
Abstract
One potential legacy of the widespread deforestation of the 1700-1800s and subsequent agricultural abandonment and reforestation across the eastern United States (U.S.) during the 1900s is the establishment of introduced apple trees (Malus domestica) exclusive of naturally occurring native crab apple species. As these apple trees might provide ecological services and a genetic reserve of long-forgotten apple cultivars, a region-wide forest inventory was used to estimate the current extent, condition, and changes in the population of apple trees across eastern forests of the U.S. It was found that hundreds of millions of apple trees currently exist in the forests of the northern U.S. with indications of population senescence as the abundance of seedlings and saplings decline while the abundance of the largest-sized trees and standing dead trees increase. The distribution of forest apple trees is only aligned with native crab apple distributions in limited areas, suggesting distinct feral apple populations derived from agricultural abandonment decades ago. As these extant apple trees are likely relics of former orchards and/or their naturally dispersed progeny, these populations may serve as an in-situ gene bank representing hundreds, if not thousands, of apple cultivars forgotten long ago but adapted to local conditions and perhaps of importance to ensuring the future resilience of domestic apple production in the context of global change.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
From the onset of the colonialization and European settlement of the United States (U.S.) starting along the eastern seaboard and progressing westward (Whitney 1994), forests were widely converted to other land uses such as settlements and agricultural uses in addition to general forest degradation (e.g., removal of high-grade sawtimber) (MacCleery 1992; Irland 1999). It has been claimed that this widespread deforestation across the eastern U.S. was so impactful that its effects can still be seen today on almost every forest acre (Fedkiw 1989). In conjunction with the settlement and deforestation of the northern U.S. from the 1600s onward, newly established agricultural areas were not only planted with a range of annual crops but also with perennial fruit crops such as apple (Russell 1982; Dolan 2009). With lone apple trees at homesteads to expansive apple orchards, the northeast U.S. became synonymous with apple cultivation and a wellspring of diverse new cultivars, founded upon material imported mostly from England and France (Beach 1905; Dolan 2009). Such a relationship between humans and apples is not unique to the northern U.S. as they have been tied together in a close relationship encompassing much of the world and spanning many centuries (Wynne 1975; Hedrick 1925). Apples readily produce offspring from seed and, where humans have carried fruit or seed, apple trees have followed (Beach 1905). As every apple tree that germinates from a seed is genetically new and unique in both tree and fruit characteristics (Beach 1905; Dolan 2009), the named apple cultivars in the commercial orchards and backyards of North America are all grafted trees representing clones of the original seedling that gave rise to each cultivar (Dolan 2009). Apples are in the genus Malus, which is composed of about forty species and many hybrid combinations ranging from tiny to small-fruited “crabapples”, most of which are native to various temperate regions of the world and some of which are cultivated for their ornamental value, to the large-fruited dessert, cider, and culinary apples (Volk et al. 2015; USDA-GRIN 2022).
The modern edible apple, Malus domestica, is the result of intermixing centuries ago of multiple apple species, primarily M. sieversii (native to China and Middle Asia especially Kazakhstan) and M. sylvestris (the European crabapple), with some M. orientalis (native to Iran, Turkey, and the Caucasus), with the crop’s primary center of diversity in Europe (Volk et al. 2015). Native crabapples of the northern U.S. (M. ioensis, M. coronaria, and M. angustifolia) do not appear to have played a notable role in the modern apple or the seedling apple populations of the twenty-first century (Cronin et al. 2020). When Europeans arrived in America more than four hundred years ago, they brought M. domestica seeds and established seedling orchards everywhere they settled from Maine to Georgia, out to the Mississippi River and beyond (Beach 1905). As indigenous people recognized the value of the fruit, they also planted apple trees from seed (Russell 1982). The resulting cultivated populations in North America became a secondary center of diversity of the crop (Volk et al. 2015). North American farmers cultivated the seedlings and then selected among resulting trees for those with desirable qualities (Beach 1905; Bunker 2007; Volk et al. 2015). By the mid-nineteenth century there were well over fifteen thousand named American apple cultivars (Dolan 2008; Bussey 2016), most growing on small farms spread over vast areas that were once forested. Once established in cultivated plantings, apple trees began to form feral seedling populations nearby (Beach 1905). Trees of M. domestica spread readily into the northeastern U.S. likely abetted by the similar latitude as the fruit’s ancestral home across Asia and Europe (Bunker 2007). As an ecotonal forest-edge species adaptable to numerous soil types, M. domestica thrived along fencerows, fields, roads, streams, rivers, and even the seashore (Bunker 2019). One fruiting tree can produce many thousands of seeds over the course of a few years (Brennan 2019) with those fruit/seeds often becoming the food source for a wide range of wildlife that distribute the seeds and thereby propagate new apple trees (Kaminsky 2016). By the early twentieth century, the days of the small, diversified farm with its multi-cultivar apple orchard was mostly a thing of the past (Dolan 2008; Bunker 2007) but those established, numerous long-lived apple cultivar trees remained and became the seed source for contemporary seedling populations (Kaminsky 2016). The seedling apple trees of today might be the only remaining indicators of once-thriving farms or entire villages (Dolan 2008) induced by colonialism centuries ago (Lenzner et al. 2022).
