
Abstract
Land-use change has led to substantial range contractions for many species. Such contractions are particularly acute for wide-ranging large carnivores in Asia’s high altitude areas, which are marked by high spatiotemporal variability in resources. Current conservation planning for human-dominated landscapes often takes one of two main approaches: a “coexistence” (land sharing) approach or a “separation” (land sparing) approach. In this study, we evaluated the effects of land-use management on a guild of large carnivores in a montane ecosystem located in northeastern Iran. We used interview surveys to collect data on Persian leopard Panthera pardus saxicolor and grey wolf Canis lupus and modeled the areas occupied by these species in a Bayesian framework. After accounting for imperfect detection, we found that wolves had a higher probability of occupying the study area than leopards (82%; 95% CI 73–90% vs. 63%; 95% CI 53–73%). Importantly, each predator showed contrasting response to land-use management. National Parks (i.e. human-free areas) had a positive association with leopard occupancy (αNational Park = 2.56, 95% CI 0.22–5.77), in contrast to wolves, which displayed a negative association with National Parks (αNational Park = − 1.62, 95% CI − 2.29 to 0.31). An opposite pattern was observed for human-dominated areas (i.e. Protected Areas and Communal Lands), where occupancy was higher for wolves but lower for leopards. Our study suggests that to protect these large carnivores, a combination of land sharing and land sparing approaches is desirable within Iran montane landscapes. Any recovery program for big cats in Iranian mountains, and likely similar mountainous landscapes in west Asia, should take into account other sympatric carnivores and how they can affect adjacent human communities. For example, conflict mitigation and compensation efforts are required to include the guild of large carnivores, instead of solely targeting the charismatic big cats.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Many of the world’s large carnivores have undergone substantial range contractions, especially in areas with high rural human population densities (Kabir et al. 2017; Petracca et al. 2018; Rasphone et al. 2019). Protected areas are one effective tool for biodiversity conservation, particularly for large carnivores (Ahmadi et al. 2020; Ashrafzadeh et al. 2020). However, landscapes with small protected areas in relation to the extensive spatial requirements of large carnivores and a lack of intermediate corridors can lead to species declines (Carter and Linnell 2016; Farr et al. 2019). Therefore, in addition to establishing protected areas, understanding the spatial patterns of large carnivores beyond the protected areas boundaries is needed to adopt proper management approaches in human-dominated landscapes (Macdonald et al. 2019; Mohammadi et al. 2021).
Two main approaches have been discussed as best solutions for protecting large carnivores under land-use changes, defined here as a process by which human activities transform natural landscapes. The “separation” (land sparing) model, which designates separate areas for human use and wildlife conservation, has been shown to be effective for a wide range of large carnivores (Packer et al. 2013; Jiang et al. 2017). In contrast, the “coexistence” (land sharing) model, in which carnivores and humans inhabit shared landscapes, is believed to have facilitated the recovery of several large carnivores (Chapron et al. 2014; Carter and Linnell 2016). Sparing extensive tracts of land for large carnivores is less plausible in many parts of the world, particularly in dry and human-dominated landscape where these animals need to range widely to access unpredictable and spatially heterogeneous resources, notably prey and water (Kabir et al. 2017; Farhadinia et al. 2018b; Mohammadi et al. 2021). In the context of Asia’s montane landscapes, a few empirical studies have evaluated these two approaches for big cats (Johansson et al. 2016; Jiang et al. 2017; Farhadinia et al. 2018b); however, research conceptualizing the interaction of these approaches for guilds of large carnivores remains uncommon, particularly in west Asia.
In this study, we assessed the effects of land-use management on a guild of large carnivores: Persian leopard Panthera pardus saxicolor and grey wolf Canis lupus in Kopet Dag Ecoregion, a region of rugged terrain along northeastern Iran and southern Turkmenistan. These species compete over limited resources, particularly prey whose availability is mandated by land-use management. For example, wild ungulates, including bezoar goat (Capra aegagrus), urial sheep (Ovis orientalis), goitered gazelle (Gazella subgutturosa) and wild pig (Sus scrofa) have higher densities in spared lands (i.e. national parks) (Farhadinia et al. 2018b; Ghoddousi et al. 2016; Hosseini-Zavarei et al. 2013; Taghdisi et al. 2013). In contrast, anthropogenic food resources, notably livestock and garbage, which contribute more to the dietary requirement of the grey wolf than the Persian leopard (Babrgir et al. 2017; Behmanesh et al. 2019; Mohammadi et al. 2019), generally occur in shared lands (i.e. communal lands).
