
Abstract
We analysed the invasion history of two North American macrophytes (Elodea canadensis and E. nuttallii) in Italy, through an accurate census of all available herbarium and field records, dating between 1850 and 2019, and a rich literature collection describing the initial introduction and naturalisation phase that supports the results obtained by the occurrence records. Elodea canadensis arrived in Italy before 1866 and had two invasion phases, between the 1890s and 1920s and between the 1990s and 2000s; E. nuttallii, probably arrived in the 1970s, started invading in 2000 and the invasion is still ongoing. Botanical gardens and fish farming played a crucial role in dispersal and naturalisation of both species. The current invasion range of both species is centred in northern Italy, with scattered occurrences of E. canadensis in central and southern regions. River Po represents a dispersal barrier to the Mediterranean region and a strategic monitoring site to prevent the invasion in the peninsula. The study detects differences in the niches of the two species during the introduction and naturalisation phase and a habitat switch occurred after 1980 in E. canadensis and after 2000 in E. nuttallii, during their expansion phases. For E. canadensis the switch corresponds to the second invasion round. Further research can clarify whether the second invasion round is due to confusion of the recently introduced E. nuttallii with E. canadensis, to a cryptic introduction of a new genotype, to post-introduction evolution, or just to an increased scientific interest in biological invasions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Biological invasions are one of the great challenges that ecosystems are facing under a scenario of global climate change and increasing human pressure (Simberloff and Rejmánek 2012; Pyšek et al. 2020). This is especially true for freshwater ecosystems, which are among the most threatened of the Earth (Brundu 2015) and most affected by biological invasions (Pyšek et al. 2010; Bolpagni 2021). On a European scale, for example, out of 36 invasive plant species of European Union concern, 13 are hydro- or hygrophilous plants (EU Regulation no. 1143/2014; European Commission 2017, 2019). Aquatic invasions are associated with the almost total alteration of aquatic habitats and trophic conditions by human activities (Wärner et al. 2011; Brundu 2015): the most invaded areas are, in fact, the most economically developed and densely populated (Malavasi et al. 2018; Boscutti et al. 2022). Human pressure and disturbance provide windows of opportunity for which alien species are extremely competitive (Pyšek and Prach 1994; Pyšek et al. 1998; Brundu et al. 2012; Bolpagni 2021). Since 1945, the number and abundance of aquatic alien species have increased because of increased trade (mostly as ornamental and aquarium species, but also for scientific research and phytoremediation—Kay and Hoyle 2001; Brundu 2015), water eutrophication and global warming (Hussner 2009; Bolpagni et al. 2015; Lazzaro et al. 2020). Shifted species assemblages have profoundly modified aquatic biodiversity and ecosystem functioning in many European wetlands (Rodríguez-Merino et al. 2018).
Diversity changes in plant communities are generally well documented in herbarium collections, because the new incomers spark scientific interest for their systematic or environmental significance, especially in the early stage of invasion (Stinca et al. 2021; Spampinato et al. 2022). Herbaria are important sources of historical data, especially for ecosystems like wetlands that have undergone dramatic changes in the last centuries (Domina et al. 2020). Despite the many biases associated with opportunistic and non-systematic plant collections (Daru et al. 2018), herbaria can be considered as unique repositories of phytogeographical data and one of the most accurate data sources to reconstruct events that occurred in the past. Therefore, the role of herbarium records has widened in recent years, from cataloging the diversity of life to documenting biodiversity changes (Muller 2016; Nualart et al. 2017; Lang et al. 2019) and tracing spatio-temporal invasion patterns (Lavoie 2013; Muller 2015).
