
Abstract
An agro-environmental payment for the management of the so-called ‘Biodiversity Promotion Areas’ (BPA) has been used to accomplish biodiversity conservation goals in Switzerland. These areas have been managed according to specific limitations on mowing dates and fertilizers. We assessed the regional-scale effectiveness of BPA implementation within Ticino Canton by answering the following questions: (i) is plant species diversity higher in BPA than in conventionally managed grasslands (CMG)? (ii) which are the differences between BPA and CMG in terms of climatic, topographical, ecological, and vegetation variables? (iii) which vegetation types, functional groups, and plant species are specifically related to BPA? A total of 242 vegetation surveys (64 in BPA and 178 in CMG, respectively) was carried out in 55 farms and the main climatic and topographic features were assessed. Differences in terms of plant diversity, ecological indicator and pastoral values, species functional groups, vegetation types, and indicator species between BPA and CMG were assessed. The BPA harboured a higher plant diversity. They were located in steeper areas, at higher elevations, and characterised by lower soil nutrient content, mowing tolerance, and pastoral value than CMG. Dry meadow species number and cover were higher in BPA, while nutrient-rich meadow species number was higher in CMG. The species associated to BPA belonged to a wider range of functional groups and 38% of them belonged to the national list for biodiversity promotion in agriculture, whereas no species associated to CMG belonged to that list. Thus, our results confirmed the effectiveness of BPA for biodiversity conservation for the Southern Swiss Alps.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Semi-natural extensive grasslands of temperate regions are amongst the most species-rich terrestrial ecosystems on the Earth (Wilson et al. 2012). A remarkable number of grassland specialists, among which many endemic and threatened species, composes a significant proportion of this high biodiversity, for both flora and fauna (Nagy 2009; WallisDeVries and Van Swaay 2009; Tocco et al. 2013; Fournier et al. 2017; Kadereit 2017). These highly diverse open habitats are the result of long-term extensive mowing and grazing regimes and most of the extreme-richness sites are managed with no fertilisation and no more than one cut per year (Dengler et al. 2014). Indeed, regular and low-intensive biomass removal reduces the competitive advantage that fast-growing plant species have in highly disturbed habitats, allowing their coexistence with low-growing species and resulting in an overall increase in species richness (Roleček et al. 2014). Moreover, species diversity is generally promoted by the spatial heterogeneity of environmental conditions (e.g. variation in temperature, solar radiation, soil depth and type, and water and nutrient availability) occurring over short distances, a common situation in mountain areas (Körner 2003). Indeed, a general increase in the richness of vascular plants was recorded from the colline to the mountain belts in Switzerland, which is also the result of the decrease in management intensity occurring at increasing elevations as a consequence of lower grassland productivity (Koordinationsstelle Biodiversitäts-Monitoring Schweiz 2009).
However, during the last century, these species-rich ecosystems have experienced a dramatic loss in their area across the entire Palearctic region, due to both agricultural intensification and abandonment processes (Dengler et al. 2014). In the most accessible and productive sites, extensive semi-natural grasslands have been often converted to arable lands or their management has been intensified by means of synthetic fertilizers and pesticides, increased cutting frequency, higher animal stocking rates, and re-sowing with a limited array of forage species (Manning et al. 2015). Consequently, intensified management has modified their botanical composition, by shifting species-rich plant communities towards less-biodiverse grasslands dominated by a specific functional group of species, namely productive grasses (Hejcman et al. 2012). Indeed, these species are able to adapt to frequent and intense defoliation regimes due to their fast leaf turnover rates, in contrast to nutrient-poor low-growing species (Pittarello et al. 2018). Conversely, in the most marginal and less productive sites, semi-natural grasslands have often been abandoned, since traditional management has become economically unviable under current socio-economic conditions, with an exacerbated trend over European montane regions (Estel et al. 2015). Therefore, the agro-pastoral abandonment has also resulted in widespread vegetation cover and composition changes, with grassland loss because of tree and shrub encroachment (Orlandi et al. 2016).
In recent decades, there has been a growing attention throughout all Europe for the preservation of grassland biodiversity, since it is a non-renewable genetic source related to the provision of several ecosystem services irreplaceable for human society, such as food production, climate regulation, nutrient cycling, pollination, soil protection, water control, wild-fire risk mitigation, and tourism and recreation opportunities (Harrison et al. 2010). In addition, a higher biodiversity generally increases stress tolerance of communities, thus enhancing the protection of ecosystems against disturbances (Hooper et al. 2005). It is also worth mentioning that grasslands are one of the most important soil cover types across all Europe. Indeed, in 2016, the area covered by permanent grasslands at a continental scale amounted to 13.2% of the total surface or 34.3% of the utilized agricultural area of the 28 member states of the European Union plus Switzerland (UST 2018a; EUROSTAT 2019). For all the above-mentioned reasons, nowadays many European agricultural and environmental policies reflect the current societal need of preserving grassland biodiversity (Habitat Directive 1992/43/EEC, Bird Directive 2009/147/EC, EU Regulation 1307/2013).