In this study, we evaluated the occupancy patterns of two sympatric species that differed in conservation status: Persian leopard (Endangered) and grey wolf (Least Concern). We used occupancy modeling, which employs repeated detection/non-detection surveys to account for imperfect detection (MacKenzie et al. 2002). We first hypothesized that leopards would have higher probability of occupancy in national parks (spared lands) that provide habitat for wild ungulates, which are the main prey for leopards (Taghdisi et al. 2013; Farhadinia et al. 2018c). In contrast, we hypothesized that grey wolves would mainly occur in communal lands (shared lands) that support abundant livestock, which are a main component of wolves diet in the study area (Hosseini-Zavarei et al. 2013; Farhadinia et al. 2017; Mohammadi et al. 2019). We also compared occupancy patterns in relation to wild prey abundance index as well as the proximity to villages (as a surrogate for human disturbance/livestock availability). We conclude by providing management implications for protecting a guild of large carnivores in multi-use landscapes of Iranian mountains.
Materials and methods
Ethics statement
The Iranian Department of Environment reviewed all sampling procedures and approved permits for the work conducted (93/16270). A descriptive summary of the research was provided to each human participant (the document was read to illiterate participants,) and all participants gave oral consent before being interviewed.
Study area
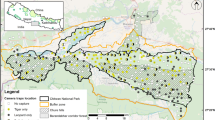
We conducted research across multiple montane reserves in northeastern Iran, including Tandoureh National Park and Protected Area (hereafter NP and PA, respectively), Salouk NP and PA, and Sarigol NP and PA. As parts of the Kopet Dag and Aladagh Mountains (Memariani et al. 2016), these coarse mountainous areas exist at the eastern extreme of the Irano-Anatolian Biodiversity Hotspot (E57°15′–E59°15′, N36° 20′–N37°20′) (Fig. 1). These semi-arid areas receive 200–300 mm of annual rainfall (Darvishsefat 2006). Urial, bezoar goat, and wild pig are the main wild ungulates in these montane reserves (Farhadinia et al. 2018c; Hosseini et al. 2019). Previous systematic camera trapping surveys revealed that high densities of leopards exist in the study areas, varying between 3.1 and 8.9 individuals/100 km2 (Farhadinia et al. 2019) due to prey availability and spatial distribution of prey. For example, in Sarigol NP and PA, prey is more concentrated in a small proportion of the area, resulting in a high density of leopards in that part (8.9 individuals/100 km2 in Sarigol NP) (Farhadinia et al. 2019). Leopards and wolves are likely killed frequently due to conflicts related to livestock depredations (Parchizadeh and Belant 2021). There are over 26 villages surrounding the three study areas with over 34,000 livestock, averaging 470 animals per herd. Grazing herds are composed largely of sheep (O. aries; 84% ± 2) with smaller numbers of goats (C. hircus). Herders adopt a combination of husbandry methods such as shepherd, herd dogs and nigh enclosures to reduce conflicts with carnivores (Farhadinia et al. 2017). In this study, NPs (625 km2) constitute spared lands while PAs (305 km2) and CLs (1820 km2) comprise shared lands, resulting in a total surveyed area of 2750 km2.

The predicted relative probability of occupancy for Persian leopard and grey wolf as calculated from models built using interview surveys in northeastern Iran. Each of 110 sampling sites (cells) is depicted at a resolution of 25 km2
Each study area (i.e. Tandoureh, Sarigol and Salouk) encompassed three different land management regimes, including (1) NPs that have stronger anti-poaching law enforcement and prohibit livestock grazing; (2) PAs that have less intense anti-poaching enforcement and permit livestock grazing during summer months (May–August) by nomadic pastoralists from surrounding settlements; and (3) CLs that have multi-use areas where human settlements are located. CLs are composed of villages, farmlands and pastures without efficient law enforcement, implying that illegal activities such as poaching are less deterred because most of the anti-poaching capacity is concentrated in NPs, and to a lesser extent in PAs.