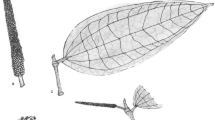
Elodea canadensis Michx. (Hydrocharitaceae; Fig. 1) is a submersed macrophyte with a long invasion history in Europe, resulting in a large number of specimens in the European and Italian public herbaria. Its invasion history is particularly interesting because, about one century after its introduction, another closely related species, E. nuttallii (Planch.) H. St. John (Fig. 2), was recorded in Europe and has ever since competed with E. canadensis for the same habitat, and even outcompeted it in many of the habitats previously invaded by the latter (Simpson 1990; Erhard and Gross 2006; Greulich and Trémolières 2006; Zehnsdorf et al. 2015). The first records of these species in Europe date back to 1836 (Northern Ireland) for E. canadensis and 1939 (Belgium) for E. nuttallii (Wolff 1980; Simpson 1984). The more aggressive invasion by E. nuttallii suggests that E. canadensis invasion should be slowing down or even declining (post-invasion status), whereas E. nuttalli should be in a phase of exponential spread. E. nuttallii is in fact shortlisted among the invasive alien species of European Union concern for which early eradication and/or control are mandatory (European Commission 2017, 2019).

Close-up of the terminal part of a sterile branch of Elodea canadensis. Note the leaves with apex obtuse or nearly rounded. Photograph taken by A. Moro—http://dryades.eu—, Madrisio (municipality of Fagagnana del Friuli), 23-04-2005. License CC BY-SA 4.0

Close-up of the terminal part of a branch of Elodea nuttalli. Note the leaves linear-lanceolate, curved, with apex acute and nearly acuminate. Photograph taken by Andrea and Riccardo Truzzi—http://dryades.eu—, Civico Museo di Storia Naturale, Milano—La Flora Esotica Lombarda. License CC BY-SA 4.0
Both species are perennial rooting hydrophytes, native to stagnant to flowing freshwaters of North America. The leaf shape can distinguish them: from ovate to linear-oblong (occasionally linear-lanceolate) with apex broadly acute to obtuse in E. canadensis, linear or linear-lanceolate with apex narrowly acute to acuminate in E. nuttallii (Simpson 1988). Apart from these slight morphological differences, the two species have analogous ecological needs: there are no precise limits between the functional niches of E. canadensis and E. nuttallii, so that they were defined as true ecological redundants (Hérault et al. 2008; Bubíková et al. 2021). Both have fast growth and spreading ability, due to their exclusively vegetative propagation (only female individuals are known in Europe, see Walters 1980), and high phenotypic plasticity (Agrawal 2001; Hérault et al. 2008; Riis et al. 2010; Kočić et al. 2014), excellent qualities for invasive species. In Italy, two morphotypes are known for E. nuttallii, often co-occurring, and cases of intra-individual heterophylly were also reported; one morphotype has short and curved leaves, the other has long and flat or more or less twisted leaves (Banfi and Galasso 2010). In Europe, the latter was sometimes interpreted as a separate species, E. callitrichoides (Rich.) Casp. (e.g. Wolff 1980; Vanderpoorten et al. 2000), but morphological and genetic analyses demonstrated that these morphotypes are two different phenotypes of the same species, E. nuttallii (Vanderpoorten et al. 2000). This phenotypic variability confounds the taxonomy of the two Elodea species and contributes to their misidentification (cfr. Kočić et al. 2014 and references therein).
Because of their large and dense monospecific populations, E. canadensis and E. nuttallii are considered ecosystem-transforming species (Buccheri et al. 2019). Communities dominated or co-dominated by E. canadensis and/or E. nuttalli are included in the Potametea pectinati Klika in Klika & V. Novák 1941 syntaxonomic class, and represent about 10% of aquatic alien-dominated plant communities in Italy (cfr. Viciani et al. 2020; Castello et al. 2021).
The aim of this study was to reconstruct the invasion history, dynamics and current distribution of E. canadensis and E. nuttallii in Italy based on occurrences from herbarium specimens, field records and historical literature.
Since many new alien species are continuously arriving in Europe and Italy, we aimed at understanding a posteriori what happened in the first invasion phases of these two hydrophytes, to learn from past invasions how to interpret, monitor and manage current and future introductions of alien aquatic plant species.
Materials and methods
Data sources
Occurrence records of E. canadensis and E. nuttallii dating between 1850 and 2019 were collected from June 2019 to November 2020 from 41 independent sources, including 36 Italian herbaria owned by Italian universities or scientific museums and 4 private herbaria (hereafter mentioned as «herbarium records»; herbarium identification codes follow Thiers 2022). Additional data were collected from published sources (floristic checklists, local floras or other floristic records; hereafter «published records») and from unpublished floristic or vegetation data (observations, relevés etc.) collected in the field by the authors in the period 1990–2020 (hereafter «field records»).
