
Abstract
Populations inhabiting the periphery of species distribution ranges may experience suboptimal environmental conditions and higher vulnerability to anthropogenic pressures. Disentangling the role of natural and human-related factors and the relationships among them in these marginal areas is thus key to understand and prevent species declines and range reductions. We analysed Eurasian otter (Lutra lutra) occurrence patterns in relation to anthropogenic pressures and natural environmental gradients in Morocco, an area constituting the arid limit of the species’ global range. The probability of otter occurrence was higher as terrain ruggedness increased and at intermediate elevations, and lower in catchments exposed to higher anthropogenic pressures. Otters tended to be rare at higher elevations and in areas with less annual precipitation, probably as a result of trophic resource limitations and large water flow fluctuations, respectively. A combination of natural and anthropogenic factors, both at drainage area and local scales, was needed to understand the current occurrence of the species. Our study highlights a need for urgent action to conserve the Eurasian otter in northern Africa, where freshwater ecosystems and their associated biodiversity are threatened by rapid human development in areas of marginal climatic conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Habitat occupancy by species may be influenced by both natural environmental variability (Gaston 2003; Oldfather et al. 2020) and by human activities (Collins and Kays 2011; Loss et al. 2015; Hill et al. 2020). Detecting the relative influence of the different factors is critical for species conservation in changing environments (Brook et al. 2008), especially for populations persisting in areas highly susceptible to climate change and under increasing anthropogenic pressure, such as freshwater habitats (e.g. Cianfrani et al. 2018). Typically, species at the margin of their distribution ranges experience suboptimal conditions near their tolerance limits. In those areas, even small variations in the environment may have detrimental effects on species persistence (Mehlman 1997; Williams et al. 2003; Ovaskainen and Meerson 2010; Yackulic et al. 2011).
The Eurasian otter (Lutra lutra, simply the otter henceforth) in Northern Africa constitutes a model to analyse the natural and anthropogenic determinants of species occurrence in the arid limit of its range. The otter is a semi-aquatic mammalian carnivore that obtains all its food from aquatic systems (Clavero et al. 2003; Krawczyk et al. 2016). It shows a wide distribution, covering most of the Palearctic and part of the Indo-Malay biogeographic realms (Roos et al. 2015). Northern Africa constitutes the south-western bound of the global otter distribution, and includes some of the driest areas where the species occurs (Riesco et al. 2020).
In recent decades, the conservation status of the otter tended to diverge between Northern Africa and Europe. In most central and western European countries, the otter declined steadily between the 1950s and the 1980s, probably due to the effects of direct persecution and environmental pollutants such as PCBs, DDT and heavy metals (Mason and Macdonald 1986; Macdonald and Mason 1994; Conroy and Chanin 2002). Since then, otter trend reversed (Conroy and Chanin 2002), probably as a result of the implementation of environmental regulations and the reduction of direct persecution (Loy and Duplaix 2020). However, otter declines are still ongoing, or at least not fully reversed, in other European and Asian developing countries (Gorgadze 2011; Balestrieri et al. 2016; Jha et al. 2020) and in Northern Africa (Delibes et al. 2012; Riesco et al. 2020).
We studied the natural and anthropogenic factors associated with otter occurrence in Northern Africa, where aridity limits the availability of suitable habitats, and where pressures associated to rapid human development are increasing. We analysed environmental correlates of otter occurrence at the drainage (i.e. the area of land that collects the water that drains to a particular site) and local (i.e. surrounding area of a river stretch) scales. At both scales we specifically hypothesized that otter occurrence would be positively associated with increasing rainfall and terrain ruggedness and negatively influenced by agricultural, industrial and urban pressures. Our results are relevant to understand the conservation status of the otter in Northern Africa and to design management strategies for the recovery of the species.
Methods
Study area
Morocco includes an appropriate representation of the climatic, orographic and human distribution gradients found in Northern Africa (Franchimont 2001). While north and north-western areas feature a Mediterranean-type climate, southern and eastern areas are characterised by semi-arid or arid conditions. Annual rainfall is highly variable within the country, ranging from relatively high precipitation up to over 1000 mm per year in the major mountain ranges—Rift and the Atlas—to < 50 mm in semi-arid and arid areas. Human population and the main productive activities are also unequally distributed, with large cities (hosting 95% of the around 37 million country’s population) and industries occupying the fertile coastal plains to the west and north of the Atlas Mountains. By contrast, south and east of these mountains human population density is very low, and the main productive activities are pastoralism and subsistence agriculture.