Sampling and environmental covariates
We surveyed 110 sampling sites across three study areas in northeastern Iran. Sampling sites were located in NPs (n = 25, 23%), PAs (n = 21, 19%), and CLs (n = 64, 58%) (Table 1 and S1). The spatial resolution of each sampling site was 25 km2, corresponding to the minimum home range size of a female leopard. This size was calculated based on the minimum convex polygon of a male leopard based on GPS telemetry in Tandoureh (63.3 km2; Farhadinia et al. 2018a) and the female/male home range ratio of 0.4 (du Preez et al. 2014). This spatial resolution allows us to interpret our findings in the context of species occupancy patterns as it was similarly used in previous occupancy modeling research for leopards (Farhadinia et al. 2018c; Soofi et al. 2018).
We divided interviewees into two groups based upon their occupation and the areas in which they worked: rangers (n = 13) and herders (n = 80). Rangers were based in NPs and PAs whereas herders were able to graze their livestock in PAs and CLs. A team of 2–3 surveyors visited a total of 26 villages bordering the study areas to interview herders. Only one person associated with each herd was interviewed (typically the only person accompanying the herd in the pasture). Each interview had two stages. We first evaluated the respondent’s ability to correctly identify the carnivores. Thus, all respondents successfully identified the three large carnivore species inhabiting the study areas, including striped hyena (Hyaena hyaena), grey wolf, and Persian leopard. Then, interviewees were asked about any encounter with leopards and wolves within their geographic area of familiarity, typically sites within their herd’s grazing pasture. We asked each interviewee to provide information from the recent past (i.e., the last 3–6 months) to minimize potential bias due to recollection of older records. We interviewed each herder between August 2013 and September 2014 and created detection records per herder. We only accepted direct sightings to minimize risk of species misidentification. On average, we conducted 6.4 (SD 4.0) interviews per sampling site. Most herders used pasture across multiple sampling sites and were thus able to provide data for more than one sampling site. Following Zeller et al. (2011), we used repeated interviews with different respondents at the same sites as survey replicates for the occupancy model. Each respondent provided information for an average of 5.8 (SD 3.4) of different sampling units.
We included the protection status of each sampling site as a categorical covariate (Table 2). The three categories of protection included NPs (human-free areas with established anti-poaching and grazing ban), PAs (limited anthropogenic land use change and over-grazing control, but with less efficient anti-poaching) and CLs (intensively human-modified areas without any efficient regulatory control (e.g. without over-grazing control)).
We also hypothesized that three environmental covariates might affect the occupancy of the two large carnivores (Table 2). As surrogates for the wild prey abundance as well as human disturbance/livestock availability, we quantified the Euclidean distance in kilometers of each sampling site centroid to the nearest active ranger stations and main village, respectively (Table S1). We expected that prey would be more abundant in closer proximity to ranger stations due to increased anti-poaching efforts (Ghoddousi et al. 2016) while human disturbance would decline with increasing distance from villages because of reduced human activity (Farhadinia et al. 2018c). To calculate the distance variables, we obtained the spatial location of human settlements (i.e. villages and ranger stations) using a handheld GPS receiver and then we visually confirmed village locations using Google Earth Pro 7.1.7.2606 (Google Inc., USA). Additionally, we used Zonal Statistics in ArcGIS 10.2 (ESRI Co., USA) to calculate a ruggedness index for each sampling site as the standard deviation of the elevation levels in meters within each site as a proxy for landscape heterogeneity (Hosseini et al. 2019; Ahmadi et al. 2020; Ashrafzadeh et al. 2020). Prior to modeling, we examined Pearson’s correlations between these covariates and found that none displayed collinearity (all pairwise r values were < 0.42).
Data analysis
We modeled leopard and wolf occupancy in a hierarchical Bayesian framework (MacKenzie et al. 2002; Tyre et al. 2003). These models estimate occupancy probability (Ψ) and detection probability (p) as well as the effects of covariates on these two parameters (Karanth et al. 2011). Given the mobile nature of the study species, we note that occupancy in this context represents the use of a given site by our focal species during the study period rather than continuous occupancy (MacKenzie et al. 2002; Mackenzie 2006). The parameter p is the probability of species detection during a survey given that it occupies that sampling site.
