.12623
Ferrier S, Faith DP, Arponen A, Drielsma M (2009) Community-level approaches to spatial conservation prioritization. In: Moilanen A, Wilson KA, Possingham HP (eds) Spatial conservation prioritization: quantitative methods and computational tools, 2nd edn. Oxford University Press, Oxford, pp 94–109
Geary WL, Tulloch AIT, Nimmo DG, Doherty TS, Ritchie EG (2019) Threat webs: reframing the co-occurrence and interactions of threats to biodiversity. J Appl Ecol 56:1992–1997. https://doi.org/10.1111/1365-2664.13427
Grantham HS, Pressey RL, Wells JA, Beattie AJ (2010) Effectiveness of biodiversity surrogates for conservation planning: different measures of effectiveness generate a kaleidoscope of variation. PLoS ONE 5(7):1–12. https://doi.org/10.1371/journal.pone.0011430
Hunter M, Westgate M, Barton P, Calhoun A, Pierson J, Tulloch A, Beger M, Branquinho C, Caro T, Gross J, Heino J, Lane P, Longo C, Martin K, Mcdowell WH, Mellin C, Salo H, Lindenmayer D (2016) Two roles for ecological surrogacy: indicator surrogates and management surrogates. Ecol Ind 63:121–125. https://doi.org/10.1016/j.ecolind.2015.11.049
Hunter ML, Gibbs JP (2006) Fundamentals of conservation biology, 3rd edn. Blackwell Publishing, Malden
Iacona GD, Sutherland WJ, Mappin B, Adams VM, Armsworth PR, Coleshaw T, Cook C, Craigie I, Dicks LV, Fitzsimons JA, McGowan J, Plumptre AJ, Polak T, Pullin AS, Ringma J, Rushworth I, Santangeli A, Stewart A, Tulloch A, Walsh JC, Possingham HP (2018) Standardized reporting of the costs of management interventions for biodiversity conservation. Conserv Biol 32(5):979–988. https://doi.org/10.1111/cobi.13195
Ives CD, Lentini PE, Threlfall CG, Ikin K, Shanahan DF, Garrard GE, Bekessy SA, Fuller RA, Mumaw L, Rayner L, Rowe R, Valentine LE, Kendal D (2016) Cities are hotspots for threatened species. Glob Ecol Biogeogr 25:117–126. https://doi.org/10.1111/geb.12404
Joseph LN, Maloney RF, Possingham HP (2009) Optimal allocation of resources among threatened species: a project prioritization protocol. Conserv Biol 23:328–338. https://doi.org/10.1111/j.1523-1739.2008.01124.x
Keith DA (2009) The interpretation, assessment and conservation of ecological communities. Ecol Manag Restor 10:S3–S15. https://doi.org/10.1111/j.1442-8903.2009.00453.x
Keith DA (2015) Assessing and managing risks to ecosystem biodiversity. Austral Ecol 40:337–346. https://doi.org/10.1111/aec.12249
Keith DA, Ferrer-Paris JR, Nicholson E, Bishop MJ, Polidoro BA, Ramirez-Llodra E, Tozer MG, Nel JL, Mac Nally R, Gregr EJ, Watermeyer KE (2022) A function-based typology for Earth’s ecosystems. Nature 610(7932):513–518. https://doi.org/10.1038/s41586-022-05318-4
Legge S, Smith JG, James A, Tuft KD, Webb T, Woinarski JCZ (2019) Interactions among threats affect conservation management outcomes: Livestock grazing removes the benefits of fire management for small mammals in Australian tropical savannas. Conserv Sci Pract 1:1–13. https://doi.org/10.1111/csp2.52
Lindenmayer DB, Likens GE (2011) Direct Measurement versus surrogate indicator species for evaluating environmental change and biodiversity loss. Ecosystems 14:47–59. https://doi.org/10.1007/s10021-010-9394-6
Lindenmayer DB, Barton PS, Lane PW, Westgate MJ, Mcburney L, Blair D, Gibbons P, Likens GE (2014) An empirical assessment and comparison of species-based and habitat-based surrogates: a case study of forest vertebrates and large old trees. PLoS ONE 9(2):1–10. https://doi.org/10.1371/journal.pone.0089807
Lombard AT, Cowling RM, Pressey RL, Rebelo AG (2003) Effectiveness of land classes as surrogates for species in conservation planning for the Cape Floristic Region. Biol Cons 112(1–2):45–62. https://doi.org/10.1016/S0006-3207(02)00422-6
Lundberg P, Arponen A (2022) An overview of reviews of conservation flagships: evaluating fundraising ability and surrogate power. Nat Conserv 49:153–188. https://doi.org/10.3897/natureconservation.49.81219
Maron M, Rhodes JR, Gibbons P (2013) Calculating the benefit of conservation actions. Conserv Lett 6:359–367. https://doi.org/10.1111/conl.12007