Otter surveys
We collected data on otter presences and absences in 270 sites distributed across 34 Moroccan river basins, visited between April 2011 and April 2013. This dataset includes 90 sites from a previous study (Delibes et al. 2012) together with 180 additional sites. The survey sites were selected a priori with the goal of covering as much of the country’s river network as feasible and being at least 30 km apart to uniformly surveying the whole region. We applied the standard otter survey method (Chanin 2003), which consists in sampling up to 600 m along the river shore searching for otter signs (spraints, anal scent gland secretions and tracks). River shores were inspected by at least two experienced surveyors in each survey. A site was coded as positive, and the survey was finished, as soon as an otter sign was found, while it was considered negative when the 600 m-long transect was completed without finding any sign. Otters use spraints and secretions for intraspecific communication placing them at conspicuous and predictable sites (e.g. prominent rocks, ledges, bridges, sand banks, etc.). Furthermore, otter activity concentrates on river shores, where tracks are frequently printed and easily detected. Therefore, the standard otter survey is a cost-effective and reliable method for presence-absence studies. The sensitivity and specificity of this protocol have shown to be very high (Kruuk et al. 1986; Mason and Macdonald 1987; Fusilo et al. 2007; Marcelli et al. 2012), especially when multiple and experienced surveyors are involved (Jeffress et al. 2011).
Spatial scales of analysis
River systems are structured in a nested hierarchical network (Fig. 1), in which ecological processes at any stretch within the network are tightly dependent upon upstream conditions and environmental pressures that may propagate across the drainage area (e.g. amount of rain, water abstraction, pollution, etc.). At the same time, otter occurrence at a particular site may be also influenced by the local ecological conditions and pressures. Therefore, we considered two spatial scales for the characterization of otter habitats: the drainage area (i.e., environmental features in the upper landscape draining to each catchment) and at the local-scale (i.e., a circular area surrounding each sampling site).

A schematic representation of the hierarchical hydrological units used in our analyses. Four catchments are represented. The drainage area for sampling point 1 is the same as the catchment (dashed). For sampling point 2, three catchments constitute the drainage area (all shaded areas). The outer boundary represents the river basin
Catchments characterize the immediate area of terrestrial influence on the aquatic environment and constitutes the basic unit in the hierarchical spatial structure of freshwater systems (e.g. the upstream–downstream gradient in river networks; Clavero et al. 2010; Linke et al. 2019). The drainage area is the land where precipitation accumulates and drains into one point in the river network. In our analyses, drainage areas are made up of the catchment where the sampling site was located together with all catchments upstream (Fig. 1). We delimited catchments using a digital elevation model at 3 arc-seconds resolution from the Radar Topography Mission (Rabus et al. 2003), applying the ARC Hydro Tools in Arc GIS 10.1. Then, we assigned the 270 transects to 205 catchments (mean area ± SD = 534 ± 340 km2) from 34 different river basins.
At the local scale, we defined a buffer area of 5-km diameter around each transect centre. This size was chosen since is likely that a large portion of the home range of an otter would fit within a 5-km river length (Erlinge 1967; Ó Néill et al. 2009) and therefore it represents the local conditions potentially affecting the home-range level of habitat selection.
Environmental predictors
We calculated two sets of variables—natural and anthropogenic—at the two spatial scales—drainage area and local buffers—for each sampled site. At the drainage area, natural variables were size of the drainage area, annual precipitation, and the precipitation coefficient of variation. We calculated size of the area (in km2) as the total area draining to each sampling site—i.e., where surface waters converge at the sampling site. This variable was log-transformed before any statistical analysis. Annual precipitation and coefficient of variation were compiled from the 1 × 1 km WorldClim database (Hijmans et al. 2005). We hypothesized that the probability of otter occurrence would increase with the total area, an indicator of the size of the aquatic system, and with precipitation, a measure of the availability of water, and it would decrease with increasing precipitation coefficient of variation due to increased instability of water availability. Anthropogenic variables included population density, night-time light intensity and crop percent cover. We used National demographic municipality statistics from 2004 (Haut Commissariat au Plan 2004) to calculate population density and converted this density to a gridded dataset with 1-km resolution. Night-time light intensity was extracted from DMPS/OLS satellite data from year 2010 using the annual average brightness of satellite-observed stable night-time lights at a spatial resolution of 30 arc-seconds (Elvidge et al. 2001). Night-time lights are generally associated with the prevalence of urban areas and economic activities (Doll et al. 2006) and they are often used as an indicator of pressures on biodiversity (Levin et al. 2020). Crop percent cover was calculated from the reclassification of the African subset of the GlobCover dataset (Arino et al. 2007) combining into one single category all crop types and mosaics between natural vegetation and crops, including: (i) irrigated and rainfed croplands and herbaceous crops; (ii) rainfed shrub or tree crops; (iii) mosaic croplands; and (iv) mosaics between cropland and grassland, shrubland or forest. We hypothesized that otter occurrence would be more frequent in drainage areas with lower human pressures (population density, nightlight intensity and crop cover).
