
Abstract
Deer populations and their impacts on forest ecosystems are increasing globally. Given the imperative and expense to mitigate impacts of invasive deer, we aimed to elucidate critical drivers of (i) deer density, (ii) deer impacts, and (iii) the relationship between them, to facilitate targeted management. We used quantile regression forests to model deer density (faecal pellet counts at 1948 locations) and impacts (browsing and other impacts on > 23,000 woody plants at 343 locations) across a mosaic of agricultural and forested ecosystems in Victoria, Australia (12,775 km2). Climate, topography, vegetation cover, and distance to water features were included as model covariates. Modelled deer density (r2 = 0.71, MAE = 0.56 pellets/m2) was most influenced by distance to waterbodies (> 10 ha, 31.2%), elevation (14.3%) and woody vegetation cover (12.9%). Modelled deer impact (r2 = 0.32, MAE = 6.9%) was most influenced by deer density (21.0%), mean annual precipitation (12.8%) and elevation (12.2%). Deer density was typically highest near large waterbodies, at low elevation, and with intermediate tree cover (40–70%). Impacts increased steadily with deer density up to ~ 2 pellets/m2. Our study demonstrates the importance of forest water and forest agricultural interfaces for both deer density and impacts. Deer are likely to be most abundant near waterbodies due to the availability of high-quality forage and water, and prefer lowland locations that have access to both open and forested habitats. Spatial models can be used to predict deer density and associated impacts to facilitate targeted invasive deer management.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Deer populations and their impacts on biodiversity and ecosystem services are increasing in many temperate forests globally (Côté et al. 2004; Davis et al. 2016; Rooney 2001), and these effects can be exacerbated outside of their native range (Wills et al. 2023). Deer impact forest biodiversity through browsing and trampling (Moser, Greet 2018), limiting woody plant recruitment (DiTommaso et al. 2014; Russell et al. 2017), promoting the spread of exotics (Baiser et al. 2008) and thus altering forest composition and structure (e.g., Eichhorn et al. 2017). Such impacts may reduce the suitability of forests as habitat for other wildlife, alter their role in water and nutrient cycling (Comte et al. 2023a), and lower their economic value (Bressette et al. 2012; Wardle et al. 2001).
Effective deer control is often difficult due to uncertainty in the location and density of deer populations, their impacts upon vegetation, and the high cost of management interventions (Bengsen et al. 2020). The design and implementation of cost-effective deer management interventions can be improved by better understanding the drivers of deer population density and impacts upon vegetation at landscape-scales.
The distribution of deer and deer impacts, and the relationships between them, are complex and contingent on climatic and landscape context (Spake et al. 2020). Several modelling approaches have been used to try and understand these relationships (e.g., light detection and ranging (lidar) and random forests, Shanley et al. 2021; and Generalised Linear Models (GLMs), Spake et al. 2020). Typically, these models relate estimates of deer abundance or density to environmental factors such as climate and land use, which are thought to be influential on deer ecology (e.g., Cunningham et al. 2022). However, such models often rely on coarse resolution spatial data or indirect estimates of deer density, for example presence-absence data (Patton et al. 2018), incidences of deer-vehicle collisions (Davies et al. 2019) or general assessments of browser impacts on vegetation (Spake et al. 2020). In this study, we use high-resolution spatial environmental data, combined with consistent and robust indices of deer density (Forsyth et al. 2007) and impact (Bennett et al. 2022), to investigate the relationships between deer density, deer impacts, and the environment.
The aims of this study were to develop spatial models and describe the key environmental drivers of (i) deer density, (ii) deer impacts on vegetation, and (iii) to examine the relationship between deer density and deer impacts to inform targeted management. We develop spatial models of deer density (based on faecal pellet counts) and deer impacts (based on impact assessments of individual woody plants) using quantile regression forests, a machine learning method capable of quantifying predictive uncertainty. Our analyses demonstrate how field observations and publicly available spatial datasets (i.e., describing topography, climate, vegetation, and surface water features) can be used to support informed decision making and provide landscape managers with valuable tools to prioritise and develop effective invasive deer management.
Materials and methods
Study area
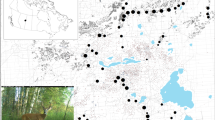
This study focuses on an agricultural and forested landscape near Melbourne, Victoria, south-east Australia, covering an area of approximately 12,775 km2 (Fig. 1). The area comprises mixed land uses and tenures including urban, peri-urban, agricultural, and native forest.

Location of deer density (n = 1948) and impact (n = 343) field surveys across the state of Victoria, Australia. Inset maps show the location of field surveys within the study area encompassing the greater Melbourne region and the location of Victoria on the Australian continent. The Melbourne Drainage and Waterway Extent boundary delineates priority areas for water management in the region
Three non-native deer species that were introduced in the late 1800s to establish populations for hunting (Moriarty 2004) are present in the study area. Sambar deer (Cervus unicolor) are the most widespread and abundant deer species across the region (Forsyth et al. 2018). Fallow deer (Dama dama) and red deer (Cervus elaphus) have much smaller and scattered distributions (Davis et al. 2016). The size of the sambar deer population has increased considerably over the last 30 years (Forsyth et al. 2018), with a corresponding rise in reports of impacts (Davis et al. 2016). Sambar deer are considered generalist browsers consuming a wide variety of trees, shrubs, grasses, and forbs (Forsyth, Davis 2011; Parker 2009; Quin et al. 2023). Fallow and red deer are predominantly grazers but supplement their diet by browsing (Davis et al. 2023; Parker 2009; Roberts et al. 2015).
Field data
Deer density
Field surveys of deer faecal pellet counts were collated from multiple datasets collected across the study area and the broader landscape comprising the state of Victoria (Fig. 1; Table 1). Deer pellets (combined across all species) were counted within 1 m radius plots located at 5 m intervals along 150 m transects (with some exceptions detailed in Table 1) using the Faecal Pellet Index (FPI) method (Forsyth 2005). This method has been widely used to index deer abundance and has a positive and approximately linear relationship with deer density (Forsyth et al. 2007). In total, we included data from 1948 transects with start coordinates, bearings, and transect distance used to spatially reference each transect accounting for bearing changes due to obstructions. The total number of pellets were summed and attributed to the centre location of each transect for spatial modelling.
