
Abstract
Determining the density trends of a guild of species can help illuminate their interactions, and the impacts that humans might have on them. We estimated the density trends from 2013 to 2017 of the clouded leopard Neofelis nebulosa, leopard cat Prionailurus bengalensis and marbled cat Pardofelis marmorata in Nam Et—Phou Louey National Park (NEPL), Laos, using camera trap data and spatial capture-recapture models. Mean (± SD) density estimates (individuals/100 km2) for all years were 1.77 ± 0.30 for clouded leopard, 1.50 ± 0.30 for leopard cat, and 3.80 ± 0.70 for marbled cat. There was a declining trend in density across the study years for all three species, with a ≥ 90% probability of decline for clouded leopard and leopard cat and an 83% probability of decline for marbled cat. There was no evidence that mesopredator release occurred as a result of tiger (Panthera tigris) and leopard (P. pardus) extirpations. We believe that snaring, the factor that led to the extirpation of tiger and leopard in NEPL, is now contributing to the decline of smaller felids, to an extent that over-rides any potential effects of mesopredator release on their densities and interactions. We recommend that the NEPL managers implement a more systematic and intensified snare removal program, in concert with extensive community outreach and engagement of local people to prevent the setting of snares. These actions might be the only hope for saving the remaining members of the felid community in NEPL.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Southeast Asia has the highest diversity of felids in the world (Luo et al., 2014) with as many as nine species occupying a diverse range of habitats on the mainland. In the evergreen forests of Southeast Asia, north of the Kra Isthmus, the felid community historically consisted of two large felids, tiger (Panthera tigris) and leopard (P. pardus), together with the “standard four” medium and small felids: clouded leopard (Neofelis nebulosa), Asian golden cat (Catopuma temminckii), marbled cat (Pardofelis marmorata), and leopard cat (Prionailurus bengalensis; Duckworth et al. 2014). Several studies have examined the mechanisms that allow such a diverse felid community to coexist, and have concluded that temporal partitioning and dietary partitioning likely help to facilitate their coexistence (Mukherjee et al. 2019; Nagy-Reis et al. 2019; Kamler et al. 2020; Rasphone et al. 2020). Additionally, smaller felid species may sometimes, but not always, behaviorally avoid larger felid species (McDougal et al. 1988; Horne et al. 2009; Foster et al. 2013; Rostro-García et al. 2015, 2021), thereby further facilitating their coexistence.
Compared to the numerous studies investigating coexistence mechanisms among felid species, few studies have investigated the effects of larger felids on the densities of smaller felids. Regarding the two large felids, some studies have shown that tiger and leopard numbers are inversely related (Harihar et al. 2011; Mondal et al. 2012; Kumar et al. 2019); thus, the tiger can suppress populations of leopard by killing and spatial displacement (Seidensticker 1976; McDougal 1988; Odden and Wegge 2009), at least in some areas. However, whether large felids suppress populations of medium and small felids has never been studied. The mesopredator release hypothesis states that the removal or reduction of large carnivores can result in population increases in medium-sized carnivores (Soulé et al. 1988; Prugh et al. 2009), which has been supported by several studies, mostly on canid communities (Crabtree and Shelden 1999; Kamler et al. 2003, 2013). In turn, an increase in mesocarnivores might cause population declines in small carnivores (Levi and Wilmers 2012).
Southeast Asia is considered one of the most biologically threatened regions in the world primarily because of widespread habitat loss and wildlife poaching (Hughes 2017). In particular, widespread and indiscriminate snaring for the illegal wildlife trade is devastating animal communities and overall biodiversity in region, in what has been called a snaring crises for Southeast Asia (Gray et al. 2017, 2018). Large felids often are targeted for the illegal wildlife trade (Ripple et al. 2016), and their populations are especially susceptible to poaching (Chapron et al. 2008). Consequently, tiger populations have crashed throughout Southeast Asia, and they have become extirpated over a large majority of the region (Goodrich et al. 2015). Similarly, leopard have recently experienced a range collapse in Southeast Asia primarily due to poaching (Rostro-García et al. 2016), and they are now extirpated from 94–98% of their former distribution in the region, and were recently classified as Critically Endangered (Rostro-García et al. 2019). Distributions of medium and small-sized felids have also been reduced in Southeast Asia, although not to the same extent as large felids. For example, the clouded leopard (16–23 kg; Lekagul and McNeely 1977) has been extirpated from most of its former distribution in Southeast Asia due to poaching and habitat fragmentation (Oswell 2010), and this species is classified as Vulnerable (Grassman et al. 2016). Similarly, the distributions of Asian golden cat (12–15 kg) and marbled cat (2–5 kg; Lekagul and McNeely 1977) also have been reduced substantially, and both species are classified as Near Threatened (McCarthy et al. 2015; Ross et al. 2016). In contrast, the leopard cat (3–5 kg; Lekagul and McNeely 1977) is still common and widespread in the region, and is classified as a Least Concern species (Ross et al. 2015).