Specifically, we modeled occupancy as a latent variable (Zi) that was Bernoulli-distributed and took a value of one when site i was occupied and zero otherwise. We modeled effect of covariates on occupancy probability using a logit link. To facilitate comparison, models for both species used the same environmental covariates. These models took the form:
where α0 is an intercept, αk is the estimated effect of the kth protection status, αranger is the estimated effect of proximity to the nearest ranger station, αvillage is the estimated effect of proximity to the nearest village, and αrugged is the estimated effect of ruggedness (Table 2). The protection status took one of three levels (CL, NP, and PA, with the CL as the reference level; see above). We also used a logit link to model the effect of covariates on p at site i and replicate j. Thus, the detection model had the form:
where β0 is an intercept and βranger is the estimated effect of ranger observers compared to herders.
To facilitate convergence and enable a comparison of effect sizes among parameters, we standardized all continuous covariates to have a mean of zero and a standard deviation of one (Kéry 2010). We analyzed both models in a Bayesian framework using Markov Chain Monte Carlo (MCMC) simulations. We generated MCMC simulations using R Studio version 1.0.136 (R Development Core Team 2013) and JAGS (Plummer 2003) via the package R2jags (Su and Yajima 2012). We used non-informative priors for all parameters that had a logistic distribution centered at zero with a scale parameter of one (Northrup and Gerber 2018). We ran three MCMC chains of 20,000 iterations each following a burn-in of 10,000 and thinned chains by five. We confirmed model convergence using R-hat statistics (i.e., all values were < 1.1; Gelman and Hill 2007) and by visually inspecting trace plots. We used Bayesian p-values to assess model goodness-of-fit (Gelman et al. 1996), where values near 0.5 indicate good model fit and extreme values (near zero or one) indicate poor model fit (Gelman et al. 1996; Kéry and Royle 2015).
Results
We interviewed a total of 93 individuals, including 13 rangers and 80 herders, resulting in a similar total number of detections for each predator (176 leopard detections vs. 184 wolf detections; Table 1). The interview surveys provided different naïve site occupancy rates for each reserve, varying between 23.3 and 51.1% and 14.0–58.1% for leopards and grey wolves, respectively. In total, 48% of leopard sighting were inside the NPs, whereas only 15.5% of wolf sighting were inside NPs.
Bayesian p-values indicated a good fit for both models (pleopard = 0.64, pwolf = 0.65). Accounting for imperfect detection, the leopard model estimated that 63% (SD 5%) out of 110 sampling sites were occupied (95% CI 53–73%; Table 3). Wolves exhibited a higher probability of occupancy (82%; SD 4%; 95% CI 73–90%; Table 3). Both predators had similar overall detection probabilities (pwolf = 0.32, 95% CI 0.27–0.35, pleopard = 0.30, 95% CI 0.26–0.34). Observer type affected the detection probability for each species. Rangers had higher detection probabilities than herders for leopards (βranger = 3.97, 95% CI 3.26–4.75), while an opposing pattern existed for wolves (βranger = − 0.94, 95% CI − 1.58 to − 0.35; Table 3).
The two predators exhibited contrasting associations with environmental covariates. Leopard occupancy had a strong positive relationship with ruggedness (αrugged = 2.20, 95% CI − 0.82 to 3.90) while wolves had no such relationship (αrugged = − 0.34, 95% CI − 0.63 to 0.54; Fig. 2). Both species had higher occupancy near ranger stations, but this effect was stronger and less variable for leopards than wolves (Table 3; Fig. 2). An opposing pattern existed for distance to village, with wolves exhibiting a stronger and less variable relationship than leopards (Table 3; Fig. 2). Importantly, the two carnivore species showed contrasting responses to land-use management. Accordingly, NPs (i.e. human-free areas) had positive effect on leopard occupancy (αNational Park = 2.56, 95% CI 0.22, 5.77), whereas wolves had a negative association with NPs (α National Park = − 1.62, 95% CI − 2.29, 0.31; Table 3; Fig. 3). An opposite pattern existed for human-dominated areas (i.e. PAs and CLs), where the occupancy was higher for wolves but lower for leopards (Table 3; Fig. 3).