Deer impact
Deer impacts on vegetation were available from a subset of 343 locations where faecal pellet counts were also collected across multiple field campaigns (Table 1). At 298 locations (Waterway Ecosystem Research Group datasets), we surveyed 120 plants along 150 m transects by selecting four plants for assessment at 5 m intervals (30 points per transect) using the point-centred quadrant method (one per quadrant; Cottam, Curtis 1956; Mitchell 2015). These points corresponded with concurrent faecal pellet counts using the FPI method (Bennett et al. 2022). Additionally, we incorporated data from 45 10 × 10 m unfenced plots (Wills et al. 2023), in which up to 15 trees or shrubs were assessed for impacts. In that study, plants were randomly selected to a maximum of 5 individuals per species, and plant height and an estimate of browser impact recorded (Wills et al. 2023).
Plants were assessed by scoring impact between 0 (no impact) and 4 (high impact; Table 2). The impact score for each plant was converted to a midpoint value based on the percent of the plant browsed or impacted. For this study, each plant > 1 m tall was assessed for impact above and below 1 m separately, and the maximum value attributed to whole plant impact. We used this distinction to account for the potentially confounding effect of native herbivore browsing, which in south-east Australian forests is primarily the swamp wallaby (Wallabia bicolor), a medium-sized macropod that occurs throughout our study area (Eldridge, Coulson 2015). We used 1 m because swamp wallabies usually browse considerably below this height (approx. 60 cm; Bennett 2008). Impacts were not differentiated by height in the study by Wills et al. (2023), and therefore impact height was assumed to be the same as the plant height. In that experimental study, co-located partial (permitting native fauna access) and full exclosures (i.e., no terrestrial fauna access) indicated that browsing impacts recorded in the unfenced plots (i.e., data included in this study) were predominantly attributable to deer, not native fauna. Plant impact percentages were averaged by height category (whole plant, < 1 m, and > 1 m) for each plot or transect. The mean impact percentage for each height category was then attributed to the plot location or centre plot of each transect.
Environmental data
Published models of deer distribution and impact have identified cross-scale interactions between climate, distance to landscape features (e.g., water sources), topography, and floristics as explanatory variables (Davies et al. 2019; Forsyth et al. 2009; Gormley et al. 2011; Moore et al. 2018; Spake et al. 2020). Equivalent variables were collated for our study area where spatially continuous data were publicly available (Table 3).
Climatic conditions could be important for habitat selection within a deer’s geographic range (Spake et al. 2020) and are also important for vegetation growth and health (Hughes 2003). Climate was characterised by mean annual temperature (Stewart, Nitschke 2017a, 2017b, 2018) and precipitation (Fedrigo et al. 2019; Stewart et al. 2020) for the period 1981–2010.
Topographic variables such as elevation have been shown to be influential on the occurrence and density of many deer species including sambar and fallow deer (Cunningham et al. 2022; Davies et al. 2019; Forsyth et al. 2009). Elevation data were sourced from the Victorian state digital terrain model (20 m; DELWP 2016a) and then smoothed using a focal mean filter with a width of 7 pixels (140 m) to capture transect-scale topographic variability. Slope and aspect were computed with the smoothed elevation model. The resulting elevation, slope, and aspect were included as model covariates.
Distances to streams and large waterbodies (> 10 ha) were both used as spatial predictors to characterise resource availability and deer mobility. Each of these variables indicate the presence of drinking water and the potential concentration of preferred food plants adjacent to water (Forsyth et al. 2009). Streams are also potentially important movement corridors (Clements et al. 2011; Opperman, Merenlender 2000; Walter et al. 2011). Two stream products and three waterbody datasets were used to best characterise surface water features across the study area. Streams were represented with (i) Vicmap Hydro 1–25,000 (DELWP 2014a) and (ii) improved regional stream network mapping (Kunapo et al. 2020). Waterbodies (> 10 ha) were represented with (i) the Victorian wetland inventory (DELWP 2016b), (ii) farm dam boundaries (DELWP 2014b), and (iii) improved regional waterbody mapping (Chee et al. 2021). Both streams and waterbodies were rasterised (20 m) and distances to each feature were calculated using the proximity function of GDAL (GDAL/OGR contributors 2022).
Deer often prefer habitat encompassing a mosaic of dense tree cover and open vegetation (Chapman, Chapman 1980; Fattebert et al. 2019; Leslie 2011). Foliage Projected Cover (FPC) was used to describe woody vegetation cover across the landscape (Gill et al. 2017). FPC is the percentage of ground area covered by the vertical projection of foliage. We used a continuous cover estimate of FPC developed using remote sensing products from Landsat satellite imagery, restricted to areas of woody vegetation (Auscover 2015; Gill et al. 2017). We calculated the mean value in a sliding focal window of varying distances from the central pixel (0.5 km, 1 km, 1.5 km, and 2 km) to represent the vegetation mosaic within a typical home range for the three deer species (Amos et al. 2014; Morse et al. 2009; Shea et al. 1990). The 1 km range consistently provided the highest predictive performance, and therefore was used to represent the scale at which deer are most likely to respond to the density of vegetation cover.
Measures such as proximity to roads and road density have been shown to affect deer habitat use (Coe et al. 2018; Comte et al. 2023b; Spake et al. 2020), but we excluded these variables because the resulting surfaces were strongly correlated with distance to streams across the study area.
Modelling and analysis
Deer density and impact on vegetation were modelled with quantile regression forests (Meinshausen 2006), a non-parametric generalisation of random forests that returns conditional quantiles rather than the conditional mean of response variables. The primary advantage of quantile regression forests is that it can be used to predict the expected distribution of the response. This means that they can be used to generate prediction intervals for quantifying uncertainty, and ultimately provide for a more informative product on which to base management decisions.
Models were fitted and evaluated in R 3.6.3 (R Core Team 2020) using the ranger package (v0.14.1; Wright, Ziegler 2017). All quantile regression forests were fitted with 500 trees, which were sufficient to minimise predictive error (Fig. S1). Model performance was evaluated using the root mean squared error (RMSE), mean absolute error (MAE), coefficient of determination (r2) and percentage bias (PBIAS), calculated from a randomly assigned ten fold cross-validation (Kohavi 1995). Detailed descriptions of each performance metric are provided by Moriasi et al. (2007). The Altmann non-parametric permutation test was used to identify variables that significantly contributed to the models (Altmann et al. 2010; p < 0.05).