In countries characterised by a higher percentage of grassland cover, such as Switzerland, this attention has become a real priority in agricultural and environmental policies. Indeed, in 2018, Swiss permanent grasslands (without considering alpine summer pastures) amounted to 607.539 ha, corresponding to 14.7% and 58.1% of total national surface and utilized agricultural area, respectively, with a remarkable proportion of montane meadows and pastures (Swiss National Agricultural Report 2019). Since the beginning of ‘90s, the Swiss government introduced an agro-environmental payment for the management of specific areas (formerly called ‘Ecological Compensation Areas’, nowadays ‘Biodiversity Promotion Areas’, BPA) to accomplish the environmental goals for agriculture in terms of biodiversity conservation (UFAG 2020a). The current legislation sets specific rules to farmers to receive agricultural direct payments, among which the obligation that at least 7% of their agricultural area must be managed as a BPA (Swiss Ordinance 910.13). For grasslands, this means that specific limitations on mowing dates and on the use of fertilizers have to be complied. Under this framework, three grassland BPA are envisaged: (i) extensive meadows, receiving no fertilizer nor manure inputs, (ii) low-intensive meadows, that can receive manure or compost input up to 30 kg N ha−1 year−1, and (iii) extensive pastures, receiving only grazing animal excreta. Meadows and pastures must be mown or grazed at least once a year, respectively, with meadows that must not be mown before July 1st or 15th at lower and higher mountain zones, respectively.
In recent years, some studies have been conducted to assess the effectiveness of this agri-environmental policy (Herzog et al. 2005; Knop et al. 2006; Dietschi et al. 2007; Kampmann et al. 2008; Peter et al. 2009; Riedel et al. 2019). These authors have generally measured positive effects of BPA management on vascular plant diversity conservation, even if with some contrasting results, depending on particular vegetation types or functional groups of species. However, the abovementioned studies have been conducted in the Northern and Eastern-Central Alps, while no specific study focused on the Southern Alps (i.e. Ticino Canton). This region is characterised by different biogeographic and socio-economic conditions and by a peculiar flora, shaped by distinct climatic conditions, which are influenced by the presence of high mountain massifs and the southern Po Basin (Ozenda 1985; Welten and Sutter 1982; Gonseth et al. 2001). Over the last decades, the Ticino Canton has faced a stronger agricultural abandonment process compared to the average Swiss trend, due to a more pronounced decrease in the number of farms, agricultural employees, and livestock units, resulting in a higher reduction in the area covered by grasslands (UST 2018b).
To fill this gap, the current study aims to assess the regional-scale effectiveness of BPA implementation for plant diversity conservation in the Southern Swiss Alps by specifically answering the following questions: (i) is vascular plant species diversity higher in grasslands managed as BPA than in conventionally managed grasslands (CMG)? (ii) which are the differences between BPA and CMG in terms of climatic, topographical, ecological, and vegetation variables? (iii) which vegetation types, functional groups, and plant species are specifically related to the BPA?
Material and methods
Study area
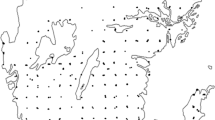
The study was conducted in the Ticino Canton (Switzerland). The surveyed area was characterised by a warm-humid insubric climate, with average annual precipitation ranging from 1318 to 1897 mm and average annual temperature ranging from 5.2 to 12.4 °C, respectively. The vegetation of 55 farms was investigated, exploring a wide range of environmental and management conditions. The farms involved in the study were selected to cover a large area within the Ticino canton, encompassing all the main valleys and a wide range of different climatic, topographic and vegetation conditions, so as to be representative of the regional scale investigated (Fig. 1). For every farm, an average number of 4.6 surveys (standard error: 0.36) was conducted, i.e. one per each homogeneous vegetation type found within the farm. The studied grasslands were located between 197 and 1611 m a.s.l. on slopes ranging from 0.6° to 37.6° and were characterized by an aspect comprised between 6.7° and 338.5°. Grasslands were managed by direct grazing (by dairy cows, sheep, or goats) and/or by cutting for fresh foddering, haymaking, and silage production. Within each farm, grasslands were alternatively considered as BPA (if that management regime had been applied at least for the five previous years) or CMG (if grasslands were not subjected to management restrictions and generally cut and/or grazed two to six times per year, depending on pedo-climatic conditions and potential forage yield).

Location of the 242 vegetation surveys carried out in the Ticino Canton, Switzerland (UTM zone 32°N, WGS84 datum). Black dots identify surveys carried out in Biodiversity Promotion Areas (BPA) while white dots those in conventionally managed grasslands (CMG). Darker shadows of grey correspond to higher elevations
Vegetation surveys
Grassland vegetation of the studied farms was assessed by means of 242 vegetation surveys, conducted once between 2012 and 2016 in either May, June, or July, with a balanced proportion of surveys carried out between BPA and CMG within each year (Fig. 1 and Table 1). A total of 64 and 178 surveys were carried out within BPA and CMG, respectively. Concerning BPA, 43% of them was composed by extensive meadows, 33% by low-intensive meadows, and 24% by extensive pastures, respectively. For each survey, elevation, slope, and aspect were calculated from the Swiss digital height model (Federal Office of Topography 2018) using QGIS 3.10 software (QGIS Development team 2019). Southness, an index of south-facing aspect, was computed as a linear transformation of aspect (180° − |aspect − 180°|; Chang et al. 2004). Average annual precipitation was calculated according to the available data from the nearest meteorological station.