Herbarium specimens were searched through the currently accepted names and their synonyms; in particular, for E. canadensis we searched for Anacharis alsinastrum Planch., A. canadensis (Michx.) Planch., Elodea canadensis Michx., Philotria canadensis (Michx.) Britton, while for E. nuttalli we searched for Anacharis nuttallii Planch. and Philotria nuttallii (Planch.) Rydb. Species identification was checked and corrected, whenever necessary, by the authors and herbarium curators.
In total, our database included 1131 records (877 of E. canadensis, 254 of E. nuttallii), of which 506 herbarium records, 584 published records and 41 field records (Supplementary material).
A parallel historical literature search, describing the introduction history of E. canadensis and E. nuttallii to Italy until their inclusion in the Italian flora, served as a reference to crosscheck introduction pathways, dates, and localities (hereafter «historical literature»). For E. canadensis we searched on Google Books (https://books.google.com) in October 2020, using the key words «Elodea canadensis» and the previously mentioned synonyms. We selected only Italian sources dating between 1850 and 1900. For E. nuttalli, that was introduced in more recent times, we also screened grey literature accompanying the first field records of the species in Italy, between 1985 and 2005.
Standardising and georeferencing data
For each occurrence record we retained the following information: coordinates (when available) and/or location (i.e. described locality), collection date, habitat and collector.
When coordinates were not available, the occurrences were georeferenced based on the locality description using Google Hybrid maps available in QGIS (Imagery 2021, © Google) and the toponym layer repository available on the Geoportale Nazionale (http://wms.pcn.minambiente.it/ogc?map=/ms_ogc/WMS_v1.3/Vettoriali/Toponimi_2011.map). If a locality corresponded to an administrative division (municipality, province, region), the coordinates were referred to the centroid of that unit. For this purpose, the ISTAT (National Institute of Statistics, www.istat.it) polygon layers of the administrative boundaries chronologically closest to the collection date of a record were used. ISTAT layers are available about every 10 years from 1861 to 2001 and every year from 2002 to today. Overall, for 632 records the coordinates were already available; for 314 records the coordinates were obtained by finding the locality using Google Hybrid; 170 records were referred to the centroid of the administrative unit: 109 to the municipality level, 48 to the province level, 13 to the region level. Only 14 records had no spatial data. The reference system used for all coordinate pairs was WGS84 (EPSG 4326). A radius was associated to each pair of coordinates in order to measure the uncertainty of a locality linked to the georeferencing procedure.
Concerning collection date, 755 records had a complete date (day, month and year), 52 had only month and year, 255 had only the year, 68 had no date.
After removing records missing spatial and/or temporal information, the final dataset consisted in 805 records of E. canadensis and 248 records of E. nuttallii.
Finally, habitat information derived from the herbarium specimen labels, which was reported in a wide range of formats, was classified into the following 13 habitat types: BAS (basins, fountains, troughs and small artificial water bodies with still water), BGA (botanical gardens), BOG (bogs and peatlands), CAN (artificial canals, including city canals, wastewater canals, agricultural ditches and any man-made canals with running water), LAK (natural lakes), MAR (marshes and swamps), PIS (pisciculture ponds), PON (natural ponds), RIV (natural rivers), RFI (rice pads and associated ditches), SPR (springs), WET (wet meadows, flooded forests) and OTH (other habitats, occasional in frequency or poorly described, like woodlands, fields, filtering systems, etc.).
Data analysis
Occurrence mapping
For each of the two Elodea species, we mapped the occurrence records in different time periods in order to observe different invasion phases. Four distribution maps were produced for E. canadensis (periods 1850–1900, 1901–1950, 1951–1980 and 1981–2019) and two for E. nuttallii (1980–2000 and 2001–2019).
