
Abstract
Forecasting habitat suitability and connectivity can be central to both controlling range expansion of invasive species and promoting native species conservation, especially under changing climate conditions. This study aimed to identify and prioritize areas in Spain to control the expansion of one of the most harmful invasive species in Europe, the American mink, while conserving its counterpart, the endangered European mink, under current and future conditions. We used ensemble habitat suitability and dynamic connectivity models to predict species ranges and movement routes considering likely climate change under three emission scenarios. Then, using habitat availability metrics, we prioritized areas for invasive mink control and native mink conservation and classified them into different management zones that reflected the overlap between species and threat from American to European minks. Results suggest that both species are likely to experience declines in habitat and connectivity under climate change scenarios with significantly larger declines by the end of the century for European minks (72 and 80% respectively) than for American minks (41 and 32%). Priority areas for management of both species varied over time and across emission scenarios, with a general shift in priority habitat towards the North-East of the study area. Our findings demonstrate how habitat suitability and dynamic connectivity approaches can guide long-term management strategies to control invasive species and conserve native species while accounting for likely landscape changes. The simultaneous study of both invasive and native species can support prioritized management action and inform management planning of the intensity, extent, and techniques of intervention depending on the overlap between species.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Climate change is projected to alter the threats posed by invasive species to native species conservation (Vilà et al. 2007; Hellmann et al. 2008; Polaina et al. 2021). Invasive species endanger native species through predation, competition for resources, and disease transmission (Genovesi et al. 2012). Because invasive species are frequently more generalist in their habitat selection and adapt more easily to changing landscape conditions than native specialist species, climate change has been found to exacerbate the detrimental effects posed by invasive species (Ravi et al. 2022). Specifically, shifts in climate have the potential to alter the introduction, establishment, distribution, and spread of invasive species (Polaina et al. 2021). These changes may also affect invasive species’ impact on native species, altering the distribution and degree of vulnerability of the native species and areas of overlap of both species (Hellmann et al. 2008). To respond to these potential shifts, both invasive species control measures and native species conservation may require dynamic approaches that can support responsive management actions across changing conditions to prevent the establishment, spread, and threat of invasive species and preserve native species over time.
The impacts of climatic changes on native and invasive species’ ability to thrive depends largely on the availability, distribution, suitability, and connectivity of their habitats. Habitat suitability and connectivity modeling are common approaches used to locate and quantify a species’ potential distribution and movements as a function of multiple biotic and abiotic environmental factors such as climate and land use or cover (Correa Ayram et al. 2015; Guisan et al. 2017). As these factors change over time (Mora et al. 2013; Song et al. 2018), so do habitat suitability and landscape connectivity (Saura et al. 2011; Rubio et al. 2012; Carvalho et al. 2021; Goicolea and Mateo-Sánchez 2022). Despite the widespread recognition that habitat suitability and associated connectivity are dynamic processes, connectivity has been more commonly modeled as a static, immutable feature (Zeller et al. 2020). However, several studies have found that static approaches to connectivity modeling fail to effectively quantify changes in landscape connectedness or identify priority areas for management measures under changing conditions (Wimberly 2006; Bishop-Taylor et al. 2018; Jennings et al. 2020; Goicolea and Mateo-Sánchez 2022). Recent advances in dynamic connectivity modeling approaches account for changes in landscape suitability and connectivity stability and can be used to identify persistent areas that are likely to support species expansion to potential future ranges (Keeley et al. 2018; Zeller et al. 2020; Goicolea and Mateo-Sánchez 2022). Some of these dynamic approaches generate predictions over multiple time periods and use future climate predictions to forecast potential changes in species expansion and adaptation to climate and related landscape changes (Beltrán et al. 2014; Bishop-Taylor et al. 2018). These dynamic models can be powerful tools to identify areas likely to persist important for focal species connectivity and thus guide relevant and sustainable conservation measures targeting long-term management (Jennings et al. 2020; Goicolea and Mateo-Sánchez 2022).
Despite the well-documented importance of maintaining or enhancing habitat integrity and connectivity in the face of habitat loss and fragmentation (Rudnick et al. 2012; Correa Ayram et al. 2015), there can also be negative consequences of landscape connectivity. Connectivity can facilitate the spread and expansion of unwanted disturbances such as catastrophic fires, pests, or invasive species, increasing the threats to species of conservation concern (Drake et al. 2017; de la Fuente et al. 2018). Previous studies have mainly focused on enhancing the distribution and movements of specialist, native, or threatened species yet rarely focus on limiting expansion of detrimental processes. However, concurrent modeling of species of conservation concern and the threats they face can be used to better identify management and conservation actions to mitigate threats and protect at-risk species. This concurrent modeling has the potential to be particularly beneficial in managing native species and non-native invasive competitors. To delineate priority conservation action areas (Wilson et al. 2009) where management can promote the persistence of native species and limit the impacts and expansion of invasive competitors, dynamic connectivity models can be combined with habitat availability metrics (Saura and Pascual-Hortal 2007; Saura and Rubio 2010). Habitat availability metrics are particularly useful for informing conservation and management efforts because they are flexible and can assess adequacy, permanence, and irreplaceability of habitat patches and linkages, three important criteria in conservation prioritization. Adequacy denotes how suitable and connected areas are to favor species populations’ survival and growth. These characteristics can arise from habitat suitability and connectivity models and depend on the environmental conditions and location of the area. The second criterion is the continuity or permanence of the adequacy of these conservation areas over time. In this sense, dynamic connectivity approaches are essential to identify areas likely to support persistently suitable and connected habitat even under projected changes. Lastly, habitat availability metrics can also be used to consider the irreplaceability of areas, i.e., whether alternative landscape elements could contribute similarly to the conservation goal. Therefore, these habitat availability metrics can be useful to guide conservation and management efforts to enhance habitat suitability and connectivity for native species and reduce that of invasive species under changing conditions.
Here, we identified and compared current and future priority areas to conserve a critically endangered native species, the European mink (Mustela lutreola), and control the expansion of its invasive competitor, the American mink (Neovison vison), in Spain. Specifically, we studied the current and potential future distribution and functional connectivity of both species with habitat suitability and dynamic connectivity models accounting for the adequacy, permanence, and irreplaceability of the areas. To address the uncertainty associated with future predictions, we employed multiple statistical techniques and evaluated suitability and connectivity over four time under three emissions scenarios. Additionally, we integrated habitat availability and connectivity results for both species to identify the most important areas to control American minks and to preserve European minks over this century.
Data and methods
Species and study area
The study area is Spain (excluding insular areas) in southwest Europe (Fig. 1). It occupies 492,175 km2 in area, and is composed of the Mediterranean forests, woodlands and scrubs, and temperate broadleaf and mixed forests biomes (Olson et al. 2001). Spain is expected to experience large climate and land-use changes (Loarie et al. 2009; Song et al. 2018) and these changes may affect population dynamics of both native and invasive species. In fact, Spain is considered one of the regions with the highest concentration of vulnerable terrestrial species to climate change (Pacifici et al. 2015).

The study area (Spain) and estimated distribution of the invasive (American mink) and the endangered native (European mink) species as of 2012 according to the Spanish inventory of terrestrial species (MITECO 2012)
The European mink (Mustela lutreola) has been recognized as one of the most endangered mammals in the world (Maran et al. 2016). In the mid 1800’s, European mink were found throughout most of continental Europe (Youngman 1990). However, in the last years, over 85% of their range has disappeared, and is currently restricted to small, isolated, and decreasing populations in eastern Europe, southern France, and northern Spain. The causes of this decline are a combination of invasive species competition and disease transmission, habitat loss and fragmentation, water pollution, and overhunting (Maran and Henttonen 1995; Mañas et al. 2001; Zuberogoitia et al. 2013). European minks in Spain (Fig. 1) occupy less than 2300 km of watercourses and have a population size of about 500 individuals with low genetic variability (Michaux et al. 2005; Palazón and Melero 2014; Cabria et al. 2015).