Botanical composition was determined using the vertical point-quadrat method (Daget and Poissonet 1971) along 25-m transects placed within vegetation patches representative of the overall grassland botanical composition. Within each transect, at every 50-cm interval, plant species touching a steel needle were identified and recorded (i.e. 50 points of vegetation measurements per transect). To account for occasional species, likely missing with the point-quadrat method, a complete list of all other species included within a 1-m buffer area surrounding the transect was also recorded. The frequency of occurrence of each plant species recorded (fi = number of occurrences/50 points of vegetation measurement), which is an estimate of species canopy cover, was calculated for each transect and converted to percentage cover (Perotti et al. 2018). To all occasional plant species, not recorded along the vegetation transect but only within the 1-m buffer area around it, a percentage cover of 0.3% was attributed (Pittarello et al. 2017). Species relative abundance (SRAi) was determined in each transect and used to detect the proportion of different species according to Daget and Poissonet (1971):
where SRAi and fi are species relative abundance and frequency of occurrence of species i. Plant species nomenclature followed Aeschimann et al. (2004).
Plant diversity was assessed for every vegetation survey by computing species richness, Shannon diversity index (Magurran 1988), and species evenness (Magurran 2004). Moreover, each plant species was classified according to the indicator values for soil moisture, soil nutrient content (mainly nitrogen), and mowing tolerance identified by Landolt et al. (2010). Each indicator value ranges from 1 (low requirement or tolerance) to 5 (high requirement or tolerance). The mean Landolt’s indicator values for soil moisture, soil nutrient content, and mowing tolerance were computed per each transect by averaging species values weighted on their relative abundance. Additionally, forage pastoral value, a synthetic index based on forage yield and quality ranging from 0 to 100, was calculated following the equation proposed by Daget and Poissonet (1971):
where SRAi and ISQi are the species relative abundance and the index of specific quality (ranging from 1 to 5, and according to Cavallero et al. 2007) of the species i, respectively.
In order to identify different functional groups of species, which are characterised by similar ecological needs, each plant species was classified according to its phytosociological optimum at the class level, as identified by Aeschimann et al. (2004). Then, species in phytosociological classes having physiognomic, ecological and floristic similarity were grouped according to Theurillat et al. (1995). Nine functional groups of species were identified: rocky, terophytic-pioneer, wetland, shrubland–woodland, heathland, edge (including dry and mesophilous edge), ruderal (including annual and perennial ruderal), pasture (including basophilous and acidophilous pasture), and meadow (including dry and nutrient-rich meadow) species. The complete species list and the corresponding phytosociological optima and functional groups are available in Online Resource 1. The total species number and cover accounting for each functional group were calculated for each vegetation survey.
Statistical analyses
Generalized linear mixed models (GLMMs) were performed in order to test for differences between BPA and CMG in terms of plant diversity indexes, climatic, topographic, and ecological features (i.e. elevation, slope, southness, precipitation, soil moisture, soil nutrient content, mowing tolerance, and pastoral value), and number and cover of species belonging to different functional groups. Management regime (i.e. BPA vs CMG) was considered as fixed factor, farm as random factor, and the vegetation survey was the statistical unit. Sequential Bonferroni correction was applied to adjust for multiple comparisons. Shannon diversity index, species evenness, climatic, topographic, and ecological features, and species covers of functional groups, being continuous variables, were modelled with both Gaussian and gamma functions, while species richness, species numbers of functional groups, being count variables, were modelled with both Poisson and negative-binomial functions. Then, the model resulting in the lowest Akaike’s Information Criterion (AIC) value for each analysis was considered as the best fitting one and retained (Zuur et al. 2009). The analyses were carried out with SPSS 25 (IBM Corp. 2017).
Botanical data were classified by hierarchical cluster analysis, in order to assign each vegetation survey to a specific vegetation type. The cluster analysis was performed using Past 4.01 software (Hammer et al. 2001), by setting Pearson’s correlation coefficient to calculate the similarity matrix and the paired group UPGMA as agglomeration method for the between-group linkage. A χ2 test with Yate’s correction for small samples was carried out to assess if each vegetation type was significantly associated to BPA or to CMG using SPSS 25.
An indicator species analysis was carried out to identify the indicator species significantly associated to BPA and CMG (Dufrêne and Legendre 1997). The analysis was carried out with R software (R Core Team 2019) using ‘multipatt’ function in ‘indicspecies’ package (De Cáceres and Legendre 2009; De Cáceres et al. 2010).
Results
A total of 389 plant species, belonging to 28 different phytosociological classes, was found during the vegetation surveys (Online Resource 1). Among them, 59 and 81 species were found exclusively within BPA and CMG (corresponding to 60.3 and 80.4 species as estimated true species richness calculated with Chao 2 estimator; Chao 1987), respectively. Average plant diversity (per 50 m2) was higher in BPA than in CMG, in terms of both species richness and Shannon index, while no difference in species evenness was detected (Table 2).