The deer density model was fitted using each of the environmental variables listed in Table 3, using faecal pellet counts at the 1948 transects as the response variable. The impact of deer on vegetation was modelled using scoring from each of the 343 locations as the response, and all environmental variables (Table 3), in addition to the modelled deer density (calibrated with the state-wide dataset) as covariates. To evaluate the effect of using modelled deer density as a spatial covariate for predicting impact, a further series of impact models were fitted with only environmental variables (Table 3). Unless otherwise specified, all results pertain to the deer impact models that include deer density as a model covariate. Models were fitted for impacts at < 1 m, > 1 m, and for the whole plant. We tested multiple impact heights to better understand how deer impacts may be distinguished from those of native herbivores that browse at lower heights. There were 18257 individual plant assessments used for the < 1 m model, 13741 for the > 1 m model, and 23144 for the whole plant model.
Several analyses were conducted to investigate any potentially confounding effects arising from spatial and/or temporal autocorrelation, as many of the observations are drawn from repeat samples over time (see Table 1). Temporal trends were investigated by regressing observations and model residuals against survey dates. Spatial autocorrelation was evaluated by calculating semi-variance at binned spatial lag distances and plotting the empirical variogram for both observations and model residuals. There was no evidence of spatial or temporal autocorrelation (Figs. S2, S3) that would lead to biased model predictions, despite the reported increase in deer abundance across the study area since 2005 (Forsyth et al. 2018). Spatial and temporal effects were therefore not included in the model design to avoid potentially confounding results, particularly given that year-to-year variability in deer management activity, survey locations (i.e., as new locations are added) and population dynamics will also have an impact on the survey data.
Results
Deer pellet density modelling
Statistical performance of the deer density model (Table 4) was strong (r2 = 0.71, MAE = 0.56 pellets/m2). There was a tendency to underpredict deer density (PBIAS = -28.0%), but this was expected given the distribution of the data (median = 0.03 pellets/m2, mean = 0.80 pellets/m2, standard deviation = 2.67 pellets/m2). The relationship between the cross-validated deer density predictions, observations, and 90% prediction intervals (5th to 95th quantile) are illustrated in Fig. 2. Observed deer density was within the 5th and 95th quantiles for 88.6% of cross-validated predictions (Fig. 2b). The high proportion of observations falling within prediction intervals less than 90% (Fig. 2b) reflect the distribution of the data and low overall density of faecal pellet counts.

Cross-validated versus observed deer density (pellets/m2; n = 1948) with 90% prediction intervals (quantile 5–95; a), and the proportion of observations within prediction intervals at different ranges centred on quantile 50 (b). Observed values are aligned vertically with the prediction interval (a) and overlap where they occur within this range
Distance to waterbodies > 10 ha was the most influential predictor of deer density, with a relative influence of 31.2% (Fig. 3a). Predicted deer densities increased with proximity to waterbodies, particularly in areas directly adjacent to them (Fig. S4). Variable contributions were also statistically significant (via permutation testing) for elevation (14.3%), woody vegetation cover (12.9%), mean annual precipitation (11.3%) and mean annual temperature (10.2%). Partial dependence plots (Fig. 4) indicate that deer pellet densities are predicted to be higher at elevations < 400 m, and at intermediate levels of woody vegetation cover (~ 40–70%). Deer pellet density tended to increase with mean annual precipitation up to 1300 mm and decrease with mean annual temperatures > 12.5°C. Slope, aspect, and distance to streams had less influence on predicted deer density.

Relative influence of predictor variables for (a) deer density (pellets/m2; n = 1948), and (b) deer impact (whole plant, %; n = 343) models. Values in parentheses are the percent relative influence of each variable to the model (scaled to total 100%). Variables with an asterisk denote significant (p < 0.05) contributions to the model determined by the Altmann non-parametric permutation test
The highest predicted densities of deer are in eastern regions of the study area (Fig. 4), which are cooler and wetter, and are often located along the interface between agricultural and forested areas, and around the perimeter of large waterbodies. Lower deer densities were predicted in drier and warmer areas in the west of the study area. Outputs from both the deer density and deer impact models are freely available on the CSIRO Data Access Portal (Fedrigo et al. 2023;https://doi.org/10.25919/7gxj-d455).

Spatial predictions of deer density modelled using quantile regression forests
Deer impact modelling
Cross-validation statistics for the deer impact models are presented in Table 4. The whole plant impact model performed best overall (r2 = 0.32, MAE = 6.86%) with the least amount of bias (PBIAS = -2.65%). The relationship between the cross-validated (whole plant) deer impact predictions, observations, and prediction intervals (5th to 95th quantile) are illustrated in Fig. 5. Observed deer impact was within the 5th and 95th quantiles for 88.6% of cross-validated predictions (Fig. 5b).

Cross-validated versus observed deer impact (whole plant, %; n = 343) with 90% prediction intervals (quantile 5–95; a), and the proportion of observations within prediction intervals at different ranges centred on quantile 50 (b). Observed values are aligned vertically with the prediction interval (a) and overlap where they occur within this range
Deer density and mean annual precipitation were statistically significant (via permutation tests) predictors of whole plant deer impact (Fig. 3b). Deer density was the most influential predictor (21.0%), followed by mean annual precipitation (12.8%), elevation (12.2%), and woody vegetation cover within 1 km (10.6%). Modelling deer impact without deer density as a predictor consistently reduced statistical performance (Δ r2 = -0.07 to -0.04, Δ MAE = -0.32 to -0.26; see Table S1), and partial dependence plots show analogous response curves for environmental variables across both whole tree impact models (Figs. S5, S6). Deer impact was predicted to increase steadily to deer densities of ~ 2 pellets/m2 (Fig. S5). Modelled deer impact was highest at elevations < 200 m and low at elevations > 600 m. Mean annual precipitation > 1350 mm was associated with lower predicted deer impact. Impact was greatest at intermediate to high levels of woody vegetation cover (above 50%). Deer impact was also predicted to be higher in areas closer to large waterbodies and streams, and on slopes with easterly or southerly aspects.
Predictions of deer impact across our study area are illustrated in Fig. 6. The highest predicted impacts are found in the eastern region of the study area, coinciding with lower elevation, intermediate forest cover, and moderate mean annual precipitation. Deer impact was predicted to be lower in drier regions in the west of the study area; however, limited field survey data were available for these areas.