Another mink species, the American mink (Neovison vison) is one of the most harmful exotic and invasive species in Europe (Genovesi et al. 2012). Both mink species are small, semi-aquatic, and carnivorous mustelids with considerable niche overlap in food consumption (Sidorovich et al. 2010; Zuberogoitia et al. 2013) and preferred habitat, i.e. habitat adjacent to water with high, dense vegetation cover (Fuller et al. 2016; Maran et al. 2016). However, American minks are more adaptable to poor habitat quality, in addition to being bigger, more aggressive, and more successful breeders than European minks (Palazón and Melero 2014; Zuberogoitia et al. 2014). American minks were introduced in Europe from North America in the first half of the twentieth century for fur farming. Escaped or released individuals formed wild populations, and rapidly spread to be widely distributed around Europe. In Spain, American minks have established feral widespread populations across 12,530 km of rivers with a population of over 30,000 individuals (Fig. 1., MAGRAMA 2014). The establishment and spread of the American mink is considered one of the main causes of decline of European mink (Sidorovich et al. 2010; Maran et al. 2016), with similar negative impacts on other native species of conservation concern such as European polecats (Mustela putorius), stoats (Mustela erminea), and sea birds (MacDonald and Harrington 2003; Ahola et al. 2006; Banks et al. 2008; Schüttler et al. 2009; Melero et al. 2012; García-díaz et al. 2013). American mink has also been linked to economic damage (Pimentel et al. 2005) and transmission of influenza and COVID-19 (ECDC 2020; Fenollar et al. 2021; Harrington et al. 2021; Sun et al. 2021).
Species occurrences and environmental variables data
Data for both mink species were obtained from the yearly updated Spanish inventory of terrestrial species (MITECO 2012). We gathered 1396 American and 197 European mink occurrences at 10-km resolution (Fig. 1). The bioclimatic variables (Table 1) were obtained from Climatologies at High resolution for the Earth’s Surface Areas (CHELSA) data (Karger et al. 2017) with 30 arcsec resolution but resampled to species occurrences resolution (10 km). We downloaded these bioclimatic variables for the historic period of 1985–2010 and three future time periods 2011–2040, 2041–2070, 2071–2100 (hereafter referred to as t1, t2, t3, and t4). The variables of the future periods were derived from the GFDL-ESM4 (National Oceanic and Atmospheric Administration, Geophysical Fluid Dynamics Laboratory, Princeton, NJ 08540, USA) Global Climatic Model under three different shared socio-economic and emissions pathways (SSP): 126, 375 and 585 (hereafter referred to as e1, e2, and e3) corresponding to the Representative Concentration Pathways (RCP) 2.6 (an optimistic and sustainable scenario), 7 (medium–high emissions scenario), and 8.5 (high fossil-fuel development) (Meinshausen et al. 2020). In total, we developed models for 10 scenarios (one historic and nine future).
Although climate variables are likely to influence species habitat selection at a coarse scale (Wiens 1989; Pearson et al. 2002; Thuiller et al. 2004), prior studies have shown that both mink species respond to finer scale habitat structure (Yamaguchi et al. 2003; Harrington and Macdonald 2008; Crego et al. 2018). Therefore, we refined our models with land use and cover variables previously identified as affecting mink habitat suitability at a finer resolution (Yamaguchi et al. 2003; Fuller et al. 2016). Specifically, we incorporated 2018 CORINE land cover maps (European Union, Copernicus Land Monitoring Service 2018) at 100-m resolution complemented with roads (ESRI and Garmin International 2021), rivers (Ministerio para la Transición Ecológica y el Reto Demográfico 2018), and tree cover density (European Environment Agency 2018). We lacked fine-scaled future forecasts for landscape variables and assumed static land use and cover over time (Costanza and Terando 2019) even though adding accurate information on land use and human pressure is of utmost importance in species’ habitat selection (Parks et al. 2020).
Habitat suitability modeling
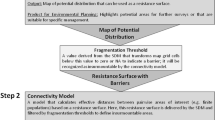
To assess connectivity dynamics over time, we started by modeling snapshots of habitat suitability at different time periods and under our different emission scenarios. Habitat suitability was calculated as the product of climate and landscape suitability measurements (Fig. 2). We calculated climate suitability by modeling species distribution based on the relationship between species occurrences and bioclimatic variables. Landscape suitability was calculated combining land use, tree cover density, and distance to water information. We determined the relationships to these landscape variables based on published literature (Yamaguchi et al. 2003; Zschille et al. 2012; Fuller et al. 2016; Halbrook and Petach 2018) as the resolution of the species occurrences (10 km) was considered too coarse to be relevant in species land use and cover selection modeling.

Diagram showing the workflow of the habitat suitability maps by combining A climate and B landscape suitability for each species. Climate suitability (A) was calculated by assembling several individual suitability models: Generalized Linear Model (GLM), generalized additive models (GAM), generalized boosting models (GBM), maximum entropy (MAXENT), multivariate adaptive regression splines (MARS), and random forests (RF). Landscape suitability (B) was calculated by combining the suitability associated with land use, distance to water, and tree cover density. Climate suitability and the final habitat suitability maps had ten different scenarios: one historic and three future scenarios, considering three socioeconomic and emissions scenarios (Shared Socioeconomic Pathways SSP 126, SSP 370, and SSP 585)
We calculated climatic suitability for each species following an ensemble species distribution modeling approach (Araújo and New 2006) considering relevant bioclimatic variables (Table 1 and Appendix S1) with the biomod2 R package (Thuiller et al. 2021). Ensemble models combine a set of individual models to account for algorithm biases (Grenouillet et al. 2011) and limit the influence of uncertainties associated with a single statistical technique for future forecasts (Araújo and New 2006; Beaumont et al. 2016). In this study, we combined six different models representing a range of statistical techniques: generalized linear models (GLM) (McCullagh and Nelder 1989), generalized additive models (GAM) (Hastie and Tibshirani 1986), generalized boosting models (GBM) (Friedman 2001), maximum entropy (MAXENT) (Phillips et al. 2006), multivariate adaptive regression splines (MARS) (Friedman 1991), and random forests (RF) (Breiman 2001). Background points were randomly selected across the study area 10-km grid at a 3:1 ratio with occurrence points (Barbet-Massin et al. 2012). We assumed there was no bias in the species occurrence data, as they were reported at a coarse, grid-based scale across all of Spain. Models were calibrated with the historic bioclimatic variables. We assessed the performance of the individual models with tenfold cross-validation (Pearce 2000) using the area under the curve (AUC) performance metric (Fan; et al. 2006). To assemble the final ensemble model for each species, we retained cross-validation runs of individual models with an AUC greater than 0.75 and averaged them in proportion to their AUC (weighted average). We also evaluated the ensemble model performance using the same cross-validation procedure, and the AUC, TSS, and Kappa evaluation statistics. Lastly, we projected the climate suitability model with clamping extrapolation to every time period (t1, t2, t3, t4) and SSP scenario (e1, e2, and e3) producing ten different climate suitability surfaces. To measure the risk of extrapolation of these projections (Owens et al. 2013), we developed a mobility-oriented parity analysis (MOP) to compare the calibration of climatic conditions (the variables at the historic scenario) with the transferred climate conditions (climate variables at future scenarios) with the kuenm R package (Cobos et al. 2019).