The BPA were steeper and located at higher elevations and were characterised by significantly lower soil nutrient content, mowing tolerance, and pastoral value than CMG (Table 3). Contrarily, no differences in southness, precipitation, and soil moisture were detected between management regimes.
The BPA were characterised by a higher number of shrubland–woodland, edge, and pasture species (Table 4). In most of the cases, the cover of the species belonging to these groups was also significantly higher in BPA than in CMG. Ruderal species number and cover, instead, were higher in CMG. Dry meadow species number and cover were higher in BPA, while nutrient-rich meadow species were higher in CMG, but only in terms of species cover. When considering total meadow species, the species number was higher in BPA, whereas the species cover was higher in CMG.
The hierarchical cluster analysis individuated six vegetation types (the complete dendrogram with the classification of the vegetation surveys is reported in Online Resource 2). These vegetation types were identified and named by the respective dominant plant species: (i) Dactylis glomerata—Poa trivialis, (ii) Festuca rubra aggr.—Brachypodium rupestre, (iii) Agrostis capillaris, (iv) Festuca rubra aggr., (v) Arrhenatherum elatius, and (vi) Anthoxanthum odoratum. The χ2 test reported significant differences among vegetation types (Pearson χ2 = 38.375; p < 0.001), highlighting that only the F. rubra—B. rupestre type was more represented in BPA than in CMG, whereas all the other five vegetation types were more frequently found in CMG than in BPA (Table 5).
The number of significant indicator species associated to BPA was higher than that of species associated to CMG (Table 6). Among the species associated to BPA, three of them were exclusively found within these grasslands, namely Anthyllis vulneraria, Potentilla rupestris, and Polygonum viviparum. Moreover, the species significantly related to BPA belonged to a wider range of different functional groups than those of CMG and 28 of them were species belonging to the national list of species of priority interest for biodiversity promotion in agriculture, whereas no species associated to CMG belonged to that list.
Discussion
Vascular plant species diversity
Although previous literature already reported positive effects related to the implementation of BPA for Swiss biodiversity conservation, the present research proved for the first time the effectiveness of this policy measure for the Southern Swiss Alps, a region characterised by different vegetation and socio-economic conditions. Indeed, grasslands managed under the extensive BPA system harboured a higher plant diversity than CMG, both in terms of species richness and Shannon diversity index. In particular, the higher plant species richness within BPA was consistent with previous findings reported for the Northern and Central Eastern Alps by Knop et al. (2006), Dietschi et al. (2007), Kampmann et al. (2008), and Peter et al. (2009). The species richness was comparable among all these studies, even if the extent of vegetation plots and the methodologies used for botanical surveys differed. In addition, the proportion of species with different percentage cover was comparable between BPA and CMG, as highlighted by the not significant evenness index of these vegetation communities, whose values were extremely comparable to the ones measured by Peter et al. (2009).
Climatic, topographic, ecological, and vegetation characteristics
Compared to CMG, the BPA were characterised by steeper slopes and higher elevations. These topographical characteristics are consistent with the ones assessed by Kampmann et al. (2008) for 12 different municipalities in Northern and Central-Eastern Alps, who also found BPA to be placed in more remote sites, located on average 300 m above CMG and on more severe slopes as well. Conversely, Dietschi et al. (2007) did not find significant differences on slope, elevation, and accessibility from farmyard sites between BPA and CMG in the valleys of Albula and Surses, in the Eastern part of Switzerland. These results highlighted the remarkable local and regional differences, likely determined by different farm distributions and holding structures, linked to socio-economic conditions and cultural heritages (Bätzing and Bartaletti 2005; Zabel 2019). However, slope and elevation are generally considered as driving forces of management intensity in mountain landscapes, since both factors are strongly related to the pedo-climatic conditions determining forage yield potential and thus the labour necessary for grassland management (Mottet et al. 2006). Kleijn and van Zuijlen (2004) pointed out the preference by farmers for the enrolment of more extensive areas in agri-environmental schemes. In that perspective, the institution of BPA proved to be an effective policy measure, since it encouraged the extensive management of the most marginal sites, which are those more subjected to agro-pastoral abandonment, above all in regions facing a stronger reduction in the number of farms, agricultural employees, and livestock units (Orlandi et al. 2016). This is particularly the case of the Ticino Canton, in which grassland cover decreased by 18% in the period 1979–2018 (against a slight increase of + 1% at a national level in the same period), due to the dramatic reduction in the number of cattle (− 44%), sheep (− 50%), and goats (− 38%) reared (data referred to 1996–2018 period; UST 2018b).
