
Abstract
The monk seal is the most endangered pinniped worldwide and the only one found in the Mediterranean, where its distribution and abundance have suffered a drastic decline in the last few decades. The limited understanding of the population demographics and conservation status of this species are due to both its rarity and evasiveness, with records biased towards occasional, mostly coastal, encounters. Current molecular techniques allow us to detect DNA traces released into the environment (eDNA) by any organism. We have developed three species-specific qPCR-assays targeting the 12S/16S rRNA mitogenome regions of the monk seal. The assays have been tested extensively on a comprehensive and diverse set of samples (n = 73), including positive controls from a breeding population in Madeira collected during their peak abundance, and two opportunistic collections of Mediterranean eDNA samples (offshore/coastal) from on-going projects. Monk seal DNA was detected in 47.2% of the samples collected from a ferry platform in the Tyrrhenian (2018–2019) and in 66.7% of those obtained in the Pelagie archipelago in the Strait of Sicily (2020). These findings anticipated subsequent visual observations in the proximity of these sampling areas by up to 2 years. At the Tyrrhenian site, monk seal detection increased between 2018 and 2019. The demonstrated approach provides a non-invasive and highly sensitive tool for defining the monk seal actual distribution and home range -enabling monitoring also in nocturnal hours-, for assessing its recovery rate and pinpointing coastal/offshore sites where prioritizing conservation, research, citizen-science, and educational opportunities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Mediterranean monk seal (Monachus monachus) is the only pinniped permanently resident in the Mediterranean basin. Originally, Mediterranean monk seals were commonly found throughout the Mediterranean, Marmara and Black Seas, stretching out into the Atlantic Ocean along the West African coast as far south as Cap Blanc, as well as around Cape Verde, Canary, Madeira and Azores Islands. In the twentieth century, these populations have dramatically fallen in size, primarily due to deliberate killing mainly by fishermen but also, due to accidental entanglement in fishing gears. Population declines over most of the Mediterranean (virtually disappearing from Egypt, Israel, Lebanon, Montenegro, the Black Sea, Italy, Corsica, France, the Balearic Islands, Spain, Tunisia and Morocco), have led to a severe population bottleneck and signatures of inbreeding depression in the refugial populations in the eastern Mediterranean (Stoffel et al. 2018; Karamanlidis et al. 2021a). However, occasional sightings in Italian coastal waters have been recorded in the last decade (Supplementary Fig. S1).
The current distribution of the Mediterranean monk seal is still only partially defined, relying on opportunistic data from sporadic (mostly coastal) sightings, some of which remain unverified. Much of the data available are retrieved from sites where adult females are known to return every year to reproduce. Thanks to the predictability of these annual events, some of the pupping caves are monitored throughout the year using camera traps (e.g. Martinez-Jauregui et al. 2012), which have provided much information about mother/pup interactions (e.g. Karamanlidis et al. 2021b). However, these marine mammals have an extended home range as adults are known to travel several tens of kilometres per day (Adamantopoulou et al. 2011). In addition, little is known about the foraging areas and social interactions, or their actual distribution in the non-breeding season. This kind of information is difficult to retrieve as monk seals are often elusive and particularly hard-to-study in offshore waters and/or during the night.
Recent advances in molecular genetic tools have opened the way to sampling and sequencing DNA from environmental samples (e.g. soil, water, air; Taberlet et al. 2012), providing a unique, sensitive and non-invasive tool for biomonitoring that circumvents the need to directly sample or even sight living organisms (Bohmann et al. 2014). Species identification by eDNA analysis relies on the amplification of taxonomically diagnostic sequences, employing pairs of specific synthetic oligonucleotides (primers). Many broad-range primer sets are currently available that can simultaneously target DNA from entire taxonomic groups (metabarcoding, e.g. for fish Miya et al. 2015). However, metabarcoding is not so efficient in the detection of rare taxa (e.g. endangered species, first appearance of invasive organisms, etc.), whose weak signals tend to be swamped, and therefore masked, by those of more abundant taxa (e.g. Stoeckle et al. 2020). In these instances, higher resolution is guaranteed by the selective amplification of the target taxon using tailored species-specific primers (barcoding).
The Mediterranean monk seal represents an ideal species to be monitored through this novel approach, due to an urgent need to fill data gaps about population demographics and because of its characteristics and habits (a large marine mammal resident of the water surface for breathing, and shores for resting and breeding) guarantee that biological material will be shed into its environment, including easily accessible superficial waters. The present study aims to: (1) identify a suitable set of specific barcode primer-pairs suitable to detect the Mediterranean monk seal mitogenome; (2) validate the assays’ performance on a suite of control samples; (3) test the efficiency of this approach on marine eDNA samples available from on-going projects in areas where the presence of Mediterranean monk seal cannot be ruled out (Tyrrhenian Sea and Strait of Sicily).
Materials and methods
Design of species-specific qPCR assays
To design monk seal specific primers we searched for candidate sequence strings within the mtDNA regions targeted by the three MarVer primer sets (Valsecchi et al. 2020), as highly variable. The 12SrDNA and 16SrDNA fragments targeted by the three primer sets -MarVer1, MarVer2 and MarVer3- were all highly polymorphic, as also evident when aligning the homologous regions in two sister species of the Mediterranean (Monachus monachus) and the Hawaiian (Neomonachus schauinslandi) monk seals (GenBankANs NC_044972 and NC_008421 respectively). In the fragments targeted by the MarVer1, MarVer2 and MarVer3 primer sets, 12 (out of 199 bp), 14 (out of 87 bp) and 24 (out of 237 bp) variable sites were found, respectively. For each amplicon, one monk-seal specific primer was designed from the internal sequence that had the largest number of diagnostic sites. Each monk-seal specific primer was paired for amplification to the corresponding MarVer universal primer on the opposite strand (Valsecchi et al. 2020). In those instances where the universal primer had one degenerate base, the mammalian variant was used (Valsecchi et al. 2020). Another prerequisite in the search for adequate primer sites was that the oligos’ pairs designed should amplify fragments of different lengths in order to: a) verify whether amplicon size matters when dealing with degraded eDNA templates; and b) test the possibility of multiplexing barcodes (see below).
Hereafter we use the terms “MarVer1”, “MarVer2” and “MarVer3” to refer to the three specific loci rather than the broad-range primer sets described previously (Valsecchi et al. 2020), while MmoMV1, MmoMV2 and MmoMV3 refer to the three new corresponding assays based on the internal monk-seal specific primers. Before carrying out the qPCR screening, the three candidate primer sets were tested for amplification using a traditional PCR approach, both individually and in pairs/triplet (multiplex).
Sample set
The assays were tested on a wide range of DNA templates, including tissue (n = 3) and residual biological—hair, faeces, food and regurgitated remains—extracts (n=12), and marine eDNA samples (n = 58). Samples (n = 73) were assigned to one of 7 categories (Table 1 and Supplementary Box S1): three kinds of positive controls, two negative controls and two trial sets of Mediterranean marine eDNA (“opportunistic” samples available from other ongoing research projects).
Category 1 DNA positive control. Mediterranean monk seal DNA extracted from tissue (spleen).