The loss of tiger and leopard in over 90% of Southeast Asia might have significant implications for the remaining felid communities, especially if mesopredator release is occurring (Macdonald et al. 2010). For example, the extirpation of these two large felids might result in population increases of medium-sized felids, such as clouded leopard and Asian golden cat, at least at the local level. In turn, increases in medium-sized felids might result in population decreases of small felids, such as marbled cat and leopard cat. However, if poaching for the illegal wildlife trade was the primary reason for the extirpation of tiger and leopard at a site, then poachers may subsequently start targeting smaller felids, which ultimately might reduce theirs populations regardless of mesopredator release. For example, clouded leopard and Asian golden cat are often targeted by poachers for their pelt and body parts, and these species are increasingly being used as substitutes for tigers in traditional Asian medicine (D’Cruze and Macdonald 2015; McCarthy et al. 2015; Grassman et al. 2016). Clearly, more research is needed on population trends of medium and small-sized felids in Southeast Asia, especially in areas where tiger and leopard have become extirpated. Such research would help show whether smaller felids benefit from the extirpation of large felids, or whether the same factors that caused the extirpation of large felids are negatively impacting small felid populations as well.
In Lao People's Democratic Republic, hereafter Laos, the Nam Et-Phou Louey National Park (NEPL) is one of the largest and best protected areas in the country. Surveys in 2003–2004 showed that the NEPL had an especially high diversity of mammals, including all six felid species mentioned above (Johnson et al. 2006). In fact, NEPL was identified as a tiger priority site because it contained one of the most important tiger populations in all of Laos, Cambodia, and Vietnam (Walston et al. 2010). However, more recent surveys in 2013–2017 showed that tiger and leopard have become extirpated in NEPL, primarily due to an exponential increase in snaring (Johnson et al. 2016; Rasphone et al. 2019). The loss of tiger and leopard on the remaining felid community is unknown because the density of smaller felids has never been determined in NEPL.
The goal of this study is to determine the density trends of clouded leopard, marbled cat, and leopard cat in NEPL, and to conclude if either mesopredator release or poaching is affecting their densities and interactions. Based on the mesopredator release hypothesis, we predict that clouded leopard densities will increase following the extirpation of tiger, which occurred during the first year of the study. If that prediction is fulfilled, we predict that densities of marbled cat and leopard cat will decrease during the study due to the increasing numbers of clouded leopard. Alternatively, if clouded leopard densities decrease, then snaring could be overriding any potential mesopredator release for this species; thus, snaring might be suppressing densities of marbled cat and leopard cat as well. Given that clouded leopard and marbled cat are a conservation concern, determining which factors most influence their densities will help inform conservation efforts in NEPL and throughout the region. Finally, we also conducted a literature review of previous density estimates for these three felid species, so that our density estimates could be put in context of the entire species’ ranges.
Material and methods
Study area
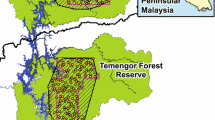
The Nam Et-Phou Louey National Park (NEPL) is located in the northern highlands of Laos (between latitude 19°50′-20°50′N and longitude 103°00′-103°53′E), bordering Vietnam along its northern boundary (Fig. 1). It spans 5,969 km2 of mountainous terrain across seven districts and three provinces (Houaphan, Luang Prabang, and Xiengkhuang). The topography of NEPL is rugged, with elevation ranging from 400–2,257 m, of which > 60% is above 1,000 m and 91% with slopes > 12% (Johnson 2012). Vegetation is largely dominated by dry evergreen and semi-evergreen forests. However, about a third of the park’s forests are degraded with a canopy cover of less than 20%. The climate is tropical monsoonal with a rainy season lasting from May to October followed by a distinct dry season for the rest of the year, and annual rainfall ranges from 1400 to 1800 mm (Johnson 2012). The fauna is described in detail by Rasphone et al. (2019). The NEPL is divided into two protected zones; a totally protected (core) zone where human access is prohibited and a peripheral managed use zone where specified livelihood activities of local people are permitted following park regulations (Fig. 1). There are 34 villages in the management zone and edge of the core zone in NEPL, but none within our survey blocks (Fig. 1).

The Nam Et—Phou Louey National Park (NEPL), Laos, showing the three blocks that were camera-trap surveyed from 2013 to 2017 (NPNV = Nam Poung—Na Vaen; PD = Pha Daeng; PS = Phoupha—Siphou). The inset map shows the location of the NEPL in Laos
Camera trapping
We used systematic camera trap surveys to estimate densities of three felid species in the NEPL. From 2013 to 2017, we carried out three surveys in each of three blocks within the NEPL core zone: Nam Poung—Na Vaen (NPNV), Pha Daeng (PD), and Phoupha—Siphou (PS; Fig. 1; Table 1). In 2014 we also surveyed a fourth block in the eastern part of the core zone (Rasphone et al. 2019), but because this block was not surveyed in subsequent years due to limited resources, we excluded data from this block in our analyses. During each year, blocks were surveyed sequentially primarily during the dry season.