Conversely to slope and elevation, the BPA did not differ from CMG in terms of soil moisture indicator, precipitation, and southness, with the latter variable being consistent with the results of Kampmann et al. (2008). Altogether, these three variables highlighted that dryer ecological conditions were not associated to any specific management regime in the studied region. Therefore, we can exclude that the higher plant diversity measured within BPA was related to ecological conditions favouring the development of dry grasslands, since it is well known that dry grasslands generally harbour the highest plant diversity, as measured also by Dietschi et al. (2007). Contrarily, the lower intensity at which BPA were managed (i.e. the combined effects of low to null fertilization and low cutting/grazing frequency) may have been the main factor determining the higher plant diversity within those areas. Indeed, the lower soil nutrient content and mowing tolerance of BPA likely resulted from the long-term implementation of these extensive management practices, as well as their lower pastoral value, which is a good proxy of management intensity, as indicated by Pittarello et al. (2018). In this paper, based on a high number of vegetation surveys on alpine grasslands across large environmental and management gradients, the authors identified the values for the Landolt’s indicator for soil nutrient content at which plant diversity and pastoral value peak. In the current study, the mean value for this variable within CMG (3.2) was very close to the value identified as peak for pastoral value in the montane and subalpine belts (3.1), while the mean value within BPA (2.9) was in between the values identified as peaks for plant diversity (2.5–2.6) and pastoral value (3.1), respectively. Therefore, in the Southern Swiss Alps, the BPA had a soil nutrient content ideal to provide both good forage quality and plant diversity levels, not so far from to the maximum values that can be found for alpine grasslands. A further prolongation of the implementation the BPA scheme in the future years within these grasslands would probably allow to reduce soil nutrient content levels and increasing vascular plant diversity as a consequence.
Vegetation types, functional groups, and plant species
The BPA were characterised by a larger proportion of species belonging to nutrient-poor (i.e. pasture species) and pre-forest (i.e. shrubland–woodland and edge species) vegetation communities than CMG, highlighting that these grasslands were characterised by a less productive vegetation, with several species typical of ecotones and forest-transition zones. At the opposite, CMG hosted a greater proportion of fast-growing and productive species, including nutrient-rich meadow species and ruderal species, an expected result for grasslands characterised by intensive cut/grazing and fertilisation regimes (Hejcman et al. 2012). Considering vegetation types, F. rubra—B. rupestre type was the only one significantly associated to BPA. It is worth mentioning that B. rupestre is a summer drought stress tolerant tall-grass encroaching mountain grasslands when agro-pastoral management is strongly reduced and/or totally abandoned, causing a sharp decline of biodiversity and forage quality when it becomes dominant within the vegetation community (Cavallero et al. 2007; Bricca et al. 2020; Tardella et al. 2020). However, a regular and extensive disturbance management, such as the cutting/grazing regimes implemented within BPA, is recommended to reduce its dominance and maintain good level of vascular plant diversity (Tardella et al. 2020). Moreover, the regular and extensive BPA disturbance management proved to be efficient in avoiding the risk of natural succession towards communities dominated by tree and shrub species, as indicated by the low number and cover of typical shrubland–woodland and heathland species (Table 4).
In their recent study conducted on 942 grasslands across Switzerland, Riedel et al. (2019) concluded that less than a half of the extensive and low-intensive BPA meadows belonged to the vegetation types that have been indicated as conservation targets by the Federal Office for Agriculture (UFAG 2020b), such as Bromus erectus-, Arrhenaterum elatius-, and Trisetum flavescens-dominated communities. Our results confirmed this conclusion, since only very few BPA belonged to these vegetation communities (e.g. those belonging to Arrhenaterum elatius type). The main reason lies in the fact that farmers are free to choose which grassland has to be indicated as BPA to have access to direct payments, regardless of site conditions, botanical composition, and thus the potential that a particular grassland has to achieve a specific vegetation community target. This consideration is particularly meaningful for nutrient-rich grasslands characterised by a low species richness, which could be difficulty shifted towards these vegetation targets, above all if a target seed bank is not present (Riedel et al. 2019). At the opposite, when considering the species significantly associated to BPA, 38% of them were species belonging to the national list of species of priority interest for biodiversity promotion in agriculture, whereas no species associated to CMG belonged to that list (Swiss Ordinance 910.13; UFAG 2020b). Moreover, the larger number of functional groups of species related to BPA confirmed the wider environmental heterogeneity of these grasslands, which was noticeable not only at species level, but also in terms of phytosociological syntaxa. Conversely, the species associated to CMG were all typical of more intensive management situations and included a large set of high-palatable grass-legume forage species (e.g. Lolium perenne, L. multiflorum, D. glomerata, Phleum pratense, Poa pratensis, Trifolium repens) or species favoured by very high fertilisation rates (e.g. Rumex acetosa, R. obtusifolius, P. trivialis) and/or disturbance regimes (e.g. Stellaria media, Taraxacum officinale).
Policy implications and future scenarios
According to the Swiss policy, farmers must manage at least 7% of their agricultural area as BPA to get access to agricultural direct payments. This system resulted to be effective to produce widespread benefits for the conservation of plant diversity at regional scale, as highlighted by our findings. Moreover, the beneficial results produced do not limit exclusively to flora, but they extend to a large number of animal taxa as well, such as butterflies, grasshoppers and ground-nesting birds, which are favoured by late mowing regimes (Herzog et al. 2005; Knop et al. 2006; Zingg 2018). For this reason, the same principle of identifying a mandatory minimum percentage threshold of agricultural surface that must be managed according to extensive rules by farmers, in order to receive agricultural direct payments, should be encouraged and implemented in other countries as well. However, one of the major limitations of BPA implementation is the lack of precise obligations to farmers for the identification of surfaces eligible for the direct payments. Consequently, grasslands characterised by low plant diversity and/or a botanical composition unfavourable to achieve specific vegetation target goals have been often indicated as BPA. To improve this situation, the agricultural advice services can be an important support, providing both ecological and economic benefits to farmers (Chevillat et al. 2017).