Comparative associations between the probability of occupancy with the distance to nearest ranger stations (as a proxy for wild prey abundance) and the distance to nearest village (as proxy for human disturbance/livestock availability) for Persian leopard and grey wolf in northeastern Iran based on interview surveys at 110 sampling sites. Black lines are model-averaged mean predictions and grey lines are model-averaged predictions from a random posterior sample of 200 iterations to depict uncertainty

Boxplots for the probability of occupancy of Persian leopards and wolves based on different land-use management estimated using interview surveys at 110 sampling sites in northeastern Iran. Parameter estimates with the 95% credible interval were estimated by Bayesian model analysis, with the central line marking the median. CL Communal Land, NP National Park, PA Protected Area
Discussion
We explored patterns of site occupancy for two sympatric carnivores in mountainous landscapes across three different land use management types in northeastern Iran. We found that leopards and wolves showed contrasting responses to land-use management and environmental covariates. Although NPs (spared lands with highest protection level) were favored by leopards, wolves more frequently occupied less protected land types (CLs that represented shared lands).
Contrasting responses to environmental covariates
The high occupancy of leopards near ranger stations and far from human-dominated areas correlates with the availability of the wild prey, notably urial and bezoar goat living in the mountainous areas of NPs (Farhadinia et al. 2018a). Ranger stations facilitated higher law-enforcement for prey species and, consequently, can provide benefits to their predators (Ghoddousi et al. 2016). In contrast to leopards, wolves showed a constant high probability of occupancy across a range of distances from villages, which is consistent with their plasticity in foraging on a wide range of food items, including mountain-dwelling ungulates such as urial and bezoar goat (Hosseini-Zavarei et al. 2013; Balajeid Lyngdoh et al. 2020), plains-occurring gazelles and livestock (Mohammadi et al. 2019; Werhahn et al. 2019), and even garbage (Hosseini-Zavarei et al. 2013; Tourani et al. 2014; Mohammadi et al. 2019). Similarly, the two predators exhibited varying responses to terrain ruggedness. In accordance to previous studies, we found that leopards often occur in rugged landscapes (Hosseini et al. 2019; Ahmadi et al. 2020; Ashrafzadeh et al. 2020). In contrast, wolves did not show any association with landscape ruggedness, but rather had a high probability of occupancy across a range of levels of terrain ruggedness.
Land sharing vs. land sparing
Our results showed that spared lands (i.e. NPs) were the main refugia for leopards, which occur in a high density varying between 3.1 and 8.9 individuals/100 km2 in these areas (Farhadinia et al. 2019). In contrast, wolves had higher probability of occupancy in shared lands (i.e. PAs and CLs), where they frequently attack people’s livestock (Hosseini-Zavarei et al. 2013; Mohammadi et al. 2019; Shahnaseri et al. 2019).
There are two non-exclusive explanations to explain the differentiated responses of our two focal carnivores to land-use management. The first is related to the association between carnivore occupancy and the selection of habitat and prey. In Iranian mountains, leopards prey mainly on cliff-dwelling wild ungulates such as urial and bezoar goat (Taghdisi et al. 2013; Sharbafi et al. 2016; Farhadinia et al. 2018a), whereas wolves mainly consume plain-dwelling ungulates, such as gazelles (which are available only in Salouk, but absent from the other two study areas), in addition to livestock and anthropogenic food resources (Hosseini-Zavarei et al. 2013; Tourani et al. 2014; Mohammadi et al. 2019; Werhahn et al. 2019; Balajeid Lyngdoh et al. 2020). A second explanation relates to intra-guild competition, which can cause different effects of land use types on the occupancy patterns of these predators. For example, wolves avoid mountainous areas with high leopard densities, a pattern which is commonly seen between sympatric gregarious canids and solitary felids (Bocci et al. 2017; Chetri et al. 2017; Elbroch and Kusler 2018; Srivathsa et al. 2019). However, a lack of historic data on wolves’ occurrence and density did not allow us to evaluate their occupancy in the absence of land sparing approach (i.e. when NPs were not designated). Overall, our data are consistent with both explanations; future research should focus on quantifying the relative contribution of these mechanisms.
National Parks (i.e., spared lands) are of paramount importance for supporting high densities of Persian leopards, insofar as they control anthropogenic causes of mortality for leopards and their prey, notably poaching (Soofi et al. 2018; Farhadinia et al. 2019). Nonetheless, there is evidence from satellite telemetry data that leopards spend around 17% of their time outside conservation areas (Farhadinia et al. 2018b). Consequently, these unprotected spaces must also be managed to minimize conflict with herders and promote the existence of carnivores in human-dominated landscapes (“land sharing”). Importantly, our findings showed that spared lands (NPs) can secure the viability of only one species in the studied carnivore guild (i.e., Persian leopard) while wolves are widely dependent on shared lands (PAs and CLs).