Category 2 DNA-traces positive control. DNA extracts from 12 environmental residual samples (hair, faeces, food and regurgitated remains).
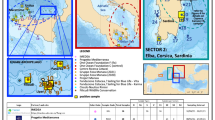
Category 3 eDNA positive control. This group included 8 samples, all collected in Madeira archipelago, where an isolated population of Mediterranean monk seals (estimated 20–30 individuals) is resident and has been protected and monitored for over 30 years (Pires et al. 2008). In the Madeira archipelago most monk seals are found in the waters surrounding Desertas Islands (37 km of coastline), that become a Nature Reserve in 1990 and where seals are constantly monitored thanks to a capillary system of several automatic cameras (taking one shot per hour) placed in the areas attended by seals. One of the eight samples was collected off the north-eastern shore of Madeira main island, while the remaining seven were collected along a 5 km-stretch of the coast of Deserta Grande Island, including the opening of a pupping cave (Tabaqueiros cave) inhabited by 8 seals at the time of sampling, as witnessed by the camera-surveillance system (Fig. 1). We opted to sample in November, when the breeding season is at its peak and the number of new born pups is maximal (Pires et al. 2008), thereby ensuring intense seal activity proximal to coastal sheltered shores and caves. The 8 seawater samples (each of 8–12 L) were collected at no more than 120 m from shore, during the 2020 breeding season, on the 10th and 12th of November (Supplementary Table S1). The 8 samples were purposely collected at points with a diversified incidence of seal presence, ranging from areas with no seal activity recorded at the time or before sampling (days to weeks) to points proximal to the entrance of caves currently inhabited by seals (Figs. 1 and 3). Water samples were collected by rangers of the National Park Service of Madeira and the expected likelihood of finding molecular signs of monk seals was initially not shared with the molecular team in order to ensure an unbiased analysis. Moreover, all 7 water samples gathered off Desertas were collected almost simultaneously, within an hour (2–3 pm, on the 12th November, 2020). This sampling protocol aimed to produce a “molecular snapshot” of the surveyed area, in order to minimize both the effect due to animals moving around and the inherent variability found in temporal eDNA replicates (Beentjes et al. 2019). The expected-positive eDNA control samples were not only used to assess the detection capability of the assays developed (quality control) but also, as term of comparison of the intensity of the molecular signal (quantitative control) when evaluating surveillance eDNA samples.

Monk seal presence in positive control samples. Picture taken on the date and hour of sampling by the autonomous surveillance and monitoring fix-camera system, showing the presence of 5 resting seals on the Tabaqueiro beach (Deserta Grande Island, Madeira archipelago). Sample Mmo + 01 was collected at sea, 70 m from the point shown in this image. Other seals were photographed within the Tabaqueiro cave (30 m from Mmo + 01 sampling point), but the imagine is not as clear as this one. The monitoring system has been installed as part of the Life13 NAT/ES/000974 Madeira Monk Seal Project
We also tested the molecular assays on two Mediterranean marine eDNA samples (Categories 4 and 5), collected as part of previous (Valsecchi et al. 2021) and ongoing projects, from areas where monk seals are considered to be extremely rare but not impossible to be found, as suggested by a few recent sightings (Supplementary Figure S1) some of which were proximal to our sampling sites, but the eDNA samples had been collected month to years previously. For each sampling event, sampling time was recorded (Supplementary Table S1): 20 (41.7%) out of 48 samples were collected during night-time.
Category 4 Mediterranean coastal eDNA samples. This subset of samples was comprised of ten eDNA samples collected between the 14th August and 25th September 2020 as part of an ongoing project from waters surrounding the Pelagie Islands, and two additional samples collected from the shore in the days following reported monk seal sightings in the proximity of the sighting point in the waters of Lampedusa island (22nd and 31st October, and 9th November, 2020: Supplementary Figure S1). These samples were used to relate the intensity of the signal to the actual presence of seals (for more details about these samples see Supplementary Table S1 and Box S1).
Category 5 Mediterranean offshore eDNA samples. These samples were collected in the summers of 2018 (n = 16) and 2019 (n = 20) from the Northern Tyrrhenian Sea as part of an ongoing project (“MeD for Med”) from marine water samples collected from operating ferries (Valsecchi et al. 2021). The route surveyed was the Livorno-Golfo Aranci run by Corsica Sardinia Ferries. Unfortunately, no 2020 sample was available due to the restrictions imposed by the COVID pandemic. The sampling involved collecting water samples from fixed stations that remained constant over the different cruises and that also included nocturnal samples. Additional samples were gathered on occasion of sightings of cetaceans operated by members of the ISPRA FLT network (Arcangeli et al. 2017). Nineteen (52.8%) of the 36 ferry samples were collected at night time.
A dual set of negative controls was included in our test both at the DNA extraction (sealed filter) and qPCR amplification (no DNA) steps. Furthermore, two classes of DNA negative controls (tissue-extracted DNA from non-target species and eDNA from marine districts where the monk seal is not found) were included:
Category 6 Tissue-extracted negative DNA controls. Since no closely related taxa to the target species are found in the Mediterranean (e.g. the Hawaiian monk seal or any other Phocidae), and considering that only one of the two PCR primers for each locus is monk-seal specific, while the other is conserved across all vertebrates, we selected negative control samples from vertebrates to be run alongside non-eDNA samples (i.e. categories 1 and 2). These negative controls were DNA extracted from two widespread vertebrate taxa in the Mediterranean, a cetacean (the bottlenose dolphin Tursiops truncatus) and a fish (the yellowfin tuna Thunnus albacares).
Category 7 Non-Mediterranean eDNA negative controls. Similarly, eDNA samples from an ongoing international projects (Maldives and Hawaii) were employed as M. monachus negative controls for environmental samples (Categories 3, 4 and 5).
DNA extraction
DNA from the tissue samples (Categories 1 and 6) was extracted using the Qiagen DNeasy® Blood and Tissue Kit, while the DNA in biological residues and environmental samples (Categories 2, 3, 4, 5 and 7) was extracted using the Qiagen DNeasy® PowerSoil® Kit, according to the manufacturer’s instructions. Both extraction protocols were carried out having cleaned the bench and equipment with bleach before each sample was processed, and blank extractions (no tissue or unused filter) were included in each extraction. When biological residual samples (MmoR, Category 2) were in liquid form (e.g., faeces stored in ethanol), an aliquot was filtered through a membrane that was subsequently rinsed with distilled water prior to DNA extraction. Tissue DNA was quantified using NanoDrop 2000 (ThermoFisher Scientific).
eDNA sample collection and processing
For each sampling station 8–12 L of seawater were collected and stored in the Bag-in-Box Sampling System (BiBSS: Valsecchi et al. 2021). Once in the laboratory (or on board the ferry for Category 5 samples), the contents of the water bags were aliquoted and filtered through 3 cellulose nitrate membranes using the Sartorius Monitor Biosart™ 100, thereby us producing 3 sample replicates for each eDNA sample collected (filters A, B and C).