Another possible limitation, in the context of future climate change scenarios, can be a too inflexible obligation on the mowing dates that are indicated for the different altitudinal belts. Indeed, the higher temperatures are determining a shift in grassland plant phenology all over the Alps, resulting in a marked advancement of the mean phenological stages for many species, accounting to 2–3 days every 10 years (Vuffray et al. 2016), and this process is foreseen to accelerate in the next decades, together with an increase of summer droughts, above all in the Southern Swiss Alps (Croci-Maspoli et al. 2018). Consequently, keeping inflexible mowing dates in the next future would probably determine a decrease in the quality of forage deriving from BPA as a result of too advanced phenological stages and a dominance of summer drought stress tolerant grasses of low forage quality, such as B. rupestre and typically associated late-flowering tall herbs (Tardella et al. 2020). For this reason, it could also have potential impacts on species community composition, resulting in a lower acceptance of this biodiversity-based scheme by farmers. As a consequence, it appears advisable to provide a more flexible range of mowing dates for the different altitudinal belts, whose indication could be based on the mean phenological stage of a set of key plant or animal species that could be communicated yearly to farmers, in order to take into account climatic interannual variations.
Conclusion
The present study highlighted the beneficial effects produced by Swiss agricultural direct payments in preserving vascular plant diversity at a landscape scale, since they encouraged extensive grassland management regime in the more marginal mountain sites, thus avoiding both agro-pastoral abandonment and intensification. These findings confirmed for the first time within the Southern Swiss Alps the results of previous research conducted over other alpine regions. According to the results of the present study, grasslands managed as ‘Biodiversity Promotion Areas’ harbour a higher plant diversity, as well as many species of priority interest for biodiversity promotion in agriculture, belonging to a wider range of different vegetation units than conventional managed grasslands. Future adaptations of this effective policy measure appear advisable to better adapt the legislation obligations to the environmental variations determined by forthcoming climatic scenarios.
Data availability
The datasets generated and analysed during the current study are available in the Zenodo repository, https://zenodo.org/record/3755786#.XpnUEcgzbIU.
Code availability
Not applicable.
References
Aeschimann D, Lauber K, Moser DM, Theurillat JP (2004) Flora alpina: atlante delle 4500 piante vascolari delle Alpi. Zanichelli, Bologna
Bätzing W, Bartaletti F (2005) Le Alpi: una regione unica al centro dell’Europa. Bollati Boringhieri, Turin
Bricca A, Tardella FM, Tolu F et al (2020) Disentangling the effects of disturbance from those of dominant tall grass features in driving the functional variation of restored grassland in a Sub-Mediterranean context. Diversity 12:11
Cavallero A, Aceto P, Gorlier A et al (2007) I tipi pastorali delle Alpi piemontesi: vegetazione e gestione dei pascoli delle Alpi occidentali. Alberto Perdisa Editore, Bologna
Chang C-R, Lee P-F, Bai M-L, Lin T-T (2004) Predicting the geographical distribution of plant communities in complex terrain—a case study in Fushian Experimental Forest, northeastern Taiwan. Ecography 27:577–588
Chao A (1987) Estimating the population size for capture-recapture data with unequal catchability. Biometrics 43:783–791
Chevillat V, Stöckli S, Birrer S et al (2017) Surfaces de promotion de la biodiversité: Amélioration quantitative et qualitative par le conseil. Recherche agronomique suisse 8:232–239
Croci-Maspoli M, Schär C, Fischer A et al (2018) CH2018-Climate Scenarios for Switzerland-Technical Report. National Centre for Climate Services, Zurich (Switzerland)
Daget P, Poissonet J (1971) Une méthode d’analyse phytosociologique des prairies. Annales Agronomiques 22:5–41
De Cáceres M, Legendre P (2009) Associations between species and groups of sites: indices and statistical inference. Ecology 90:3566–3574
De Cáceres M, Legendre P, Moretti M (2010) Improving indicator species analysis by combining groups of sites. Oikos 119:1674–1684. https://doi.org/10.1111/j.1600-0706.2010.18334.x
Dengler J, Janišová M, Török P, Wellstein C (2014) Biodiversity of Palaearctic grasslands: a synthesis. Agric Ecosyst Environ 182:1–14. https://doi.org/10.1016/j.agee.2013.12.015
Dietschi S, Holderegger R, Schmidt SG, Linder P (2007) Agri-environment incentive payments and plant species richness under different management intensities in mountain meadows of Switzerland. Acta Oecologica 31:216–222
Dufrêne M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol Monogr 67:345–366