Trees of M. domestica have now naturalized into much of the landscape of North America. Although it is clear from where they originated and how they became established, their status and abundance has not been assessed. Such an assessment would document the outcomes of past cultural land use and is an important basis for understanding the biodiversity of pollinator habitat, the availability of forage for wildlife, and the degree of useful genetic diversity represented by feral apple populations that have become subsumed by the widespread reforestation of the eastern U.S. throughout the 1900s to the present (Thompson et al. 2013; Oswalt et al. 2019). Therefore, the goal of this study was to estimate the current extent, condition, and changes in the population of apple trees (Malus spp.) in U.S. forests as a basis for utility considerations. This study focused primarily on the eastern U.S. where there is notable abundance. Specific objectives were to: (1) assess the current distribution of apple trees in U.S. forests in contrast to native crabapple species, (2) estimate changes from 1998 to 2012 to 2013–2020 in apple tree abundance in U.S. forests by tree size and in comparison to all tree species, (3) examine apple tree abundance by tree size class and components of tree biomass change (e.g., gross growth, mortality, harvests) by U.S. state, and (4) evaluate the sustainability of apple trees in forested land uses via indicators of tree regeneration and hotspots of change relative to competing tree species.
Methods
Forest Inventory Sample Design and Field Protocols
The study relied on the plot network and associated land use and forest attribute data collected by the USDA Forest Service’s Forest Inventory and Analysis (FIA) program, which is the primary source for information about the extent, condition, status, and trends of forest resources across the U.S. (Oswalt et al. 2019). The FIA program applies a nationally consistent sampling protocol using a systematic design covering all ownerships across the U.S. with a national sample intensity of one plot per 2,428 ha using a multi-phase inventory (Bechtold and Patterson 2005). In the first phase, the population of interest is stratified (e.g., forest canopy cover classes) and plots are assigned to strata to increase the precision of estimates. Remotely sensed data may also be used to determine if plot locations have forest cover; only forested land is measured in the field component of the inventory and is defined as being at least 10% canopy coverage with tree species, at least 0.4 ha in size, and at least 36.6 m wide (Bechtold and Patterson 2005). In the second phase, tree and site attributes are measured for plots established in the 2,428-ha hexagons. FIA inventory plots established in forested conditions consist of four, 7.32-m fixed-radius subplots spaced 36.6 m apart in a triangular arrangement with one subplot in the center (USDA 2021, Burrill et al. 2021). All trees (live and standing dead) with a diameter of at least 12.7 cm (at a point 1.37 m above the ground) are inventoried with in situ measurement of diameter, species, height among a host of other attributes (USDA 2021) on forested subplots. A standing dead tree is considered downed dead wood when the lean angle of its central trunk is greater than 45 degrees from vertical (USDA 2021). Within each subplot, a 2.07 m microplot offset 3.66 m from subplot center is established where only live trees with a diameter between 2.5 and 12.7 cm are inventoried.