To calculate landscape suitability, we considered three factors within each 100 m pixel across our study area: type of land use, tree cover density, and distance to rivers and water bodies (Fuller et al. 2016). We assigned land use suitability according to the degree of human pressure for each land cover type (Table S2 in supplementary material): artificial land uses were considered unsuitable (0), agricultural uses moderately suitable (0.75), and natural areas assigned maximum suitability (1) (Fuller et al. 2016). For the tree cover component of landscape suitability, we assigned suitability values according to quartiles of tree cover density (Yamaguchi et al. 2003): a coverage of 100–75% was associated with the highest suitability (1), 75–50% with 0.75 suitability, 50–25% with 0.5 suitability, and 0–25% as unsuitable (0). For the final component of landscape suitability, we transformed distance to water into a suitability percentage with a negative exponential function assigning pixels adjacent to rivers or water bodies with a maximum suitability value (1) and with values decreasing exponentially for pixels farther from water (Zschille et al. 2012; Fuller et al. 2016; Halbrook and Petach 2018). To fit the exponential function, we assigned a suitability value of 0.05 to 900 m, the maximum distance of American minks to water sources reported in the literature (Crego et al. 2018).
To calculate the final habitat suitability value of each cell, we multiplied the climate suitability and the three components of landscape suitability. We obtained ten different habitat suitability surfaces at 100 m resolution, one for each period and emissions scenario considered. We also transformed the habitat suitability surfaces to binary surfaces of habitat or non-habitat using the true skill statistic (TSS) threshold (Somodi et al. 2017) of the climate suitability models. We used this binary surface to delineate habitat patches (Fig. 3) in ArcGIS software (Esri, Redlands, California, USA) by aggregating suitable habitat cells within 1 km and assembling polygons of habitat with a minimum area of 70 ha. This 70 ha threshold is the estimated minimum area required for populations’ long-term survival, reproduction, and proliferation, and was calculated as the longitudinal home range (7 km) multiplied by the pixel width (100 m). We verified that previous studies obtained similar home ranges (Yamaguchi and MacDonald 2003; Zabala et al. 2007; Fournier et al. 2008; Harrington and Macdonald 2008; Peters et al. 2009; Zschille et al. 2012; Palomares et al. 2017; Halbrook and Petach 2018). Lastly, we used the area of these habitat patches to estimate species abundance assuming a proportional relationship to patch size (Drake et al. 2021). Aside from the ten sets of habitat patches for each species (one per period and scenario), we calculated the overall potential habitat by aggregating the habitat patches of all periods and scenarios to represent the total area that could be colonized at any time by each species.

Connectivity analysis workflow. Asterisks denote 10 different scenarios: one historic and three future scenarios, each of the future ones with three emissions scenarios
Habitat connectivity modeling
We used the delineated habitat patches and resistance surfaces derived from the habitat suitability surfaces as the basis for least-cost analyses and connectivity metrics (Fig. 3). We calculated the resistance surface, which represents the difficulty for the species to move across each cell of the landscape matrix (Spear et al. 2010; Zeller et al. 2012) for each species, period, and SSP scenario using the negative exponential transformation described in Keeley et al. (2016). We considered that both species would likely be more tolerant of moderately suitable areas during dispersal than when selecting habitat; therefore, we set an intermediate value for the parameter c (c = 2) to transform from habitat suitability to resistance to movement surfaces as the inverse exponential of the habitat suitability surfaces (Mateo-Sánchez et al. 2015; Keeley et al. 2016; Jennings et al. 2020) with the formula:
We then calculated potential links or corridors (least-cost paths) (Adriaensen et al. 2003), as well as their Euclidean and effective (cost weighted) distances between patch edges and across the resistance surface with Linkage Mapper software (McRae and Kavanagh 2011). We set a cutoff maximum bounding circle buffer distance of 50 km (Harrington et al. 2014; Oliver et al. 2016) around every habitat patch to increase computability. We also obtained the ratio of the effective to Euclidean distances of all links and calculated the mean of all the ratios in the historic scenario as an estimate to transform from Euclidean to effective distance.
To evaluate connectivity over time for both species, we calculated two connectivity metrics: the overall connectivity of the ecological network and the importance (adequacy and irreplaceability) of each patch and link to maintain it. To calculate these metrics, we used graph-based habitat availability metrics (Appendix S3 in supplementary material, Pascual-Hortal and Saura 2006; Saura and Pascual-Hortal 2007; Saura and Rubio 2010) with the command line version of Conefor software (Saura and Torné 2009, www.conefor.org). These connectivity metrics were calculated separately for each species, time period, and SSP scenario. They account for both the habitat available within a patch (intrapatch connectivity) and the habitat available through the connections with other patches (interpatch connectivity).
Overall landscape connectivity
To calculate overall landscape connectivity, we used the Equivalent Connected Area index (ECA), which is defined as the area of a single habitat patch that would provide the same level of connectivity as the actual pattern of habitat patches in the whole landscape (Saura et al. 2011). This metric is a function of the dispersal capacity of the focal species (given in this case by its maximum dispersal distance), the landscape configuration (represented by the effective distance between patches), and patch attributes (in this case patch area) and is directly comparable among scenarios. We used a maximum dispersal distance of 40 km (Mitchell 1961; Harrington et al. 2014; Oliver et al. 2016) converted into an effective distance (i.e., multiplied by the mean effective to Euclidean ratio obtained previously). We matched this distance to a probability of 0.05 to fit a negative exponential function that represented the probability of connection between each pair of patches in relation to the effective distance between them.
Priority conservation and control areas
To assess patch importance, we calculated the change in the overall connectivity when systematically removing each patch (see appendix S3 in the supplementary material and Saura and Pascual-Hortal 2007). The most important (adequate and irreplaceable) patches are those whose removal would imply a bigger loss of overall connectivity. We calculated patch importance for each of the ten scenarios, but also calculated a generalized patch importance value by min–max normalizing and summing the importance across all the scenarios for each species to find the areas that remained stable in their importance despite global changes.
To most effectively manage areas of overlap between competing native and invasive species, it is important to evaluate changes in habitat suitability and connectivity relative to one another. To do that, we integrated the results for both species and classified potential habitat into three different management zones: the overlap zone, the expansion zone, and the low-risk zone. The overlap zone corresponds to the areas where overall potential habitat of both species co-occur and therefore management of both species should be considered as top priority. The expansion zone represents areas of overall potential American mink habitat that do not directly overlap with potential habitat of European minks, but is within a buffer of 40 km, the maximum dispersal distance. The expansion zone is where controlling the spread of American minks is most important. Finally, we defined the low-risk zone as the area where American mink overall potential habitat exceeds the maximum dispersal distance (> 40 km) from European mink habitat, or European mink overall potential habitat does not overlap with that of American minks. Within these three management zones, we ranked the importance of the areas based on generalized patch importance. Lastly, we determined the percentage of overlap of each management zone within the network for existing protected areas (UNEP-WCMC and IUCN 2022).
Results
Habitat suitability
Every individual and ensemble climate suitability model achieved an AUC greater than 0.85 (Table S4). In the historic period (t1), the amount of suitable area (total patch area) was 19,418 km2 for the American mink and 6171 km2 for the European mink. Suitable habitat was predicted to decrease for both species across all future time periods and emission scenarios (Fig. 4 and Table S6). However, the pattern of decline was different for the two species. The suitable area for the American mink was predicted to decrease more drastically in the two first future periods (t2 and t3) while the biggest decrease in habitat suitability for the European mink was predicted at the end of the century (t4). Overall, results showed a greater proportional decrease in habitat amount for the European minks; the mean decrease in suitable area across the three emission scenarios by t4 was 41% for American minks and 72% for European minks. For both species, the number of habitat patches generally decreased over time and with the increase of expected emissions. The size of these habitat patches also declined with the time and emissions for the European mink while the size of American mink patches was more variable (Table S6 of supplementary material). The overlap in suitable habitat remained constant across all periods and emission scenarios (Fig. S7). The overall potential habitat aggregated across all time periods and emission scenarios was 2,756,673 ha and 828,606 ha (141% and 134% of the amount of suitable area in the historic period) for American and European mink respectively. The risk of extrapolation increased with the time period and emissions scenario (Fig. S5).