The four sets of eDNA samples (Categories 3 to 5 and 7) that came from different projects, were not always homogeneous in terms of the amount of water processed and the porosity of the filters (see Supplementary Table S1). However, they were all collected from the sea surface, with the exception of the ferry samples collected from sea water intake ports 4.5 m below sea level.
All laboratory procedures were carried out in separate rooms for the pre- and post- amplification steps, with dedicated personal protective equipment. Tissue-extracted DNA and eDNA extractions were carried out in the same lab, but in temporally distant (months to years apart) events: for example, the ferry eDNA samples (Category 5) were processed and extracted long before the monk seal tissue sample arrived at our laboratory. Moreover, before commencing this study our research group had not carried out any projects on pinnipeds, reducing the risk of cross contamination. All qPCR reactions were set-up following strict protocols in a designated clean laboratory, under a UV-sterilised laminar flow cabinet, in a facility located apart from the extraction laboratory.
The two samples collected directly from shore (MmoMc11 and MnoMc12, Category 4) underwent a tenfold template dilution prior to amplification, a procedure used to minimize any interference of PCR inhibitors (Gasparini et al. 2020; McKee et al. 2015; Wang et al. 2017) known to be particularly abundant in the proximity of sediments (Lance and Guan, 2019).
Experimental design
The three species-specific assays were first tested using traditional PCR on a subset of samples that included tissue-extracted DNA, to verify successful amplification, the size of the fragments amplified and the possibility to multiplexing the three loci. The PCR temperature cycling conditions used were: an initial denaturation at 95 °C for 3 min; 40 cycles of denaturation at 94 °C for 30 s, annealing at 56 °C for 30 s and elongation at 72 °C for 30 s; the final cycle was followed by extension at 72 °C for 7 min.
Quantitative Real Time PCR (qPCR) assays were performed on an AB 7500 (Applied Biosystem) to test the three primer pairs. First, each primer pair was used on M. monachus tissue samples to determine the amplification efficiency (E), limit of detection (LOD) and limit of quantification (LOQ), according to Bustin et al. 2009 and Klymus et al. 2019. To generate the standard curve, tenfold serial dilutions of quantified positive tissue controls were used. All 73 samples (tissue, residual and eDNA) were run in triplicate, using the following qPCR conditions: an initial denaturation at 95 °C for 10 min; 40 cycles of denaturation at 95 °C for 15 s and annealing-elongation at 56 °C for 1 min; and a final dissociation stage. The amplification reaction contained 5.0 μl SsoFast™ EvaGreen® Supermix with Low ROX (Bio-Rad), 0.1 μl of each [10 μM] primer solution, 2 μl DNA and 2.8 μl of Milli-Q water. For each run, positive controls, negative tissue/eDNA controls, and no-template negative controls were also included in triplicate. The Ct (Cycle threshold) values were converted into counts (DNA copies: Bruno et al. 2017). When the qPCR copy number output was below the LOQ but above the theoretical qPCR limit (three copies per reaction according to Bustin et al. 2009), they were considered as ‘detectable but not quantifiable’ (DBNQ). For all the reactions, primer specificity was verified by the calculated melting temperature (Tm).
Testing the correlation between amplicon size and DNA degradation
Since the primer set producing the larger amplicon (MarVer3, 216 bp) was found to perform less efficiently than the other two sets (see Results), we tested whether the signals obtained from a high-quality DNA sample and a degraded DNA sample (as eDNA) were similar for all three loci, regardless of fragment size. This was possible thanks to the availability of two exceptional samples: (1) a high molecular weight DNA sample, MmoT01, freshly extracted from tissue collected immediately post-mortem; and (2) a high quality/quantity monk seal eDNA sample, MmoM + 01, collected at a site with a high concentration of monk seal at the time of sampling. This sample was collected at a site characterized as proximal to a cave contextually inhabited by many seals and where there was limited mixing with surrounding waters (reducing signal dispersion) due to a counter-current that pushed the water towards the opening of the cave (see Discussion). All three loci produced a strong signal in both samples and all replicates, allowing the correlation between amplicon size and DNA quality to be tested. We applied a Kruskal–Wallis test that did not require the data to be normally distributed and similar levels of variance.
Results
Primer sets
At each of the three MarVer loci (Valsecchi et al. 2020), one Monachus monachus specific primer site was identified that fulfilled the specificity criteria established for the target species (Table 2). In order to increase specificity, the three species-specific primers were designed slightly longer than the universal MarVer primers used in each primer set. As a result, the three species-specific priming sites were well differentiated between the Mediterranean monk seal (GenBankAN: NC044972) and its closest relative, the Hawaiian monk seal (GenBankAN: NC_008421). The number of variable bases at the priming sites were: 6 for primer MmoMV1F (24%), 9 for primer MmoMV2R (32.1%) and 8 for primer MmoMV3F (22.2%), in the MarVer1, MarVer2 and MarVer3 regions respectively (Table 2). The sequences of three monk-seal specific oligos were fully conserved (i.e., invariable) among the five complete mitogenomes deposited in GenBank (Accession Numbers MG570471- MG570475) by Gaubert et al. (2019), covering all extant and historical haplotypes described for this species. All three assays successfully amplified PCR products of the expected size and could be multiplexed (Figure S4). These primer sequences have been deposited in the BOLD System repository (Ratnasingham and Hebert, 2007).
qPCR outcomes
A total of 1860 qPCR reactions were run on the 73 samples attributed to the seven sample Categories (Table 1). The amplification efficiency of the MarVer1 primers was 92.6%, with a LOQ that corresponded to a Ct = 34.75 (16 DNA counts/µL of reaction). The MarVer2 amplification efficiency was 100.7%, with a LOQ that corresponded to Ct = 35.8 (9 DNA counts/µL of reaction), while the MarVer3 primers had an amplification efficiency that reached 92.2% and a LOQ that corresponded to a Ct = 34 (25 DNA counts/µL of reaction) (R2 ≥ 0.99 for each assay). The TMs were 75.4 ± 0.3 for MarVer1, 71.5 ± 0.3 for MarVer2, and 78.5 ± 0.2 °C for MarVer3. Based on the LOQ calculated for each locus, the qPCR DNA detection outcomes were divided in three classes: (1) no signal; (2) monk-seal DNA detectable but not quantifiable (DBNQ); or (3) positive quantifiable detection (PQD). Of the 73 samples screened, 26 were classified as PQD for at least one of the three markers, of which 19 (90.5%) were from the positive control group (n = 21) and 7 (14.6%) from the trial eDNA samples (n = 48; Fig. 2 and Supplementary Table S1). In addition, there were a total of 19 DBNQ positive reactions for at least one of the three loci, of which only 1 (4.7%) corresponded to the positive control group, with the remaining 18 (37.5%) corresponding to the Mediterranean trial eDNA samples: 2 from the Strait of Sicily and 16 along the Northern Tyrrhenian ferry track. In 28 samples (38.4%) there was no monk seal DNA signal, these including one positive control (eDNA), MmoM + 02 sample, and all four negative controls (sample Categories 6 and 7). The remaining 23 samples were Mediterranean eDNA samples, namely 19 (52.7%) from the Tyrrhenian route and 4 (33.3%) from the Strait of Sicily.