Dataset and statistical analysis
Field data for this study were taken entirely from the publicly available FIA database (Burrill et al. 2021). The complete field dataset used is available for download at https://apps.fs.usda.gov/fia/datamart/datamart.html (FIA Datamart; USDA 2022). Because the distribution of apple trees in U.S. forests was found to be mostly focused in the northern U.S., spatially tiered analysis was conducted as follows: (1) For the purposes of national-scale analysis, all coterminous U.S. states were included; (2) For spatial displays of current apple tree distribution in U.S. forests along with hotspots of change, only the eastern U.S. was included given the relatively infrequent identification of apple trees in the western U.S.; and (3) For highlighting the states with the highest occurrence of apple trees in northern forests of the eastern U.S., only the following states were analyzed: Delaware (DE), Illinois (IL), Indiana (IN), Iowa (IA), Kentucky (KY), Maine (ME), Maryland (MD), Massachusetts (MA), Michigan (MI), Minnesota (MN), New Jersey (NJ), New York (NY), Ohio (OH), Pennsylvania (PA), Vermont (VT), Virginia (VA), West Virginia (WV), and Wisconsin (WI). All analyses were conducted using R version 3.6.3 (R Core Team 2020) and the package ggmap (Kahle and Wickham 2013). To better assess the status of apple regeneration and recruitment in forests, proportions of apple tree abundance to the abundance of all other competing tree species was calculated. Percentages of total apple basal area occupancy to total live tree basal area were also calculated to enable examination of apple abundance ratios by both demographic (e.g., small trees versus large trees) and site occupancy classes.
Results
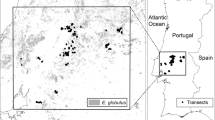
The current range of apple trees in U.S. forests indicated widespread occurrence across the eastern U.S. (Fig. 1) with no suggestion of such a phenomenon in the western U.S. where apple orchards also exist. In the eastern U.S., forest inventory plot network observations of apple trees were prevalent in the northeastern U.S., across the Alleghany plateau of western PA, northern WV, and northeastern OH extending into southern MI, and across the driftless region of western WI and southeastern MN. There was widespread overlap identified between feral apple tree observations and the expected range of M. coronaria especially in southern MI and the Allegheny plateau. Minimal overlap was detected, mostly limited to southern WI, between apple trees and the natural range of M. ioensis. Minimal overlap was detected between apple trees and M. angustifolia, with the latter native crabapple existing in the southeastern U.S. where few apple tree observations were identified.

Forest inventory network observations (red dots, 2013–2020) of apple trees (i.e., feral Malus domestica) in forests of the eastern U.S. in contrast to native ranges of three crab apple species (M. ioensis, M. coronaria, M. angustifolia; distributions derived from Little et al. 1971)
Since the 1998–2012 forest inventory, apple tree abundance has decreased 24% across eastern U.S. forests from an estimated 342 to 259 million trees (Fig. 2). Although there was a decrease in total abundance, net growth on remaining trees resulted in a minimal increase in total above ground apple tree carbon stocks to 4.9 million metric tons representing a 3% increase but reducing its national ranking among eastern tree species from #102 to #108 out of 223 species. In terms of apple tree demography (i.e., seedling regeneration to standing dead trees), changes in abundance across tree size classes and/or life stages indicated a senescing resource as the abundance of apple tree seedlings, small/medium/large trees, and standing dead trees changed by -35%, -30%, 0%, 15%, and 17%, respectively (Fig. 2). Furthermore, compared to all other forest tree species across the eastern U.S. study region, M. domestica had substantial reductions in rankings for seedlings and small trees, from #104 to #112 and #78 to #92, respectively for 1998–2012 vs. 2013–2020 (Fig. 2).

Estimates of apple tree population attributes (total carbon, total number of trees, and abundance by demographic stage) across time compared (by rank and percentage change in population totals) to all other tree species (n = 223) in eastern U.S. forests. Time 1: 1998–2012 and Time 2: 2013–2020, total carbon is in Mg C, total trees and tree sizes are total abundance estimates, tree diameter classes are small 2.54–12.67 cm, medium 12.68–25.37 cm, large > 25.37 cm
Apple tree abundance among tree size classes was highly variable across study states where abundance was appreciable (Fig. 3a). New York was determined to have by far the highest abundance (including seedlings) of forest apple trees at more than 150 million, followed by PA, WI, IL, and MI at 75, 60, 40, and 25 million, respectively. However, most of the abundance was composed of seedlings and saplings (diameter < 12.67 cm), with only ME, MI, NY, OH, and PA possessing large numbers of large trees. Evaluation of components of biomass change between measurement cycles (Fig. 3b) identified that states with the highest ratio of gross biomass growth were IA, KY, MN, NJ, NY, VT, and WI. In contrast, states with the highest ratio of biomass mortality were IL, ME, MD, OH, PA, VA, and WV. The state with the highest ratio of biomass removal (attributed to land use change) was MA at nearly 0.7 followed by ME at 0.2. In terms of forest harvest, states with the highest rates of apple tree harvest activity were NJ, MI, VA, and NY.