Martin TG, Kehoe L, Mantyka-Pringle C, Chades I, Wilson S, Bloom RG, Davis SK, Fisher R, Keith J, Mehl K, Diaz BP, Wayland ME, Wellicome TI, Zimmer KP, Smith PA (2018) Prioritizing recovery funding to maximize conservation of endangered species. Conserv Lett 11:1–9. https://doi.org/10.1111/conl.12604
Moilanen A, Wilson KA, Possingham HP (2009) Spatial conservation prioritization: quantitative methods and computational tools, 1st edn. Oxford University Press, Oxford
Murdoch W, Polasky S, Wilson KA, Possingham HP, Kareiva P, Shaw R (2007) Maximizing return on investment in conservation. Biol Cons 139:375–388. https://doi.org/10.1016/j.biocon.2007.07.011
Nicholson E, Keith DA, Wilcove DS (2009) Assessing the threat status of ecological communities. Conserv Biol 23:259–274. https://doi.org/10.1111/j.1523-1739.2008.01158.x
Nicholson E, Watermeyer KE, Rowland JA, Sato CF, Stevenson SL, Andrade A, Brooks TM, Burgess ND, Cheng S, Grantham HS, Hill SL, Keith DA, Maron M, Metzke D, Murray NJ, Nelson CR, Obura D, Plumptre A, Skowno AL, Watson JEM (2021) Scientific foundations for an ecosystem goal, milestones and indicators for the post-2020 global biodiversity framework. Nat Ecol Evol 5:1338–1349. https://doi.org/10.1038/s41559-021-01538-5
Noss RF (1987) From plant communities to landscapes in conservation inventories: a look at the nature conservancy (USA). Biol Cons 41:11–37. https://doi.org/10.1016/0006-3207(87)90045-0
Noss RF (1990) Indicators for monitoring biodiversity: a hierarchical approach. Conserv Biol 4:355–364. https://doi.org/10.1111/j.1523-1739.1990.tb00309.x
Noss RF (1996) Ecosystems as conservation targets. Trends Ecol Evol 11:351
Olson DM, Dinerstein E (2002) The global 200: priority ecoregions for global conservation. Ann Mo Bot Gard 89:199–224. https://doi.org/10.2307/3298564
Pebesma E (2018) Simple features for R: standardized support for spatial vector data. R Journal 10:439–446
R Core Team (2020) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [WWW Document]. https://www.r-project.org/
Renwick AR, Robinson CJ, Garnett ST, Leiper I, Possingham P, Carwardine J (2017) Mapping Indigenous land management for threatened species conservation: an Australian case-study. PLoS ONE 12:1–11. https://doi.org/10.1371/journal.pone.0173876
Roberge JM, Angelstam P (2004) Usefulness of the umbrella species concept as a conservation tool. Conserv Biol 18:76–85. https://doi.org/10.1111/j.1523-1739.2004.00450.x
Rodrigues ASL, Brooks TM (2007) Shortcuts for biodiversity conservation planning: the effectiveness of surrogates. Annu Rev Ecol Evol Syst 38:713–737. https://doi.org/10.1146/annurev.ecolsys.38.091206.095737
Salgado-Rojas J, Hermoso V, Álvarez-Miranda E (2023) Prioriactions: multi-action management planning in R. Methods Ecol Evol. https://doi.org/10.1111/2041-210X.14220
Tulloch AIT, Barnes MD, Ringma J, Fuller RA, Watson JEM (2016) Understanding the importance of small patches of habitat for conservation. J Appl Ecol 53:418–429. https://doi.org/10.1111/1365-2664.12547
Waldron A, Miller DC, Redding D, Mooers A, Kuhn TS, Nibbelink N, Roberts JT, Tobias JA, Gittleman JL (2017) Reductions in global biodiversity loss predicted from conservation spending. Nature 551:364–367. https://doi.org/10.1038/nature24295
Ward M, Simmonds JS, Reside AE, Watson JEM, Rhodes JR, Possingham HP, Trezise J, Fletcher R, File L, Taylor M (2019) Lots of loss with little scrutiny: the attrition of habitat critical for threatened species in Australia. Conserv Sci Pract 1:1–13. https://doi.org/10.1111/csp2.117
Ward M, Rhodes JR, Watson JEM, Lefevre J, Atkinson S, Possingham HP (2020) Use of surrogate species to cost-effectively prioritize conservation actions. Conserv Biol 34:600–610. https://doi.org/10.1111/cobi.13430
Ward M, Carwardine J, Yong C, Watson J, Silcock J, Taylor G, Lintermans M, Gillespie G, Garnett S, Woinarski J, Tingley R, Fensham R, Hoskin C, Hines H, Roberts J, Kennard M, Harvey M, Chapple D, Reside A (2021) A national-scale dataset for threats impacting Australia’s imperilled flora and fauna. Ecol Evol 00:1–13. https://doi.org/10.1002/ece3.7920