Positive quantifiable detections (PQDs) found in seven exploratory Mediterranean eDNA samples, shown in relation to positive control samples. The values are expressed in Log2 DNA-copies, quantified per litre of marine water in the case of eDNA samples. Note that Detectable But Not Quantifiable (DBNQ) signals are not shown here but they are detailed in Table 1S and Fig. 2S
Only positive control samples (Categories 1, 2 and 3) produced a positive signal for monk seal DNA for all three loci (and experimental replicates). Most eDNA samples, including 5 of the 8 positive-control eDNA samples from Madeira (Category 3), generated a positive result for monk seal DNA above the LOD for only one or two of the three loci (Fig. 2, Supplementary Table S1). The MarVer1 locus detected monk seal DNA in most samples (n = 22), followed by MarVer2 (n = 20). The MarVer3 16SrDNA primer set returned the lowest number of PQDs (n = 12), all of which were found in the positive control samples and only one of which was an eDNA sample (MmoM + 01). The complete list of Log2 DNA copies recovered in the whole data set, for all three sample and experimental replicates and markers, is shown in Supplementary Fig. S2.
We considered the results obtained in terms of the categories to which the samples were assigned.
Positive-control samples (Categories 1 to 3)
Category 1
DNA was extracted from Mediterranean monk seal tissue (MmoT1) at a concentration of 284 ng/μl (NanoDrop 2000, ThermoFisher Scientific) and used as reference sample. All three primer sets produced amplicons of the expected size in all 3 reaction replicates with this DNA, allowing detection up to 5.7 × 10–8 mg/L.
Category 2
All 12 DNA extracts from the biological residues (MmoR samples in Table 1 and Supplementary Table S1) produced at least one PQD in each of the assays, with 11 PQDs in all three replicates with MarVer1, 10 with MarVer2 and 8 with MarVer3. Thus, MarVer1 produced more overall PQDs from Category 2 samples (34 out or 36) than the other two loci (31/36 for MarVer2 and 27/36 for MarVer3), although the number of estimated DNA copies was highest for MarVer2.
Category 3
The results obtained with the expected positive control eDNA samples collected off the Madeira archipelago and related to the field observations reported by the rangers at the time of the water sample collection are shown in Fig. 3. Sample MmoM + 01, was collected at the entrance of the Tabaqueiro cave (used by approximately 8 seals) and it was the control eDNA sample that produced the strongest signal in all three sample replicates, for all the experimental triplicates and for all three markers (Supplementary Table S1, Fig. 2 and Supplementary Fig. S2).

Map of Madeira eDNA positive-control samples (Category 3) and field annotations. The eight sampling sites are indicated with color-coded circles according to which monk seal DNA marker produced positive quantifiable detections (PQDs). The table at the bottom shows the field notes for each sampling site as reported by the rangers at the moment of sample collection, and released to the molecular team only when the genetic analysis was completed
Mediterranean eDNA sample sets (Categories 4 and 5)
In the Mediterranean eDNA samples PQD (n = 7) and DBNQ (n = 18) monk seal DNA was detected in the samples from both sample Categories 4 and 5 (Fig. 4). Monk seal eDNA was recovered in 47.2% of the ferry samples (Tyrrhenian, n = 36) and in 66.7% of the coastal samples (Strait of Sicily, n = 12). The proportion of PQR and DBNQ detection was much higher in the Pelagian samples (6 PQD and 2 DBNQ detections) than in the ferry samples (1 PQD and 16 DBNQ detections). Moreover, excluding the two samples collected on the days following sightings (MmoMc11 and MmoMc12), Pelagian monk seal DNA recovery (n = 6) was more frequent in waters far from the main island of Lampedusa (83.3%, n = 5), 4 of which were proximal to Lampione, rather than in the waters surrounding Lampedusa itself (n = 1). As for the ferry samples, monk seal DNA was detected more commonly at night (64.7%, n = 11) than in samples collected during daylight hours (35.3%, n = 6), despite diurnal (n = 17, 47.2%) and nocturnal (n = 19, 52.8%) ferry samples were roughly equally represented.

Calendar of sample collection and monk seal sightings, along with the molecular results in the two Mediterranean areas tested for which eDNA samples were available from on-going projects: offshore Tyrrhenian (left) and coastal Strait of Sicily, Pelagie archipelago (right). In the tables, the black squares indicate positive and quantifiable monk seal DNA detections (PQD) at one or more loci (red symbols in the maps). The grey squares show samples where weak (below the detection threshold) monk-seal signals (yellow symbols in the maps), defined in the text as DBNQ (detectable but not quantifiable). The number within the black/grey squares reflects the number of sample replicates with a positive detection. Coloured cells indicate sampling events, with darker shading reflecting nocturnal samples. The positions and dates of recent monk seal sightings in the two study areas are reported (in red in the calendar tables, asterisks in the maps)
Negative-control samples (Categories 6 and 7)
The negative controls for the tissue and eDNA samples both produced no signals in all qPCR runs.
A significant difference was found between the three loci in terms of the drop of intensity in the molecular signal (ΔLog2 DNA-copies) from the tissue (MmoT01) to the eDNA (MmoM + 01) control samples (Kruskal–Wallis test: χ2 = 13.18, df = 2, p-value = 0.001374). The major difference was associated with the MarVer3 locus, which showed a significantly stronger drop in the signal than the two 12SrDNA markers, which both displayed a similar decrease in their signal (Supplementary Fig. S3).
Discussion
There is widespread concern about the fate of the Mediterranean monk seal (Saydam 2018). Yet, despite concerted efforts over the last few decades, many gaps remain in our understanding of the distribution of this threatened species (Mo, 2011; La Mesa et al. 2021). For such endangered and elusive organisms, eDNA analysis may represent a very effective surveillance tool as it does not rely on visual observations (Bohmann et al. 2014). In the present study we have designed three species-specific quantitative molecular assays that target the 12S (two) and 16S (one) rDNA sequences, allowing Mediterranean monk-seal DNA to be detected both from biological residuals in soil and water environmental matrices.
Our results show that the described assays successfully detect small amounts of monk seal eDNA (e.g., 5.7 × 10–8 mg/L), with the two 12S markers outperforming the 16S marker. In addition, the screening of opportunistic Mediterranean marine eDNA samples (from previous and ongoing studies) detected monk seal DNA in about 50% of the samples, evidence that eDNA analysis can detect the presence of monk seal even in the absence of visual confirmation. The approach proposed can potentially shed light on as yet unknown aspects of the habitat use of this charismatic marine mammal species, through its detection in otherwise inaccessible contexts (i.e., offshore waters and at night). Finally, the possibility of monitoring the presence of monk seals in offshore waters provides useful spatial data for identifying potential areas for conservation targets.