Spatial predictions of whole plant deer impact modelled using quantile regression forests
Deer density and impact relationship
Deer density, as measured by faecal pellet counts, had a significant (p < 0.001) weak positive correlation (Kendall’s τ = 0.28) with deer impact (see Fig. S7) and contributed 21.0% relative variable importance to the whole plant impact model. Deer impact was modelled with 6 times fewer data points than deer density, concentrated within a smaller geographic area, and were additionally influenced by other environmental predictors. Modelling deer impact with and without deer density shows that deer impacts were constrained in areas of low deer density and increased in areas of high deer density (Fig. 4; Fig. S8).
Forested areas near large waterbodies or agricultural land and with intermediate levels of forest cover were often associated with both high deer density and impacts (Fig. 7). For example, the margins of the large Upper Yarra Reservoir (750 ha; Fig. 7a, top row) experience alternating periods of water inundation and exposure that supports dense herbaceous vegetation (Bennett 2008). Deer density and impacts are both predicted to be high in this region (Fig. 7a, centre and bottom rows), likely due to the accessibility of high-quality forage, water, and forest cover. Conversely, modelled deer density and impact did not respond to the sharp forest-agriculture boundary (e.g., at Bunyip State Park; Fig. 7b, top row). Both deer density and impacts were low within densely forested areas (> 70% woody vegetation cover) but were high in patches of forest along riparian corridors, commonly within mosaics of forest and agricultural land (Fig. 7b).

Regions of high deer density and impact modelled across (a) forest–water reservoir (Upper Yarra Reservoir) and (b) forest–agriculture (Bunyip State Forest) boundaries. Satellite imagery (top row) provides context for deer density (centre row) and impact (bottom row). Inset maps show the location of each region within the study area and broader Australian extent
Discussion
Deer density and impact modelling
Our modelled predictions, using high-resolution spatial data and extensive field data, demonstrate the importance of forest water and forest agricultural interfaces for both deer density and impacts. Deer are likely to be abundant in the vicinity of waterbodies due the availability of high-quality forage and water, and prefer locations with access to both open and forested habitats. At low to moderate deer densities, deer impacts are likely to increase with small increases in density, while at high densities, impacts become dependent on environmental and landscape context.
Tree cover within 1 km was influential on both deer density and deer impacts. The critical importance of woody vegetation to various deer species globally for providing forage, cover and bedding is well established (Avey et al. 2003; Borkowski, Ukalska 2008; Coe et al. 2018). Our finding that deer prefer intermediate levels of tree cover concurs with studies of deer populations in North America (Avey et al. 2003; Coe et al. 2018) and deer impacts in Britain (Spake et al. 2020). The ‘humped’ response of deer density to vegetation cover density (Figs. S4, S5) is likely due to deer preferring locations with access to both forested (for cover during the daytime) and open habitats (for nocturnal foraging; Borkowski, Pudełko 2007; Comte et al. 2023b; Fattebert et al. 2019).
Proximity to large waterbodies was highly influential on both deer density and, to a lesser extent, on deer impacts. The importance of water for deer is well known (Brunjes et al. 2006; Dinerstein 1979; McKay, Eisenberg 1974), including for sambar deer (Comte et al. 2022; Forsyth et al. 2009), the most abundant deer species in our study area. As well as providing water for deer to drink, areas surrounding large waterbodies also support an abundance of high-quality forage (Brunjes et al. 2006; Forsyth et al. 2009).
Lower elevations were preferred by deer in our study area and were subject to greater impact. Elevation preferences of deer vary between deer species and seasonally according to snow cover, food availability and breeding status (Brunjes et al. 2006; Comte et al. 2022; Zhang et al. 2013). Fallow deer prefer flat, lowland areas (Chapman, Chapman 1980; Cunningham et al. 2022), but sambar and red deer occur across a wide range of elevations within their native habitat (Leslie 2011; Luccarini et al. 2006; Whitehead 1993) and, in the Australian Alps, sambar deer descend from higher to lower elevations in colder months to avoid snow cover (Comte et al. 2022). The higher elevation locations within our study area (and throughout most of south-east Australia) are characterised by continuous, dense forest where snowfalls are uncommon, highlighting the greater importance of proximity to waterbodies and intermediate levels of woody vegetation cover to provide habitat.
Drier areas (mean annual precipitation < 700 mm) in the west of our study area were least preferred, while wetter areas (mean annual precipitation > 1350 mm) had low predicted deer impacts, potentially due to the reduced suitability of these areas to fallow deer (Cunningham et al. 2022). Other geographic variables we used as model covariates (e.g., slope, aspect) were only moderately, at best, associated with deer density and impact. However, the tendency for higher deer density and impact on easterly and southerly aspects is consistent with previous modelling of sambar density in the study area (Forsyth et al. 2009). It is possible that preferences for these factors vary seasonally, or that clear relationships were obscured by deer species-specific preferences.
Our model predicted that deer impact increases steadily from low to moderate deer density (up to ~ 2 pellets/m2), but that at high deer density impact was dependent on environmental and landscape context. This indicates that the native woody vegetation assessed is highly vulnerable to impacts from non-native deer (Norbury et al. 2015; Nugent et al. 2001), and concords with recent experimental findings in our study area (Wills et al. 2023). While both deer density and impacts were predicted to be greatest along forest water and forest agricultural interfaces, these relationships were stronger for density, with a wider range of environmental factors influencing impacts.
Our models include predictions of deer density and impact for unsurveyed habitats including locations where access by deer is somewhat restricted, such as French and Phillip Islands located in Westernport Bay, and the Central Business District (CBD) of Melbourne. Deer are established on French Island (Davies et al. 2022) and periodic incursions may occur on both islands (deer can purportedly swim to these sites; Forsyth et al. 2015). Deer have also been reported < 5 km from Melbourne’s CBD, most likely using the Yarra River as a corridor for movement (DELWP 2021). Collection of additional field data from currently unsampled habitats with unique combinations of environmental and climatic data (e.g., coastal areas) would assist to refine and improve certainty of the density and impact predictions in these locations.
Our approach and predictions represent an improvement on previous attempts to model deer density and impacts. We used high-resolution climatic, topographic, and remote sensing derived variables that well represent environmental gradients in mosaic landscapes with topographically variable terrain. Quantile regression forests provided a method for capturing complex, non-linear responses, and variable interactions, and furthermore, enabled the generation of prediction intervals that can be used to quantify uncertainty in model predictions. The strong response of deer density to waterbodies and vegetation cover suggests that access to resources and suitable habitat are more important constraints than broad-scale climatic variables typically used in species distribution models.