Total (A.1 and B.1) and change (A.2 and B.2) in suitable habitat (A) and overall connectivity (B) for each period t1, t2, t3, and t4 (1985–2010, 2011–2040, 2041–2070, 2071–2100) and species (American and European mink). Overall connectivity is measured with the metric Equivalent Connected Area (ECA). The three future periods (t2, t3, and t4) have three emissions scenarios. In A.1 and B.1, central lines show the mean across the three emission scenarios and the shaded areas represent the range between the minimum and the maximum habitat area (A) and ECA (B) according to the three scenarios. A.2 and B.2 show the proportion of suitable area and ECA of future periods (t2, t3, and t4) in relation to the historic period (t1)
Connectivity analyses
The effective to Euclidean distance ratio was 78 for the American and 71.6 for the European mink, and the maximum effective dispersal distances were 2864 km and 3120 km for each species respectively. In mapping our connectivity results, we observed that links connecting habitat patches for one species often crossed habitat patches or links of the other species (Fig. S8). In fact, in the historic period t1 26.3% of European mink links passed through American mink habitat patches or links. The predicted overall habitat connectivity (ECA) decreased for both species over time (Fig. 4). American mink’s greatest loss of habitat connectivity was from time t2 to time t3 while for the European mink was from time t3 to time t4. By the end of the century, the impacts of climate change are expected to be worse for the European mink: connectivity decreased on average across the three emission scenarios 32% for American minks and 80% for European minks by t4. Also, increasing emissions were generally associated with a greater decline in habitat connectivity for both species.
Prioritization of action areas
We identified clear separation in the priority areas to maintain the overall species connectivity (i.e., areas with the greatest generalized patch importance) for the two species, with the northwest of Spain being most important for American minks and north central for European minks (Fig. 6). These priority areas for both species are predicted to shrink towards the north and east over time (Fig. S9) and with increasing emissions (Fig. S10).
Figure 6 shows the three zones of conservation and the order of priority within each zone to control American minks and to preserve European minks. Of the overall potential habitat for American mink, the overlap zone represented 18%, the expansion zone 25%, and the low threat zone 57%. 17.4% of this overall potential habitat is inside Protected areas (19% in the overlap zone, 27% in the expansion zone, and 13% in low threat zone). For the European mink, 60% of their overall potential habitat belongs to the overlap zone and 40% to the low exposure zone. 19.5% of the potential European mink habitat is protected (19% in the case of the overlap zone and 20% in the low exposure zone).
Discussion
Climate change, loss of connectivity through habitat fragmentation, and establishment and expansion of invasive species are interrelated phenomena that pose a threat to biodiversity and challenge conservation efforts. Effectively planning for and responding to these threats requires consideration of the complex and dynamic nature of the interactions among them. For example, enhancing connectivity is one of the most recommended strategies to support species' adaptation to climate change (Welch 2005; Heller and Zavaleta 2009; Keeley et al. 2018), but it also has the potential to facilitate the spread of invasive species. Additionally, climate change may affect both the threat posed by invasive species on ecosystems and native species (Hellmann et al. 2008; MITECO 2020) and species’ habitat availability and connectivity (Keeley et al. 2018; Zeller et al. 2020; Goicolea and Mateo-Sánchez 2022). In this study, we presented a spatially explicit and dynamic approach to integrate all three phenomena to guide long-term and climate-wise measures to preserve a native species and control its invasive competitor in a changing landscape. The combination of the habitat suitability and connectivity results from multiple time periods and scenarios allowed us to identify consistently important areas likely to enhance current and future habitat connectivity between contemporary patches, but also areas and routes that promote species’ expansion to new areas (Goicolea and Mateo-Sánchez 2022). By focusing on one of the most worrying invasive species in Europe, the American mink, and one endangered native species affected by its invasion, the European mink, our findings demonstrated the importance of dynamic, integrated analyses. This approach also serves as a transferable framework to support to climate-wise conservation planning for other invasive species and their threatened native counterparts.
Decreasing habitat suitability and connectivity for both species under climate projections
Our results confirmed that habitat availability and connectivity for both native and invasive species are dynamic phenomena likely to be affected by projected climatic changes over time (Figs. 4 and 5). By 2071–2100 European mink habitat is expected to experience more substantial negative impacts under changing climate conditions, probably due to their higher degree of habitat selection specialization (Pacifici et al. 2015). American minks were also projected to experience habitat reduction, even higher than that of European minks between 2041 and 2070, which is notable given their ecological plasticity (Palazón and Melero 2014; Zuberogoitia et al. 2014) and the observed rapid range expansion in recent years (Põdra and Gómez 2018). This result suggests that even generalist species currently undergoing range expansion might be substantially affected by global changes. In this sense, climate change is likely to lead to a gradual decline in the amount of suitable area and potentially the distribution of American minks in Spain. Other studies have also predicted a contraction of the American mink range in Europe in future decades (Polaina et al. 2021). The period with the highest predicted loss of habitat and connectivity for the American mink (2041–2070) may offer an opportunity to intensify the control measures during a time when populations contract and control measures might be more effective. Additionally, minimizing the impact of the invasive species during this time could reduce other stressors and help European minks adapt to the greater loss of habitat availability and connectivity predicted by the end of the century (2071–2100).

Generalized importance of habitat patches for A American mink and B European mink. The generalized patch importance was calculated as the sum of min–max normalized patch importance (see appendix S3 in the supplementary material and Saura and Pascual-Hortal 2007) across all the scenarios
Addressing future uncertainties
Climate suitability and connectivity models projected to future climate conditions can be associated with several sources of uncertainty (Heikkinen et al. 2006; Pacifici et al. 2015; Keeley et al. 2021). First, these models produce very different prediction and performance values depending on the statistical method used (Grenouillet et al. 2011; Pacifici et al. 2015; Beaumont et al. 2016). Given this variability, we applied multiple statistical methods and assembled their forecasts to reduce the uncertainty associated with any single modeling approach (Araújo and New 2006). Another common source of uncertainty in ecological models is the existence of future environmental conditions outside the training or calibration range (Zurell et al. 2012). Model projections to such novel future conditions can lead to erroneous extrapolations. Making cross-validation model evaluations is a recommended strategy to test how well the model performs when extrapolating outside the training range (Pearce and Ferrier 2000; Guisan et al. 2017). It is also recommended to check for the existence of novel environmental conditions to identify the areas where predictions could be spurious (Zurell et al. 2012; Owens et al. 2013). In this study we identified these areas with an MOP analysis (Fig. S5 in supplementary material).
Habitat and connectivity models projected to future scenarios are also subject to climatic uncertainties as different general circulation models, time periods, and emission scenarios forecast fluctuating climate conditions (Fernández et al. 2017). In fact, we obtained varying results depending on the time and emission scenario for both species (Figs. 4 and 5). To address this source of uncertainty, we separately considered the different periods and emission scenarios to obtain a range of potential outcomes that could be used to inform management strategies. The climatic uncertainty was also accounted for when prioritizing conservation areas. We found that the key areas to preserve the native species and control the invasive one did not remain static over time (Fig. S9) and the emissions scenario considered (Fig. S10). We found a trend of habitat reduction towards the east of the study area for both species over time and with higher emissions (Fig. S10). The similarities in the patterns we observed across time and emission scenarios suggest that higher greenhouse gas emissions may lead to an acceleration of climatic changes and associated effects. The independent study of habitat suitability at multiple periods let us identify both emerging and disappearing habitat and corridors over time that may inform improved management strategies that account for habitat and connectivity persistence. In the case of the invasive species, the early detection of novel habitats and corridors connecting them may guide more efficient measures that prevent invasive species’ spread and establishment in advance instead of controlling and eradicating them from already invaded areas. It can also be useful to identify ephemerally suitable areas for the invasive or the native species to avoid focusing control and conservation resources and efforts on only temporarily suitable areas.