Estel S, Kuemmerle T, Alcántara C et al (2015) Mapping farmland abandonment and recultivation across Europe using MODIS NDVI time series. Remote Sens Environ 163:312–325. https://doi.org/10.1016/j.rse.2015.03.028
EUROSTAT (2019) Agri-environmental indicator—cropping patterns—statistics explained. https://ec.europa.eu/eurostat/statistics-explained/index.php/Agri-environmental_indicator_-_cropping_patterns. Accessed 20 Apr 2020
Federal Office of Topography (2018) Digital height model DHM25. https://shop.swisstopo.admin.ch/en/products/height_models/dhm25. Accessed 10 Apr 2020
Fournier B, Mouly A, Moretti M, Gillet F (2017) Contrasting processes drive alpha and beta taxonomic, functional and phylogenetic diversity of orthopteran communities in grasslands. Agric Ecosyst Environ 242:43–52
Gonseth Y, Wohlgemuth T, Sansonnens B, Buttler A (2001) Die biogeographischen regionen der Schweiz. Erläuterungen und Einteilungsstandard Umwelt Materialien 137, Swiss Agency for Environment, Forests and Landscape, Bern
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4:1–9
Harrison PA, Vandewalle M, Sykes MT et al (2010) Identifying and prioritising services in European terrestrial and freshwater ecosystems. Biodivers Conserv 19:2791–2821
Hejcman M, Strnad L, Hejcmanová P, Pavlů V (2012) Response of plant species composition, biomass production and biomass chemical properties to high N, P and K application rates in Dactylis glomerata-and Festuca arundinacea-dominated grassland. Grass Forage Sci 67:488–506
Herzog F, Dreier S, Hofer G et al (2005) Effect of ecological compensation areas on floristic and breeding bird diversity in Swiss agricultural landscapes. Agric Ecosyst Environ 108:189–204
Hooper DU, Chapin FS, Ewel JJ et al (2005) Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol Monogr 75:3–35
IBM Corp. (2017) IBM SPSS Statistics for Windows, Version 25.0. Version 25.0. IBM Corporation, Armonk, NY
Kadereit JW (2017) The role of in situ species diversification for the evolution of high vascular plant species diversity in the European Alps—a review and interpretation of phylogenetic studies of the endemic flora of the Alps. Perspect Plant Ecol Evol Syst 26:28–38
Kampmann D, Herzog F, Jeanneret P et al (2008) Mountain grassland biodiversity: impact of site conditions versus management type. J Nat Conserv 16:12–25
Kleijn D, van Zuijlen GJ (2004) The conservation effects of meadow bird agreements on farmland in Zeeland, The Netherlands, in the period 1989–1995. Biol Conserv 117:443–451
Knop E, Kleijn D, Herzog F, Schmid B (2006) Effectiveness of the Swiss agri-environment scheme in promoting biodiversity. J Appl Ecol 43:120–127. https://doi.org/10.1111/j.1365-2664.2005.01113.x
Koordinationsstelle Biodiversitäts-Monitoring Schweiz (2009) Koordinationsstelle Biodiversitäts-Monitoring Schweiz, 2009. Zustand der Biodiversität in der Schweiz—Ergebnisse des Biodiversitätsmonitorings Schweiz (BDM) im Überblick. Stand: Mai 2009. Umwelt-Zustand 09/11. Federal Office for the Environment
Körner C (2003) Alpine plant life: functional plant ecology of high mountain ecosystems. Springer Berlin Heidelberg, Berlin, Heidelberg
Landolt E, Bäumler B, Erhardt A et al (2010) Flora indicativa: ökologische Zeigerwerte und biologische Kennzeichen zur Flora der Schweiz und der Alpen = ecological indicator values and biological attributes of the flora of Switzerland and the Alps. Editions des Conservatoire et Jardin botaniques de la Ville de Genève & HauptVerlag, Bern, Stuttgart, Vienna
Magurran AE (1988) Ecological diversity and its measurement. Princeton University Press, Princeton
Magurran AE (2004) Measuring biological diversity. Blackwell Publishing, Oxford
Manning P, Gossner MM, Bossdorf O et al (2015) Grassland management intensification weakens the associations among the diversities of multiple plant and animal taxa. Ecology 96:1492–1501. https://doi.org/10.1890/14-1307.1
Mottet A, Ladet S, Coqué N, Gibon A (2006) Agricultural land-use change and its drivers in mountain landscapes: a case study in the Pyrenees. Agric Ecosyst Environ 114:296–310
Nagy S (2009) Grasslands as a bird habitat. In: Veen P, Jeferson R, de Smidt J, and van der Straaten J (eds) Grasslands in Europe. KNNV Publishing, pp 35–41
Orlandi S, Probo M, Sitzia T et al (2016) Environmental and land use determinants of grassland patch diversity in the western and eastern Alps under agro-pastoral abandonment. Biodivers Conserv 25:275–293. https://doi.org/10.1007/s10531-016-1046-5
Ozenda P (1985) La végétation de la chaîne alpine dans l’espace montagnard européen. Masson, Paris
Perotti E, Probo M, Pittarello M et al (2018) A five-year rotational grazing changes the botanical composition of sub-alpine and alpine grasslands. Appl Veg Sci 21:647–657
Peter M, Gigon A, Edwards PJ, Lüscher A (2009) Changes over three decades in the floristic composition of nutrient-poor grasslands in the Swiss Alps. Biodivers Conserv 18:547–567. https://doi.org/10.1007/s10531-008-9521-2
