
Abstract
Riparian ecosystems have unique biodiversity, are highly sensitive to disturbance and anthropogenic influence. As world water resources become scarcer, scientists predict greater competition among species for water resources. Indeed, increased encroachment of upland plants into the riparian zone is already occurring, irreversibly changing riparian plant communities. Since semi-arid regions such as Mediterranean-type ecosystems are likely to follow this same trajectory, assessing the contributions of riparian versus upland (sclerophyllous) plants to community composition is important. A survey of seventy 2 km-long riparian transects on the Sado and Guadiana watersheds in southern Portugal assessed (1) the woody riparian plant community composition, (2) how much richness is due to strictly riparian plants versus sclerophyllous upland plants, and (3) which combinations of biotic and abiotic factors allow higher species richness in the strictly riparian, sclerophyllous, and overall plant communities. The survey detected 53 different woody plant species (28 endemic) across all communities. Riparian community richness was on average 16 species, seven of which were strictly riparian and the remainder being sclerophyllous, exotic species or fruit trees. Sclerophyllous plant species occurred consistently across sampling units (90% of transects). On average, 46% of the total woody plant community richness was due to strictly riparian plants and 28% was due to sclerophyllous plants. Community richness was positively affected by the area of shrubs in the riparian zone and by the absence of human activities and goats. Surrounding landscape pattern only affected the strictly riparian plant richness. These results suggest that natural and human-mediated disturbances in riparian ecosystems create gaps and clearings for which riparian and sclerophyllous plants compete. Establishment success seems to be related to the propagule pressure of the neighbouring landscape, its diversity and density, as well as the presence of herbivores. Preserving strictly riparian plants, removing exotic species, preventing grazing, and promoting riparian values (recreation, aesthetics and the provision of ecosystem services) will aid the future conservation of the unique biodiversity of riparian ecosystems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Riparian ecosystems are highly diversified, dynamic and complex biophysical terrestrial ecosystems (Miller 2002; Naiman et al. 2005). These systems are transitional zones between aquatic and upland terrestrial environments with a linear spatial configuration. Riparian ecosystems contain a high and unique number of plant species (Sabo et al. 2005), adapted to disturbance (e.g., floods, drought) (Lyon and Gross 2005; Malanson 1993), in a restricted area of land (Lyon and Gross 2005; Malanson 1993). Riparian ecosystems also provide aquatic, water-land interface and terrestrial habitats for animal species, as well as drinking water for upland animals (Brookshire et al. 2002; Hilty and Merenlender 2004; Iverson et al. 2001; Machtans et al. 1996; Matos et al. 2008; Spackman and Hughes 1994; Virgós 2001; Williams et al. 2003). Despite their high biological value, riparian ecosystems have seldom been included in systematic conservation planning (Nel et al. 2009), and are becoming increasingly threatened by human activities (Salinas et al. 2000) and upland plant encroachment (Huxman et al. 2005), especially in the semi-arid Mediterranean region (Nel et al. 2009).
Riparian plant communities in Mediterranean climates have been impoverished and threatened by human activities (Aguiar et al. 2006; Schnitzler et al. 2007) such as agriculture (Aguiar and Ferreira 2005; Salinas et al. 2000; Tabacchi et al. 2002), land development for industry or tourism, and transportation infrastructures (Jongman and Pungetti 2003; Scarascia-Mugnozza et al. 2000). These changes led to the loss of unique riparian species (Sabo et al. 2005; Salinas et al. 2000) and likely resulted in woody plant encroachment in the riparian ecosystem (Huxman et al. 2005). Woody plant encroachment causes major shifts in hydrological dynamics by decreasing surface and subsurface flow, which decreases scouring flows, leading to an increase in woody plant survival. This results in higher forest cover along the channel, which intensifies water loss through increased transpiration, and decreases water availability to other plant and wildlife species, and other riparian functions (for a review see Huxman et al. 2005).
The impacts of woody plant encroachment on water availability are exacerbated by climate change impacts on riparian areas. Rivers have already been influenced by changing precipitation regimes resulting from climate change (Schröter et al. 2005), especially in areas like the Iberian Peninsula which have become more arid. It is likely that such areas will be more susceptible to incursion by sclerophyllous plants (species which have developed leathery leaves to minimize water loss, as a response to nutrient poor soil, or herbivory) that may eventually completely replace strictly riparian plants (species with a life cycle that requires an inundated period for seed establishment and germination). Such a transition is likely to occur when space becomes available to sclerophyllous species, spatially segregated from the strictly riparian plant patches. To test if this is this pattern holds along two main watersheds in southern Portugal, I investigated the ratio of strictly riparian and sclerophyllous species. More specifically, the study asked (1) how much of the riparian richness is due to strictly riparian species and how much is due to sclerophyllous woody plant species; (2) are the strictly riparian and sclerophyllous woody plant species spatially segregated; and (3) which combination of biotic and abiotic factors allows for a richer, more intact (i.e., more strictly riparian plants) woody riparian community. Based on the results of this survey, this paper assesses watershed level impacts in riparian ecosystems and proposes measures that enhance conservation of these unique riparian ecosystems in face of current threats and climate change.