At the local scale, natural variables were maximum elevation and ruggedness within the 5-km buffer. Both were calculated using the data from the Radar Topography Mission described above. We hypothesized that the probability of otter occurrence would increase with increasing terrain ruggedness since rougher terrain make access to river stretches more difficult for humans. We also hypothesised that otter occurrence would have a unimodal relationship with elevation (i.e. peaking at intermediate elevations), following results from studies in Mediterranean and arid areas (Clavero et al. 2010; Riesco et al. 2020). Local-scale human variables were population density, night-time light intensity and crop percent cover.
Modelling strategy
We specified a priori a set of nine different plausible models that resulted from the combinations of the different groups of factors involved in our hypotheses. i.e., anthropogenic and natural, at the two scales—drainage area and local buffers. Specifically, we tested if otter occurrence was associated to anthropogenic vs. natural factors observed at one or both scales, respectively, or by a combination of the different factors and scales (Table 1; Appendix Table A1). We fitted all models using Generalized Linear Mixed Models (GLMM; Bolker et al. 2009) by the Lapplace approximation, including the catchment nested within the drainage area as a random term to account for the non-independence of observations from sites within the same hydrological unit. The size of the drainage area and population density variables were log-transformed. In order to prevent collinearity effects, we excluded from a priori models variables with correlation rs > 0.50. After preliminary exploration of the models, we eliminated the variable from each pair that contributed the least to explain otter occurrence probability. Following these criteria, we excluded the population density and proportion of crops at the drainage area from models combining natural and human variables; and the local-scale intensity of nightlights from all models.
We ranked models using the second-order Akaike’s Information Criterion AICc and calculated their differences (ΔAICc) and the associated model selection probabilities (wi; Anderson 2008). In addition, we estimated Pseudo-R2 statistics as an indicator of the model’s goodness of fit, separating the marginal component (i.e. variability explained by the fixed factors; \({R}_{m}^{2}\)) and the conditional component (i.e. variability explained by both fixed and random factors; \({R}_{c}^{2}\); Nakagawa and Schielzeth 2013). Finally, we draw inference from the best approximating model by exploring the relative influence of each predictor in the model from the scaled parameter estimates and associated standard errors.
Results
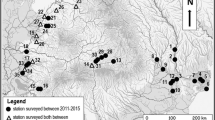
Otter occurrence was detected in 141 out of 270 survey sites (52%; Fig. 2; Appendix 1 Table A2), along a wide range of environmental conditions. For example, otter occupied sites with annual precipitation ranging from 50 to 875 mm, elevations from sea level up to ca. 2300 m, and human population densities between 1.5 and 2700 inhabitants/km2.

The distribution of positive (black circles) and negative (white circles) otter surveys. Main rivers and tributaries are also shown. The grey scale of the map represents the mean annual precipitation
In general, models that included both natural and anthropogenic predictors and integrating the two scales of analysis performed better than models using one set of variables and/or one spatial scale (Table 1). In general, models reflecting the anthropogenic influence had a stronger support than natural ones at the scale of drainage areas, while the contrary happened at the local scale (although ruggedness may be a proxy for both human and natural effects; see the “Discussion” section). The best approximating model to describe otter occurrence in Morocco combined human and natural variables at both spatial scales. This model was clearly superior, resulting in ΔAICc > 15 units than the second competing model and wi > 0.99.
According to scaled estimates of the best model (Table 2), we found strong positive associations between otter occurrence and the annual precipitation and the size of the drainage area, and a strong negative association with night-time lights intensity. The influence of night-light intensity and precipitation were independent, since sites with otters tended to have lower light intensities than sites without otters for any value along the annual precipitation gradient of our dataset (Fig. 3). The effect of the precipitation coefficient of variation in the drainage areas was weak. At the local scale, site elevation had a very strong quadratic influence on otter occurrence, followed by ruggedness (Table 2). The positive association between otter occurrence and the local population density was weak and resulted in high estimate uncertainty. Figure 4 illustrates the most important associations resulting from the selected model.

The relationship between the average precipitation (mm) and night-lights intensity at the drainage area in positive otter sites (black dots, solid line) and in negative sites (white dots, dashed line). Sites with otter: \(y=-0.79+9\times {10}^{-4}\); without otter: \(y=-0.45+9\times {10}^{-4}\). Anova test for the difference between intercepts: F = 19.6; P < 0.001

Natural and anthropogenic factors influencing otter occurrence at the drainage area and local scales. Fitted probabilities ± SE are represented for the most influential variables according to the best approximating model. Each prediction represents the full range of variation of the predictor observed in our study, while keeping the rest of predictors constant at the average value observed in otter positive sites
Discussion
Otter distribution at the arid limit of its distribution range was influenced by both natural and anthropogenic factors. The species was more likely to occur in less humanized, more humid and more rugged environments at intermediate elevations.