Estimates of eastern forest apple population characteristics: (a) Apple tree abundance by state and tree size class; (b) Apple tree biomass by components of change (gross growth, mortality, other removals [i.e., forest land use conversion], and cutting) (tree diameter classes: small 2.54–12.67 cm, medium 12.68–25.37 cm, large > 25.37 cm). No estimates of apple population change are provided for Delaware as no inventory plots containing apple trees were remeasured
Where percentage of apple tree basal area compared to total live tree basal area was less than 10%, the abundance of apple trees by size class was negligible, except for medium- and large-sized trees where it was approximately 5% of total forest abundance (Fig. 4). In forests where the percentage of apple basal area compared to total live tree basal area was between 10 and 50%, apple tree abundance was less than 25% of total live tree abundance especially for seedlings where it was negligible. In forests where apple tree basal area site occupancy percentage was greater than 50%, the abundance of apple seedlings compared to all seedlings was minor at approximately 4% while small- and medium-sized apple tree abundance nearly exceeded 75% of all live trees.

Estimates of the proportions of apple tree abundance (apple trees versus inventory plot totals for all tree species) by tree demographic classes for three levels of total site occupancy as indicated by the proportion of apple tree basal area compared to forest total for trees with a diameter exceeding 12.7 cm) in eastern U.S. forests, 2013–2020
In terms of a cursory examination of spatial hotspots of apple tree change, strong indications of gain were only identified for seedlings in WI, large trees in NY and PA, and standing dead trees in OH and PA (Fig. 5). In contrast, indications of apple tree loss hotspots were common and larger in magnitude, with widespread mild to moderate loss of seedlings across the study region. There were strong indications of small-, medium-, and large-sized apple tree loss in NY and PA (Fig. 5).

Hotspots of gross gain and loss for apple tree abundance by individual apple tree demographic stages (seedlings through standing dead trees), Time 1: 1998–2012 and Time 2: 2013–2020, eastern U.S. forests. Axes are latitude and longitude. Legend scale indicates apple tree population abundance (blue to red), (i.e., the interpolated spatial density of inventory observations) with higher values indicating greater abundance (tree diameter classes: small 2.54–12.67 cm, medium 12.68–25.37 cm, large > 25.37 cm)
Discussion
Although the location and prevalence of current apple orchards across the U.S. is well understood and recognized by society, the ubiquitous presence of apple trees comingled with native crabapples across large swaths of U.S. forests has rarely been noted, let alone quantified. In this study, the presence of hundreds of millions of M. domestica apple trees was estimated across eastern U.S. forests (with indications in the literature and via genotypic evaluations of very limited hybridization with native crabapple populations). Across eastern U.S. forests, M. domestica trees ranked 96 out of 223 tree species in terms of total abundance, suggesting that this resource is not a minor aberration in the inventory of forest resources on a national scale. Although apple trees ranked consistently in the top 100 across all eastern U.S. forest tree species by demographic classes, the highest ranking was for standing dead trees (#65) and the lowest was for seedlings (#112), which suggests population establishment events were decades to more than a century ago. Hence, such trees represent relict abandoned orchard or homestead trees or descendants thereof. As noted by Beach (1905), Bunker (2007), Dolan (2008), and Bussey (2016), M. domestica apple trees were widely established across the eastern tier of the U.S. in the 1700 and 1800 s, which this study found to be widely dispersed throughout contemporary eastern U.S. forests. As M. domestica trees can be one of the first perennials to dominate former pasture sites in former agricultural settings that occur in the northern U.S. (Stover and Marks 1998), perhaps the agricultural abandonment of the 1800 to 1900s (Thompson et al. 2013) led to an increased proliferation of apple seedlings derived from cultivars (which would now be considered heirloom cultivars; Dunbar-Wallis et al. 2022) across the eastern U.S. Given the indicators documented here of M. domestica tree population decline in eastern U.S. forests such as relatively high rates of apple tree mortality, low levels of regeneration, and prevalence of standing dead trees even in sites highly occupied by apple trees, it can be hypothesized that such senescence chronologically aligns with periods of agricultural abandonment (40–140 years ago) across the region. The observed changes in apple tree demography, especially the increase in large-sized trees in states such as NY and PA, would be expected as the forest canopy closed during advanced stages of forest stand development in recent decades (Woodall and Weiskittel 2021).