Materials and methods
Study area
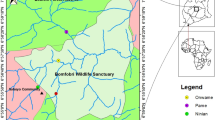
The study area was a 6,400 km2 region located within the Alentejo province of southern Portugal (Fig. 1). Topography in this area ranges from coastal areas to low altitude mountains (400 m). Climate varies from cold to mild during the winter to hot and dry during the summer (with a temperature range of −8 to 45.2°C, Portuguese Meteorology Institute; Carmel and Flather 2004). Mean precipitation is 500 mm/year, with a dry period from June to September (Rivas-Martinez 1987). Dominant plant communities are characterized by cork oak (Quercus suber) and holm oak (Q. rotundifolia) woodlands with terrestrial shrubs such as rock-rose (Cistus spp.) dominating the sub-canopy (Chícharo et al. 2001). The majority of watercourses are intermittent, drying during the summer. Along watercourses, the dominant riparian species are the white poplar (Populus alba), raywood ash (Fraxinus angustifolia), grey willow (Salix atrocinerea), African tamarisk (Tamarix africana), oleander (Nerium oleander) and alder (Alnus glutinosa) (Aguiar et al. 2006; Chícharo et al. 2001). Human activities include traditional agriculture (olive groves, vineyards, cereal production and cork extraction) and livestock production (cattle, sheep, goats and pigs), which require water extraction from neighbouring watercourses, and result in reduction of riparian vegetation by clear-cutting or grazing.

Location of the study area in Alentejo province, southern Portugal. Highlighted areas indicate the riparian ecosystems sampled (creeks n = 24, streams n = 24 and rivers n = 22)
Sampling design
I used a stratified random sampling method to select 72 water stretches. The stretches were equally distributed among creeks (1–5 m wide and lower order waterways), streams (5–10 m wide, and intermediate order waterways) and rivers (>10 m wide and high order waterways), and among surrounding dominant land-cover (cork oak woodland, holm oak woodland, and agriculture) giving eight replicates of each waterway-type/landscape-context combination (Fig. 1). Creeks, streams and rivers were defined by their progressively higher order, and this classification was confirmed by testing if the classified stretches had significantly different river bed width. Since there was a clear significant difference in river bed width between creeks, streams and rivers, the distinction was considered reliable. I derived five land-cover classes from the 1990 CORINE land-cover data (derived from classification of Landsat TM 30m resolution multispectral imagery) within a 1.5 km wide buffer of the waterway. The classes are: extensive agriculture (cereal plantations) (58%), cork oak woodland (23%), holm oak woodland (6%), intensive agriculture (e.g., tomato, corn; 1%), and other (including Eucalyptus spp. and Pinus spp. plantations, urban areas, etc.; 12%).
I used a digital data layer of watercourses in the study area overlain on the land-cover data to identify all possible 2 km stretches dominated by a single land-cover type within the waterway buffers. Seventy-two sampling sites were randomly selected from this layer and screened for site accessibility. Two river transects surrounded by holm oak woodlands were inaccessible, resulting in a final sample of 70 transects.
Field data collection
I visited all sites once for plant identification between December 2003 to February 2004, and revisited each transect between June to September 2004 to assess any change in environmental context variables (see below). The two seasons represent the variability of surface water in the watercourses, a key factor affecting plant establishment and growth.
Each 2 km transect was subdivided into 200 m segments, in which plant species presence was recorded. This distance was selected as subsamples because it matches the minimum resolvable unit in the land cover map (approximately 200 m2), and is comparable to similar surveys along riparian systems in the Iberian Peninsula (Aguiar and Ferreira 2005). Each waterway was surveyed using a transect parallel to the right waterway margin, which I walked while recording the presence or absence of every woody plant species within 5 m of the bank. All plant species were identified in the field, and samples of unknown species were collected and identified in the laboratory. The identification was resolved to the finest taxonomic status possible, with all specimens categorized at genus or species levels, especially in the case of the willow, moor and heath species, which lacked diagnostic features during the sampling months. Herbaceous species were excluded from the analysis because of the lack of consistently identifiable features (due to either phenology or herbivory).
Environmental variables
To assess the role of biotic and abiotic factors in determining the richness of the woody riparian plant communities, each of the sampling sites was characterized according to characteristics of the riparian zone itself, water availability, characteristics of the surrounding landscape, and the presence of human disturbance (Table 1). Characteristics of the riparian zone included the structure of the riparian area (presence of tree and shrub layers, along with their length, width and area, calculated by multiplying length by width; Table 1) and the continuity of the riparian vegetation (vegetation was defined as sparse when >100 m were traversed without finding woody species). I measured the length and average width of the tree and shrub layers within each transect. I measured water depth and width every 200 m along the transect (a total of 11 measurements per transect), and used the average value to characterise the water availability of each transect. Water characteristics were measured twice at the same locations, once in the wet season (winter) and once in the dry season (summer).