Pittarello M, Lonati M, Gorlier A et al (2017) Species-rich Nardus stricta grasslands host a higher vascular plant diversity on calcareous than on siliceous bedrock. Plant Ecol Divers 10:343–351
Pittarello M, Lonati M, Gorlier A et al (2018) Plant diversity and pastoral value in alpine pastures are maximized at different nutrient indicator values. Ecol Indic 85:518–524
QGIS Development team (2019) QGIS Geographic Information System. Open Source Geospatial Foundation Project
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Riedel S, Lüscher G, Meier E et al (2019) Qualité écologique des prairies qui bénéficient des contributions à la biodiversité. Recherche Agronomique Suisse 10:80–87
Roleček J, Čornej II, Tokarjuk AI (2014) Understanding the extreme species richness of semi-dry grasslands in east-central Europe: a comparative approach. Preslia 86:5–27
Swiss National Agricultural Report (2019) Superficie agricola utile. https://www.agrarbericht.ch/it/azienda/strutture/superficie-agricola-utile. Accessed 8 Apr 2020
Tardella FM, Bricca A, Goia IG, Catorci A (2020) How mowing restores montane Mediterranean grasslands following cessation of traditional livestock grazing. Agric Ecosyst Environ 295:106880
Theurillat J-P, Aeschimann D, Küpfer P, Spichiger R (1995) The higher vegetation units of the Alps. In: Géhu J-M (ed) Colloques Phytosociologiques. Verlag nicht ermittelbar, Bailleul, pp 189–239
Tocco C, Negro M, Rolando A, Palestrini C (2013) Does natural reforestation represent a potential threat to dung beetle diversity in the Alps? J Insect Conserv 17:207–217
UFAG-Federal office for agriculture (2020a) Politica agricola. https://www.blw.admin.ch/blw/it/home/politik/agrarpolitik.html. Accessed 20 Apr 2020
UFAG-Federal office for agriculture (2020b) Contributo per la qualità. https://www.blw.admin.ch/blw/it/home/instrumente/direktzahlungen/biodiversitaetsbeitraege/qualitaetsbeitrag.html. Accessed 9 Apr 2020
UST-Federal statistical office (2018a) Farm land uses and utilised agricultural area (UAA) according to classification level 3 by Canton. In: PX-Web. https://www.pxweb.bfs.admin.ch/pxweb/en/px-x-0702000000_106/px-x-0702000000_106/px-x-0702000000_106.px/. Accessed 20 Apr 2020
UST-Federal Statistical Office (2018b) Agriculture and forestry. https://www.bfs.admin.ch/bfs/en/home/statistiken/land-forstwirtschaft.html. Accessed 10 Apr 2020
Vuffray Z, Deléglise C, Amaudruz M et al (2016) Phänologische Entwicklung von Mähwiesen—21 Beobachtungsjahre. Agrarforschung Schweiz 7:322–329
WallisDeVries MF, Van Swaay CA (2009) Grasslands as habitats for butterflies in Europe. In: Veen P, Jeferson R, de Smidt J, and van der Straaten J (eds) Grasslands in Europe. KNNV publishing, pp 26–34
Welten M, Sutter R (1982) Verbreitungsatlas der Farn-und Blütenpflanzen der Schweiz/Atlas de Distribution des Pteridophytes et des Phanerogames de la Suisse/Atlante della Distribuzione delle Pteridofite e Fanerogame della Svizzera. Birkhäuser, Basel
Wilson JB, Peet RK, Dengler J, Pärtel M (2012) Plant species richness: the world records. J Veg Sci 23:796–802. https://doi.org/10.1111/j.1654-1103.2012.01400.x
Zabel A (2019) Biodiversity-based payments on Swiss alpine pastures. Land Use Policy 81:153–159
Zingg S (2018) Integrating food production and biodiversity conservation in temperate agricultural landscapes. Inauguraldissertation der Philosophisch-naturwissenschaftlichen Fakultät der Universität Bern. PhD, Universität Bern
Zuur A, Ieno EN, Walker N et al (2009) Mixed effects models and extensions in ecology with R. Springer Science & Business Media, New York
Acknowledgements
Authors would like to thank the Swiss Grassland Society that funded the project PRATIVA (Grant Number 15-314) in the context of which the vegetation surveys were carried out, the Cantonal Office for Agricultural Advisory of Ticino, and all the farmers visited during the project. Authors also thank prof. Andrea Cavallero for inspiring this work and providing us the necessary knowledge.
Funding
Open access funding provided by Agroscope This research was founded by the Swiss Grassland Society under the project PRATIVA (Grant Number 15-314).
Author information
Authors and Affiliations
Contributions
All authors contributed equally to this work.
Corresponding author
Ethics declarations
Conflict of interest
Authors disclose all relationships or interests that could have direct or potential influence or impart bias on the work.
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Communicated by Daniel Sanchez Mata.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article belongs to the Topical Collection: Biodiversity appreciation and engagement.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ravetto Enri, S., Nucera, E., Lonati, M. et al. The Biodiversity Promotion Areas: effectiveness of agricultural direct payments on plant diversity conservation in the semi-natural grasslands of the Southern Swiss Alps. Biodivers Conserv 29, 4155–4172 (2020). https://doi.org/10.1007/s10531-020-02069-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-020-02069-4