While the individual results from each time period and scenario informed our understanding of habitat suitability and connectivity dynamics for each species, the integration of these results into a generalized adequacy and irreplaceability value allowed us to account for the likelihood of permanence of these parameters over time for both species. Those areas that appeared to be highly adequate (suitable and connected) and irreplaceable in multiple periods and scenarios are more likely to keep being important for the species in the long-term. Additionally, priority areas that enhance connectivity at multiple time periods may also promote connectivity of species to their future suitable ranges which could inform management of expansion routes of both invasive and native species (Goicolea and Mateo-Sánchez 2022).
Integrating native and invasive species modeling to prioritize management actions
Since the invasive and the native species in our study have similar habitat requirements (Zuberogoitia et al. 2013, 2014; Zschille et al. 2014; Fuller et al. 2016; Halbrook and Petach 2018), their distribution may overlap and they may be affected similarly by the upcoming global changes and by management measures. The results from this and similarly designed studies can be used in planning for control of invasive species to avoid negatively affecting native species populations. Equally, these results can guide preservation measures that avoid promoting the expansion and improvement of invasive species populations. One main feature of this study is the prioritization of conservation and control areas considering the invasive and the threatened species together, to avoid misplacing resources that would have unintentional results on the other species. Despite the different rates of habitat loss for the two species and the past increasing overlap distributions between the two species found in previous studies (Põdra and Gómez 2018), here we found nearly constant overlapping habitat patches (nearly one-third) between the two species over time (Fig. S7), consistent with previously observed degrees of overlap (Põdra and Gómez 2018). However, when we consider the overall potential habitat aggregated across all time periods and emission scenarios, a much higher amount of habitat for European mink, nearly 60%, overlaps with that of American minks. Therefore, some areas may be suitable for both species in different periods.
We classified different management zones to control American minks and to preserve the European mink depending on the threat posed to the native species (Fig. 6). These categories divide the study area in localized areas that could be subjected to different timing, intensities of intervention, and types of measures to manage both the invasive and the native species (MAGRAMA 2014). The overlap zone comprises areas with likely direct competition between both species that may lead to a displacement or population decline of the native species. Managers should prioritize locations and management actions with higher intensities in these overlap zone to avoid further deleterious consequences for European minks. Managers can also target more holistic approaches in overlapping zones involving both measures to preserve and enhance European mink populations and to remove American minks (MacDonald and Harrington 2003). Measures to preserve the European mink in these areas will likely fail to protect the species if they are not combined with measures to reduce the threat of American minks. Additionally, control measures such as live-trapping and selective control procedures could be employed more judiciously and carried out by trained and experienced staff in the overlap zone due to the risk of harming European minks (MacDonald and Harrington 2003; Reynolds et al. 2013). These intensive and selective measures are usually expensive and difficult to implement and therefore are more likely to be successful when planned for localized and manageable areas such as this overlap management zone. More practical measures at broad scales could be used on the expansion, and low-risk zones where there is no threat of overlap between species. Control measures in the expansion and low-risk zones could include removal traps, castration measures, the use of dogs (Fuller et al. 2016), scent repellents (Baker and Macdonald 1999), or fences to avoid their expansion or access to specific areas (e.g., farms). However, careful consideration of the potential effects of management actions on other mustelid or native species remains a critical step in evaluating tools for controlling invasive species. The overall potential habitat of European minks inside the low-risk zone could undergo conservation measures that directly benefit European minks, such as habitat restoration, and recovery and artificial reintroduction of the species (Skorupski 2020). Other measures however should be applied throughout the study area to avoid further increasing the competition and disease transmission from American to European minks, such as the prevention of American mink escapes (MAGRAMA 2014) and eradication of diseases in fur farms (Mañas et al. 2001).

Importance of areas to A control American minks and B conserve European mink in terms of connectivity separated in different management areas: overlap zone (potential habitat for both species), expansion zone (potential American mink habitat that within a buffer of 40 km of potential European mink habitat), and low risk zone (either potential American mink habitat farther away than 40 km from European mink habitat, or potential European mink habitat that does not overlap with that of American mink)
Our categorization of the priority areas aligns with the goals of the Spanish strategy to control American minks (MAGRAMA 2014) that differentiates two intensity levels and types of management depending on the distance to populations of European minks: first, eliminating the current American mink populations from the overlapping and peripheral areas of European mink distribution; and second, reducing their populations in the other areas of the country to minimize their colonizing potential. However, the Spanish strategy only considers the current distribution of both species, while this study considers both current and future distributions and connectivity. With integration of these future projections, conservation and control efforts can incorporate more proactive strategies.
Considering the overlap between management zones and protected areas also gives valuable information for management of both species as conservation plans to counteract climate change and invasive species effects can be implemented more easily there. The protected areas that overlap with American mink overall potential habitat can aid in control efforts by developing special programs to prevent species arrival, monitor their presence, remove individuals, or lessen the impact of their populations depending on the state of their distribution. Lastly, to enhance conservation efforts, the priority areas to preserve European minks could be evaluated for inclusion in the net of protected areas to avoid their exposure to land-use changes or anthropogenic pressure that would further threaten the fragile status of the species. Although future projections of land use change are not currently available for Spain at a scale relevant to our analyses, if produced in the future, this type of information could be overlaid and evaluated in the context of the management zones to determine where land-use decisions could affect European mink conservation. Such landscape scale decisions are particularly important given that 80% of European minks’ potential habitat lies outside protected areas. The overall potential habitat and the generalized importance (Fig. 5) of the species should be considered in future protected area delineation to preserve highly adequate (suitable and connected), irreplaceable, and persistent areas for the European minks.
Conclusions
Climate change is likely to exacerbate the negative effects of invasive species while concurrently posing a threat to species of conservation concern. Habitat suitability and dynamic connectivity approaches can be useful tools to forecast species distributions and movements to anticipate management measures within the context of climate change and support adaptive planning that accounts for a changing landscape. Specifically, these tools can improve our understanding of how the landscape facilitates the functionality of ecological networks over time. In this study, climate change is expected to negatively affect American minks and European minks by 2100, with poorer outcomes for the native mink. However, results from the different emissions scenarios highlight the uncertainty surrounding habitat suitability projections. This uncertainty can be addressed by integrating multiple statistical techniques, periods, and emission scenarios to identify a range of potential outcomes for the species of interest and develop flexible and responsive management measures to adapt management strategies to new knowledge. Lastly, habitat suitability and connectivity models can guide both conservation and control measures, and integrating the results of invasive and threatened species to differentiate management zones can support the development of more effective management measures. Although our study was focused on the case of the American and European mink in Spain, our analytical framework can readily be applied to similar studies in different locales or with different focal species.
Data availability
The datasets generated and analysed during the current study are available from the corresponding author on reasonable request.