Although M. domestica apple trees in U.S. forests are an introduced non-native species, they might provide ecosystem services such as wildlife forage or pollinator services. Watson et al. (2011) found that the proportion of forest in proximity to apple orchards in Wisconsin was the strongest predictor of wild bee abundance with positive effects in terms of pollination services (Martins et al. 2015) found that the complementary habitats of meadows and forest in proximity to orchards was positively associated with wild bee species diversity with positive effects on fruit production in southern Quebec. Although our study did not explicitly estimate pollination services provided by apple trees in U.S. forests, their long-standing role in supporting higher levels of pollinator diversity and population abundance across mixed land uses suggests that they supplement pollinator diversity in northern U.S. forests especially when comingled with agricultural land uses.
This study’s estimate of over 250 million M. domestica trees in eastern U.S. forests suggest a resource that might serve as an unintended gene bank of long-forgotten apple cultivars. Some trees will probably be historically documented cultivars not available anywhere else, sought-after but otherwise lost. Known cultivars can be identified among the extant apple trees in abandoned homesteads and orchards, but not all of the grafted trees can yet be assigned to a known cultivar (Routson et al. 2009; Magby et al. 2019; Volk et al. 2021; Dunbar-Wallis et al. 2022). Because such trees represent the outcomes of deliberate selection and probably represent named cultivars in Bussey (2016), they are prime candidates for conservation and further evaluation. Sometimes old orchard or homestead trees that have long since succumbed to reforestation might turn out to be the missing links in the pedigree network that ultimately connects all apple cultivars (Dunbar-Wallis et al. 2022). Pedigree knowledge is powerful for understanding, conserving, managing, and utilizing apple genetic resources (Peace et al. 2019). Apple trees in U.S. forest trees may also have genetic value as the direct descendants of lost cultivars and/or missing pedigree links. With advances in new DNA profiling techniques (e.g., Cronin et al. 2020; Howard & Peace et al. 2021), the parentage and recent ancestry of any apple tree (with at least some M. domestica ancestry) in the North American landscape can now be determined (Dunbar-Wallis et al. 2022). Each seedling tree can also have its own value as a potential new cultivar or as a parent of next-generation cultivars. Contemporary feral apple trees are essentially the same as those in seedling orchards from which classic North American cultivars arose (Gross 2019), other than growing uncultivated. Just as the original spread of seedling trees created a secondary center of diversity of apple beyond Europe and led directly to cultivars and their offspring that now represent the large majority of cultivated apple worldwide, so too do feral populations have the potential for valuable contributions to future apple crop production. Such populations have even advanced further generations and survived more seasons of North American conditions than had the gene pool from which arose ‘Golden Delicious’, ‘McIntosh’, ‘Red Delicious’, and ‘Ralls Genet’ (the latter two being the parents of the most-produced cultivar in the world, ‘Fuji’), Each new seedling represents a unique genetic combination, a key initial step of breeding, but the hundreds of millions of feral trees are far beyond what breeding programs can generate and maintain. For as many U.S. forest apple trees as possible, systematic evaluation for desirable attributes (such as tree hardiness, pest resistance, disease resistance, and fruit appearance, flavor, and yield) would inform selection decisions and could identify some astounding unique individuals.