Overview of the assays
Specificity and sensitivity
The degree of molecular differentiation of the three monk seal specific primers relative to homologous regions in the genome of sister species, such as the Hawaiian monk seal, reflects the high specificity of the proposed assays, an assumption that was confirmed in all the positive and negative control samples. High specificity for the target species is a fundamental prerequisite for the uptake of an innovative method, particularly for monitoring the conservation status of rare species. Our experimental and trial design suggest that there is little room for false positives (except those attributable to laboratory contamination, for which maximum caution should be taken as in any experiment that is based on eDNA analysis), although perhaps the incidence of false negatives should be of more concern. Indeed, PCR inhibitors in environmental matrices have been recognized as one of the main limitations of eDNA-based detection approaches (Loeza-Quintana et al. 2020).
Coverage
The tested monk-seal assays are conserved and thus suitable for the detection of all -present and historical- haplotypes described for this species (Gaubert et al. 2019), thus they can be used across its whole distribution range.
Reliability
Samples that generated a positive result in only 1 or 2 of the three amplification replicates were considered to indicate a positive monk seal detection. This is based firstly on the high specificity of the species-specific primers to Mediterranean monk seal DNA, as supported by the specific melting temperature (Tm) recorded, suggesting that any positive detection denotes the presence of monk seal DNA. Secondly, failing to amplify DNA in replicates of a positive sample may be due to the extremely low concentration of the target molecule. Some studies suggest that marine vertebrate DNA accounts for only 0.004% of the eDNA retrieved from marine samples (Stat et al. 2017) and, although metabarcode vs barcode data are difficult to compare, it is noteworthy that in a DNA metabarcoding analysis on a subset of the same ferry samples (only from 2018), using the MarVer1 and MarVer3 universal primer pairs, over 90% of the reads were attributed to bony fish and no reads were attributed to the monk seal (Valsecchi et al. 2021). Indeed, this figure decreased more than one order of magnitude when considering the incidence of rare vertebrates like marine mammals. The use of replicates (from sampling to DNA amplification) reduces the rate of false negatives, yet the optimal level of replicates is difficult to estimate as it strongly depends on the detection probability of the target taxon.
As for eDNA expected positive samples (Category 3), with the exception of sample MmoM + 1 that was collected proximal to the entrance of a cave densely populated by monk seals (n = 8) and returned a positive result for all three loci and replicates, all the remaining eDNA samples from Madeira (n = 7, 87.5%) were positive for monk seal DNA, either in some replicates and/or for some of the loci (typically the two 12SrDNA ones), even though seals were definitely present in and around these waters, as evidenced by the continuous photographic monitoring system. In some cases, positive identification was only made in one of the sample replicates (each originating from one of the three filters used to process a single sample). This effect can be more easily explained if we consider the specific characteristics of the animal and the release of eDNA (Lacoursière‐Roussel and Deiner, 2019; Turner et al. 2014). Large animals like marine mammals release biological traces into the surrounding water that likely consists of both large clusters of cells -sloughed skin- as well as single or small groups of cells (Valsecchi et al. 1998). In this sense, when a sample is divided into two or more subsamples, the clustered cells may not be distributed homogeneously among the sub-samples. Thus, imperfect detection is an unavoidable feature of the data on the presence/absence of a species even in traditional ecological field studies when individuals and species present at a site are not always detected. Several recent studies addressed the imperfect detection issue, highlighting the role of abiotic factors, such as temperature, pH, UV (Kasai et al. 2020; Pilliod et al. 2014; Strickler et al. 2015), organic matter and DNA polymerase inhibitors (van Bochove et al. 2020); biotic factors (Tsuji et al. 2017); but also, DNA fragment size (Bylemans et al. 2018). All these factors, together with low target DNA concentration, may have a role in false negative results. Indeed, such failures to account for imperfect detection may result in biased inferences, which is particularly relevant to eDNA assays that rely on small amounts of degraded DNA. Moreover, laboratory and sequencing costs limit the replication of this data (Ficetola et al. 2015). However, the high sensitivity and specificity of the eDNA-based approaches developed can overcome these limitations.
Performance of the three barcode assays
The primer set producing the largest amplicon (locus MarVer3, 216 bp) was amplified with more difficulty and/or produced fewer DNA copies than the other two primer sets (MarVer1, 146 bp and MarVer2, 71 bp). The same pattern was observed when employing high molecular weight DNA (extracted from tissue) as the template. The rate of drop in signal between a high concentration/quality DNA sample and eDNA was significantly higher than that observed in the two markers targeting smaller fragments, suggesting that, besides eDNA being affected by factors such as inhibition, template concentration and template competition, the reduction in amplification yield is also related to eDNA integrity and thus, to the amplicon size rather than to the intrinsic characteristic of the MarVer3 locus (e.g. primer efficiency/specificity of the gene targeted). However, no difference in performance was evident when comparing 12 species-specific primer pairs that produce amplicons from the Cyt b gene ranging from 76 to 249 bp in Yangtze finless porpoise (Neophocoena asiaeorientalis asiaeorientalis: Ma et al. 2016). However, a meta-analysis of eDNA-based studies (Saito and Doi, 2021), showed that eDNA degradation was accelerated in amplicons with higher length. Thus, the hypothesis of a correlation between the drop in the DNA signal recorded in relation to DNA degradation and hence, to temporal variables (e.g. the time since DNA release into the environment), should be explored more deeply in future studies.
Our study suggests that the most informative and efficient markers are the two 12SrDNA loci (MarVer1 and MarVer2), whereas MarVer3 (16SrDNA) could be employed in the marker panel, possibly providing some evidence of the integrity/freshness of the sample/signal, especially if a correlation between amplicon size and eDNA degradation is ascertained.
Traditional PCR and assays multiplexing
The possibility of visualizing the assay outcomes by traditional PCR rather than qPCR is desirable, as it allows tests to be carried out in any structure equipped with essential molecular biology laboratory apparatus, and it is cheaper. Although the diagnostic sensitivity decreases in the case of traditional PCR, when the signal is not too weak (see Supplementary Fig. S4) even basic, cost-effective, PCR allows traces of monk seal DNA to be identified, without the necessity of relying on qPCR multiplexed assays (such as Taqman). A preliminary multiplex test indicates that all combinations of the three assays proposed are compatible with multiplexing and that the approach is even efficient with eDNA samples, providing a relatively high (say > 10 log2DNA copies/L) monk seal DNA content, such as that seen in the Madeira control sample, MmoM + 01 (Supplementary Fig. S4).
Current matters
In the field of marine biology there are many and increasing applications that rely on the information obtained from the study of eDNA (Bohmann et al. 2014; Ruppert et al. 2019; Eble et al. 2020). However, this heterogeneous complex of DNA molecules is susceptible to diverse environmental factors, such as temperature (and hence seasonal variation) and microhabitat heterogeneity (Troth et al. 2021), as well as vertical zonation patterns (Jeunen et al. 2020; Moyer et al. 2014), meteorological conditions, biotic/abiotic factors and spatiotemporal dynamics that determine the presence, persistence and fate of eDNA (Collins et al. 2018; Stewart 2019).