In order to avoid redundancy and reduce the bias associated with different collection efforts in different areas, while maintaining spatial information (Antunes and Schamp 2017), the georeferenced records of each species were thinned by overlaying a grid and extracting only one record per grid-cell per year. For this purpose we used the Italian 177 cells of the 10 × 10 km grid by Cervellini et al. (2020), that follows the requirements of article 17 of the Habitats Directive (92/43/EEC). Each grid-cell is assigned to one European biogeographical region and indicates Nature 2000 habitats that it contains (Cervellini et al. 2020). We used gridded data for the period 2000–2019, to test differences between the two species in biogeographic distribution, habitats threatened by the occurrence of the species within the cell, and main land use. For land use we extrapolated the amount of urban and agricultural areas in cells where the two species occur or co-occur by intersecting the grid cells with the 2012 CORINE Land Cover layer (www.isprambiente.gov.it). Differences between agricultural and urban areas were tested between species with a one way-ANOVA (PAST 4.03; Hammer et al. 2001). Data mapping and spatial analysis were performed with QGIS version 3.12.2 (www.qgis.org).
Invasion curves
Invasion curves based on cumulative numbers of occurrence records over time can define three main invasion phases: lag, exponential growth and plateau (Pyšek and Prach 1993; Blackburn et al. 2011; Antunes and Schamp 2017). The temporal length of these three phases and the rate of spread, defined by the derivative of the curve, can vary from species to species (Crawford and Hoagland 2009; Antunes and Schamp 2017; Ceschin et al. 2018) and can be useful to understand, a posteriori, how invasions evolved. We used the thinned and gridded dataset of records to construct the invasion curves of E. canadensis and E. nuttallii in Italy and calculate the curve slope of their linear models.
Results
Invasion history from occurrence records and historical literature
A herbarium specimen of E. canadensis with a hypothetical date (1880?) from the Province of Mantova in northern Italy (Leg. F. Masè, s.d., in MSPC; Table 1) might be the most ancient specimen on a national scale if the collection date could be confirmed. The first certain herbarium accessions are two samples of 1888 collected in northern Italy, around Lake Como (Leg. M. Longa, 7–1888, in FI; Leg. M. Longa, det. G. Camperio, 7–1888, in FI; Table 1). The first Italian published record that we could trace in the historical literature, documenting the occurrence of E. canadensis in the wild, dates back to 1873 (Goiran 1897: San Michele Extra, near Verona, northern Italy). However, for some years before 1873 the species had been the object of an intense samples’ exchange for scientific research: in 1866 E. canadensis was sent to Mantova botanic garden, in 1867 it was introduced in the Botanic Garden of Padova and, shortly thereafter, it started naturalising in the Italian territory (Paglia 1879; Bozzi 1888; Banfi and Galasso 2010). The species was included in the Italian national flora in 1908 (Fiori and Paoletti 1896–1908). The historical literature shows that the species was present in Europe in the nineteenth century and was cited for the first time in Italian literature by Antonio Stoppani (1873), who reported a possible introduction to northern Europe from North America with timber rafts and documented the invasive spreading in European freshwater causing problems to river navigation. Between 1873 and 1908, the species was used in fish farming and its response to carbon and light was tested experimentally (Tolomei 1893). Some scientists even suggested its introduction in malarial wetlands (Gasperini 1890) as a natural remedy against mosquitoes (note that in Italy malaria was eradicated only in the 1950s).
The herbarium records show that in 1900 E. canadensis was already naturalised and widespread in northern Italy, while in the Mediterranean region it was cultivated in the Botanic Gardens of Pisa (Leg. P. Pellegrini, 6–1892, in PI) and Rome (Leg. E. Chiovenda, 23-5-1899, in BOLO) and occurred in canals nearby and fishponds of the Royal Palace of Caserta, in southern Italy (Fiori 1895; Fig. 3a). At the end of 1800 it was naturalised in ditches in the territory of Padova, in northern Italy: three herbarium specimens prove the occurrence of E. canadensis in the countryside ca. 25 km from Padova (Leg. U. Ugolini, 1892, in PAD), in town (Leg. Adr. Fiori, 6–1894, in FI), and in the ditches of Padova Agricultural Garden (Leg. L. Vaccari, 11–1895, in FI; Table 1). In the subsequent years the expansion was more intense in the continental (northern Italy) than in the Mediterranean region (Fig. 3b, c, d). Additional herbarium specimens come from new streams close to Padova Botanic Garden and the Royal Palace of Caserta (1901–1950, Fig. 3b) and from streams close to Pisa Botanic Garden (1901–2019, Fig. 3b, c, d). According to our reconstruction, the most aggressive invasion phase was in the period 1980–2019 (Fig. 3d), that counts 545 more herbarium records than in the period 1950–1980 (Fig. 3c). In the most recent map (1981–2019, Fig. 33333d) it is possible to identify long river stretches invaded by E. canadensis along the north–south valleys of big Italian rivers that originate in the Alps, and scattered occurrences in the Mediterranean region that cannot be obviously connected to botanic gardens or other sources.