For each of block (~ 200 km2 each), cameras were set in pairs at 80 stations (a total of 160 cameras) in a regular grid with 1.0–1.5 km spacing between stations for 49–54 days (Table 1). The distance between stations was based on previous camera-trapping studies that targeted clouded leopards, as this distance was shown to be adequate for estimating density of this species given its home range size (Singh and Macdonald 2017; Naing et al. 2018). This distance also was shown to be adequate for estimating densities of marbled cats and leopard cats based on their home range sizes (Mohamed et al. 2013; Srivathsa et al. 2015; Hearn et al. 2016; Singh and Macdonald 2017). We assumed our survey period of 1.5–2 months was adequate for population closure in the felid populations, because previous studies on felids suggested survey periods of 2–3 months for population closure (Karanth and Nichols 1998; Weingarth et al. 2015; Rostro-García et al. 2018). A recent study suggests that longer survey periods can increase detection success and lead to lower estimation bias for slow-living species, unless the survey period overlaps the reproductive season of the target species (Dupont et al. 2019). In our case, we chose to sample more areas of the park by surveying different blocks sequentially within the dry season, instead of setting cameras in one block for the entire period. We feel that this was a good compromise given our limited number of cameras (n = 160), lack of knowledge about the reproductive season of the felid species, and the relatively large area of the NEPL core zone. We used a mixture of infrared camera trap models: CuddeBack Ambush IR (infrared)–Model 1187 (CuddeBack Digital, USA), Reconyx HYPERFIRE HC500 (Reconyx, USA), and MAGINON—WK 3 HD (Supra Foto-Elektronik-Vertriebs-GmbH, Germany).
Elevation of stations ranged from 629–2,185 m (Table 1); 16% of the stations were below 1000 m, 16% were between 1000 to 1500 m, and 68% were above 1500 m. About 63% of the stations were in closed evergreen and high density mixed deciduous forests while the rest were in shrub/bamboo forest, grassland and regeneration/agricultural areas. Elevation ranges and habitat were relatively similar among the survey blocks (Table 1). The cameras were placed mostly along ridge lines and animal trails, and at points where trails met streams. Each camera was mounted onto a tree and its motion sensor was set at 35 cm above ground (approximate shoulder height of clouded leopard).
Analysis
We used spatially explicit capture-recapture (SCR) models to estimate population density of the three felid species from our camera-trap data from three survey periods (2013–2014, 2015–2016, and 2017). We identified individuals of the clouded leopard, marbled cat and leopard cat by comparing and matching their pelage patterns from all the camera trap photographs. Records with both right and left flank photographs were matched first and verified based on the location, date and time of encounters before assigning their final identities. In a second step we matched all encounters where we only had either the left or the right flanks. To estimate density, the encounters of all individuals with full identities along with encounters of the flank that had the largest number of captures (either only lefts or only rights) were used. Images of a fourth felid species, the Asian golden cat, were captured, but distinguishing among individuals of this species is difficult due to lack of unique pelage markings; therefore, this species was not included in the analysis.
The densities of the clouded leopard, leopard cat and marbled cat were estimated using a SCR model in a Bayesian framework adapted from Royle and Gardner (2011). While open-population SCR models exist that can directly model survival and recruitment (Gardner et al. 2010), we had too few re-captures of individuals across years to use these models. Instead, we extended the basic SCR model to analyse data from three survey years by stacking three independent models that can share parameters. Given similar habitat types and our use of a standardized survey design, we assumed detection parameters were comparable across sampling blocks and survey years. This allowed us to hold \({\lambda }_{0}\) (the probability of capture when the distance between the animal’s activity center and the trap is zero) and σ (the movement parameters determining the rate of decline in detection probability as function of distance from activity center to the trap) constant for each species across the three years while allowing for a difference in density. Sharing parameters across survey years in this manner should lead to improved parameters estimates, particularly when recapture data from individual sampling blocks or years are too limited to analyse separately (Sollmann et al. 2014). We did not consider sex as a covariate because the sex of many individuals was unknown (Table 2), therefore only the basic model described above was used for the density estimations. In our analysis, we used buffers of 15 km for clouded leopard and 10 km for both marbled cat and leopard cat to define habitat masks required for our SCR models.
We implemented the models in a Bayesian framework and ran them in JAGS (Plummer 2003) using jagsUI in R (R Core Team 2017). We used a half-normal detection function and a Poisson encounter model. The Markov Chain Monte Carlo (MCMC) algorithm was run with 3 MCMC chains with 100, 000 iterations and a thinning rate of 10 after discarding the first 50,000 samples.
After estimating a density for each year, we then calculated the trend by taking the ratio between the first and the last survey periods (D1: 2013/14, D2: 2017).
An important advantage of implementing the model in a Bayesian framework is the ability to obtain a posterior distribution on the change in density, enabling us to derive a point estimate of trend as well as drawing probabilistic conclusions of the genre “We are 90% certain that the population has increased/decreased by 30%”.
Results
The cumulative survey effort across years and blocks was 29,244 trap nights (Table 1). Camera theft increased across years for all blocks (Table 1), and the overall percentage of camera theft was 5.0%, 10.5% and 15.8% of stations per year. The surveys yielded photographs of 10–19 individuals/year for clouded leopards, 9–18 individual/year for leopard cats, and 11–19 individuals/year for marbled cats (Table 2). Across species, only 1–4 individuals were recaptured during the second year, and 0–2 individuals were recaptured the third year (Table 2), indicating high annual turnover in the population for all three species. Only one individual, a clouded leopard, was photographed during all three survey years. Naïve occupancy across years ranged from 7–22% for clouded leopard, 6–31% for leopard cat, and 5–14% for marbled cat, and naïve occupancy was lowest in the third year of the survey for all three felids (Table 2).