The anthropogenic influence on otter the distribution, revealed from the intensity of night lights, should warn on the potential consequences of ongoing accelerated changes in Morocco, if mitigation measures are not put in place. The development of roads, urban areas and industrialization have all increased rapidly in recent years as a consequence of the socioeconomic change experienced by the country. Between 1961 and 2012 the population in Morocco has grown by 256% (Abécassis 2009; World Bank 2014). Agriculture has developed extensively during the last quarter of the twentieth century with agricultural lands covering 68.5% of the country by the mid-2010s, a figure that was 55% in the 1960s (World Bank 2014). The spread of agriculture has also increased the pressure on freshwater ecosystems through growing water withdrawals, river regulation and pollution (Kettle et al. 2011; El Alami and Fattah 2020), including increases in pollutants such as PCBs, DDT and mercury (Berkat and Tazi 2006; Benbakhta et al. 2014).
We did not detect the expected direct negative relationship between the distribution of agricultural land and otter presence. In our dataset, the detrimental effects of agriculture could have been masked by the relationship between agriculture and rainfall (rs = 0.82), the latter being also a major direct driver of otter occurrence. In any case, water-demanding agriculture may still seriously imperil otter and other freshwater-dependent species in Morocco. As observed in Southern Europe during previous decades, the expansion of intensive agriculture is often followed by increases in the amount of pollutants dumped into freshwater ecosystems, severe changes in river hydrology, and a reduction of fish availability (Reid et al. 2019). However, there may be a significant time-lag between the incidence of those impacts and their detection through presence-absence surveys, because these do not allow recording changes in population abundance that may precede local extinctions (Ceballos et al. 2017).
As in our study area, precipitation has been identified as a climatic predictor of otter presence in Europe at a continental scale (Cianfrani et al. 2011), and particularly in the Mediterranean climate region (Prenda et al. 2001; Clavero et al. 2010), where high and unpredictable seasonal and inter-annual precipitation variability influences the structure and functioning of freshwater ecosystems (Gasith and Resh 1999). Rainfall seems to be decreasing and is predicted to further decline in Morocco due to global climate change (Filahi et al. 2017; Driouech et al. 2020). Therefore, some habitats where otter still occurs may soon become unsuitable due to aridification.
As in the case of precipitation, the positive relationship between the size of the drainage area and otter occurrence may also be reflecting the large water availability of major river systems. Large drainage basins generate more productive riverine environments than small watercourses, allowing the occurrence of rich and diverse aquatic communities, and buffer environmental variability, reducing the probability of local extinctions (Eadie et al. 1986; Watters 1992; Carl et al. 2021). Species-area patterns described for relevant otter prey in river systems, particularly fish, have been proposed to drive area-related variation in otter trophic ecology (e.g. Clavero et al. 2008) and arguably are the mechanism explaining the positive effect of drainage area on otter occurrence found in our results.
Otters were more likely to occur at intermediate altitudes. The scarcity or lack of prey in the oligotrophic high mountain aquatic systems explains the species absence at high altitudes (Barbosa et al. 2003; Clavero et al. 2010). However, the scarcity of trophic resources does not seem a plausible explanation for the low prevalence of the otter at low altitudes in Morocco. In European rivers, otters have often been found also at lower reaches down to the sea level (Heggberget and Moseid 1994; Clavero et al. 2006; Parry et al. 2011), and the same was occasionally recorded in our surveys. The frequent absence of otters at low elevations in Morocco may be explained by the intermittency of waters of the downstream reaches in small and medium-sized rivers, making these rivers saltier and prone to drought during the summer (Riesco et al. 2020). However, otters were also very rare in lower reaches of most large rivers despite they maintaining permanent water flows and having abundant fish populations (Azeroual et al. 2000; Nouiri et al. 2007). Here, the pattern resembles the one reported in Southern Europe during the 1980s when the species was mainly confined to mountainous areas (Elliot 1983; Delibes 1990). We argue that the demographic, industrial and agricultural development of Morocco have altered freshwater systems (Mehdaoui et al. 2000; Tahiri et al. 2005; El Morhit et al. 2009), and that this degradation is magnified in lowlands due to the directional interconnectivity of the freshwater systems. Moreover, lowlands are usually more urbanised and altered than hilly and mountain areas.
Otters were found more frequently in areas with high local ruggedness. We treated ruggedness as a natural predictor, but this variable could be related to both natural and human influences on the species occurrence. On the one hand, terrain roughness may be a proxy for the availability of refuge, which favours otter occurrence (Prenda and Granado-Lorencio 1996; Marcelli and Fusillo 2009). But, on the other hand, ruggedness may also indicate the intensity of human presence, since it is related to the accessibility of people to the river. Higher ruggedness could thus reduce direct otter persecution and competition between otter and people for resources such as water, fish and riparian vegetation (El Alami et al. 2020).