There are strong indications that the U.S. forest apple resource of M. domestica is in decline. It is important to examine apple trees compared to competing tree species across the landscape as well as the components of its change. Numerous indicators of tree population sustainability pointed to a decline, such as the reduction in total tree abundance, especially in seedlings and smallest trees, and an increase in standing dead trees. Even in forests where apple trees accounted for the majority of the total forest tree basal area, there was determined to be minimal apple tree regeneration with little indication of spatial hotspots of regeneration across the eastern U.S. Furthermore, high rates of mortality relative to gross growth for apple trees in eastern forests were identified. This finding contrasts with wild populations of M. sylvestris in Europe, where large-sized trees (diameter > 44 cm) were very rare (Schnitzler et al. 2014). However, the lack of U.S. forest feral apple regeneration does appear to align with the European wild apple populations where Schnitzler et al. (2014) found rare germination and high sapling mortality, compensated for by periodic fruiting in optimal germination years. Furthermore, Schnitzler et al. (2014) suggested that wild European apple populations might have developed a unique colonizing strategy to take advantage of small-scale, high-severity disturbance events to compensate for the species’ shade tolerance and low growth capacity. As apple trees in forests are expected to be influenced by forest management activities like all other tree species, changes in forest management practices from individual tree selection approaches (i.e., canopy gap formation) to larger regeneration clear-cuts might have deleterious effects on wild apple populations (Schnitzler et al. 2014). In the northern U.S., uneven-aged forestry systems (e.g., irregular shelterwoods) are quite commonly employed compared to the even-aged systems (e.g., clear cutting) of industrial timberlands of the southern U.S., with the exception being spruce/fir forests of ME and aspen/birch types of the upper Midwest. Therefore, a few processes operating at the landscape level (e.g., agricultural abandonment) to stand-level (e.g., individual to group selection silvicultural practices) might have coincided over the past century or more to establish M. domestica trees across northern forests that are now in decline as some of those processes may cease.
Historic orchards can serve as conservation targets where land managers, historic preservation experts, and gene bank collection scientists can collaborate on the conservation of culturally and economically important species (Volk et al. 2021). As M. domestica apple trees in U.S. forests represent a non-native species with some provisioning of ecosystem services, while not markedly displacing native tree species, cultural heritage considerations might be important (Lenzner et al. 2022). Public land managers in the U.S. have a mandate to preserve historic resources including old apple orchards that are threatened by encroaching vegetation, structural instability, disease, and intentional removal (Volk et al. 2021). The identification of a rare apple cultivars on public lands could make an otherwise unremarkable old orchard a candidate for listing in the National Register of Historic Places (Volk et al. 2021).
With the results of this study indicating the forest feral apple (M. domestica) resource is a minor to moderate resource across the eastern U.S. but potentially in decline, conservation strategies may be considered especially in locations where there is still an abundance of medium to large-sized living trees (e.g., NY, PA, WI, MI, ME) but with indications of appreciable levels of mortality. Limited gene flow from orchards to native apple populations coupled with a weak native population structure (i.e., population only found in gaps, forest edges, or poor-quality sites) (Schnitzler et al. 2014) suggests there are few opportunities to reverse the decline in the U.S. forest apple resource that do not involve active forest management. Assuming the factors that led to the North American apple tree resource being established can never be redone, perhaps conservation strategies would usefully focus on targeting populations of genetic importance and remaining large-sized trees with pending senescence and little to no regeneration or recruitment. Several widespread areas of New England (ME, NH, VT, MA, CT, RI), NY, PA, and Lake States (WI, MI, MN) are estimated to possess numerous populations of M. domestica trees in forests with many being composed of larger-sized individuals. If the conservation of these populations and/or genetic material is of priority then conservation could be focused in these areas before these populations further deteriorate.
Conclusion
Non-native apple trees in U.S. forests, descended from reverted orchards and other agricultural land decades and centuries ago, have proliferated to now exist in the hundreds of millions, especially in the northern U.S. This cohort of apple trees, once most likely pioneer trees in abandoned agricultural landscapes, might now be an only minor component of an otherwise fully stocked and maturing forest stand. Such a situation is indicated by the current estimates in this study of a forest apple population that is in decline with potentially senescing larger-sized trees and inadequate regeneration and recruitment of the next generation. Not only are forest apple trees relics of past centuries, they might also serve as a vast repository representing thousands of apple cultivars from previous agrarian communities. Such a forest “gene bank” might be key to increasing the genetic diversity and associated valuable attributes of future apple orchards and their fruit bounty and enhance the resilience of apple production in the face of global change while offering opportunities to revive the production of once popular (and often now lost) cultivars. Without conservation efforts aimed at replicating some of the original population establishment conditions and/or preserving worthwhile genetic material, it is expected that the apple tree population in U.S. forests will continue to decline until they are one of the rarest of U.S. forest relics.