Management implications
Successful management of invasive deer populations typically occur over medium- to long-term time frames, and are influenced by landscape context including size and shape of the management area and proximity to other habitat features such as adjacent refugia (Bengsen et al. 2020; Comte et al. 2023a). Land managers require spatial information (i.e., maps) to understand the landscape context, and support informed decision making and the implementation of cost-effective deer control strategies (Putman et al. 2011).
We found that deer have strong preferences for forest water and forest agricultural interfaces. This is in accordance with studies that have identified the attractiveness of landscapes with a mosaic of vegetation types to various deer species (Avey et al. 2003; Borkowski, Ukalska 2008; Brunjes et al. 2006; Fattebert et al. 2019). Such landscapes are thus likely to require targeted efforts to reduce invasive or overabundant deer populations and their impacts.
The affinity of deer to forest water and forest agricultural interfaces indicates that human land use change is an important driver of deer population distribution and a process by which many deer populations have become overabundant (Côté et al. 2004). Human settlement and agriculture typically require access to permanent water sources and the increase in the abundance and extent of waterbodies such as dams and reservoirs are likely to also increase the favourability of landscapes to deer. Similarly, increased fragmentation of treed landscapes through forest harvesting, urbanisation or fire is likely to increase the suitability of landscapes to deer, promoting the potential for deer invasion or overabundance (Côté et al. 2004; Fattebert et al. 2019).
Our finding that deer impact increases steadily from low to moderate deer density suggests that reduction of deer population to very low densities are required to reduce impact to low levels (Nugent et al. 2001). This concurs with previous findings that, even at low densities, introduced herbivores may restrict ecosystem recovery (e.g., Tanentzap et al. 2009). Targeted impact mitigation may be necessary to prioritise threatened plant species or communities where management resources are limited. Predictive spatial models, such as presented here, can help to prioritise deer management actions.
Our models did not consider several potentially important variables, including roads and seasonal variation. Deer may use different habitats seasonally (e.g., Comte et al. 2022; Zhang et al. 2013), but we did not include seasonality in our models because deer faecal pellets can persist for ~ 12 months (Davis, Coulson 2016) and therefore we could not reliably attribute them to one month or season. For local management decisions, seasonal and fine-scale variation in habitat use by deer, as well as potential interactions with sympatric deer (Brunjes et al. 2006) and native browser species may be important to consider.
Conclusion
The design and implementation of cost-effective deer management can be supported with an improved understanding of deer population density and corresponding impacts on vegetation. This study demonstrates a spatial modelling approach to mapping deer density and impact using field-based measurements in conjunction with machine learning. We found that faecal pellets, as a proxy for deer density, can be modelled with moderate to high predictive accuracy at broad spatial scales. Predictions of deer impact were highly influenced by deer density, although these models did not perform as well statistically. The resulting spatial products (including estimates of uncertainty) provide land and water managers with valuable resources for developing targeted and cost-effective deer management strategies.
Data availability
The deer density and impact model outputs generated as part of this study are accessible on the CSIRO Data Access Portal (Fedrigo et al. 2023; https://doi.org/10.25919/7gxj-d455).
References
Altmann A, Toloşi L, Sander O et al (2010) Permutation importance: a corrected feature importance measure. Bioinformatics 26:1340–1347. https://doi.org/10.1093/bioinformatics/btq134
Amos M, Baxter G, Finch N et al (2014) At home in a new range: wild red deer in south-eastern Queensland. Wildl Res 41:258–265. https://doi.org/10.1071/WR14034
Avey J, Ballard W, Wallace M et al (2003) Habitat relationships between a sympatric mule and white-tailed deer Texas. Southwest Nat 48:644–653. https://doi.org/10.1894/0038-4909(2003)048%3c0644:HRBSMD%3e2.0.CO;2
Baiser B, Lockwood JL, La Puma D et al (2008) A perfect storm: two ecosystem engineers interact to degrade deciduous forests of New Jersey. Biol Invasions 10:785–795. https://doi.org/10.1007/s10530-008-9247-9
Bengsen AJ, Forsyth DM, Harris S et al (2020) A systematic review of ground-based shooting to control overabundant mammal populations. Wildl Res 47:197–207. https://doi.org/10.1071/WR19129
Bennett A (2008) The impacts of sambar (Cervus unicolor) in the Yarra Ranges National Park. PhD Thesis. The University of Melbourne, Parkville, Australia
Bennett A, Fedrigo M, Greet J (2022) A field method for rapidly assessing deer density and impacts in forested ecosystems. Ecol Manag Restor 23:81–88. https://doi.org/10.1111/emr.12518
Borkowski J, Pudełko M (2007) Forest habitat use and home-range size in radio-collared fallow deer. Ann Zool Fenn 44:107–114
Borkowski J, Ukalska J (2008) Winter habitat use by red and roe deer in pine-dominated forest. For Ecol Manage 255:468–475. https://doi.org/10.1016/j.foreco.2007.09.013
Bressette JW, Beck H, Beauchamp VB (2012) Beyond the browse line: complex cascade effects mediated by white-tailed deer. Oikos 121:1749–1760. https://doi.org/10.1111/j.1600-0706.2011.20305.x
Brunjes KJ, Ballard WB, Humphrey MH et al (2006) Habitat use by sympatric mule and white-tailed deer in Texas. J Wildl Manag 70:1351–1359. https://doi.org/10.2193/0022-541X(2006)70[1351:HUBSMA]2.0.CO;2
Chapman NG, Chapman DI (1980) The distribution of fallow deer: a worldwide review. Mammal Rev 10:61–138. https://doi.org/10.1111/j.1365-2907.1980.tb00234.x
Clements GM, Hygnstrom SE, Gilsdorf JM et al (2011) Movements of white-tailed deer in riparian habitat: implications for infectious diseases. J Wildl Manag 75:1436–1442. https://doi.org/10.1002/jwmg.183