Noteworthy, in the present study we found discordant signals from two sampling sites expected to be positive for M. monachus eDNA detection. Indeed, Madeira samples from the stations 1 (MmoM + 01) and 2 (MmoM + 02) are only 112 m apart, and seawater samples were collected at the two spots just 5 min apart. However, in the first case (30 m from the entrance of the cave frequented by > 8 seals) the molecular signal was the strongest of all our eDNA samples, whereas in the latter no signal was found in 27 reactions (3 filters, 3 replicates, 3 markers). Thus, along this 112 m transect some physical factor, possibly a current barrier, seems to prevent the two water masses from mixing (Supplementary Fig. S5). This fortuitous circumstance suggests including currents among the factors that should be considered when collecting water samples for eDNA-based assays. Conversely, it adds reassurance that when a signal is indeed intercepted it probably reflects the presence of the animal over a limited spatio-temporal range, as signals that disperse with currents are rapidly diluted and thus remain undetected.
Detection of monk seal in Mediterranean eDNA samples prior visual observation
Monk seal DNA was detected in both the Tyrrhenian and Strait of Sicily samples, both marine areas where monk seals are occasionally encountered, although DNA was more often recovered in the Pelagian (especially around Lampione Islet) than in the ferry samples. In the last two decades, monk seal sightings have been rare, yet steadily more common in waters surrounding the Italian peninsula (Supplementary Fig. S1). According to the published data, no sighting was recorded in 2019 compared to at least 9 events recorded in 2020. Although these opportunistic data must be taken with due caution, they suggest an increase in the occurrence of monk seals in Italian waters. The same trend, albeit anticipated by a year, was found in our two-year spanning Tyrrhenian sample. These observations are consistent with the increased sighting/pupping rate recently recorded in Cyprus (Nicolaou et al. 2019), which may reflect a westward species expansion.
The strongest recovery of monk seal DNA in the ferry-route eDNA was found in sample MmoMo17 (sample 19-1LiGA1), collected on the 20th June, 2019 approximately 11 km from Elba Island and 26 km from Capraia Island, where monk seals were sighted at least twice in summer 2020 (June/July). At the same site, exactly one year previously (18th of June 2018), a detectable but non-quantifiable signal was identified, providing tentative evidence of the presence of monk seals in the Tuscany archipelagos two years prior to the recent de visu identification in the waters surrounding Capraia Island (June-July 2020).
In the Pelagian data, both samples related to sightings (MmoMc09, and MmoMc11) gave the strongest detection signals. Sample MmoMc09 was collected in the northern shore of Lampedusa, close to the location of two consecutive sightings two months later. By contrast, sample MmoMc11 was collected two days after a sighting occurred nearby. In this case, the signal was the strongest within the Pelagian data, at least for MarVer2, and comparable to the signal obtained in the control eDNA from Madeira MmoM + 01.
Relevance of offshore and nocturnal samples
Although recognized as the most endangered pinniped species worldwide (Karamanlidis and Dendrinos, 2015), the status of the Mediterranean monk seal population is currently considered “data-deficient”, relying only on a partial view of its distribution range due to the lack of specific large-scale spatial studies (La Mesa et al. 2021). Indeed, to date the study of this species has focused on resting/reproductive coastal areas where paradoxically, research is necessarily limited by the concern of disturbing such a vulnerable species, hence the use of camera traps. Yet, despite the aforementioned constraints, limiting the study of the monk seals to their coastal occurrence would inevitably restrict and bias our knowledge of the species, as it only offers insight into limited aspects of its life cycle. This study provides the first example of the molecular detection of monk seals based on seawater eDNA analysis and also represents the first case where monk seals are looked for in open waters at any time of day, allowing to assess diel variations in habitat and home range use. Ferry-based eDNA sampling allows to focus on the structure of offshore biological communities. For example, in our ferry sample (Category 5) the points sampled outside the continental shelf are the majority (22 out of 36) and homogeneously distributed between nocturnal and diurnal samples. It is interesting to note, within offshore samples, the prevalence of signals (albeit weak) attributable to the presence of monk seals in nocturnal samples. According to an eDNA metabarcoding study (Valsecchi et al. 2021) carried out on a subset of Category 5 samples targeting vertebrates’ MarVer1 and MarVer3 regions, vertebrate reads (> 95% of which are attributable to bony fish) are significantly higher in nocturnal as opposed to diurnal samples, thus suggesting prey abundance. Future offshore screenings will allow to confirm or reject the hypothesis that monk seals are likely to frequent deep waters mostly at night -probably foraging-, while they have more coastal habits during day time.
The relevance of molecular detection to monk seal conservation
There are at least three circumstances in which molecular monitoring via species specific molecular assays (such as those described here) represent a valid approach: (1) for regular monitoring of priority conservation areas or other defined areas (e.g. near breeding/pupping sites or expected underwater resting/sleeping sites); (2) to validate observations outside monitoring areas; and (3) to investigate range expansions as well as exploring yet unstudied facets of the biology of this threatened species, such as habitat use, feeding habits and movements during non-reproductive seasons, with a particular focus on offshore waters. The main applications of the molecular approach for monk seal detection are summarized in Fig. 5.

Possible applications of monk-seal molecular monitoring (photo credit Emanuele Coppola)
In conclusion, three molecular assays tailored for the detection of monk seal DNA were validated on three sets of control samples, including eDNA samples collected during the 2020 monk seal breeding season off Madeira. The eDNA-based assays proved to successfully detect the presence of this threatened species in both coastal and offshore Mediterranean eDNA samples. Regular and well-designed eDNA sampling shows promise for detecting the presence of monk seals in areas not routinely covered by visual observations. Not only could this approach represent an innovative and non-invasive means to monitor the conservation status of this unique pinniped but also, it has the potential to highlight unstudied aspects of their behaviour, especially in offshore and/or nocturnal contexts. Overall, the data presented encourages the study of eDNA as a molecular surveillance tool to be coupled with traditional ecological surveys in order to gather more accurate predictions of species distribution, leading to potentially more effective marine conservation planning.
Availability of data and material
Should the manuscript be accepted for publication, the data supporting the results will be archived in an appropriate repository (Dryad, Figshare or Hal) and the data DOI will be included at the end of the article.