It is likely that surrounding land cover contributes a significant source of seeds and propagules to riparian areas. Therefore, I characterized the surrounding landscape using a suite of landscape metrics calculated from the available digital CORINE landcover data following their formulation in McGarigal and Marks (1995) (Table 1). All the metrics were calculated for a buffer of 1.5 km from each side of the riparian zone because this was the average distance from the waterway to the top of the nearest hill. As a proxy for the effect of propagule connectivity (Li and Wu 2004), I assessed the potential impact of type of surrounding landscape (area of cork oak, holm oak, dry agriculture, irrigated agriculture and others), and for each of the land cover types its extent (patch size), configuration (number of patches), its degree of contact with the riparian area (edge density) and its shape complexity (area weighted mean shape index and area weighted mean fractal dimension). Further, to assess the effect of the presence of multiple surrounding landscapes on the seed sources to surveyed patches in the riparian areas, the landscape diversity index and landscape equitability were calculated using Shannon-Wiener (H’) and Simpson (D) diversity indexes, which account for both the abundance and evenness of landscape (Krebs 1998). The Shannon diversity index emphasizes rare landscapes whereas the Simpson diversity index more heavily weights common landscapes. H’ varies between 0 to log(k), where k is the number of classes, and D varies from 0 to +∞. I also calculated the evenness of the landscapes using the Shannon’s equitability index (J’). Equitability assumes values between 0 and 1, with 1 being complete evenness in landscape composition and corresponds to samples receiving the maximum value of the Shannon-Wiener index. Landscape metrics were calculated using the “Patch Analyst v. 3.1” (Rempel and Carr 2003) extension for ArcView 3.2 (ESRI 1996). Finally, I made qualitative assessments of the degree of human presence through registering presence and absence of human activities (houses, livestock, hunting, farming, etc.), development (houses, fences and roads), and livestock along each transect (Table 1).
Data analysis
Woody plant richness was defined as the total number of identified species per sampling unit (per transect). Richness values for strictly riparian species (species with a life cycle that requires an inundated period for seed establishment and germination) and sclerophyllous species (species which have developed leathery leaves to minimize water loss, and as a response to poor nutrient soils and herbivory) were also calculated.
In order to assess if the samples were sufficient to describe study-area-wide riparian vegetation richness I used a species transect curve. A sample was considered sufficient when the curve of the cumulative number of identified species plotted against the number of samples reaches an asymptote, i.e., the more samples collected the fewer new species are expected to be found. The number of samples at which the asymptote is reached corresponds to the sufficient sample size required (Krebs 1998). Species-transect curves were calculated in PC-ORD (McCune and Grace 2002), and an asymptote was reached with 22 sampling transects, even when separating between creeks (n = 24), streams (n = 24) and rivers (n = 22). This indicates that the sample size was sufficient to characterize the variability in the study area.
The effects of spatial autocorrelation on transect location were tested using Moran’s I index (Moran 1950). This index measures the similarity in the spatial patterns of the variable (Fortin et al. 1989), in our case woody species richness, and varies from −1 (perfect negative spatial autocorrelation) to 1 (perfect positive spatial autocorrelation), with values close to 0 representing no spatial autocorrelation. To estimate the distance threshold at which spatial autocorrelation could be considered negligible, the neighborhood distance was progressively increased from a radius of 1000–5000 m in 1000 m increments and I measured Moran’s I index for each radius distances. Spatial autocorrelation was calculated using ROOKCASE Microsoft Excel Add-in (Sawada 1999). Since no significant spatial autocorrelation was found at distances above 1.5 km, it was concluded that spatial autocorrelation was not affecting the data and therefore it could be used for further analysis.
One-way ANOVA was used to determine if the riparian plant community richness was a function of the watercourse type, after testing for normality in the distribution of the variables and transforming accordingly (log transforming area of landcover) (Zar 1999). To test how much of the total richness is a function of the riparian and the sclerophyllous plants, a regression was fitted between the total species richness and the richness of riparian and sclerophyllous plants. The slope of the regression line indicates additive richness (slope = 1), complete replacement (slope = 0) or partial replacement (0 < slope < 1). To assess the proportion of the richness that each of these groups contributes, the percentage of the total richness due to riparian and sclerophyllous plant species was estimated and a linear regression fitted between the total richness and the percent of riparian and sclerophyllous in the community. A linear regression was also fitted between the richness of riparian and sclerophyllous plants to identify a relationship between the two.