Coe PK, Clark DA, Nielson RM, Gregory SC, Cupples JB, Hedrick MJ, Johnson BK, Jackson DH (2018) Multiscale models of habitat use by mule deer in winter. J Wildl Manag 82(6):1285–1299. https://doi.org/10.1002/jwmg.21484
Comte S, Thomas E, Bengsen AJ et al (2022) Seasonal and daily activity of non-native sambar deer in and around high-elevation peatlands, south-eastern Australia. Wildl Res 49:659–672. https://doi.org/10.1071/WR21147
Comte S, Bengsen AJ, Thomas E et al (2023a) A before-after control-impact experiment reveals that culling reduces the impacts of invasive deer on endangered peatlands. J Appl Ecol 60:2340–2350. https://doi.org/10.1111/1365-2664.14498
Comte S, Thomas E, Bengsen AJ et al (2023b) Cost-effectiveness of volunteer and contract ground-based shooting of sambar deer in Australia. Wildl Res 50:642–656. https://doi.org/10.1071/WR22030
Côté SD, Rooney TP, Tremblay J-P et al (2004) Ecological impacts of deer overabundance. Annu Rev Ecol Evol Syst 35:113–147. https://doi.org/10.1146/annurev.ecolsys.35.021103.105725
Cottam G, Curtis JT (1956) The use of distance measures in phytosociological sampling. Ecology 37:451–460. https://doi.org/10.2307/1930167
Cunningham C, Perry G, Bowman D et al (2022) Dynamics and predicted distribution of an irrupting ‘sleeper’ population: fallow deer in Tasmania. Biol Invasions 24:1131–1147. https://doi.org/10.1007/s10530-021-02703-4
Davies C, Wright W, Hogan F et al (2019) Predicting deer–vehicle collision risk across Victoria, Australia. Aust Mammalogy 42:293–301. https://doi.org/10.1071/AM19042
Davies C, Wright W, Wedrowicz F et al (2022) Delineating genetic management units of sambar deer (Rusa unicolor) in south-eastern Australia, using opportunistic tissue sampling and targeted scat collection. Wildl Res 49:147–157. https://doi.org/10.1071/WR19235
Davis NE, Bennett A, Forsyth DM et al (2016) A systematic review of the impacts and management of introduced deer (family Cervidae) in Australia. Wildl Res 43:515–532. https://doi.org/10.1071/WR16148
Davis NE, Coulson G (2016) Habitat-specific and season-specific faecal pellet decay rates for five mammalian herbivores in south-eastern Australia. Aust Mammal 38:105–116. https://doi.org/10.1071/AM15007
Davis NE, Forsyth DM, Bengsen AJ (2023) Diet and impacts of non-native fallow deer (Dama dama) on pastoral properties during severe drought. Wildl Res 50:701–715. https://doi.org/10.1071/WR22106
DELWP (2021). Peri-urban deer control plan 2021–26. Department of environment, land, water and planning, state of Victoria, Australia. Accessed on 15 Oct 2023. https://www.environment.vic.gov.au/__data/assets/pdf_file/0026/563660/Peri-Urban-Deer-Control-Plan-2021-2026.pdf
Dinerstein E (1979) An ecological survey of the royal Karnali-Bardia wildlife reserve, Nepal. Part II: habitat/animal interactions. Biol Conserv 16:265–300. https://doi.org/10.1016/0006-3207(79)90055-7
DiTommaso A, Morris SH, Parker JD et al (2014) Deer browsing delays succession by altering aboveground vegetation and belowground seed banks. PLoS ONE 9:e91155. https://doi.org/10.1371/journal.pone.0091155
Eichhorn MP, Ryding J, Smith MJ et al (2017) Effects of deer on woodland structure revealed through terrestrial laser scanning. J Appl Ecol 54:1615–1626. https://doi.org/10.1111/1365-2664.12902
Eldridge MDB, Coulson GM (2015) Family macropodidae (Kangaroos and Wallabies). In: Wilson DE and Mittermeier RA (eds) Handbook of the Mammals of the World. Vol. 5 Monotremes and Marsupials Lynx Edicions, Barcelona, Spain, pp. 630–735
Fattebert J, Morelle K, Jurkiewicz J et al (2019) Safety first: seasonal and diel habitat selection patterns by red deer in a contrasted landscape. J Zool 308:111–120. https://doi.org/10.1111/jzo.12657
Fedrigo M, Stewart SB, Roxburgh SH et al (2019) Predictive ecosystem mapping of south-eastern Australian temperate forests using lidar-derived structural profiles and species distribution models. Remote Sensing 11:93. https://doi.org/10.3390/rs11010093
Forsyth DM (2005) Protocol for estimating changes in the abundance of deer in New Zealand forests using the Faecal Pellet Index (FPI). New Zealand department of conservation, Landcare Research New Zealand Ltd, Wellington, New Zealand
Forsyth DM, Barker RJ, Morriss G et al (2007) Modeling the relationships between fecal pellet indices and deer density. J Wildl Manag 71:964–970. https://doi.org/10.2193/2005-695
Forsyth DM, McLeod SR, Scroggie MP et al (2009) Modelling the abundance of wildlife using field surveys and GIS: non-native sambar deer (Cervus unicolor) in the Yarra ranges, south-eastern Australia. Wildl Res 36:231–241. https://doi.org/10.1071/WR08075
Forsyth DM, Davis NE (2011) Diets of non-native deer in Australia estimated by macroscopic versus microhistological rumen analysis. J Wildl Manag 75:1488–1497. https://doi.org/10.1002/jwmg.179
Forsyth DM, Caley P, Davis NE et al (2018) Functional responses of an apex predator and a mesopredator to an invading ungulate: dingoes, red foxes and sambar deer in south-east Australia. Austral Ecol 43:375–385. https://doi.org/10.1111/aec.12575
Forsyth DM, Stamation K, Woodford L (2015) Distributions of sambar deer, rusa deer and sika deer in Victoria. Arthur Rylah institute for environmental research unpublished client report for the biosecurity branch, department of economic development, jobs, transport and resources. Department of environment, land, water and planning, Heidelberg, Victoria
GDAL/OGR contributors (2022) GDAL/OGR geospatial data abstraction software library. Open source geospatial foundation, URL: https://gdal.org; https://doi.org/10.5281/zenodo.5884351
GHD (2015) Pest animal and riparian vegetation condition monitoring and management in closed catchments: development of a conceptual model and monitoring framework for impacts of deer. Melbourne, Australia
Gill T, Johansen K, Phinn S et al (2017) A method for mapping Australian woody vegetation cover by linking continental-scale field data and long-term Landsat time series. Int J Remote Sens 38:679–705. https://doi.org/10.1080/01431161.2016.1266112