References
Adamantopoulou S, Androukaki E, Dendrinos P, Kotomas S, Paravas V, Psaradellis M, Tounta E, Karamanlidis AA (2011) Movements of Mediterranean Monk Seals (Monachus monachus) in the Eastern Mediterranean Sea. Aquat Mamm 37:256–261. https://doi.org/10.1578/AM.37.3.2011.256
Arcangeli A, Campana I, Bologna MA (2017) Influence of seasonality on cetacean diversity, abundance, distribution and habitat use in the western Mediterranean Sea: implications for conservation. Aquat Conserv Mar Freshw Ecosyst 27:995–1010. https://doi.org/10.1002/aqc.2758
Bohmann K, Evans A, Gilbert MTP, Carvalho GR, Creer S, Knapp M, Yu DW, de Bruyn M (2014) Environmental DNA for wildlife biology and biodiversity monitoring. Trends Ecol Evol 29:358–367. https://doi.org/10.1016/j.tree.2014.04.003
Bruno A, Sandionigi A, Galimberti A, Siani E, Labra M, Cocuzza C, Casiraghi M (2017) One step forward for the routine use of high-throughput DNA sequencing in environmental monitoring. An efficient and standardizable method to maximize the detection of environmental bacteria. Microb Open 6:e00421. https://doi.org/10.1002/mbo3.421
Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL, Vandesompele J, Wittwer CT (2009) The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem 55:611–622. https://doi.org/10.1373/clinchem.2008.112797
Bylemans J, Furlan EM, Gleeson DM, Hardy CM, Duncan RP (2018) Does size matter? An experimental evaluation of the relative abundance and decay rates of aquatic environmental DNA. Environ Sci Technol 52(11):6408–6416. https://doi.org/10.1021/acs.est.8b01071
Collins RA, Wangensteen OS, O’Gorman EJ, Mariani S, Sims DW, Genner MJ (2018) Persistence of environmental DNA in marine systems. Commun Biol 1:1–11. https://doi.org/10.1038/s42003-018-0192-6
Eble JA, Daly-Engel TS, DiBattista JD, Koziol A, Gaither MR (2020) Marine environmental DNA: approaches, applications, and opportunities. In: Sheppard C (ed) Advances in marine biology, 1st edn, vol 86, issue 1. Academic Press, New York, pp 141–169
Ellison SL, English CA, Burns MJ, Keer JT (2006) Routes to improving the reliability of low level DNA analysis using real-time PCR. BMC Biotechnol 6:1–11. https://doi.org/10.1186/1472-6750-6-33
Ficetola GF, Pansu J, Bonin A, Coissac E, Giguet-Covex C, De Barba M, Taberlet P (2015) Replication levels, false presences and the estimation of the presence/absence from eDNA metabarcoding data. Mol Ecol Resour 15:543–556. https://doi.org/10.1111/1755-0998.12338
Gasparini L, Crookes S, Prosser SR, Hanner R (2020) Detection of freshwater mussels (Unionidae) using environmental DNA in riverine systems. Environ DNA 2:321–329. https://doi.org/10.1002/edn3.71
Gaubert P, Justy F, Mo G, Aguilar A, Danyer E, Borrell A, Dendrinos P, Öztürk B, Improta R, Tonay AM, Karamanlidis AA (2019) Insights from 180 years of mitochondrial variability in the endangered Mediterranean monk seal (Monachus monachus). Mar Mammal Sci 35:1489–1511. https://doi.org/10.1111/mms.12604
Jeunen G, Lamare MD, Knapp M, Spencer HG, Taylor HR, Stat M, Bunce M, Gemmell NJ (2020) Water stratification in the marine biome restricts vertical environmental DNA (eDNA) signal dispersal. Environ DNA 2(1):99–111. https://doi.org/10.1002/edn3.49
Karamanlidis AA, Skrbinšek T, Amato G, Dendrinos P, Gaughran S, Kasapidis P, Kopatz A, Stronen AV (2021a) Genetic and demographic history define a conservation strategy for earth’s most endangered pinniped, the Mediterranean monk seal Monachus monachus. Sci Rep 11:1–10. https://doi.org/10.1038/s41598-020-79712-1
Karamanlidis A, Dendrinos P, Trillmich F (2021b) Maternal behavior and early behavioral ontogeny of the Mediterranean monk seal Monachus monachus in Greece. Endanger Species Res 45:13–20. https://doi.org/10.3354/esr01114
Karamanlidis AA, Dendrinos P (2015) Monachus monachus. The IUCN Red List of Threatened Species. e.T13653A45227543
Kasai A, Takada S, Yamazaki A, Masuda R, Yamanaka H (2020) The effect of temperature on environmental DNA degradation of Japanese eel. Fisheries Sci. https://doi.org/10.1371/journal.pone.0255576
Klymus KE, Merkes CM, Allison MJ, Goldberg CS, Helbing CC, Hunter ME, Piaggio AJ (2019) Reporting the limits of detection and quantification for environmental DNA assays. Environ DNA 2:271–282. https://doi.org/10.1002/edn3.29
La Mesa G, Lauriano G, Mo G, Paglialonga A, Tunesi L (2021) Assessment of the conservation status of marine species of the Habitats Directive (92/43/EEC) in Italy: results, drawbacks and perspectives of the fourth national report (2013–2018). Biodivers Conserv 30:4251–4264. https://doi.org/10.1007/s10531-021-02303-7
Lacoursière-Roussel A, Deiner K (2019) Environmental DNA is not the tool by itself. J Fish Biol 1:1–4. https://doi.org/10.1111/jfb.14177
Lance R, Guan X (2019) Variation in inhibitor effects on qPCR assays and implications for eDNA surveys. Can J Fish Aquat Sci 77:23–33. https://doi.org/10.1139/cjfas-2018-0263
Loeza-Quintana T, Abbott CL, Heath DD, Bernatchez L, Hanner RH (2020) Pathway to Increase Standards and Competency of eDNA Surveys (PISCeS)—advancing collaboration and standardization efforts in the field of eDNA. Environ DNA 2:255–260. https://doi.org/10.1002/edn3.112
Ma H, Stewart K, Lougheed S, Zheng J, Wang Y, Zhao J (2016) Characterization, optimization, and validation of environmental DNA (eDNA) markers to detect an endangered aquatic mammal. Conserv Genet Resour 8:561–568. https://doi.org/10.1007/s12686-016-0597-9
Martínez-Jauregui M, Tavecchia G, Cedenilla MA, Coulson T, Fernández de Larrinoa M, Muñoz M, González LM (2012) Population resilience of the Mediterranean monk seal Monachus monachus at Cabo Blanco peninsula. Mar Ecol Prog Ser 461:273–281. https://doi.org/10.3354/meps09811
McKee AM, Spear SF, Pierson TW (2015) The effect of dilution and the use of a post-extraction nucleic acid purification column on the accuracy, precision, and inhibition of environmental DNA samples. Biol Conserv 183:70–76. https://doi.org/10.1016/j.biocon.2014.11.031
Mo G (2011) Mediterranean Monk Seal (Monachus monachus) Sightings in Italy (1998–2010) and Implications for Conservation. Aquat Mamm 37(3):236–240. https://doi.org/10.1578/AM.37.3.2011.236
Moyer GR, Diaz-Ferguson E, Hill JE, Shea C (2014) Assessing environmental DNA detection in controlled lentic systems. PLoS ONE 9(7):e103767. https://doi.org/10.1371/journal.pone.0103767
Nicolaou H, Dendrinos P, Marcou M, Michaelides S, Karamanlidis AA (2019) Re-establishment of the Mediterranean monk seal Monachus monachus in Cyprus: priorities for conservation. Oryx 55(4):1–3. https://doi.org/10.1017/S0030605319000759
Pilliod DS, Goldberg CS, Arkle RS, Waits LP (2014) Factors influencing detection of eDNA from a stream-dwelling amphibian. Mol Ecol Res 14(1):109–116. https://doi.org/10.1111/1755-0998.12159
Pires R, Neves HC, Karamanlidis AA (2008) The Critically Endangered Mediterranean monk seal Monachus monachus in the archipelago of Madeira: priorities for conservation. Oryx 42:278–285. https://doi.org/10.1017/S0030605308006704