References
Adriaensen F, Chardon JP, De Blust G et al (2003) The application of “least-cost” modelling as a functional landscape model. Landsc Urban Plan 64:233–247. https://doi.org/10.1016/S0169-2046(02)00242-6
Ahola M, Nordström M, Banks PB et al (2006) Alien mink predation induces prolonged declines in archipelago amphibians. Proc R Soc B Biol Sci 273:1261–1265. https://doi.org/10.1098/rspb.2005.3455
Araújo MB, New M (2006) Ensemble forecasting of species distributions. Trends Ecol Evol 22:42–47. https://doi.org/10.1016/j.tree.2006.09.010
Baker SE, Macdonald DW (1999) Non-lethal predator control: exploring the options. In: Advances in vertebrate pest management. pp 251–256
Banks PB, Nordström M, Ahola M et al (2008) Impacts of alien mink predation on island vertebrate communities of the Baltic sea archipelago: review of a long-term experimental study. Boreal Environ Res 13:3–16
Barbet-Massin M, Jiguet F, Albert CH, Thuiller W (2012) Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol Evol 3:327–338. https://doi.org/10.1111/j.2041-210X.2011.00172.x
Beaumont LJ, Graham E, Duursma DE et al (2016) Which species distribution models are more (or less) likely to project broad-scale, climate-induced shifts in species ranges? Ecol Modell 342:135–146. https://doi.org/10.1016/j.ecolmodel.2016.10.004
Beltrán BJ, Franklin J, Syphard AD et al (2014) Effects of climate change and urban development on the distribution and conservation of vegetation in a Mediterranean type ecosystem. Int J Geogr Inf Sci 28:1561–1589. https://doi.org/10.1080/13658816.2013.846472
Bishop-Taylor R, Tulbure MG, Broich M (2018) Evaluating static and dynamic landscape connectivity modelling using a 25-year remote sensing time series. Landsc Ecol 33:625–640. https://doi.org/10.1007/s10980-018-0624-1
Breiman L (2001) Random forests. Mach Learn 45:5–32. https://doi.org/10.1007/9781441993267_5
Cabria MT, Gonzalez EG, Gomez-Moliner BJ et al (2015) Patterns of genetic variation in the endangered European mink (Mustela lutreola L., 1761) Phylogenetics and Phylogeography. BMC Evol Biol 15:1–15. https://doi.org/10.1186/s12862-015-0427-9
Carvalho JS, Graham B, Bocksberger G et al (2021) Predicting range shifts of African apes under global change scenarios. Divers Distrib 27:1663–1679. https://doi.org/10.1111/ddi.13358
Cobos ME, Townsend Peterson A, Barve N, Osorio-Olvera L (2019) Kuenm: an R package for detailed development of ecological niche models using Maxent. PeerJ 2019:1–15. https://doi.org/10.7717/peerj.6281
Correa Ayram CA, Mendoza ME, Etter A, Pérez Salicrup DR (2015) Habitat connectivity in biodiversity conservation: a review of recent studies and applications. Prog Phys Geogr 40:1–32. https://doi.org/10.1177/0309133315598713
Costanza JK, Terando AJ (2019) Landscape connectivity planning for adaptation to future climate and land-use change. Curr Landsc Ecol Reports 4:1–13. https://doi.org/10.1007/s40823-019-0035-2LANDSCAPE
Crego RD, Jiménez JE, Rozzi R (2018) Potential niche expansion of the American mink invading a remote island free of nativepredatory mammals. PLoS ONE 13:1–18. https://doi.org/10.1371/journal.pone.0194745
de la Fuente B, Saura S, Beck PSA (2018) Predicting the spread of an invasive tree pest: the pine wood nematode in Southern Europe. J Appl Ecol 55:2374–2385. https://doi.org/10.1111/1365-2664.13177
Drake JC, Griffis-Kyle KL, McIntyre NE (2017) Graph theory as an invasive species management tool: case study in the Sonoran Desert. Landsc Ecol 32:1739–1752. https://doi.org/10.1007/s10980-017-0539-2
Drake J, Lambin X, Sutherland C (2021) The value of considering demographic contributions to connectivity: a review. Ecography (Cop). https://doi.org/10.1111/ecog.05552
ECDC (2020) Detection of new SARS-CoV-2 variants related to mink
ESRI, Garmin International I (2021) World Roads. https://www.arcgis.com/home/item.html?id=83535020ce154bd5a498957c159e3a99. Accessed 23 Oct 2021
European Environment Agency (2018) High Resolution Layer: Tree Cover Density (TCD) 2018. https://land.copernicus.eu/pan-european/high-resolution-layers/forests/tree-cover-density/status-maps/tree-cover-density-2018?tab=metadata. Accessed 13 Oct 2021
Fan J, Upadhye S, Worster A (2006) Understanding receiver operating characteristic (ROC) curves. Can J Emerg Med 8:19–20
Fenollar F, Mediannikov O, Maurin M et al (2021) Mink, SARS-CoV-2, and the human-animal interface. Front Microbiol. https://doi.org/10.3389/fmicb.2021.663815
Fernández J, Casanueva A, Montávez JP et al (2017) Regional climate projections over Spain: atmosphere. Present climate evaluation. CLIVAR Exch No 73(2050):39–44
Fournier P, Maizeret C, Fournier-Chambrillon C et al (2008) Spatial behaviour of European mink Mustela lutreola and polecat Mustela putorius in southwestern France. Acta Theriol (warsz) 53:343–354. https://doi.org/10.1007/bf03195195
Friedman JH (1991) Multivariate adaptive regression splines. Ann Stat 19:1–67
Friedman JH (2001) Greedy function approximation: a gradient boosting machine. Ann Stat 29:1189–1232. https://doi.org/10.1017/CBO9781107415324.004
Fuller AK, Sutherland CS, Royle JA, Hare MP (2016) Estimating population density and connectivity of American mink using spatial capture-recapture. Ecol Appl 26:1125–1135. https://doi.org/10.1890/15-0315.1
García-díaz P, Arévalo V, Vicente R, Lizana M (2013) The impact of the American mink (Neovison vison) on native vertebrates in mountainous streams in Central Spain. Eur J Wildl Res 59:823–831. https://doi.org/10.1007/s10344-013-0736-5
Genovesi P, Carnevali L, Alonzi A, Scalera R (2012) Alien mammals in Europe: updated numbers and trends, and assessment of the effects on biodiversity. Integr Zool 7:247–253. https://doi.org/10.1111/j.1749-4877.2012.00309.x
Goicolea T, Mateo-Sánchez MC (2022) Static vs. dynamic connectivity: how landscape changes affect connectivity predictions in the Iberian Peninsula. Landsc Ecol. https://doi.org/10.1007/s10980-022-01445-5
Grenouillet G, Buisson L, Casajus N, Lek S (2011) Ensemble modelling of species distribution: the effects of geographical and environmental ranges. Ecography (cop) 34:9–17. https://doi.org/10.1111/j.1600-0587.2010.06152.x
Guisan A, Thuiller W, Zimmermann N (2017) Habitat suitability and distribution models: with applications in R. Cambridge University Press
Halbrook RS, Petach M (2018) Estimated mink home ranges using various home-range estimators. Wildl Soc Bull 42:656–666. https://doi.org/10.1002/wsb.924
Harrington LA, Macdonald DW (2008) Spatial and temporal relationships between invasive American mink and native European polecats in the southern United Kingdom. J Mammal 89:991–1000. https://doi.org/10.1644/07-MAMM-A-292.1
Harrington LA, Põdra M, Macdonald DW, Maran T (2014) Post-release movements of captive-born European mink Mustela lutreola. Endanger Spec Res 24:137–148. https://doi.org/10.3354/esr00590
Harrington LA, Díez-León M, Gómez A et al (2021) Wild American mink (Neovison vison) may pose a COVID-19 threat. Front Ecol Environ 19:266–267. https://doi.org/10.1002/fee.2344
Hastie T, Tibshirani R (1986) Generalized additive models. Stat Sci 1:297–318
Heikkinen RK, Luoto M, Araújo MB et al (2006) Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog Phys Geogr 30:751–777. https://doi.org/10.1177/0309133306071957
Heller NE, Zavaleta ES (2009) Biodiversity management in the face of climate change: a review of 22 years of recommendations. Biol Conserv 142:14–32. https://doi.org/10.1016/j.biocon.2008.10.006