In our density analysis, we assessed model convergence based on Gelman-Rubin diagnostics and the Rhat values from MCMC results (at convergence, Rhat = 1) and found that all SCR models for the three species had successfully converged. For the clouded leopard, the posterior mean densities for the first and second survey years were similar, whereas density appeared to decrease the third survey year (Fig. 2; Table 3). The average density estimate across years was 1.70 ± 0.30 individuals/100 km2. Similarly, for the leopard cat, the estimated mean densities were relatively similar for the first two survey years, whereas density appeared to decline the third survey year (Fig. 2; Table 3). The average density across years was 1.50 ± 0.30 individuals/100 km2. For the marbled cat, its estimated density declined steadily during all three survey years (Fig. 2; Table 3), and its average density across years was 3.80 ± 0.70 individuals/100 km2.

Density (individuals/100 km2) trends of clouded leopard, leopard cat, and marbled cat based on three camera-trap surveys conducted from 2013 to 2017 in Nam Et—Phou Louey National Park, Laos. Bars represent 95% credible intervals for the Bayesian density estimates
Based on the posterior distribution of the change in density, there was a 90% probability of a decline in density for the clouded leopard, 93% for the leopard cat, and 83% for the marbled cat. Analysis indicated that we were 83% certain that the clouded leopard population decreased at least 10% during the study. Similarly, we were 88% certain that the leopard cat population declined at least 10%, whereas we were 75% certain that the marbled cat population declined at least 10%.
The estimates of movement parameters were similar for the clouded leopard and leopard cat, which both were more than twice as high as for the marbled cat (Table 3). The baseline encounter rates were similar for all three species (Table 3).
Discussion
We present the first density estimates of clouded leopard, leopard cat and marbled cat in Laos, and the first density trends of these species from anywhere in Asia. Compared to previous studies, our overall density estimate for clouded leopard was within the range reported for this species, whereas overall density estimates for both leopard cat and marbled cat were the lowest ever reported for these species (Table 4). Across four years in one of the most important protected areas in Southeast Asia, we found high (≥ 90%) probability of density declines for clouded leopard and leopard cat, with evidence, albeit a lower (83%) probability, of a decline for marbled cat. The declining trend for all three species was consistent with the high annual turn-over of the individuals we recorded, indicating a high annual mortality for all three species. Additionally, naïve occupancy was lowest for all three felid species during the last survey year. These declines were against a background of evidence that snaring began and exponentially increased in NEPL from 2008 to 2012 (Johnson et al. 2016). Snaring has not been quantified in NEPL since 2013, however, the opinion amongst protected area staff and our field teams in NEPL is that snaring has continued to proliferate since the exponential increase recorded from 2008 to 2012, which is consistent with trends of increased snaring recorded throughout Southeast Asia (Gray et al. 2017, 2018). In support of this view, we photographed poachers on numerous occasions and the increasing theft of cameras across years in all blocks likely indicated an increasing trend of illegal human activity in NEPL during our study. Therefore, we conclude that a continuous increase in snaring within NEPL for the illegal wildlife trade was the most likely cause for the decline of all three felid species.
We are unsure if poachers in NEPL targeted medium and small-sized felids, or if snaring was indiscriminate and felids were simply by-catch in snares set for more common species, such as muntjac and wild pig. Clouded leopards are targeted by poachers for the illegal wildlife trade (D’Cruze and Macdonald 2015), and their bones are used as substitutes for tigers in traditional Asian medicine (Oswell 2010; Grassman et al. 2016). Consequently poaching has been shown to decrease clouded leopard densities in Thailand (Petersen et al. 2020). Even smaller felids such as marbled cat and leopard cat are valued for their skin, meat, and bones (Ross et al. 2015, 2016). After the extirpation of the tiger in NEPL, we suspect that most snaring was relatively indiscriminate, which is typical of that reported for other areas of Southeast Asia (Gray et al. 2017, 2018), with a primary focus on medium and small-sized ungulates and carnivores. Although clouded leopard and marbled cat are considered more arboreal than leopard cat (Grassman et al. 2016; Ross et al. 2016), the former two species used trails extensively in NEPL, and we recorded a similar number of all three felid species in our cameras, which were set only along trails. Thus, despite presumed difference in behaviour and ecology among the felid species, their susceptibility to snares likely was similar in NEPL probably because all three species appeared to regularly travel along animal trails, which is typically where poachers set snares. We expect the densities of all three felid species to continue to decrease unless snaring is reduced within NEPL.