Management implications
Our study provided two key management implications. First, the recovery of leopards could be associated with the exclusion of grey wolves in northeastern Iran, a pattern that might also occur in other west Asian mountains. In response to leopard recovery, wolves might have shifted their spatial patterns to human-dominated habitats, which can result in escalated conflict with communities due to livestock depredation (Tourani et al. 2014; Mohammadi et al. 2019) which can be evaluated through monitoring of both carnivore population trends and their conflict levels with communities. Therefore, although conservation agencies might perceive their management plans to be successful in recovering leopards with minimum level of conflict with communities, people will likely experience high conflict with the other species, such as wolves. Nonetheless, this dynamic is not often considered in management plans (Farhadinia et al. 2017; Kusi et al. 2019; Trajçe et al. 2019). Therefore, any recovery program for big cats in Asian mountains should take into account the other sympatric carnivores and how they can affect adjacent human communities. For example, conflict mitigation and compensation efforts are required to include the guild of large carnivores, instead of solely targeting the charismatic big cats (Srivathsa et al. 2019; Werhahn et al. 2019).
Second, land sparing, which often involves protecting larger and better-connected areas, is preferred for supporting breeding nuclei of large cats in Asia’s rugged landscapes. However, this strategy is unlikely to be equally effective for safeguarding other large carnivores. Therefore, in Iranian mountains with scattered resource availability, a combined land sharing (i.e. establishing NPs and PAs) and land sparing (i.e. encouraging coexistence approaches such as conflict mitigation and community-based engagement) is envisaged to support the viability of the guild of large carnivores.
References
Ahmadi M, Farhadinia MS, Cushman SAM et al (2020) Species and space: a combined gap analysis to guide management planning of conservation areas. Landsc Ecol 35:1505–1517
Ashrafzadeh MR, Khosravi R, Adibi MA et al (2020) A multi-scale, multi-species approach for assessing effectiveness of habitat and connectivity conservation for endangered felids. Biol Conserv 245:108523
Babrgir S, Farhadinia MS, Moqanaki EM (2017) Socio-economic consequences of cattle predation by the endangered Persian leopard Panthera pardus saxicolor in a Caucasian conflict hotspot, northern Iran. Oryx 51:124–130
Balajeid Lyngdoh S, Habib B, Shrotriya S (2020) Dietary spectrum in Himalayan wolves: comparative analysis of prey choice in conspecifics across high-elevation rangelands of Asia. J Zool 310:24–33
Behmanesh M, Malekian M, Hemami MR, Fakheran S (2019) Patterns and determinants of human-carnivore conflicts in Central Iran: realities and perceptions behind the conflict. Hum Dimens Wildl 24:14–30
Bocci A, Lovari S, Khan MZ, Mori E (2017) Sympatric snow leopards and Tibetan wolves: coexistence of large carnivores with human-driven potential competition. Eur J Wildl Res 63:92
Carter NH, Linnell JDC (2016) Co-adaptation is key to coexisting with large carnivores. Trends Ecol Evol 31:575–578
Chapron G, Kaczensky P, Linnell JDC et al (2014) Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 346:1517–1519
Chetri M, Odden M, Wegge P (2017) Snow leopard and Himalayan wolf: food habits and prey selection in the Central Himalayas, Nepal. PLoS One 12:e0170549
Darvishsefat AA (2006) Atlas of protected areas of Iran. Ravi, Tehran
du Preez B, Loveridge AJ, Macdonald DW (2014) To bait or not to bait: a comparison of camera-trapping methods for estimating leopard (Panthera pardus) density. Biol Conserv 176:153–161
Elbroch LM, Kusler A (2018) Are pumas subordinate carnivores, and does it matter? PeerJ 6:e4293
Farhadinia MS, Johnson PJ, Hunter LTB, Macdonald DW (2017) Wolves can suppress goodwill for leopards: patterns of human-predator coexistence in northeastern Iran. Biol Conserv 213:210–217