Hellmann JJ, Byers JE, Bierwagen BG, Dukes JS (2008) Five potential consequences of climate change for invasive species. Conserv Biol 22:534–543. https://doi.org/10.1111/j.1523-1739.2008.00951.x
Jennings MK, Haeuser E, Foote D et al (2020) Planning for dynamic connectivity: operationalizing robust decision-making and prioritization across landscapes experiencing climate and land-use change. Land. https://doi.org/10.3390/LAND9100341
Karger DN, Conrad O, Böhner J et al (2017) Climatologies at high resolution for the earth’s land surface areas. Sci Data 4:1–20. https://doi.org/10.1038/sdata.2017.122
Keeley ATH, Beier P, Gagnon JW (2016) Estimating landscape resistance from habitat suitability: effects of data source and nonlinearities. Landsc Ecol 31:2151–2162. https://doi.org/10.1007/s10980-016-0387-5
Keeley ATH, Ackerly DD, Cameron DR et al (2018) New concepts, models, and assessments of climate-wise connectivity new concepts, models, and assessments of climate-wise connectivity. Environ Res Lett 13:1–18
Keeley ATH, Beier P, Jenness JS (2021) Connectivity metrics for conservation planning and monitoring. Biol Conserv. https://doi.org/10.1016/j.biocon.2021.109008
Loarie SR, Duffy PB, Hamilton H et al (2009) The velocity of climate change. Nature 462:1052–1055. https://doi.org/10.1038/nature08649
MacDonald DW, Harrington LA (2003) The American mink: the triumph and tragedy of adaptation out of context. New Zeal J Zool 30:421–441. https://doi.org/10.1080/03014223.2003.9518350
MAGRAMA (2014) Estrategia de gestión, control y erradicación del visón americano (Neovison vison) en España. In: Minist. Agric. Aliment. y Medio Ambient. https://www.miteco.gob.es/es/biodiversidad/publicaciones/pbl-fauna-flora-estrategias-eei-vison-americano.aspx. Accessed 13 Apr 2022
Mañas S, Ceña JC, Ruiz-Olmo J et al (2001) Aleutian mink disease parvovirus in wild riparian carnivores in Spain. J Wildl Dis 37:138–144. https://doi.org/10.7589/0090-3558-37.1.138
Maran T, Skumatov Valentinovich D, Gómez A, et al (2016) Mustela lutreola
Maran T, Henttonen H (1995) Why is the European mink (Mustela lutreola) disappearing? - A review of the process and hypotheses. Ann Zool Fennici 32:47–54
Mateo-Sánchez MC, Balkenhol N, Cushman S et al (2015) A comparative framework to infer landscape effects on population genetic structure: are habitat suitability models effective in explaining gene flow? Landsc Ecol 30:1405–1420. https://doi.org/10.1007/s10980-015-0194-4
McCullagh P, Nelder JA (1989) Generalized linear models. Chapman & Hall, London
McRae BH, Kavanagh DM (2011) Linkage mapper connectivity analysis software
Meinshausen M, Nicholls ZRJ, Lewis J et al (2020) The shared socio-economic pathway (SSP) greenhouse gas concentrations and their extensions to 2500. Geosci Model Dev 13:3571–3605. https://doi.org/10.5194/gmd-13-3571-2020
Melero Y, Plaza M, Santulli G et al (2012) Evaluating the effect of American mink, an alien invasive species, on the abundance of a native community: Is coexistence possible? Biodivers Conserv 21:1795–1809. https://doi.org/10.1007/s10531-012-0277-3
Michaux JR, Hardy OJ, Justy F et al (2005) Conservation genetics and population history of the threatened European mink Mustela lutreola, with an emphasis on the west European population. Mol Ecol 14:2373–2388. https://doi.org/10.1111/j.1365-294X.2005.02597.x
Ministerio para la Transición Ecológica y el Reto Demográfico (2018) Ríos completos clasificados según Pfafstetter modificado. https://www.miteco.gob.es/es/cartografia-y-sig/ide/descargas/agua/red-hidrografica.aspx
Mitchell JL (1961) Mink movements and populations on a montana river. J Wildl Manage 25:48–54
MITECO (2012) Riqueza de especies. Inventario Español de Especies Terrestres: Malla 10 x 10 km
MITECO (2020) Plan Nacional de Adaptación al Cambio Climático 2021–2030. Madrid
Mora C, Frazier AG, Longman RJ et al (2013) The projected timing of climate departure from recent variability. Nature 502:183–187. https://doi.org/10.1038/nature12540
Oliver MK, Piertney SB, Zalewski A et al (2016) The compensatory potential of increased immigration following intensive American mink population control is diluted by male-biased dispersal. Biol Invasions 18:3047–3061. https://doi.org/10.1007/s10530-016-1199-x
Olson DM, Dinerstein E, Wikramanayake ED et al (2001) Terrestrial ecoregions of the world: a new map of life on Earth. Bioscience 51:933–938. https://doi.org/10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2
Owens HL, Campbell LP, Dornak LL et al (2013) Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecol Modell 263:10–18. https://doi.org/10.1016/j.ecolmodel.2013.04.011
Pacifici M, Foden WB, Visconti P et al (2015) Assessing species vulnerability to climate change. Nat Clim Chang 5:215–225. https://doi.org/10.1038/nclimate2448
Palazón S, Melero Y (2014) Status, threats and management actions on the European mink Mustela lutreola (Linnaeus, 1761) in Spain: a review of the studies performed since 1992. Munibe Monogr Nat Ser 3:109–118. https://doi.org/10.21630/mmns.2014.3.09
Palomares F, López-Bao JV, Telletxea G et al (2017) Activity and home range in a recently widespread European mink population in Western Europe. Eur J Wildl Res. https://doi.org/10.1007/s10344-017-1135-0
Parks SA, Carroll C, Dobrowski SZ, Allred BW (2020) Human land uses reduce climate connectivity across North America. Glob Chang Biol 26:2944–2955. https://doi.org/10.1111/gcb.15009
Pascual-Hortal L, Saura S (2006) Comparison and development of new graph-based landscape connectivity indices: towards the priorization of habitat patches and corridors for conservation. Landsc Ecol 21:959–967. https://doi.org/10.1007/s10980-006-0013-z
Pearce J, Ferrier S (2000) Evaluating the predictive performance of habitat models developed using logistic regression. Ecol Modell 133:225–245. https://doi.org/10.1016/S0304-3800(00)00322-7
Pearson RG, Dawson TP, Berry PM, Harrison PA (2002) SPECIES: a spatial evaluation of climate impact on the envelope of species. Ecol Modell 154:289–300. https://doi.org/10.1016/S0304-3800(02)00056-X
Peters E, Brinkmann I, Krüger F et al (2009) Reintroduction of the European mink Mustela lutreola in Saarland, Germany. Preliminary data on the use of space and activity as revealed by radio-tracking and live-trapping. Endanger Species Res 10:305–320. https://doi.org/10.3354/esr00180
Phillips SB, Aneja VP, Kang D, Arya SP (2006) Modelling and analysis of the atmospheric nitrogen deposition in North Carolina. Int J Glob Environ Issues 6:231–252. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Pimentel D, Zuniga R, Morrison D (2005) Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol Econ 52:273–288. https://doi.org/10.1016/j.ecolecon.2004.10.002
Põdra M, Gómez A (2018) Rapid expansion of the American mink poses a serious threat to the European mink in Spain. Mammalia 82:580–588. https://doi.org/10.1515/mammalia-2017-0013
Polaina E, Soultan A, Part T, Recio MR (2021) The future of invasive terrestrial vertebrates in Europe under climate and land-use change. Environ Res Lett. https://doi.org/10.1088/1748-9326/abe95e
Ravi S, Law DJ, Caplan JS et al (2022) Biological invasions and climate change amplify each other’s effects on dryland degradation. Glob Chang Biol 28:285–295. https://doi.org/10.1111/gcb.15919
Reynolds JC, Richardson SM, Rodgers BJE, Rodgers ORK (2013) Effective control of non-native American mink by strategic trapping in a river catchment in mainland Britain. J Wildl Manage 77:545–554. https://doi.org/10.1002/jwmg.500