We found no evidence of mesopredator release following the extirpation of tiger in NEPL (Rasphone et al. 2019), but we suspect that any such effect could have been masked by the impact of snaring. Based on experimental research on other carnivores (Kamler et al. 2003), the extirpation of large felids should have resulted in an immediate increase in clouded leopard if mesopredator suppression was occurring. Our literature review showed that the highest densities of clouded leopard were reported from sites where tiger and leopard were absent (Table 4), which is consistent with the mesopredator release hypothesis. This is supported by research in India and Thailand, which showed the highest reported densities of clouded leopard were thought to be due to mesopredator release because the local tiger and leopard populations had become extirpated or had very low densities (Singh and Macdonald 2017; Petersen et al. 2020). Regarding marbled cat and leopard cat, the pattern across studies appeared to show that the highest reported densities of these felids also were from sites where tiger and leopard were absent (Table 4) which is unexpected under the mesopredator release hypothesis that predicts small felids should benefit from the presence of large felids via the suppression of mesofelids. Nonetheless, consistent with the pattern shown in Table 4, tiger and leopard regularly kill and consume medium and small-sized felids (Hayward et al. 2006; Simcharoen et al. 2018), including leopard cat (Rostro-García et al. 2018), suggesting that all smaller felid species might benefit numerically from the absence of large felids. Overall, it appears that densities of medium and small felids tend to be higher at sites were large felids are absent, unless poaching is relatively high. However, future research is needed to test this hypothesis.
Alternatively, factors other than poaching, such as habitat loss or prey declines, could have caused the decline in medium and small-sized felids in NEPL. Although habitat loss and fragmentation reportedly are significant threats to both clouded leopards and marbled cats in Southeast Asia (Grassman et al. 2016; Ross et al. 2016; Cushman et al. 2017), there was no large scale logging or other major new human disturbance of habitats within NEPL during our study. Additionally, recent range-wide habitat modelling of clouded leopard indicated that NEPL is still suitable for them (Macdonald et al. 2019). Therefore, habitat loss was not likely a factor in the decline of felids in NEPL. Alarmingly, our density estimate for the leopard cat was the lowest recorded for this species, which raises further concern about the impact of snaring in NEPL, especially given that leopard cat is a generalist species that attain relatively high densities in other parts of their range, even in habitats that are fragmented and heavily altered by humans (Mohamed et al. 2013; Chua et al. 2016; Petersen et al. 2019).
Clouded leopards reportedly have a diverse diet that includes small- and medium-sized ungulates, primates, brush-tailed porcupines (Atherurus macrourus), pangolins (Manis javanica), squirrels, and pheasants (Nowell and Jackson 1996; Grassman et al. 2005). It is doubtful that all these prey species significantly declined in our study. Even if all of these potential prey species declined during our study, then clouded leopards would have declined in concert with them only if prey populations were limiting clouded leopard densities. Given the relatively low density of clouded leopards in NEPL, and the high diversity of potential prey that we recorded, it is highly unlikely that the clouded leopard population was limited by prey. The diets of marbled cats and leopard cats also are diverse, and reportedly consist primarily of small rodents (Nowell and Jackson 1996; Kamler et al. 2020; Rostro-García et al. 2021). It is doubtful that all species of small rodents significantly declined during our study, especially because small rodents are not targeted by poachers because these species are too small to be captured in snares. Also, given the low densities of marbled cats and leopard cats in our study, and the relatively high diversity of small rodents potentially occurring in NEPL (about 30 species from 16 genera; Lunde and Son 2001), it is highly unlikely that small rodent abundance was limiting the populations of either small felid species.
Immediate actions are needed to reduce poaching in NEPL, although effective anti-poaching programs typically are difficult to implement. Nonetheless, in other areas of Southeast Asia, poaching within protected areas has been reduced using several methods. In Kuiburi National Park, Thailand, a community outreach program resulted in a decrease in poaching and recovery of hunted wildlife populations within the park (Steinmetz et al. 2014). In Huai Kha Khaeng Wildlife Sanctuary, Thailand, the development of systematic and intensified enforcement regime was developed, which included the collection of detailed data by rangers in the field, along with increased patrol days and patrol distances (Duangchantrasiri et al. 2016). This effective enforcement program was successful in reducing poaching and maintaining the tiger density in the sanctuary (Duangchantrasiri et al. 2016). However, snare detectability by rangers in tropical forests of Southeast Asia is relatively low (Ibbett et al. 2020), consequently snaring can increase exponentially in a park even with greater funding for enforcement and increases in days patrolled, as observed in NEPL (Johnson et al. 2016). Therefore, we recommend that the NEPL managers implement a more systematic and intensified snare removal program, in concert with extensive community outreach and engagement of local people to prevent the setting of snares. Carrying out this two-pronged strategy may be the only hope to avert the extirpations of the remaining felid species in NEPL.