Gormley AM, Forsyth DM, Griffioen P et al (2011) Using presence-only and presence–absence data to estimate the current and potential distributions of established invasive species. J Appl Ecol 48:25–34. https://doi.org/10.1111/j.1365-2664.2010.01911.x
Hughes L (2003) Climate change and Australia: trends, projections and impacts. Austral Ecol 28(4):423–443. https://doi.org/10.1046/j.1442-9993.2003.01300.x
Kohavi R (1995) A study of cross-validation and bootstrap for accuracy estimation and model selection. Int Joint Conf Artif Intell 14:1137–1145
Leslie DM (2011) Rusa unicolor (Artiodactyla: Cervidae). Mamm Species 43:1–30. https://doi.org/10.1644/871.1
Luccarini S, Mauri L, Apollonio M et al (2006) Red deer (Cervus elaphus) spatial use in the Italian Alps: home range patterns, seasonal migrations, and effects of snow and winter feeding. Ethol Ecol Evol 18:127–145. https://doi.org/10.1080/08927014.2006.9522718
McKay GM, Eisenberg JF (1974) Movement patterns and habitat utilization of ungulates in Ceylon. In: Geist V, Walther F (eds) The behaviour of ungulates and its relation to management, vol 2. IUCN. Morges, Switzerland, pp 708–721
Meinshausen N (2006) Quantile regression forests. J Mach Learn Res 7:983–999
Mitchell K (2015) Quantitative analysis by the point-centered quarter method. Available at: https://arxiv.org/abs/1010.3303. Department of mathematics and computer science, Hobart and William Smith Colleges, Geneva, New York
Moore EK, Iason GR, Pemberton JM et al (2018) Habitat impact assessment detects spatially driven patterns of grazing impacts in habitat mosaics but overestimates damage. J Nat Conserv 45:20–29. https://doi.org/10.1016/j.jnc.2018.07.005
Moriarty A (2004) The liberation, distribution, abundance and management of wild deer in Australia. Wildl Res 31:291–299. https://doi.org/10.1071/WR02100
Moriasi DN, Arnold JG, Van Liew MW et al (2007) Model evaluation guidelines for systematic quantification of accuracy in watershed simulations. Trans ASABE 50:885–900. https://doi.org/10.13031/2013.23153
Morse BW, Nibbelink NP, Osborn DA et al (2009) Home range and habitat selection of an insular fallow deer (Dama dama L.) population on Little St. Simons Island, Georgia, USA. Eur J Wildl Res 55:325–332. https://doi.org/10.1007/s10344-008-0245-0
Moser S, Greet J (2018) Unpalatable neighbours reduce browsing on woody seedlings. For Ecol Manag 414:41–46. https://doi.org/10.1016/j.foreco.2018.02.015
Norbury GL, Pech RP, Byrom AE et al (2015) Density-impact functions for terrestrial vertebrate pests and indigenous biota: guidelines for conservation managers. Biol Conserv 191:409–420. https://doi.org/10.1016/j.biocon.2015.07.031
Nugent G, Fraser W, Sweetapple P (2001) Top down or bottom up? Comparing the impacts of introduced arboreal possums and “terrestrial” ruminants on native forests in New Zealand. Biol Conserv 99:65–79. https://doi.org/10.1016/S0006-3207(00)00188-9
Opperman JJ, Merenlender AM (2000) Deer herbivory as an ecological constraint to restoration of degraded riparian corridors. Restor Ecol 8:41–47. https://doi.org/10.1046/j.1526-100x.2000.80006.x
Parker BD (2009) Feeding biology of sympatric sambar (Cervus unicolor) and fallow deer (Dama dama) in NE Victoria. PhD Thesis. The University of Melbourne, Creswick, Australia
Victoria P (1998) French Island national park management plan. Parks Victoria, Melbourne
Patton SR, Russell MB, Windmuller-Campione MA et al (2018) Quantifying impacts of white-tailed deer (Odocoileus virginianus Zimmerman) browse using forest inventory and socio-environmental datasets. PLoS ONE 13:e0201334. https://doi.org/10.1371/journal.pone.0201334
Putman R, Watson P, Langbein J (2011) Assessing deer densities and impacts at the appropriate level for management: a review of methodologies for use beyond the site scale. Mammal Rev 41:197–219. https://doi.org/10.1111/j.1365-2907.2010.00172.x
Quin MJ, Morgan JW, Murphy NP (2023) Spatial and temporal variation in the diet of introduced sambar deer (Cervus unicolor) in an alpine landscape. Wildl Res. https://doi.org/10.1071/WR23017
R Core Team (2020) R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. URL: https://www.R-project.org/
Roberts C, Westbrooke M, Florentine S et al (2015) Winter diet of introduced red deer (Cervus elaphus) in woodland vegetation in Grampians National Park, western Victoria. Aust Mammal 37:107–112. https://doi.org/10.1071/AM14013
Rooney TP (2001) Deer impacts on forest ecosystems: a North American perspective. Forestry 74:201–208. https://doi.org/10.1093/forestry/74.3.201
Russell MB, Westfall JA, Woodall CW (2017) Modeling browse impacts on sapling and tree recruitment across forests in the northern United States. Can J For Res 47:1474–1481. https://doi.org/10.1139/cjfr-2017-0155
Shanley CS, Eacker DR, Reynolds CP et al (2021) Using LiDAR and random forest to improve deer habitat models in a managed forest landscape. For Ecol Manag 499:119580. https://doi.org/10.1016/j.foreco.2021.119580
Shea SM, Flynn LB, Marchinton RL, et al. (1990) Part II: social behaviour, movement ecology and food habits. In: Tall Timbers Research Station (ed) Ecology of sambar deer on St. Vincent national wildlife refuge, Florida. Tall Timbers Research Station, Tallahassee, USA, pp. 13–62
Spake R, Bellamy C, Gill R et al (2020) Forest damage by deer depends on cross-scale interactions between climate, deer density and landscape structure. J Appl Ecol 57:1376–1390. https://doi.org/10.1111/1365-2664.13622
Stewart SB, Nitschke CR (2017a) Improving temperature interpolation using MODIS LST and local topography: a comparison of methods in south east Australia. Int J Climatol 37:3098–3110. https://doi.org/10.1002/joc.4902
Tanentzap AJ, Larry EB, William GL et al (2009) Landscape-level vegetation recovery from herbivory: progress after four decades of invasive red deer control. J Appl Ecol 46:1064–1072. https://doi.org/10.1111/j.1365-2664.2009.01683.x
Walter WD, Baasch DM, Hygnstrom SE et al (2011) Space use of sympatric deer in a riparian ecosystem in an area where chronic wasting disease is endemic. Wildl Biol 17:191–209. https://doi.org/10.2981/10-055
Wardle DA, Barker GM, Yeates GW et al (2001) Introduced browsing mammals in New Zealand natural forests: aboveground and belowground consequences. Ecol Monogr 71:587–614. https://doi.org/10.1890/0012-9615(2001)071[0587:IBMINZ]2.0.CO;2