The patch structure of the riparian zones was analysed by comparing the segments of each transect in terms of their riparian and sclerophyllous composition. I tested whether the two vegetation types were present in the same spatial location (i.e., the same 200 m sample) or spatially segregated in the same riparian zone. Linear regression was used to test if within each segment higher richness of strictly riparian plants was correlated with higher richness of sclerophyllous vegetation. If the slope of the regression was negative it would indicate spatial segregation. For these tests a significance level of 0.05 was used, and Bonferroni corrections were applied to correct significance values for multiple comparisons (Zar 1999).
The correlation between each of the environmental context variables (Table 1) was tested using Pearson correlation coefficients (Zar 1999). Since there was not significant collinearity between any of the predictor variables, they were maintained for further analysis. A generalized linear model (GLM) was used to test the effect of each of the environmental context variables in the total riparian plant richness, richness of strictly riparian plants and richness of sclerophyllous plant species. Model significance was assessed using F-test values, and for statistically significant models (α = 0.1), model fit (explanatory power) was assessed using R-square values. All statistical analyses were performed using JMP 5.0 (SAS Institute) for Windows.
Results
Riparian plant richness
Riparian plant communities were composed of 53 different woody plant species, which included strictly riparian and sclerophyllous plant species (Appendix Table 3). Raywood ash (60.6%), cork oak (40.7%), willows (40.1%), black poplar (33.1%), olive tree (31%), and holm oak (30.2%) were the most common tree species, and blackberry (79.5%) and rockrose (36.1%) were the most common shrubs. Strictly riparian species included white willow and other willows, African tamarisk, black poplar, and raywood ash. Sclerophyllous species included cork and holm oak, lentisc and rock-roses. Sclerophyllous plant species were consistently found across all sampling units, except for 10% of transects (7 out of 70) where no sclerophyllous species were detected. Exotic species such as acacia and eucalyptus were also commonly found, and so were fruit trees, including pears, quinces, and others (see Appendix Table 3).
Species richness had a mean of 15.6 ± 7.3 species, with a maximum of 33 different species in one transect and a minimum of two species. Strictly riparian species richness was significantly higher than sclerophyllous plants (F = 6.46, d.f. = 138, P = 0.01). Strictly riparian had a mean richness of 6.6 ± 2.5 species per transect, whereas sclerophyllous species had a mean richness of 5.2 ± 3.9 species.
When considering all species, species richness did not vary as a function of watercourse type, as they were not significantly different among creeks, streams and rivers (P > 0.05 for all tests). Sclerophyllous plants richness alone was also not significantly different along watercourses (F = 0.51, d.f. = 69, P = 0.6). Riparian species richness, when considered alone, was significantly higher along rivers (F = 5.02, d.f. = 69, P = 0.009) than either creeks or streams.
On average, 46% of the woody plant species were strictly riparian plants, and 28% were sclerophyllous plants. However, there is a stronger relationship between total riparian richness and the sclerophyllous plant richness (R 2 = 0.84) than that between total riparian richness and strictly riparian plant species (R 2 = 0.51) (Fig. 2a), indicating that most of the total riparian richness is due to sclerophyllous plant species. The remainder of the variability was accounted for by exotic and fruit trees. As the total richness of the community increased, the percentage of strictly riparian plants significantly decreased (P < 0.0001) and the percentage of sclerophyllous plants significantly increased (P < 0.0001; Fig. 2b). Lowest total richness was associated with a community dominated by strictly riparian and high total richness was due to the combined presence of strictly riparian and encroachment by sclerophyllous species. There was a weak (R 2 = 0.19) but significant positive correlation between strictly riparian and sclerophyllous plant species richness within riparian areas (Fig. 3). Regression between strictly riparian and sclerophyllous plants in each 200 m segment was not significant (P > 0.05), indicating no spatial segregation.

(a) Regression of strictly riparian (closed circles, left axis, dotted line) and sclerophyllous (open circles, right axis, full line) plant species with total plant species richness. The stronger explanatory power of the sclerophyllous regression indicates an additive effect of sclerophyllous species to total richness. (b) Regression of % strictly riparian (closed circles, left axis, dotted line) and sclerophyllous (open circles, right axis, full line) plant species with total plant species richness. The proportion of each group changes as total richness increases

Relationship between strictly riparian and sclerophyllous species richness. There is a positive relationship between the two plant groups, but highly variable
Environmental variables associated with riparian plant richness
Higher total woody plant richness, as well as strictly riparian and sclerophyllous richness were mainly a function of the areas of shrubs in the riparian ecosystem (except for sclerophyllous plants richness), as well as the absence of human activities and goats (Table 2). Strictly riparian plant richness was also significantly increased by a surrounding landscape with high number of large holm oak patches, and with low diversity of land covers (Table 2). Sclerophyllous plants richness increased in reduced areas of agriculture and reduced human activities and goats (Table 2). The final statistical model explained about 70% of the variability in total woody species richness, and similar values were attained for both strictly riparian (69%) and sclerophyllous species (71%). All GLMs were significant (Table 2).