References
Bashir T, Bhattacharya T, Poudyal K, Sathyakumar S, Qureshi Q (2013) Estimating leopard cat Prionailurus bengalensis densities using photographic captures and recaptures. Wildl Biol 19:462–472
Borah J, Sharma T, Das D et al (2013) Abundance and density estimates for common leopard Panthera pardus and clouded leopard Neofelis nebulosa in Manas National Park, Assam, India. Oryx 48:149–155
Chapron G, Miquelle DG, Lambert A, Goodrich JM, Legendre S, Clobert J (2008) The impact on tigers of poaching versus prey depletion. J Appl Ecol 45:1667–1674
Chua MAH, Sivasothi N, Meier R (2016) Population density, spatiotemporal use and diet of the leopard cat (Prionailurus bengalensis) in a human-modified succession forest landscape of Singapore. Mammal Res 61:99–108
Crabtree RL, Sheldon JW (1999) Coyotes and canid coexistence in Yellowstone. In: Clark TM, Curlee AP, Minta SC, Kareiva PM (eds) Carnivores in ecosystems: the Yellowstone experience. Yale University Press, New Haven, pp 127–163
Cushman SA, Macdonald EA, Landguth EL, Malhi Y, Macdonald DW (2017) Multiple-scale prediction of forest loss risk across Borneo. Landsc Ecol 32:1581–1598
D’Cruze N, Macdonald DW (2015) Clouded in mystery: the global trade in clouded leopards. Biodivers Conser 24:3505–3526
Duangchantrasiri S, Umponjan M, Simcharoen S et al (2016) Dynamics of a low-density tiger population in Southeast Asia in the context of improved law enforcement. Conserv Biol 30:639–648
Duckworth JW, Lynam AJ, Breitenmoser-Würsten C (2014) Non-Panthera cats in South-east Asia: present knowledge and recommendations. Cat News S8:62–67
Dupont P, Milleret C, Gimenez O, Bischof R (2019) Population closure and bias-precision trade-off in spatial capture-recapture. Methods Ecol Evol 10:661–672
Foster VC, Sarmento P, Sollmann R et al (2013) Jaguar and puma activity patterns and predator-prey interactions in four Brazilian biomes. Biotropica 45:373–379
Gardner B, Reppucci J, Lucherini M, Royle JA (2010) Spatially explicit inference for open populations: estimating demographic parameters from camera-trap studies. Ecology 91:3376–3383
Grassman L, Lynam A, Mohamad S et al (2016) Neofelis nebulosa. The IUCN Red List of Threatened Species 2016:e.T14519A97215090. https://dx.doi.org/https://doi.org/10.2305/IUCN.UK.2016-1.RLTS.T14519A97215090.en. Accessed 10 Jan 2020
Grassman LI, Tewes ME, Silvy NJ, Kreetiyutanont K (2005) Ecology of three sympatric felids in a mixed evergreen forest in north-central Thailand. J Mammal 86:29–38
Gray TN, Hughes AC, Laurance WF et al (2018) The wildlife snaring crisis: an insidious and pervasive threat to biodiversity in Southeast Asia. Biodivers Conserv 27:1031–1037
Gray TN, Lynam AJ, Seng T, Laurance WF, Long B, Scotson L, Ripple WJ (2017) Wildlife-snaring crisis in Asian forests. Science 355:255–256
Harihar A, Pandav B, Goyal SP (2011) Responses of leopard Panthera pardus to the recovery of a tiger Panthera tigris population. J Appl Ecol 48:806–814
Hayward MW, Henschel P, O’Brien J, Hofmeyr M, Balme G, Kerley GIH (2006) Prey preferences of the leopard (Panthera pardus). J Zool (Lond) 270:298–313
Hearn AJ, Ross J, Bernard H, Bakar SA, Hunter LTB, Macdonald DW (2016) The first estimates of marbled cat Pardofelis marmorata population density from Bornean primary and selectively logged forest. PLoS ONE 11:e0151046
Horne JS, Haines AM, Tewes ME, Laack LL (2009) Habitat partitioning by sympatric ocelots and bobcats: implications for recovery of ocelots in southern Texas. Southwest Nat 54:119–126
Hughes AC (2017) Understanding the drivers of Southeast Asian biodiversity loss. Ecosphere 8:e01624
Hunter L, Barrett P (2011) A field guide to the carnivores of the world. New Holland, London
Johnson A (2012) A landscape summary for the Nam Et-Phou Louey National Protected Area, Lao PDR. In: Sunderland TCH, Sayer J, Minh-Ha H (eds) Evidence-based conservation: lessons from the lower Mekong. Earthscan, London, pp 73–90
Johnson A, Goodrich J, Hansel T et al (2016) To protect or neglect? Design, monitoring, and evaluation of a law enforcement strategy to recover small populations of wild tigers and their prey. Biol Conserv 202:99–109
Johnson A, Vongkhamheng C, Hedemark M, Saithongdam T (2006) Effects of human–carnivore conflict on tiger (Panthera tigris) and prey populations in Lao PDR. Anim Conserv 9:421–430
Kamler JF, Ballard WB, Gilliland RL, Lemons PR, Mote K (2003) Impacts of coyotes on swift foxes in northwestern Texas. J Wildl Manage 63:317–323
Kamler JF, Inthapanya X, Rasphone A, Bousa A, Vongkhamheng C, Johnson A, Macdonald DW (2020) Diet, prey selection, and activity of Asian golden cats and leopard cats in northern Laos. J Mammal 101:1267–1278