Conclusions
Morocco and other countries in Northern Africa are immersed in a rapid process of development that leads to socio-economic growth and improvement in the quality of life of their people. These processes, if they occur without effective conservation planning, can generate an undesired degradation of ecosystems and loss of biodiversity, especially affecting freshwater biodiversity in semiarid and arid regions. We have shown that human pressures directly limit otter distribution in Morocco, with an impact on species occurrence comparable to that of precipitation. With increasing levels of urbanization and industrialization in a rapidly developing country, our results suggest a serious and widespread threat to otter populations in Northern Africa, where aridity conditions already impose a sharp constraint on the natural distribution of wetlands. However, the persistence of the species in some agriculture-dominated lands detected in this study also point out to opportunities for reconciling conservation in human-dominated landscapes, as has already occurred in Europe. An increased protection of the species and sustainable management of rivers, including precluding the use of pesticides that imperil freshwater ecosystems, have already shown to be effective in reversing the decline of otters and associated freshwater ecological communities in other countries.
Data availability
The otter sampling data used for this study is included as electronic supplementary material. Also, we will make this data available via GBIF.
Code availability
Not applicable.
References
Abécassis F (2009) La mise en place du réseau routier marocain: Aperçu historique. In: L’Atlas à l’orient musulman Contrib. en hommage à Daniel Rivet. Available at https://halshs.archives-ouvertes.fr/halshs-00435869
Anderson DR (2008) Model based inference in the life sciences: a primer on evidence. Springer, New York
Arino O, Gross D, Ranera F et al (2007) GlobCover: ESA service for Global land cover from MERIS. Int Geosci Remote Sens Symp. https://doi.org/10.1109/IGARSS.2007.4423328
Azeroual A, Crivelli AJ, Yahyaoui A, Dakki M (2000) L’ichtyofaune des eaux continentales du Maroc. Cybium 24:17–22
Balestrieri A, Messina S, Pella F et al (2016) Eurasian otter Lutra lutra in developing countries: a resurvey of Albania 22 years after the fall of communism. Oryx 50:368–373. https://doi.org/10.1017/S0030605314000921
Barbosa AM, Real R, Olivero J, Vargas JM (2003) Otter (Lutra lutra) distribution modeling at two resolution scales suited to conservation planning in the Iberian Peninsula. Biol Conserv 114:377–387. https://doi.org/10.1016/S0006-3207(03)00066-1
Benbakhta B, Khallaf M, Fekhaoui M et al (2014) Organochlorine pesticides in sediments from the atlantic coast of Morocco. Int J Innov Appl Stud 6:1129–1137
Berkat O, Tazi M (2006) Morocco. Country pasture/forage resource profiles. Food and Agriculture Organization of the United Nations (FAO), Rome
Bolker BM, Brooks ME, Clark CJ et al (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol 24:127–135. https://doi.org/10.1016/j.tree.2008.10.008
Brook BW, Sodhi NS, Bradshaw CJA (2008) Synergies among extinction drivers under global change. Trends Ecol Evol. https://doi.org/10.1016/j.tree.2008.03.011
Carl LM, Esselman PC, Sparks-Jackson BL, Wilson CC (2021) The species—area relationship for a highly fragmented temperate river system. Ecosphere 12:e03411. https://doi.org/10.1002/ecs2.3411
Ceballos G, Ehrlich PR, Dirzo R (2017) Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc Natl Acad Sci USA 114:E6089–E6096. https://doi.org/10.1073/pnas.1704949114
Chanin P (2003) Monitoring the Otter: Lutra lutra. Conserving Natura 2000 10. English Nature, Peterborough
Cianfrani C, Le LG, Maiorano L et al (2011) Adapting global conservation strategies to climate change at the European scale: the otter as a flagship species. Biol Conserv 144:2068–2080. https://doi.org/10.1016/j.biocon.2011.03.027
Cianfrani C, Broennimann O, Loy A, Guisan A (2018) More than range exposure: global otter vulnerability to climate change. Biol Conserv 221:103–113. https://doi.org/10.1016/J.BIOCON.2018.02.031
Clavero M, Prenda J, Delibes M (2003) Trophic diversity of the otter (Lutra lutra L.) in temperate and Mediterranean freshwater habitats. J Biogeogr 30:761–769. https://doi.org/10.1046/j.1365-2699.2003.00865.x
Clavero M, Prenda J, Delibes M (2006) Seasonal use of coastal resources by otters: comparing sandy and rocky stretches. Estuar Coast Shelf Sci 66:387–394. https://doi.org/10.1016/j.ecss.2005.09.011