Discussion
Riparian plant richness
Previous studies of richness of comparable riparian systems in the Iberian Peninsula have shown that in the last 5 years, these communities have on average 16 woody riparian plant species in 100 m (Aguiar and Ferreira 2005; Aguiar et al. 2006). The results presented in this study show lower values (average richness of eight species per 100 m, Table 2), with less than half of the sites (31 out of 70) having more than 15 species. Several factors contribute to richness in riparian plant communities, such as productivity (Pollock et al. 1998), flow-facilitated dispersal of seeds and propagules (Deferrari and Naiman 1994), soil variability (Pollock et al. 1998), geographical and topographical variability (Naiman and Décamps 1997), disturbance (Pollock et al. 1998), and diversity of interfaces between aquatic and terrestrial habitats (Naiman and Décamps 1997). Since the areas that were surveyed in this study are similar to those studied previously and, in some cases, at the same locations of studies from other authors, it is not likely that the discrepancy in the results is due to differences in productivity, flow-facilitated dispersal of seeds and propagules, soil variability, and geographical and topographical variability. However, the degree of disturbance of the sites in the current study may be higher than that of previous studies. A recent study focusing on ranking the health of riparian systems in southern Portugal found that most riparian stretches are in poor condition (Matos et al. 2008). Furthermore, current riparian clear cut practices, such as those present in the study area, have been identified as a cause of decreasing riparian species richness in the Iberian Peninsula (Salinas et al. 2000).
Higher total plant richness was recorded when more of the sclerophyllous species were present. This is not unexpected as riparian plant richness is unique (Sabo et al. 2005), and thus contribution from upland plants can only result in an increase in local richness. On average 48% of the total plant richness was due to the presence of strictly riparian plants, which was almost twice the average contribution of the sclerophyllous plants (28%). This indicates that the riparian community is mainly dominated by strictly riparian plants. Since strictly riparian plants occur in limited numbers (Pollock et al. 1998; Sabo et al. 2005) the maximum richness is truncated, and increases are only possible with the addition of sclerophyllous species. In fact, higher absolute numbers of sclerophyllous than riparian plant species were recorded, and the limitation on maximum richness of strictly riparian species explains the negative slope in the regression between total species richness and strictly riparian plants. However, these results also indicate a constant presence of sclerophyllous species in riparian ecosystems, which may be explained by encroachment of upland species as a response to climate induced reduction in water levels in the upland habitats (Gasith and Resh 1999). This shift towards a water resistant community was also observed in the Tagus river system in Portugal (Aguiar et al. 2006). Alternatively, upland and exotic species are known to use riparian ecosystems for dispersal (Schnitzler et al. 2007), and some of those seeds could have become established either naturally or following disturbance (Aguiar et al. 2001).
Spatial segregation between strictly riparian and sclerophyllous plant patches may be an important factor in determining propagule pressure from sclerophyllous plants species into riparian areas. My results however, only show spatial segregation between transects (between 2 km transects) and not within transects (between 200 m segments of each transect). These results may be explained by the heterogeneity of the riparian ecosystem itself at the finer scale (segments) and that of the landscape at the larger scale (transects). As both natural and human-mediated disturbances create gaps in the riparian ecosystem (Naiman and Décamps 1997; Salinas et al. 2000), this creates opportunities to the establishment of both riparian and sclerophyllous plants in similar locations (Tabacchi et al. 2002).
Environmental variables associated with riparian plant richness
Pollock et al. (1998) demonstrated that species richness in riparian ecosystems is a dynamic equilibrium between disturbance frequency and community productivity. These authors suggested that richness is increased by the presence of patches of different seral stages, which contain different suites of species. In the context of this study, I predicted that a more heterogeneous riparian ecosystem would have higher total woody species richness, which would be mostly due to the presence of sclerophyllous plants in addition to (rather than replacing) strictly riparian plants. The findings in this study corroborate this prediction; as total richness increases, sclerophyllous species richness increases at a similar rate, while riparian species richness has a lower effect (Fig. 2). However, from the negative relationship between richness and presence of human activities it can be inferred that increased sclerophyllus richness does not seem to be a function of the structure of the riparian ecosystem. Human activities in the riparian ecosystem included development of roads, fences, walls, houses, and artificial water channels, which in turn create higher fragmentation and gaps within the riparian vegetation. Furthermore, changes in water rights policies have altered the management prescriptions for riparian zones, allowing neighbouring land-owners to clear-cut riparian trees for easier access to water. These factors have also been identified by other authors as major causes of the decrease in strictly riparian richness in other riparian areas (Aguiar and Ferreira 2005; Hilty and Merenlender 2004; Malanson 1993; Miller 2002; Pollock et al. 1998; Salinas et al. 2000; Tabacchi et al. 2002). However, this effect may be only temporary, matching Pollock et al. (1998) pattern of different seral stages. Younger seral stages will be dominated by riparian plants, and as sclerophyllous species may colonize gaps, mixed mosaics of riparian and sclerophyllous plant species appear as older seral stages, resulting ultimately in an increase in total species richness.