Kamler JF, Stenkewitz U, Macdonald DW (2013) Lethal and sublethal effects of black-backed jackals on cape foxes and bat-eared foxes. J Mammal 94:295–306
Karanth KU, Nichols JD (1998) Estimation of tiger densities in India using photographic captures and recaptures. Ecology 79:2852–2862
Kumar U, Awasthi N, Qureshi Q, Jhala Y (2019) Do conservation strategies that increase tiger populations have consequences for other wild carnivores like leopards? Sci Rep 9:14673
Lekagul B, McNeely JA (1977) Mammals of Thailand. Association for the Conservation of Wildlife, Bangkok
Levi T, Wilmers CC (2012) Wolves–coyotes–foxes: a cascade among carnivores. Ecology 93:921–929
Lunde D, Son NT (2001) An identification guide to the rodents of Viet Nam. American Museum of Natural History, New York
Luo SJ, Zhang Y, Johnson WE et al (2014) Sympatric Asian felid phylogeography reveals a major Indochinese-Sundaic divergence. Mol Ecol 23:2072–2092
Macdonald DW, Bothwell HM, Kaszta Ż et al (2019) Multi-scale habitat modelling identifies spatial conservation priorities for mainland clouded leopards (Neofelis nebulosa). Divers Distrib 25:1639–1654
Macdonald DW, Loveridge AJ, Nowell K (2010) Dramatis personae: An introduction to the wild felids. In: Macdonald DW, Loveridge AJ (eds) Biology and Conservation of Wild Felids. Oxford University Press, Oxford, pp 3–58
McCarthy J, Dahal S, Dhendup T et al. (2015) Catopuma temminckii (errata version published in 2016). The IUCN Red List of Threatened Species 2016:e.T T4038A97165437. https://doi.org/https://doi.org/10.2305/IUCN.UK.2015-4.RLTS.T4038A50651004.en. Accessed 10 Jan 2020
McDougal C (1988) Leopard and tiger interactions at Royal Chitwan National Park. Nepal J Bombay Nat Hist Soc 85:609–611
Mohamad SW, Rayan DM, Christopher WCT, Hamirul M, Mohamed A, Lau CF, Siwan ES (2015) The first description of population density and habitat use of the mainland clouded leopard Neofelis nebulosa within a logged-primary forest in South East Asia. Popul Ecol 57:495–503
Mohamed A, Sollmann R, Bernard H et al (2013) Density and habitat use of the leopard cat (Prionailurus bengalensis) in three commercial forest reserves in Sabah, Malaysian Borneo. J Mammal 94:82–89
Mondal K, Gupta S, Bhattacharjee S, Qureshi Q, Sankar K (2012) Response of leopards to re-introduced tigers in Sariska Tiger Reserve, Western India. Int J Biodivers Conserv 4:228–236
Mukherjee S, Singh P, Pinto Silva A et al (2019) Activity patterns of the small and medium felid (Mammalia: Carnivora: Felidae) guild in northeastern India. J Threat Taxa 11:13432–13447
Nagy-Reis MB, Iwakami VHS, Estevo CA, Setz EZF (2019) Temporal and dietary segregation in a neotropical small-felid assemblage and its relation to prey activity. Mamm Biol 95:1–8
Naing H, Ross J, Burnham D, Htun S, Macdonald DW (2019) Population density estimates and conservation concern for clouded leopards Neofelis nebulosa, marbled cats Pardofelis marmorata and tigers Panthera tigris in Htamanthi Wildlife Sanctuary, Sagaing, Myanmar. Oryx 53:654–662
Nowell K, Jackson P (1996) Wild cats: status survey and conservation action plan. IUCN, Gland
Odden M, Wegge P (2009) Kill rates and food consumption of leopards in Bardia National Park. Nepal Mammal Res 54:23–30
Oswell AH (2010) The big cat trade in Myanmar and Thailand. TRAFFIC Southeast Asia, Petaling Jaya
Penjor U, Macdonald DW, Wangchuk S, Tandin T, Tan CKW (2018) Identifying important conservation areas for the clouded leopard Neofelis nebulosa in a mountainous landscape: Inference from spatial modeling techniques. Ecol Evol 8:4278–4291
Petersen WJ, Savini T, Steinmetz R, Ngoprasert D (2019) Estimating Leopard Cat Prionailurus bengalensis Kerr, 1792 (Carnivora: Felidae) density in a degraded tropical forest fragment in northeastern Thailand. J Threat Taxa 11:13448–13458
Peterson WJ, Steinmetz R, Sribuarod K, Ngoprasert D (2020) Density and movements of mainland clouded leopards (Neofelis nebulosa) under conditions of high and low poaching pressure. Glob Ecol Conserv 23:e01117
Plummer M (2003) JAGS: A program for analysis of Bayesian graphical models using Gibbs sampling. In: Proceedings of the 3rd International Workshop on Distributed Statistical Computing, vol 124, pp 1–10
Prugh LR, Stoner CJ, Epps CW, Bean WT, Ripple WJ, Laliberte AS, Brashares JS (2009) The rise of the mesopredator. Bioscience 59:779–791
R Core Team (2017) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rasphone A, Kamler JF, Macdonald DW (2020) Temporal partitioning by felids, dholes, and their potential prey in northern Laos. Mammal Res 65:679–689
Rasphone A, Kéry M, Kamler JF, Macdonald DW (2019) Documenting the demise of tiger and leopard, and the status of other carnivores and prey, in Lao PDR’s most prized protected area: Nam Et - Phou Louey. Glob Ecol Conserv 20:e00766