Farhadinia MS, Johnson PJ, Hunter LTB, Macdonald DW (2018a) Persian leopard predation patterns and kill rates in the Iran–Turkmenistan borderland. J Mammal 99:713–723
Farhadinia MS, Johnson PJ, Macdonald DW, Hunter LTB (2018b) Anchoring and adjusting amidst humans: ranging behavior of Persian leopards along the Iran-Turkmenistan borderland. PLoS One 13:e0196602
Farhadinia MS, Moll RJ, Montgomery RA et al (2018c) Citizen science data facilitate monitoring of rare large carnivores in remote montane landscapes. Ecol Indic 94:283–291
Farhadinia MS, McClintock BT, Johnson PJ et al (2019) A paradox of local abundance amidst regional rarity: the value of montane refugia for Persian leopard conservation. Sci Rep 9:14622
Farr MT, Green DS, Holekamp KE et al (2019) Multispecies hierarchical modeling reveals variable responses of African carnivores to management alternatives. Ecol Appl 29:e01845
Gelman A, Hill J (2007) Data analysis using regression and multilevel/hierarchical models. Cambridge University Press, New York
Gelman A, Meng X-L, Stern H (1996) Posterior predictive assessment of model fitness via realized discrepancies. Stat Sin 6:733–807
Ghoddousi A, Soofi M, Khorozyan I et al (2016) Effects of ranger stations on predator and prey distribution and abundance in an Iranian steppe landscape. Anim Conserv 19:273–280
Hosseini-Zavarei F, Farhadinia MS, Beheshti-Zavareh M, Abdoli A (2013) Predation by grey wolf on wild ungulates and livestock in central Iran. J Zool 290:127–134
Hosseini M, Farashi A, Khani A, Farhadinia MS (2019) Landscape connectivity for mammalian megafauna along the Iran-Turkmenistan-Afghanistan borderland. J Nat Conserv. https://doi.org/10.1016/j.jnc.2019.125735
Jiang G, Wang G, Holyoak M et al (2017) Land sharing and land sparing reveal social and ecological synergy in big cat conservation. Biol Conserv 211:142–149
Johansson Ö, Rauset GR, Samelius G et al (2016) Land sharing is essential for snow leopard conservation. Biol Conserv 203:1–7
Kabir M, Hameed S, Ali H et al (2017) Habitat suitability and movement corridors of grey wolf (Canis lupus) in Northern Pakistan. PLoS One 12:e0187027
Karanth KU, Gopalaswamy AM, Kumar NS et al (2011) Monitoring carnivore populations at the landscape scale: occupancy modelling of tigers from sign surveys. J Appl Ecol 48:1048–1056
Kéry M (2010) Introduction to WinBUGS for ecologists: Bayesian approach to regression, ANOVA, mixed models and related analyses. Academic Press, London
Kéry M, Royle JA (2015) Applied hierarchical modeling in ecology: analysis of distribution, abundance and species richness in R and BUGS: Volume 1: Prelude and static models. Academic Press, London
Kusi N, Sillero-Zubiri C, Macdonald DW et al (2019) Perspectives of traditional Himalayan communities on fostering coexistence with Himalayan wolf and snow leopard. Conserv Sci Pract 2:e165
Macdonald DW, Bothwell HM, Kaszta Ż et al (2019) Multi-scale habitat modelling identifies spatial conservation priorities for mainland clouded leopards (Neofelis nebulosa). Divers Distrib 25:1639–1654
Mackenzie DI (2006) Modeling the probability of resource use: the effect of, and dealing with, detecting a species imperfectly. J Wildl Manag 70:367–374
MacKenzie DI, Nichols JD, Lachman GB et al (2002) Estimating site occupancy rates when detection probabilities are less than one. Ecology 83:2248–2255
Memariani F, Zarrinpour V, Akhani H (2016) A review of plant diversity, vegetation, and phytogeography of the Khorassan-Kopet Dagh floristic province in the Irano-Turanian region (northeastern Iran–southern Turkmenistan). Phytotaxa 249:8–30
Mohammadi A, Kaboli M, Sazatornil V, López-Bao JV (2019) Anthropogenic food resources sustain wolves in conflict scenarios of Western Iran. PLoS One 14:e0218345
Mohammadi A, Almasieh K, Nayeri D et al (2021) Identifying priority core habitats and corridors for effective conservation of brown bears in Iran. Sci Rep 11:1–13
Northrup JM, Gerber BD (2018) A comment on priors for Bayesian occupancy models. PLoS One 13:e0192819