This study results also revealed that riparian species richness (total and strictly riparian) was positively affected by the presence of a developed shrub layer and it was negatively affected by the presence of goats. The most commonly found shrub species in the study area were blackberry shrubs (79.5%), and rock-rose (36.1%). While the first is mostly found in riparian areas, the second is a sclerophyllous shrub. Blackberry shrubs are probably the most related to the observed positive influence on riparian richness, since they are the ones most detected. Blackberry shrubs tend to create a very dense canopy, which may prevent light from reaching the riparian species seeds; however, willows and poplar seeds are known to germinate in the dark (Karrenberg et al. 2002). Thus, blackberry bushes may facilitate the germination seeds from these species, which occurs in a short period (a few days), and also prevent seed mortality from desiccation by providing shade (Karrenberg et al. 2002). At the same time, the thorns and the dense canopy of these shrubs may prevent the saplings from being eaten by goats, which had a marked negative effect on the overall plant richness. In the Mediterranean, cows, sheep and goats share the same forage areas and are separated temporally and behaviorally (Vallentine 2001) by different foraging preferences. Cows are grazers that consume grasses and avoid woody species, sheep are intermediate foragers that consume grasses, forbs and woody species, and goats are browsers that consume forbs and woody species and avoid grasses (Vallentine 2001). Goat foraging period (May–June; Portuguese Associations for Bovine and Ovine and Caprice livestock production, unpublished data) coincides with the time when young woody riparian plants have reached the sapling stage and become more conspicuous, making them more vulnerable to herbivores.
The results showed that strictly riparian plant richness was positively affected by fragmentation (higher number of patches) of the surrounding landscape, and it was negatively affected by the presence of patches of different landscapes (as measured by the landscape diversity indexes). Three factors may contribute to this pattern: the total area covered by the different land covers, diversity of land covers and their density. First, the results indicate that fewer riparian plants are found when larger sclerophyllous patches surround the riparian ecosystem, suggesting that these fewer larger patches may be contributing greater numbers of sclerophyllous plant propagules to the riparian ecosystem. Furthermore, patches of a variety of different land covers (holm oak, cork oak woodlands, olive yards, etc.) have a very negative effect on the strictly riparian plant richness, as the total riparian community is inundated by propagules from different types of plant species, which may have different establishment success rates in the different open patches within the riparian area. Finally, if the surrounding land cover is mainly holm oak woodlands, the frequency of seeds and propagules may actually be reduced since this landscape is characterized by a sparse canopy that is experiencing a decreasing trend in recruitment (Plieninger et al. 2004; Ramirez and Diaz 2008), currently below replenishment rates, and holm oak woodlands do not seem to be exporting seeds elsewhere. This can also explain the negative effect of the area of agriculture on the richness of sclerophyllous plants in the riparian ecosystem. As more agricultural land exists around the riparian area, reduced sclerophyllous seeds exist in the seed pool to colonize the riparian zone.
Data quality assessment
The quality of the interpretation of the results also depends upon the quality of the data input to the models. It is acknowledged that some underestimation may have occurred of species richness as some species lacked key characters that allowed their differentiation. Even though this underestimation may make comparison of these results to those of other authors more difficult, its effect is likely negligible. For both ecophysiological groups of plants (strictly riparian and sclerophyllous) all the species that had diagnostic features were identified, and willow, heath and moor species were the ones harder to diagnose (see Appendix Table 3), which resulted in comparable underestimations. If all the taxa were identified to species level, richness estimates would be increased at most by eight species. Furthermore, the range of variability among riparian systems found in Mediterranean climates was covered, as a large sample size (n = 70) for a study of this type was achieved, and the statistical models were significant and returned a very good fit to the data (ca. 70% of explained variability).
Implications for conservation of riparian ecosystems in semi-arid Mediterranean climates
The change in the composition and structure of riparian ecosystems observed in this study may result in the loss of the suite of ecosystem services they provide, including soil and bank stabilization, water temperature control, water quality regulation, water storage, microclimate moderation, creation of distinct habitats and communities, shelter and nutrients for aquatic and terrestrial organisms, and service as corridors for wildlife (Miller 2002). The maintenance of riparian ecosystems and their role as ecosystem service providers may require creating a riparian plant seed bank and nursery, which could be used for restoration and reforestation of riparian zones. At the larger scale, conservation of riparian ecosystems through limitations in grazing and tramping by livestock and water use regimes is recommended. Future preservation of riparian ecosystems should include monitoring of edge encroachment by upland plants and management to prevent their establishment within a buffer zone around the riparian system, remove existing exotic plants, and preventing or limiting grazing. Further conservation measures should promote riparian ecosystem aesthetic, recreational and service provider values through economic incentives and subsidies, and collaborative campaigns to increase the awareness of these valuable resources that involve land-owners, managers and users. All these measures can be included in a more encompassing and stronger protection policy especially designed for the conservation of the unique biodiversity and services of riparian ecosystems.