Data Availability
All data used in this study are publicly available at the US Department of Agriculture, Forest Service forest inventory online datamart: https://apps.fs.usda.gov/fia/datamart/datamart.html. All SQL code used to query the aforementioned database, comma delimited output files derived from the SQL source code, and associated R code for analysis/visualization have been provided.
Change history
26 May 2023
A Correction to this paper has been published: https://doi.org/10.1007/s10531-023-02622-x
References
Beach SA (1905) The Apples of New York. 1st edition. J.B. Lyon. 532 p
Brennan A (2019) Uncultivated: wild apples, real cider, and the complicated art of making a living. Chelsea Green Publishing, p 288
Bunker JP Not Far from the Tree: A brief history of the apples and the orchards of Palermo Maine 1804–2004., Bunker JP (2007) Palermo, ME. 190 p
Bunker JP (2019) Apples and the Art of Detection: Tracking Down, Identifying, and Preserving Rare Apples. Out on a Limb Apples. 407 p
Burrill EA, DiTommaso AM, Turner JA, Pugh SA, Menlove J, Christiansen G, Perry CJ, Conkling BL (2021) The Forest Inventory and Analysis Database: database description and user guide version 9.0.1 for Phase 2. U.S. Department of Agriculture, Forest Service. 1026 p. [Online]. Available at: http://www.fia.fs.fed.us/library/database-documentation/
Bussey DJ (2016) The Illustrated History of Apples in the United States and Canada. JAK JAW Press
Cronin D, Kron P, Husband BC (2020) The origins and evolutionary history of feral apples in southern Canada. Mol Ecol 29:1776–1790
Dolan SA (2009) Fruitful legacy: a historic context of orchards in the United States, with technical information for registering orchards in the National Register of historic places. Interior Dept., National Park Service, Olmsted Center for Landscape Preservation, Pacific West Regional Office, p 229
Dunbar-Wallis A, Volk GM, Johnson AM, Schuenemeyer A, Bunker J, Castro D, Little-Siebold T, Pendergast L et al (2022) What’s in a name? The importance of identity in heirloom apple tree preservation. Plants People Planet 1–8. https://doi.org/10.1002/ppp3.10307
Fedkiw J (1989) The evolving use and management of the nation’s forests, grasslands, croplands, and related resources. Gen. Tech. Rep. RM-175. USDA for. Serv. Rocky Mtn Research Station, Fort Collins, CO
Gross BL (2020) How the apple escaped cultivation in North America. Mol Ecol 29:1761–1763
Hedrick UP (1925) Systematic pomology. Macmillan Co., p 488
Howard NP, Peace C, Silverstein KAT, Poets A, Luby JJ, Vanderzande S, Durel C-E, Muranty H et al (2021) The use of shared haplotype length information for pedigree reconstruction in asexually propagated outbreeding crops, demonstrated for apple and sweet cherry. Hortic Res 8:202
Irland LC (1999) The Northeast’s changing forests. Harvard University Press, Cambridge, MA
Kahle D, Wickham H (2013) GGmap: spatial visualization with ggplot2. R J 5:144–161
Kaminsky M (2016) The Wild Apple Forager’s Guide. 84 p
Lenzner B, Latombe G, Schertler A, Seebens H, Yang Q, Winter M, Weigelt P, van Kleunen M et al (2022) Naturalized alien floras still carry the legacy of european colonialism. Nat Ecol Evol 6:1723–1732
Little EL, Jr (1971) Atlas of United States Trees. Volume 1: conifers and important hardwoods. US Department of Agriculture, Forest Service. Misc. Pub. 1146
MacCleery DW (1992) America Forests – A History of Resiliency and Recovery. FS-540. USDA For. Serv., Washington, DC
Magby J, Volk GM, Henk A, Miller S (2019) Identification of historic homestead and orchard apple cultivars in Wyoming. HortScience 54:8–16
Martins KT, Gonzalez A, Lechowicz MJ (2015) Agriculture, Ecosystems, and the Environment. 200: 12–20
Nowacki GJ, Abrams MD (2015) Is climate an important driver of post-european vegetation change in the Eastern United States. Glob Change Biol 21:314–334
Oswalt SN, Smith WB, Miles PD, Pugh SA (2019) Forest Resources of the United States, 2017: a technical document supporting the Forest Services 2020 RPA Assessment. Gen. Tech. Rep. WO-097. U.S. Department of Agriculture, Forest Service, Washington Office, Washington, DC
Peace CP, Bianco L, Troggio M, van de Weg E, Howard NP, Cornille A, Durel C-E, Myles S (2019) Apple whole genome sequences: recent advances and new prospects. Hortic Res 6:59