Ratnasingham S, Hebert PDN (2007) BOLD: The Barcode of Life Data System (www.barcodinglife.org). Mol Eco Notes 7:355–364. https://doi.org/10.1111/j.1471-8286.2007.01678.x
Ruppert KM, Kline RJ, Rahman MS (2019) Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: a systematic review in methods, monitoring, and applications of global eDNA. Glob Ecol Conserv 17:e00547. https://doi.org/10.1016/j.gecco.2019.e00547
Saito T, Doi H (2021) A model and simulation of the influence of temperature and amplicon length on environmental DNA degradation rates: a meta-analysis approach. Front Ecol Evol 9:139. https://doi.org/10.3389/fevo.2021.623831
Saydam E (2018) A survival blueprint for the Mediterranean monk seal. Mediterranean Conservation Society, Zoological Society of London, Izmir, Monachus monchus
Stat M, Huggett MJ, Bernasconi R, DiBattista J, Berry TE, Newman SJ, Harvey ES, Bunce M (2017) Ecosystem biomonitoring with eDNA: metabarcoding across the tree of life in a tropical marine environment. Sci Rep 7:1–11. https://doi.org/10.1038/s41598-017-12501-5
Stewart KA (2019) Understanding the effects of biotic and abiotic factors on sources of aquatic environmental DNA. Biodivers Conserv 28(5):983–1001. https://doi.org/10.1007/s10531-019-01709-8
Stoeckle MY, Das Mishu M, Charlop-Powers Z (2020) Improved environmental DNA reference library detects overlooked marine fishes in New Jersey. US Front Mar Sci 7:226. https://doi.org/10.3389/fmars.2020.00226
Stoffel MA, Humble E, Paijmans AJ, Acevedo-Whitehouse K, Chilvers BL, Dickerson B, Galimberti F, Gemmell NJ, Goldsworthy SD, Nichols HJ, Krüger O, Negro S, Osborne A, Pastor T, Robertson BC, Sanvito S, Schultz JK, Shafer ABA, Wolf JBW, Hoffman JI (2018) Demographic histories and genetic diversity across pinnipeds are shaped by human exploitation, ecology and life-history. Nat Commun 9:1–12. https://doi.org/10.1038/s41467-018-06695-z
Strickler KM, Fremier AK, Goldberg CS (2015) Quantifying effects of UV-B, temperature, and pH on eDNA degradation in aquatic microcosms. Biol Conserv 183:85–92. https://doi.org/10.1016/j.biocon.2014.11.038
Taberlet P, Coissac E, Hajibabaei M, Rieseberg LH (2012) Environmental DNA. Mol Ecol 21:1789–1793. https://doi.org/10.1111/j.1365-294X.2012.05542.x
Troth CR, Sweet MJ, Nightingale J, Burian A (2021) Seasonality, DNA degradation and spatial heterogeneity as drivers of eDNA detection dynamics. Sci Total Environ 768:144466. https://doi.org/10.1016/j.scitotenv.2020.144466
Tsuji S, Ushio M, Sakurai S, Minamoto T, Yamanaka H (2017) Water temperature-dependent degradation of environmental DNA and its relation to bacterial abundance. PLoS ONE 12(4):e0176608. https://doi.org/10.1371/journal.pone.0176608
Valsecchi E, Glockner-Ferrari D, Ferrari M, Amos W (1998) Molecular analysis of the efficiency of sloughed skin sampling in whale population genetics. Mol Ecol 7:1419–1422. https://doi.org/10.1046/j.1365-294x.1998.00446.x
Valsecchi E, Bylemans J, Goodman SJ, Lombardi R, Carr I, Castellano L, Galimberti A, Galli P (2020) Novel universal primers for metabarcoding environmental DNA surveys of marine mammals and other marine vertebrates. Environ DNA 2:460–476. https://doi.org/10.1002/edn3.72
Valsecchi E, Arcangeli A, Lombardi R, Boyse E, Carr IM, Galli P, Goodman SJ (2021) Ferries and environmental DNA: underway sampling from commercial vessels provides new opportunities for systematic genetic surveys of marine biodiversity. Front Mar Sci 8:1136. https://doi.org/10.3389/fmars.2021.704786
van Bochove K, Bakker FT, Beentjes KK, Hemerik L, Vos RA, Gravendeel B (2020) Organic matter reduces the amount of detectable environmental DNA in freshwater. Ecol Evol 10(8):3647–3654. https://doi.org/10.1002/ece3.6123
Wang H, Qi J, Xiao D, Wang Z, Tian K (2017) A re-evaluation of dilution for eliminating PCR inhibition in soil DNA samples. Soil Biol Biochem 106:109–118. https://doi.org/10.1016/j.soilbio.2016.12.011
Acknowledgements
We thank Giuseppe Sciancalepore of the Dept. of Comparative Biomedicine and Food Science of the University of Padova for providing the monk seal archive tissue sample from the Mediterranean marine mammal tissue bank (www.marinemammals.eu). We are grateful to Corsica and Sardinia Ferries, particularly to Cristina Pizzutti, for her willingness to explore the potential of eDNA sampling from their fleet, and we thank Dr Antonella Arcangeli of ISPRA who set up the FLT network within which our protocol was first tested. Thanks to the rangers from the IFCN for the sample collection off the Desertas Islands, to Moby Diving and Triton Research for support in the sample collection off Lampedusa, and Franco D’Urso of the KRCA for prompt supply of the sampling bags (BiBSS). We thank Gruppo Foca Monaca APS-Rome—and Grupa Sredozemna Medvjedica-Zagabira—for providing the biological residues from their archives and the data on occasional observations over the past decades. We also thank the University of Bicocca Marine Science, past and current master students Roberto Lombardi, Sofia Frappi, Simone Innocente and Francesca Gagliardi, for aliquots of the samples collected for their theses that were used to test the assays presented in this study. Finally, we would like to thank all the supporters of the “MeD for Med” (“Marine eDNA for the Mediterranean”) crowdfunding project, who enthusiastically believed in our initiative even though the fundraising campaign was launched during the COVID pandemic, restricting our sampling. We are also greatly thankful to the four anonymous referees whose constructive criticism and valuable comments have helped us to improve the manuscript, and to Mark Sefton (BioemdRed SL) for author editing of our final draft.
Funding
The research was funded by Bicocca University Crowdfund project MeD for Med (Marine eDNA for the Mediterranean): Fund “2020-CONT-0312 Bicocca Università Del Crowdfunding—MeD for Med”.
Author information
Authors and Affiliations
Contributions
Conceptualization: EV; methodology: EV, AB; formal analysis and investigation: EV, AB; writing—original draft preparation: EV; writing—review and editing: EC, RP, AP, MC; PG, AB; funding acquisition: EV, PG, MC; resources: EC, RP, AP; supervision: EV.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Additional information
Communicated by James Tony Lee.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article belongs to the Topical Collection: Coastal and marine biodiversity.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Valsecchi, E., Coppola, E., Pires, R. et al. A species-specific qPCR assay provides novel insight into range expansion of the Mediterranean monk seal (Monachus monachus) by means of eDNA analysis. Biodivers Conserv 31, 1175–1196 (2022). https://doi.org/10.1007/s10531-022-02382-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-022-02382-0