References
Aguiar FC, Ferreira MT (2005) Human-disturbed landscapes: effects on composition and integrity of riparian woody vegetation in the Tagus River basin, Portugal. Environ Conserv 32:30–41
Aguiar FC, Ferreira MT, Moreira I (2001) Exotic and native vegetation establishment following channelization of a western Iberian river. Reg Rivers Res Manag 17:509–526
Aguiar FC, Ferreira MT, Albuquerque A (2006) Patterns of exotic and native plant species richness and cover along a semi-arid Iberian river and across its floodplain. Plant Ecol 184:189–202
Bernez I, Ferreira M, Albuquerque A, Aguiar F (2005) Relations between river plant richness in the Portuguese floodplains and the widespread water knotgrass (Paspalum paspalodes). Hydrobiologia 551:121–130
Blondel J (2006) The ‘design’ of mediterranean landscapes: a millennial story of humans and ecological systems during the historic period. Human Ecol 34:713–729
Brookshire ENJ, Kauffman JB, Lytjen D, Otting N (2002) Cumulative effects of wild ungulate and livestock herbivory on riparian willows. Oecologia 132:559–566
Carmel Y, Flather CH (2004) Comparing landscape scale vegetation dynamics following recent disturbance in climatically similar sites in California and the Mediterranean basin. Land Ecol 19:573–590
Chícharo MA, Chícharo LM, Galvão H, Barbosa A, Marques MH, Andrade JP, Esteves E, Miguel C, Gouveia I (2001) Status of the Guadiana Estuary (south Portugal) during 1996–1998: an ecohydrological approach. Aqua Ecos Health Manag: 73–89
Deferrari CM, Naiman RJ (1994) A multiscale assessment of the occurrence of exotic plants on the Olympic Peninsula, Washington. J Veg Sci 5:247–258
ESRI (1996) Arcview GIS––user’s guide version 3.1. Environmental Systems Research Institute Inc
Fortin MJ, Drapeau P, Legendre P (1989) Spatial autocorrelation and sampling design in plant ecology. Vegetatio 83
Gasith A, Resh VH (1999) Streams in Mediterranean climate regions: abiotic influences and biotic responses to predictable seasonal events. Ann Rev Ecol Syst 30:51–81
Hilty JA, Merenlender AM (2004) Use of riparian corridors and vineyards by mammalian predators in northern California. Conserv Biol 18:126–135
Huxman TE, Wilcox BP, Breshears DD, Scott RL, Snyder KA, Small EE, Hultine K, Pockman WT, Jackson RB (2005) Ecohydrological implications of woody plant encroachment. Ecology 86:308–319
Iverson LR, Szafoni DL, Baum SE, Cook EA (2001) A riparian wildlife habitat evaluation scheme developed using GIS. Environ Manag 28:639–654
Jongman R, Pungetti G (2003) Ecological networks and greenways, concept, design, implementation. Cambridge University Press, Cambridge, UK
Karrenberg S, Edwards PJ, Kollmann J (2002) The life history of Salicaceae living in the active zone of floodplains. Freshw Biol 47:733–748
Krebs CJ (1998) Ecological methodology. Addison-Welsey Publishers, Menlo Park, CA., USA
Li H, Wu J (2004) Use and misuse of landscape indices. Land Ecol 19:389–399
Lyon J, Gross NM (2005) Patterns of plant diversity and plant-environmental relationships across three riparian corridors. For Ecol Manag 204:267–278
Machtans CS, Villard M-A, Hannon SJ (1996) Use of riparian buffer strips as movement corridors by forest birds. Conserv Biol 10:1366–1379
Malanson GP (1993) Riparian landscapes. Cambridge University Press, Cambridge, UK
Matos HM, Santos MJ, Palomares F, Santos-Reis M (2008) Does riparian habitat condition influence mammalian carnivore abundance in Mediterranean ecosystems? Biodiv Conserv 18:373–386
McCune B, Grace J (2002) Analysis of ecological communities. MjM Publishers, Gleneden Beach, OR
McGarigal K, Marks BJ (1995) FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. In Gen Tech Rep PNW-GTR-351. USDO Agriculture, F. Service, P.N.R. Station, Portland, OR
Miller C (2002) Conservation of Riparian Forest remnants, West Coast, New Zealand. Land Res 27
Moran PAP (1950) A test for the serial independence of residuals. Biometrika 37:178–181
Naiman RJ, Décamps H (1997) The ecology of interfaces: riparian zones. Ann Rev Ecol Syst 28:621–658
Naiman RJ, Décamps H, McClain ME (2005) Riparia: ecology, conservation, and management of streamside communities. Burlington