Packer C, Loveridge A, Canney S et al (2013) Conserving large carnivores: dollars and fence. Ecol Lett 16:635–641
Parchizadeh J, Belant JL (2021) Human-caused mortality of large carnivores in Iran during 1980–2021. Glob Ecol Conserv 27:e01618
Petracca LS, Frair JL, Cohen JB et al (2018) Robust inference on large-scale species habitat use with interview data: the status of jaguars outside protected areas in Central America. J Appl Ecol 55:723–734
Plummer M (2003) JAGS: a program for analysis of Bayesian graphical models using Gibbs sampling. In: Proceedings of the 3rd international workshop on distributed statistical computing, vol 124, pp 1–8
R Development Core Team (2013) R: a language and environment for statistical computing
Rasphone A, Kéry M, Kamler JF, Macdonald DW (2019) Documenting the demise of tiger and leopard, and the status of other carnivores and prey, in Lao PDR’s most prized protected area: Nam Et-Phou Louey. Glob Ecol Conserv 20:e00766
Shahnaseri G, Hemami M-R, Khosravi R et al (2019) Contrasting use of habitat, landscape elements, and corridors by grey wolf and golden jackal in central Iran. Landsc Ecol 34:1263–1277
Sharbafi E, Farhadinia MS, Rezaie HR, Braczkowski AR (2016) Prey of the Persian leopard (Panthera pardus saxicolor) in a mixed forest-steppe landscape in northeastern Iran (Mammalia: Felidae). Zool Middle East 62:1–8
Soofi M, Ghoddousi A, Zeppenfeld T et al (2018) Livestock grazing in protected areas and its effects on large mammals in the Hyrcanian forest, Iran. Biol Conserv 217:377–382
Srivathsa A, Karanth KU, Kumar NS, Oli MK (2019) Insights from distribution dynamics inform strategies to conserve a dhole Cuon alpinus metapopulation in India. Sci Rep 9:3081
Su YS, Yajima M (2012) R2jags: a package for running jags from R
Taghdisi M, Mohammadi A, Nourani E et al (2013) Diet and habitat use of the endangered Persian leopard (Panthera pardus saxicolor) in northeastern Iran. Turk J Zool 37:554–561
Tourani M, Moqanaki EM, Boitani L, Ciucci P (2014) Anthropogenic effects on the feeding habits of wolves in an altered arid landscape of central Iran. Mammalia 78:117–121
Trajçe A, Ivanov G, Keçi E et al (2019) All carnivores are not equal in the rural people’s view. Should we develop conservation plans for functional guilds or individual species in the face of conflicts? Glob Ecol Conserv 19:e00677
Tyre AJ, Tenhumberg B, Field SA et al (2003) Improving precision and reducing bias in biological surveys: estimating false-negative error rates. Ecol Appl 13:1790–1801
Werhahn G, Kusi N, Li X et al (2019) Himalayan wolf foraging ecology and the importance of wild prey. Glob Ecol Conserv 20:e00780
Zeller KA, Nijhawan S, Salom-Perez R et al (2011) Integrating occupancy modeling and interview data for corridor identification: a case study for jaguars in Nicaragua. Biol Conserv 144:892–901
Acknowledgements
We sincerely thank Iranian Department of Environment for administrative support. We are also grateful to rangers within investigated areas, particularly M. Taghdisi, H. Fakhrani, A. Hosseinzadeh, M. Ahmadi, S. Alizadeh, A. Hosseini, V. Alizadeh, B. Moradian, S. Gerivani, H. Mohebi, G. Pishghadam, A. Daneshfar, S. Firouzi, H. Nourani, H. Eslahi, M. Teymouri and Z. Salahshour for their field assistance. MSF was supported by a research fellowship from the Oxford Martin School at the University of Oxford.
Author information
Authors and Affiliations
Contributions
MSF conceived the ideas and designed methodology; MSF, KH, PB and PM collected the data; MSF, CT, RJM, CL and AM analyzed the data; AM, MSF, CL and DWM led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Adeline Loyau.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mohammadi, A., Lunnon, C., Moll, R.J. et al. Contrasting responses of large carnivores to land use management across an Asian montane landscape in Iran. Biodivers Conserv 30, 4023–4037 (2021). https://doi.org/10.1007/s10531-021-02290-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-021-02290-9