R Core Team (2020) R: A Language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Routson KJ, Reilley AA, Henk AD, Volk GM (2009) Identification of historic apple trees in the Southwestern United States and implications for conservation. HortScience 44:589–594
Russell HS (1982) A long, deep furrow: three centuries of farming in New England. University Press of New England, p 394
Schnitzler A, Arnold C, Cornille A, Bachmann O, Schnitzler C (2014) Wild european apple (Malus sylvestris (L.) Mill.) Population dynamics: insight from genetics and ecology in the Rhine Valley. Priorities for a future conservation programme. PLoS ONE 9:e96596
Stover ME, Marks PL (1998) Successional vegetation on abandoned cultivated and pastured land in Tompkins County, New York. Journal of the Torrey Botanical Society 125: 150–164
Thompson JR, Carpenter DN, Cogbill CV, Foster DR (2013) Four centuries of change in northeastern United States forests. PLoS ONE 8:e72540
USDA-GRIN (2022) U.S. Department of Agriculture. U.S. National Plant Germplasm System. Version 2.2.2.2. https://npgsweb.ars-grin.gov/gringlobal/taxon/taxonomysearch
USDA, Forest Service. Forest inventory and analysis national core field guide, Volume 1: Field data collection procedures for phase 2 plots, Version 9.1. Available at https://www.fia.fs.fed.us/library/field-guides-methods-proc/index.php
USDA Forest Service (2022) Forest Inventory and Analysis Datamart. U.S. Department of Agriculture, Forest Service. Washington, DC. https://apps.fs.usda.gov/fia/datamart/datamart.html
Volk GM, Chao CT, Norelli J, Brown SK, Fazio G, Peace C, McFerson J, Zhong G-Y et al (2015) The vulnerability of US apple (Malus) genetic resources. Genet Resour Crop Evol 62:765–794
Volk GM, Henk AD (2016) Historic american apple cultivars: identification and availability. J Am Soc Hortic Sci 141:292–301
Volk GM, McCardle K, Serin J, Magby J, Mazur R (2021) Genetic fingerprinting identifies apple (Malus domestica Borkh.) Fruit cultivars in historic orchards on public lands. Genet Resour Crop Evol 68:3133–3144
Watson JC, Wolf AT, Ascher JS (2011) Forested landscapes promote richness and abundance of native bees (Hymenoptera: Apoidea: Anthophila) in Wisconsin apple orchards. Environ Entomol 40:621–632
Whitney GG (1994) From coastal wilderness to fruited plains: a history of environmental change in temperate North America from 1500 to the Present. Cambridge University Press, New York
Woodall CW, Weiskittel AR (2021) Relative density of United States forests has shifted to higher levels over last two decades with important implications for future dynamics. Sci Rep 11:18848
Wynne P (1975) Apples: History, Folklore, Horticulture, and Gastronomy. Hawthorn Books. 280 p
Acknowledgements
The authors wish to recognize the hundreds of foresters and associated field crews over two decades who collected the inventory data used in this study that covers the entire eastern United States. Special thanks are extended to Anthea Prasad for providing digitized crabapple distributions based on Little’s 1971 works and for his foundational research on tree range science.
Funding
CPP was supported by the USDA National Institute of Food and Agriculture Hatch project 1014919, Crop Improvement and Sustainable Production Systems (WSU reference 00011).
Author information
Authors and Affiliations
Contributions
CWW conceived the study, performed data analyses, and wrote the manuscript. GCL assisted with data analyses and manuscript editing. JPB assisted with conceiving the study, writing the introduction, and general manuscript editing. CPP assisted with manuscript writing/editing, especially the introduction and discussion. JMF assisted with identifying initial study results as part of larger joint federal/state Maine forest analyses, along with manuscript editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval
Not applicable.
Additional information
Communicated by David Hawksworth.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Woodall, C., Liknes, G.C., Bunker, J.P. et al. Forgotten forest relics: Apple trees (Malus spp.) in eastern U.S. forests. Biodivers Conserv 32, 1657–1670 (2023). https://doi.org/10.1007/s10531-023-02569-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-023-02569-z