Clavero M, Prenda J, Blanco-Garrido F, Delibes M (2008) Hydrological stability and otter trophic diversity: a scale-insensitive pattern? Can J Zool Can Zool 86:1152–1158. https://doi.org/10.1139/z08-09
Clavero M, Hermoso V, Brotons L, Delibes M (2010) Natural, human and spatial constraints to expanding populations of otters in the Iberian Peninsula. J Biogeogr 37:2345–2357. https://doi.org/10.1111/j.1365-2699.2010.02377.x
Collins C, Kays R (2011) Causes of mortality in North American populations of large and medium-sized mammals. Anim Conserv 14:474–483. https://doi.org/10.1111/j.1469-1795.2011.00458.x
Conroy JWH, Chanin PRF (2002) The status of the Eurasian otter (Lutra lutra) in Europe—a review. In: Conroy JWH, Yoxon P, Gutleb AC (eds) Proceedings of the first otter toxicology conference. International Otter Survival Fund, Skye, pp 7–28
Delibes M (1990) La nutria (Lutra lutra) en España. Serie Técnica. ICONA, Madrid
Delibes M, Calzada J, Clavero M et al (2012) The near threatened Eurasian otter Lutra lutra in Morocco: no sign of recovery. Oryx 46:249–252. https://doi.org/10.1017/S0030605311001517
Doll CNH, Muller JP, Morley JG (2006) Mapping regional economic activity from night-time light satellite imagery. Ecol Econ 57:75–92. https://doi.org/10.1016/j.ecolecon.2005.03.007
Driouech F, Stafi H, Khouakhi A et al (2020) Recent observed country-wide climate trends in Morocco. Int J Climatol. https://doi.org/10.1002/joc.6734
Eadie JM, Hurly TA, Montgomerie RD, Teather KL (1986) Lakes and rivers as islands: species-area relationships in the fish faunas of Ontario. Environ Biol Fishes 15:81–89. https://doi.org/10.1007/BF00005423
El Alami A, Fattah A (2020) Olive mill wastewater causing pollution in the Oum Er Rbia River and potential environmental effects and impact on the Eurasian Otter. J Anal Sci Appl Biotechnol 2:110–115. https://doi.org/10.48402/IMIST.PRSM/jasab-v2i2.24324
El Alami A, Fattah A, Chait A (2020) A survey of the Eurasian Otter Lutra lutra and human-otter interaction in the middle Oum Er Rbia river, Morocco. IUCN Otter Spec Gr Bull 37:219–231
El Morhit M, Fekhaoui M, Elie P et al (2009) Heavy metals in sediment, water and the European glass eel, Anguilla anguilla (Osteichthyes: Anguillidae), from Loukkos River estuary (Morocco, eastern Atlantic). Cybium 33:219–228
Elliot KM (1983) The otter (Lutra lutra L.) in Spain. Mamm Rev 13:25–34. https://doi.org/10.1111/j.1365-2907.1983.tb00262.x
Elvidge CD, Imhoff ML, Baugh KE et al (2001) Night-time lights of the world: 1994–1995. Isprs J Photogramm Remote Sens 56:81–99. https://doi.org/10.1016/s0924-2716(01)00040-5
Erlinge S (1967) Home range of the otter Lutra lutra L. in Southern Sweden. Oikos 18:186. https://doi.org/10.2307/3565098
Filahi S, Tramblay Y, Mouhir L, Diaconescu EP (2017) Projected changes in temperature and precipitation indices in Morocco from high-resolution regional climate models. Int J Climatol 37:4846–4863. https://doi.org/10.1002/joc.5127
Franchimont J (2001) National study on biodiversity. Synthesis report. Kingdom of Morocco. Ministery of territorial Planing, Water and Environment. Department of Environment, Rabat
Fusillo R, Marcelli M, Boitani L (2007) Survey of an otter Lutra lutra population in Southern Italy: site occupancy and influence of sampling season on species detection. Acta Theriol (Warsz) 52:251–260. https://doi.org/10.1007/BF03194221
Gasith A, Resh VH (1999) Streams in Mediterranean climate regions: abiotic influences and biotic responses to predictable seasonal events. Annu Rev Ecol Syst 30:51–81. https://doi.org/10.1146/annurev.ecolsys.30.1.51
Gaston KJ (2003) The structure and dynamics of geographic ranges. Oxford University Press, Oxford
Gorgadze G (2011) Current status and threats affecting otter (Lutra lutra) population in the South Caucasus. In: Prigioni C, Loy A, Balestrieri A, Remonti L (eds) XIth International Otter Colloquium. Hystrix, The Italian Journal of Mammalogy
Haut Commissariat au Plan (2004) Recensement general de la population et de l’habitat 2004: caracteristiques demographiques et socio economiques de la population. Rapport national. Royaume du Maroc. Haut Commissariat au Plan
Heggberget TM, Moseid K-E (1994) Prey selection in coastal Eurasian otters Lutra lutra. Ecography (Cop) 17:331–338. https://doi.org/10.1111/j.1600-0587.1994.tb00110.x