Ware C, Williams KJ, Harding J, Hawkins B, Harwood T, Manion G, Perkins GC, Ferrier S (2018) Improving biodiversity surrogates for conservation assessment: a test of methods and the value of targeted biological surveys. Divers Distrib 24(9):1333–1346
Wickham H, Averick M, Bryan J, Chang W, McGowan L, François R, Grolemund G, Hayes A, Henry L, Hester J, Kuhn M, Pedersen T, Miller E, Bache S, Müller K, Ooms J, Robinson D, Seidel D, Spinu V, Yutani H (2019) Welcome to the Tidyverse. J Open Source Softw 4:1686
Wiens JA, Hayward GD, Holthausen RS, Wisdom MJ (2008) Using surrogate species and groups for conservation planning and management. Bioscience 58:241–252. https://doi.org/10.1641/B580310
Wilcox BA (1984) In situ conservation of genetic resources: determinants of minimum area requirements. Conservation and Development, National Parks, pp 639–647. https://doi.org/10.13140/2.1.4879.2322
Wilson KA, Underwood EC, Morrison SA, Klausmeyer KR, Murdoch WW, Reyers B, Wardell-Johnson G, Marquet PA, Rundel PW, McBride MF, Pressey RL, Bode M, Hoekstra JM, Andelman S, Looker M, Rondinini C, Kareiva P, Shaw MR, Possingham HP (2007) Conserving biodiversity efficiently: what to do, where, and when. PLoS Biol 5:1850–1861. https://doi.org/10.1371/journal.pbio.0050223
Wilson KA, Cabeza M, Klein CJ (2009a) Fundamental concepts of spatial conservation prioritization. Spatial conservation prioritization: quantitative methods and computational tools. Oxford University Press, Oxford, pp 16–27
Wilson KA, Carwardine J, Possingham HP (2009b) Setting conservation priorities. Ann NY Acad Sci 1162:237–264. https://doi.org/10.1111/j.1749-6632.2009.04149.x
Wintle BA, Cadenhead NCR, Morgain RA, Legge SM, Bekessy SA, Cantele M, Possingham HP, Watson JEM, Maron M, Keith DA, Garnett ST, Woinarski JCZ, Lindenmayer DB (2019) Spending to save: what will it cost to halt Australia’s extinction crisis? Conserv Lett 12:1–7. https://doi.org/10.1111/conl.12682
Yong C, Ward M, Watson JEM, Reside AE, van Leeuwen S, Legge S, Geary WL, Lintermans M, Kennard MJ, Stuart S, Carwardine J (2023) The costs of managing key threats to Australia’s biodiversity. J Appl Ecol 00:1–13. https://doi.org/10.1111/1365-2664.14377
Acknowledgements
We thank C.J. Yong for guidance on the cost modeling methods and providing these datasets. D. Duncan contributed to the compilation of threats for TECs supported by the National Environmental Science Programme Threatened Species Recovery Hub (Project 7.6). Species and ecological communities’ distribution data were supplied by the Australian Government Department of Agriculture, Water and Environment. J.O.R. was supported by Monash International Tuition Scholarship (MITS). J.C.W. is funded by an Australian Research Council Discovery Early Career Researcher Award, DE210101030.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. J.O.R. was supported by Monash International Tuition Scholarship (MITS). J.C.W. is funded by an Australian Research Council Discovery Early Career Researcher Award, DE210101030.
Author information
Authors and Affiliations
Contributions
J.OR., J.C.W. and C.N.C. conceived the study conception and design. J.OR., J.C.W. and M.S.W. designed the methodology with significant contributions from C.N.C. Material preparation, data collection and analysis were performed by J.OR. The first draft of the manuscript was written by J.OR. and all authors contributed critically to the writing and editing of manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Communicated by Ricardo Correia.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Olivares-Rojas, J., Cook, C.N., Ward, M.S. et al. Species and ecological communities as management surrogates for threatened biodiversity. Biodivers Conserv 33, 987–1008 (2024). https://doi.org/10.1007/s10531-023-02773-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-023-02773-x