Rubio L, Rodriguez-Freire M, Mateo Sánchez MC et al (2012) Sustaining forest landscape connectivity under different land cover change scenarios. For Syst 21:223–235. https://doi.org/10.5424/fs/2012212-02568
Rudnick DA, Ryan SJ, Beier P, et al (2012) The role of landscape connectivity in planning and implementing conservation and restoration priorities. Issues Ecol Fall:1–20
Saura S, Pascual-Hortal L (2007) A new habitat availability index to integrate connectivity in landscape conservation planning: comparison with existing indices and application to a case study. Landsc Urban Plan 83:91–103. https://doi.org/10.1016/j.landurbplan.2007.03.005
Saura S, Rubio L (2010) A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography (cop) 33:523–537. https://doi.org/10.1111/j.1600-0587.2009.05760.x
Saura S, Torné J (2009) Conefor Sensinode 2.2: a software package for quantifying the importance of habitat patches for landscape connectivity. Environ Model Softw 24:135–139. https://doi.org/10.1016/j.envsoft.2008.05.005
Saura S, Estreguil C, Mouton C, Rodríguez-Freire M (2011) Network analysis to assess landscape connectivity trends: application to European forests (1990–2000). Ecol Indic 11:407–416. https://doi.org/10.1016/j.ecolind.2010.06.011
Schüttler E, Klenke R, McGehee S et al (2009) Vulnerability of ground-nesting waterbirds to predation by invasive American mink in the Cape Horn Biosphere Reserve, Chile. Biol Conserv 142:1450–1460. https://doi.org/10.1016/j.biocon.2009.02.013
Sidorovich VE, Polozov AG, Zalewski A (2010) Food niche variation of European and American mink during the American mink invasion in north-eastern Belarus. Biol Invasions 12:2207–2217. https://doi.org/10.1007/s10530-009-9631-0
Skorupski J (2020) Fifty years of research on european mink mustela lutreola l., 1761 genetics: Where are we now in studies on one of the most endangered mammals? Genes (basel) 11:1–27. https://doi.org/10.3390/genes11111332
Somodi I, Lepesi N, Botta-Dukát Z (2017) Prevalence dependence in model goodness measures with special emphasis on true skill statistics. Ecol Evol 7:863–872. https://doi.org/10.1002/ece3.2654
Song X, Hansen MC, Stehman SV et al (2018) Global land change 1982–2016. Nature 560:639–643. https://doi.org/10.1038/s41586-018-0411-9.Global
Spear SF, Balkenhol N, Fortin MJ et al (2010) Use of resistance surfaces for landscape genetic studies: considerations for parameterization and analysis. Mol Ecol 19:3576–3591. https://doi.org/10.1111/j.1365-294X.2010.04657.x
Sun H, Li F, Liu Q et al (2021) Mink is a highly susceptible host species to circulating human and avian influenza viruses. Emerg Microbes Infect 10:472–480. https://doi.org/10.1080/22221751.2021.1899058
Thuiller W, Araújo MB, Lavorel S (2004) Do we need land-cover data to model species distributions in Europe? J Biogeogr 31:353–361. https://doi.org/10.1046/j.0305-0270.2003.00991.x
Thuiller W, Georges D, Gueguen M, et al (2021) biomod2: ensemble platform for species distribution modeling
UNEP-WCMC, IUCN (2022) Protected planet: the world database on protected areas (WDPA) and world database on other effective area-based conservation measures (WD-OECM) [Online]. In: Cambridge, UK UNEP-WCMC IUCN. www.protectedplanet.net
European Union, Copernicus Land Monitoring Service 2018 EEA (EEA) CORINE Land Cover
Vilà M, Corbin JD, Dukes JS, et al (2007) Linking plant invasions to global environmental change chapter 8 linking plant invasions to global environmental change. In: Terrestrial ecosystems in a changing world. pp 93–102
Welch D (2005) What should protected areas managers do in the face of climate change. George Wright Forum 22:76–93
Wiens JA (1989) Spatial scaling in ecology published by: British ecological society stable. Funct Ecol 3:385–397
Wilson KA, Cabeza M, Klein CJ (2009) Fundamental concepts of spatial conservation prioritization. In: Spatial conservation prioritization: quantitative methods and computational tools. pp 16–27
Wimberly MC (2006) Species dynamics in disturbed landscapes: When does a shifting habitat mosaic enhance connectivity? Landsc Ecol 21:35–46. https://doi.org/10.1007/s10980-005-7757-8
Yamaguchi N, MacDonald DW (2003) The burden of co-occupancy: intraspecific resource competition and spacing patterns in American mink, Mustela vison. J Mammal 84:1341–1355
Yamaguchi N, Rushton S, Macdonald DW (2003) Habitat preferences of feral American mink in the upper thames. J Mammal 84:1356–1373. https://doi.org/10.1644/1545-1542(2003)084
Youngman PM (1990) Mustela lutreola. Am Soc Mammal Stable 1–3
Zabala J, Zuberogoitia I, Martínez-Climent JA (2007) Spacing pattern, intersexual competition and niche segregation in American mink. Ann Zool Fennici 44:249–258
Zeller KA, McGarigal K, Whiteley AR (2012) Estimating landscape resistance to movement: a review. Landsc Ecol 27:777–797. https://doi.org/10.1007/s10980-012-9737-0
Zeller KA, Lewsion R, Fletcher RJ et al (2020) Understanding the importance of dynamic landscape connectivity. Land 9:1–15. https://doi.org/10.1093/oso/9780198838388.003.0005
Zschille J, Stier N, Roth M, Berger U (2012) Dynamics in space use of American mink (Neovison vison) in a fishpond area in Northern Germany. Eur J Wildl Res 58:955–968. https://doi.org/10.1007/s10344-012-0638-y
Zschille J, Stier N, Roth M, Mayer R (2014) Feeding habits of invasive American mink (Neovison vison) in northern Germany-potential implications for fishery and waterfowl. Acta Theriol (warsz) 59:25–34. https://doi.org/10.1007/s13364-012-0126-5
Zuberogoitia I, Zalewska H, Zabala J, Zalewski A (2013) The impact of river fragmentation on the population persistence of native and alien mink: an ecological trap for the endangered European mink. Biodivers Conserv 22:169–186. https://doi.org/10.1007/s10531-012-0410-3
Zuberogoitia I, Manuel J, De AP (2014) Population trends and evolution of the knowledge of European Mustela lutreola (Linnaeus, 1761) and American mink Neovison vison (Schreber, 1777) in Bizkaia Mustela. Munibe Monogr Nat Ser 3:119–131
Zurell D, Elith J, Schröder B (2012) Predicting to new environments: tools for visualizing model behaviour and impacts on mapped distributions. Divers Distrib 18:628–634. https://doi.org/10.1111/j.1472-4642.2012.00887.x
Acknowledgements
We would like to thank the contribution of Nima Farchadi, Erica Mills, Greta Schmidt, and Eve Bohnett for their advice on the manuscript writing. We also gratefully acknowledge the Universidad Politécnica de Madrid for the funding and for providing computing resources on Magerit Supercomputer. We thank the valuable feedback from the two anonymous reviewers.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This study was supported by funding from the Universidad Politécnica de Madrid.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Data collection and analysis were performed by TG. The first draft of the manuscript was written by TG and all authors commented on previous versions of the manuscript. All authors read thoroughly and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Goicolea, T., Lewison, R.L., Mateo-Sánchez, M.C. et al. Dynamic connectivity analyses to inform management of the invasive American mink and its native competitor, the European mink. Biol Invasions 25, 3583–3601 (2023). https://doi.org/10.1007/s10530-023-03128-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-023-03128-x