Nel JL, Reyers B, Roux DJ, Cowling RM (2009) Expanding protected areas beyond their terrestrial comfort zone: identifying spatial options for river conservation. Biol Conserv 142:1605–1616
Plieninger T, Pulido FJ, Schaich H (2004) Effects of land-use and landscape structure on holm oak recruitment and regeneration at farm level in Quercus ilex L. dehesas. J Arid Environ 57:345–364
Pollock MM, Naiman RJ, Hanley TA (1998) Plant richness in riparian wetlands––a test of biodiversity theory. Ecol Lett 79:194–195
Ramirez JA, Diaz M (2008) The role of temporal shrub encroachment for the maintenance of Spanish holm oak Quercus ilex dehesas. For Ecol Manag 255:1976–1983
Rempel RS, Carr AP (2003) Patch analyst extension for ArcView. Version 3
Rivas-Martinez S (1987) Memoria del Mapa de Series de Vegetación de España. In: Tecnica Serie (ed) I.C.O.N.A.-M.d. Agricultura, P.y. Alimentación. Madrid, Spain
Sabo JL, Sponseller R, Dixon M, Gade K, Harms T, Hefferman J, Jani A, Katz G, Soykan C, Watts J, Welter J (2005) Riparian zones increase regional species richness by harbouring different, not more, species. Ecol Lett 86:56–62
Salinas MJ, Blanca G, Romero AT (2000) Evaluating riparian vegetation in semi-arid Mediterranean watercourses in the south-eastern Iberian Peninsula. Environ Conserv 27:24–35
Sawada M (1999) ROOKCASE: an excel 97/2000 Visual Basic (VB) add-in for exploring global and local spatial autocorrelation. Bull Ecol Soc Am 80:231–234
Scarascia-Mugnozza G, Oswald H, Piussi P, Radoglou K (2000) Forests of the Mediterranean region: gaps in knowledge and research needs. For Ecol Manag 132:97–109
Schnitzler A, Hale BW, Alsum EM (2007) Examining native and exotic species diversity in European riparian forests. Biol Conserv 138:146–156
Schröter D, Cramer W, Leemans R, Prentice C, Araújo MB, Arnell NW, Bondeau A, Bugmann H, Carter TR, Gracia CA, Vega-Leinert ACdl, Erhard M, Ewert F, Glendining M, House JI, Kankaanpää S, Klein RJT, Lavorel S, Lindner M, Metzger MJ, Meyer J, Mitchell TD, Reginster I, Rounsevell M, Sabaté S, Sitch S, Smith B, Smith J, Smith P, Sykes MT, Thonicke K, Thuiller W, Tuck G, Zaehle S, Zierl B (2005) Ecosystem service supply and vulnerability to global change in Europe. Science 310:1333–1337
Spackman SC, Hughes JW (1994) Assessment of minimum stream corridor width for biological conservation: species richness and distribution along mid-order streams in Vermont, USA. Biol Conserv 71:325–332
Tabacchi E, Correll DL, Hauer R, Pinay G, Planty-Tabacchi A-M, Wissmar RC (2002) Development, maintenance and role of riparian vegetation in the river landscape. Freshw Biol 40:497–516
Vallentine JF (2001) Grazing management. Academic Press, San Diego
Virgós E (2001) Relative value of riparian woodlands in landscapes with different forest cover for medium-sized Iberian carnivores. Biodiv Conserv 10:1039–1049
Williams P, Whitfield M, Biggs J, Bray S, Fox G, Nicolet P, Sear D (2003) Comparative biodiversity of rivers, streams, ditches and ponds in an agricultural landscape in Southern England. Biol Conserv 115:329–341
Zar JH (1999) Biostatistical analysis. New Jersey
Acknowledgments
The present paper was funded by the Fundação para Ciência e Tecnologia project “Riparian galleries as Corridors and Linkage Habitats in the Fragmented Landscape of Southern Portugal: Applications to Conservation Planning” (POCTI/MGS/47435/2002; MJS: POCTI/MGS/47435/2002). The author is extremely grateful to Hugo M. Matos for the field work and to Margarida Santos-Reis for the logistic and financial support. Further thanks are due to Ana Rita Alves and Luís Miguel Rosalino for their help during fieldwork, and J. Judson Wynne, Alex Fremier, Margaret Andrew, Francis Bozzolo and Erin Hestir provided helpful improvements to this manuscript.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Santos, M.J. Encroachment of upland Mediterranean plant species in riparian ecosystems of southern Portugal. Biodivers Conserv 19, 2667–2684 (2010). https://doi.org/10.1007/s10531-010-9866-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-010-9866-1