Ripple WJ, Abernethy K, Betts MG et al (2016) Bushmeat hunting and extinction risk to the world’s mammals. R Soc Open Sci 3:160498
Ross J, Brodie J, Cheyne S et al (2016) Pardofelis marmorata. The IUCN Red List of Threatened Species 2016:e.TI6218A97164299. https://dx.doi.org/https://doi.org/10.2305/IUCN.UK.2016-1.RLTS.TI6218A97164299.en. Accessed 10 Jan 2020
Ross J, Brodie J, Cheyne S et al (2015) Prionailurus bengalensis. The IUCN Red List of Threatened Species 2105:e.T18146A50661611. https://dx.doi.org/https://doi.org/10.2305/IUCN.UK.2016-1.RLTS.T18146A50661611.en. Accessed 10 Jan 2020
Rostro-García S, Kamler JF, Ash E et al (2016) Endangered leopards: Range collapse of the Indochinese leopard (Panthera pardus delacouri) in Southeast Asia. Biol Conserv 201:293–300
Rostro-García S, Kamler JF, Clements GR, Lynam AJ, Naing H (2019) Panthera pardus spp. delacouri. The IUCN Red List of Threatened Species 2019:e.T124159083A124159128. https://doi.org/https://doi.org/10.2305/IUCN.UK.2019-3.RLTS.T124159083A163986056.en. Accessed 10 Jan 2020
Rostro-García S, Kamler JF, Crouthers R et al (2018) An adaptable but threatened big cat: density, diet and prey selection of the Indochinese leopard (Panthera pardus delacouri) in eastern Cambodia. R Soc Open Sci 5:171187
Rostro-García S, Kamler JF, Hunter LTB (2015) To kill, stay or flee: the effects of lions and landscape factors on habitat and kill site selection of cheetahs in South Africa. PLoS ONE 10:e0117743
Rostro-García S, Kamler JF, Minge A et al (2021) Small cats in big trouble? Diet, activity, habitat use, and occupancy of jungle cats and leopard cats in threatened dry deciduous forests, Cambodia. Ecol Evol 11:In press.
Royle JA, Gardner B (2011) Hierarchical spatial capture–recapture models for estimating density from trapping arrays. In: O’Connell A, Nichols JD, Karanth KU (eds) Camera traps in animal ecology: methods and analyses. Springer, New York, pp 163–190
Seidensticker J (1976) On the ecological separation between tigers and leopards. Biotropica 8:225–234
Selvan M, Lyngdoh S, Gopi Govindan V, Habib B (2014) Density estimation of leopard cat Prionailurus bengalensis using capture-recaptures sampling in lowland forest of Pakke Tiger Reserve, Arunachal Pradesh, India. Mammalia 78:555–559
Simcharoen A, Simcharoen S, Duangchantrasiri S, Bump J, Smith JLD (2018) Tiger and leopard diets in western Thailand: Evidence for overlap and potential consequences. Food Webs 15:e00085
Singh P, Macdonald DW (2017) Populations and activity patterns of clouded leopards and marbled cats in Dampa Tiger Reserve, India. J Mammal 98:1453–1462
Sollmann R, Linkie M, Haidir IA, Macdonald DW (2014) Bringing clarity to the clouded leopard Neofelis diardi: first density estimates from Sumatra. Oryx 48:536–539
Soulé ME, Bolger DT, Alberts AC, Wrights J, Sorice M, Hill S (1988) Reconstructed dynamics of rapid extinctions of chaparral-requiring birds in urban habitat islands. Conserv Biol 2:75–92
Srivathsa A, Parameshwaran R, Sharma S, Karanth KU (2015) Estimating population sizes of leopard cats in the Western Ghats using camera surveys. J Mammal 96:742–750
Steinmetz R, Srirattanaporn S, Mor-Tip J, Seuaturien N (2014) Can community outreach alleviate poaching pressure and recover wildlife in South-East Asian protected areas? J Appl Ecol 51:1469–1478
Walston J, Robinson JG, Bennett EL et al (2010) Bringing the tiger back from the brink-the six percent solution. PLoS Biol 8:e1000485
Weingarth K, Zeppenfeld T, Heibl C, Heurich M, Bufka L, Daniszová K, Müller J (2015) Hide and seek: extended camera-trap session lengths and autumn provide best parameters for estimating lynx densities in mountainous areas. Biodivers Conserv 24:2935–2952
Acknowledgements
We thank the Wildlife Conservation Society, Laos, and the Nam Et—Phou Louey National Park for their extensive in-kind support and for their efforts towards planning and implementing the research. For data collection and management, we thank Mr. Phonesouk Chanthalavong for his excellent assistance in field work preparation and implementation, and data management. We further thank Mike Meredith, and Ngumbang Juat for their guidance. The research was supported by a grant to DWM from the Robertson Foundation and National Geographic Society, for which we are deeply grateful.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Grzegorz Mikusinski.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rasphone, A., Kamler, J.F., Tobler, M. et al. Density trends of wild felids in northern Laos. Biodivers Conserv 30, 1881–1897 (2021). https://doi.org/10.1007/s10531-021-02172-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-021-02172-0