Hijmans RJ, Cameron SE, Parra JL et al (2005) Very high resolution interpolated climate surfaces for global land areas. Int J Climatol 25:1965–1978. https://doi.org/10.1002/joc.1276
Hill JE, DeVault TL, Wang G, Belant JL (2020) Anthropogenic mortality in mammals increases with the human footprint. Front Ecol Environ 18:13–18. https://doi.org/10.1002/fee.2127
Jeffress MR, Paukert CP, Sandercock BK, Gipson PS (2011) Factors affecting detectability of river otters during sign surveys. J Wildl Manag 75:144–150. https://doi.org/10.1002/jwmg.12
Jha RR, Silwal T, Yoxon GM et al (2020) Status of otters in Nepal: a link with ancient waterways and people. In: Regmi G, Huettmann F (eds) Hindu Kush-Himalaya watersheds downhill: landscape ecology and conservation perspectives. Springer, Cham, pp 409–418
Kettle AJ, Asbjørn Vøllestad L, Wibig J (2011) Where once the eel and the elephant were together: decline of the European eel because of changing hydrology in southwest Europe and northwest Africa? Fish Fish 12(4):380–411
Krawczyk AJ, Bogdziewicz M, Majkowska K, Glazaczow A (2016) Diet composition of the Eurasian otter Lutra lutra in different freshwater habitats of temperate Europe: a review and meta-analysis. Mamm Rev 46:106–113. https://doi.org/10.1111/mam.12054
Kruuk H, Conroy JWH, Glimmerveen U, Ouwerkerk EJ (1986) The use of spraints to survey populations of otters Lutra lutra. Biol Conserv 35:187–194. https://doi.org/10.1016/0006-3207(86)90050-9
Levin N, Kyba CCM, Zhang Q et al (2020) Remote sensing of night lights: a review and an outlook for the future. Remote Sens Environ 237:111443. https://doi.org/10.1016/J.RSE.2019.111443
Linke S, Lehner B, Ouellet Dallaire C et al (2019) Global hydro-environmental sub-basin and river reach characteristics at high spatial resolution. Sci Data 6:1–15. https://doi.org/10.1038/s41597-019-0300-6
Loss SR, Will T, Marra PP (2015) Direct mortality of birds from anthropogenic causes. Annu Rev Ecol Evol Syst 46:99–120. https://doi.org/10.1146/annurev-ecolsys-112414-054133
Loy A, Duplaix N (2020) Decline and recovery of the otter in Europe. Lessons learned and future challenges. J Mt Ecol 13:1–8
Macdonald SM, Mason CF (1994) Status and conservation need of the otter (Lutra lutra) in the Western Palaearctic. Council of Europe, Strasbourg
Marcelli M, Fusillo R (2009) Assessing range re-expansion and recolonization of human-impacted landscapes by threatened species: a case study of the otter (Lutra lutra) in Italy. Biodivers Conserv 18:2941–2959. https://doi.org/10.1007/s10531-009-9618-2
Marcelli M, Poledník L, Poledníková K, Fusillo R (2012) Land use drivers of species re-expansion: inferring colonization dynamics in Eurasian otters. Divers Distrib 18:1001–1012. https://doi.org/10.1111/j.1472-4642.2012.00898.x
Mason CF, Macdonald SM (1986) Otters: ecology and conservation. Cambridge University Press, Cambridge
Mason CF, Macdonald SM (1987) The use of spraints for surveying otter Lutra lutra populations: an evaluation. Biol Conserv 41:167–177. https://doi.org/10.1016/0006-3207(87)90100-5
Mehdaoui O, Fekhaoui M, Descoíns C (2000) Accumulation and biomagnification of organochlorine insecticides in molluscs and fish of the Moulay Bouselham lagoon, Morocco. Cah Sante 10:373–379
Mehlman DW (1997) Change in avian abundance across the geographic range in response to environmental change. Ecol Appl 7:614–624. https://doi.org/10.2307/2269525
Nakagawa S, Schielzeth H (2013) A general and simple method for obtaining R 2 from generalized linear mixed-effects models. Methods Ecol Evol 4:133–142. https://doi.org/10.1111/j.2041-210x.2012.00261.x
Nouiri H, Berrebi P, Lbadaoui K et al (2007) Seasonal variations of abundance and size of recruits of Mugilidae in the coastal lagoon of Merja Zerga (Atlantic coast of Morocco). Cybium 31:427–434
Ó Néill L, Veldhuizen T, de Jongh A et al (2009) Ranging behaviour and socio-biology of Eurasian otters (Lutra lutra) on lowland mesotrophic river systems. Eur J Wildl Res 55:363–370. https://doi.org/10.1007/s10344-009-0252-9
Oldfather MF, Kling MM, Sheth SN et al (2020) Range edges in heterogeneous landscapes: integrating geographic scale and climate complexity into range dynamics. Glob Chang Biol 26:1055–1067. https://doi.org/10.1111/gcb.14897
Ovaskainen O, Meerson B (2010) Stochastic models of population extinction. Trends Ecol Evol 25:643–652. https://doi.org/10.1016/j.tree.2010.07.009