Whitehead GK (1993) The whitehead encyclopedia of deer. Swan Hill Press, Shrewsbury
Wills TJ, Retallick RWR, Greet J et al (2023) Browsing by non-native invasive sambar deer dramatically impacts forest structure. For Ecol Manag 543:121153. https://doi.org/10.1016/j.foreco.2023.121153
Wright MN, Ziegler A (2017) Ranger: a fast implementation of random forests for high dimensional data in C++ and R. J Stat Soft 77:1–17. https://doi.org/10.18637/jss.v077.i01
Zhang M, Liu Z, Teng L (2013) Seasonal habitat selection Cervus elaphus alxaicus in the Helan mountains, China. Zoologia 30:24–34. https://doi.org/10.1590/S1984-46702013000100003
DATA SOURCES
Auscover (2015). Foliage projective cover. Download dm7** products. terrestrial ecosystem research network Auscover. http://qld.auscover.org.au/public/data/remote-sensing/landsat/persistent_green/v2/
Bennett A, Haydon S, Stevens M et al (2015) Culling reduces fecal pellet deposition by introduced sambar (Rusa unicolor) in a protected water catchment. Wildl Soc Bull 39:268–275. https://doi.org/10.1002/wsb.522
Bennett A (2013) Deer in Yellingbo nature conservation reserve, Victoria. a report prepared for melbourne water. Unpublished client report for Melbourne water. The University of Melbourne, Parkville
Chee YE, Kunapo J, Steele W, van Burm E, Shackleton M, and Walsh C (2021) Waterbodies spatial database for the Melbourne water region (version: mWregion_Waterbodies_v1.3)—metadata report. Melbourne waterway research-practice partnership. School of ecosystem and forest sciences, The University of Melbourne, Australia
DELWP (2014a). Vicmap hydro 1:25,000. Department of environment, land, water and planning, state of Victoria, Australia. Published 01/08/2014. https://discover.data.vic.gov.au/dataset/vicmap-hydro-1-25-000
DELWP (2014b). Farm dam boundaries. Department of environment, land, water and planning, state of Victoria, Australia. Published on 01/08/2014. https://discover.data.vic.gov.au/dataset/farm-dam-boundaries
DELWP (2016a). Vicmap elevation dtm 20m. Department of environment, land, water and planning, state of Victoria, Australia. Published on 24/03/2016. http://discover.data.vic.gov.au/dataset/vicmap-elevation-dtm-20m
DELWP (2016b). Victorian wetland inventory current. Department of environment, land, water and planning, state of Victoria, Australia. Published on 08/11/2016. https://discover.data.vic.gov.au/dataset/victorian-wetland-inventory-current
Fedrigo M, Bennett A, Stewart S, Forsyth D, Greet J (2023): Modelled deer density and impact on vegetation across the Melbourne drainage and waterway extent, Victoria. v4. CSIRO. Data Collection. https://doi.org/10.25919/7gxj-d455
Forsyth DM, Gormley AM, Woodford L et al (2012) Effects of large-scale high-severity fire on occupancy and abundances of an invasive large mammal in south-eastern Australia. Wildl Res 39:555–564. https://doi.org/10.1071/WR12033
Gormley A, Jameson G, Scroggie MP, et al. (2009) Recent (2005−2008) trends in the abundance of deer at eight sites in Victoria. Arthur Rylah institute for environmental research, department of sustainability and environment, Heidelberg, Victoria
Greet J, Fedrigo M, Bennett A (2022) Deer impacts on the vegetation composition and structure of wet forests in the Yarra Ranges. The waterway ecosystem research group. Technical report 22.3. The University of Melbourne, Richmond, Victoria
Kunapo J, Walsh CJ, and Sammonds MJ (2020) The Melbourne water stream network. Version 1.1. melbourne waterway research-practice partnership report 19.4a. School of ecosystem and forest sciences, The University of Melbourne, Melbourne
Stewart SB, Fedrigo M, Roxburgh SH, Kasel S, and Nitschke CR (2020) Climate Victoria: precipitation (9 second, approx. 250 m). v2. CSIRO. Data collection. https://doi.org/10.25919/5e3be5193e301
Stewart SB, and Nitschke CR (2017b) Climate Victoria: maximum temperature (3DS-M; 9 second, approx. 250 m). v3. CSIRO. Data Collection. https://doi.org/10.25919/5e5d9033d8cc7
Stewart SB, and Nitschke CR (2018) Climate Victoria: minimum temperature (3DS-TM; 9 second, approx. 250 m). v3. CSIRO. Data collection. https://doi.org/10.25919/5e5dc18621a12
Acknowledgements
We acknowledge the Wurundjeri Woi-wurrung and Bunurong people as the Traditional Owners of the land on which our fieldwork was undertaken. We acknowledge the contribution of Lee Hazel to the initial stages of this project, and the field assistance of Danny White, Scott McKendrick, April Gloury, Elise King, Sarah Fischer, Paul Hanley, Claudia Nicklason and Bradley Jenner. We greatly appreciate the contribution of field-measured deer faecal pellet count data to this project by individuals and organisations including Parks Victoria, Arthur Rylah Institute for Environmental Research, DELWP, Melbourne Water, GHD Group and The University of Melbourne. Fieldwork was conducted under DELWP Research Permits No. 10009140 and No. 10009437. We thank three anonymous reviewers for comments that greatly improved this paper.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This research was funded by Melbourne Water through the Melbourne Water Research Practice Partnership with the Waterway Ecosystem Research Group (University of Melbourne). Field surveys were jointly funded by Melbourne Water and the Victorian Department of Environment, Land, Water and Planning (DELWP), with additional funding and logistical support provided by Parks Victoria. Financial support was also awarded to MF from the Albert Shimmins Fund at the University of Melbourne.
Author information
Authors and Affiliations
Contributions
MF, AB, SBS and JG conceived the study. AB, DMF and JG designed the field methods and collected the data; MF and SBS led the modelling and analyses. All authors contributed critically to drafting this manuscript and gave final approval for its publication.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fedrigo, M., Bennett, A., Stewart, S.B. et al. Modelling the spatial abundance of invasive deer and their impacts on vegetation at the landscape scale. Biol Invasions 26, 1901–1918 (2024). https://doi.org/10.1007/s10530-024-03282-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-024-03282-w