Parry GS, Burton S, Cox B, Forman DW (2011) Diet of coastal foraging Eurasian otters (Lutra lutra L.) in Pembrokeshire south-west Wales. Eur J Wildl Res 57:485–494. https://doi.org/10.1007/s10344-010-0457-y
Prenda J, Granado-Lorencio C (1996) The relative influence of riparian habitat structure and fish availability on otter Lutra lutra L. sprainting activity in a small mediterranean catchment. Biol Conserv 76:9–15. https://doi.org/10.1016/0006-3207(95)00080-1
Prenda J, López-Nieves P, Bravo R (2001) Conservation of otter (Lutra lutra) in a Mediterranean area: the importance of habitat quality and temporal variation in water availability. Aquat Conserv Mar Freshw Ecosyst 11:343–355. https://doi.org/10.1002/aqc.454
Rabus B, Eineder M, Roth A, Bamler R (2003) The shuttle radar topography mission—a new class of digital elevation models acquired by spaceborne radar. ISPRS J Photogramm Remote Sens 57:241–262. https://doi.org/10.1016/S0924-2716(02)00124-7
Reid AJ, Carlson AK, Creed IF et al (2019) Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol Rev 94:849–873. https://doi.org/10.1111/brv.12480
Riesco M, Delibes M, Calzada J et al (2020) Desert otters: Distribution, habitat use and feeding ecology in arid rivers of Morocco. J Arid Environ 178:104165. https://doi.org/10.1016/j.jaridenv.2020.104165
Roos A, Loy A, de Silva P, et al (2015) Lutra lutra. The IUCN red list of threatened species. https://doi.org/10.2305/IUCN.UK.2015-2.RLTS.T12419A21935287.en
Tahiri L, Bennasser L, Idrissi L et al (2005) Metal contamination of Mytilus galloprovincialis and sediments in the Bouregreg Estuary (Morocco). Water Qual Res J Canada 40:111–119. https://doi.org/10.2166/wqrj.2005.011
Watters GT (1992) Unionids, fishes, and the species-area curve. J Biogeogr 19:481–490. https://doi.org/10.2307/2845767
Williams CK, Ives AR, Applegate RD (2003) Population dynamics across geographical ranges: time-series analyses of three small game species. Ecology 84:2654–2667. https://doi.org/10.1890/03-0038
World Bank (2014) World development indicators: Morocco. Available at https://databank.worldbank.org/source/world-development-indicators. Accessed 11 Sep 2014
Yackulic CB, Sanderson EW, Uriarte M (2011) Anthropogenic and environmental drivers of modern range loss in large mammals. Proc Natl Acad Sci USA 108:4024–4029. https://doi.org/10.1073/pnas.1015097108
Acknowledgements
We acknowledge the help of our colleagues in conducting the field sampling: Begoña Adrados, Seila Alvaré, Manuela González, Carlos Gutiérrez-Expósito, Miguel Jácome, Francisco Palomares, Carli Pérez, Eloy Revilla, Jacinto Román and Pablo Villalba, and the scientific advice of Abdeljebbar Qninba and his help in processing the permits necessary to conduct the study. We thank three anonymous referees for their constructive comments on an earlier manuscript. The picture of an otter used in Figure 4 was donated by Juan Matutano. Sampling permissions were obtained from the Haut Commissariat aux Eaux et Forêts et a la Lutte contre la Désertification du Royaume du Maroc. Financial support was received from Estación Biológica de Doñana-CSIC. N.F. acknowledges the support of iDiv funded by the German Research Foundation (DFG–FZT 118, 202548816). Funding for Open Access charge: Universidad de Huelva / CBUA.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. Partial financial support was received from Estación Biológica de Doñana, CSIC. N.F. acknowledges the support of iDiv funded by the German Research Foundation (DFG–FZT 118, 202548816).
Author information
Authors and Affiliations
Contributions
Conceptualization: JC, MC, MD, NF. Methodology: JC, MC, MD, NF. Investigation: JC, MC, MD, NF. Writing—Review & Editing: JC, MC, MD, NF. Formal analyses: NF, MC.
Corresponding author
Ethics declarations
Conflict of interest
Authors declare no conflict of interest.
Ethical approval
Not applicable.
Consent to participate
All authors have approved the current contents of the manuscript and its submission to Biodiversity and Conservation.
Consent for publication
All authors have approved the current contents of the manuscript and its submission to Biodiversity and Conservation.
Additional information
Communicated by Karen E. Hodges.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Calzada, J., Clavero, M., Delibes, M. et al. Human pressures constrain Eurasian otter occurrence in semiarid Northern Africa. Biodivers Conserv 31, 1519–1533 (2022). https://doi.org/10.1007/s10531-022-02405-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-022-02405-w