Reconstruction based on herbarium records of the distribution of Elodea canadensis in different temporal ranges. Botanic gardens and the Royal Park of Caserta, in which, according to the literature, the species was cultivated, are also shown with red and blue diamonds respectively. NE is the Botanic Garden of Padova, W-centre is the Botanic Garden of Pisa and SW is the Botanic Garden of Rome

Cumulative distribution of Elodea nuttallii records from 1980 to 2000 (A) and from 2001 to 2019 (B)

Invasion curves of Elodea canadensis and E. nuttallii in Italy, based on herbarium records, published records and field records. Trend line and equation of the curve are also shown
The first Italian herbarium record of E. nuttalli dates back to 1989 and comes from the Lake Idro, in northern Italy; it is preserved at the Herbarium of Rovereto (ROV) and was first cited in the literature some years later (Desfayes 1995; Selvaggi and Dellavedova 2016). The species was subsequently observed in nearby areas (Zanotti 2000) and was included in the checklist of the Italian vascular flora in 2005 (Conti et al. 2005). In Italy, the introduction of E. nuttallii was due to the trade of ornamental species for artificial ponds and aquaria (Banfi and Galasso 2010). In other parts of Europe, the species was first found in Belgium in 1939, likely carried by migrating waterfowls (Verloove 2006); however, it is plausible that the species went unobserved for a long time, given the morphological and taxonomic similarity to E. canadensis (Walters 1980) and the presence of two different E. canadensis phenotypes (Banfi and Galasso 2010).
The invasion of Elodea nuttallii has been observed in northern Italy for the last 40 years (1980–2019). No occurrences were recorded in the Mediterranean biogeographic region of Italy so far (Fig. 4).
Invasion curves
Elodea canadensis invasion curve shows a lag phase from 1866 to 1892, an expansion phase from 1893 to 1923, and a plateau phase from 1925 to 1971 (Fig. 5). After this plateau, the curve has a second exponential rise, indicating a second phase of expansion from 1977 to today, suggesting that the invasion has not come to a halt yet. However, since 2005 the slope of this second expansion has become less steep, suggesting that the invasion is approaching the second plateau phase. The slope of the whole curve from 1866 to 2019 is quite moderate (angular coefficient of the trend line 3.03).
Elodea nuttallii had a lag phase between 1989 and 2000 and is still in the expansion phase (Fig. 5). The slope was most steep between 2000 and 2003 and the curve shows a breaking point around 2003, after which the expansion has been slowing down, suggesting that E. nuttalli is reaching the plateau phase too. The slope of the curve from 1989 to 2019 is quite steep (angular coefficient of the trend line 6.26). It has however to be noted that the cumulative record number remarkably differs between the two species: for E. canadensis it is 549, whereas for E. nuttallii it is only 152 (Fig. 5).
Biogeography and habitats
Both species occur in the Continental and Alpine biogeographic regions within Italy, although with notable differences in the number of occupied cells (the Continental region is more invaded than Alpine one). E. nuttalli occurs in the same grid cells as E. canadensis and only 3 cells are occupied by E. nuttalli alone. Only E. canadensis occurs in the Mediterranean region, and the number of cells occupied in the continental region in northern Italy is an order of magnitude higher than that of E. nuttallii (deduced by the number of cells assigned to continental, alpine and mediterranean biogeographic regions occupied by the two species).
Artificial canals (including ditches; habitat code CAN), streams (RIV) and lakes (LAK) are the most common water bodies where E. canadensis and E. nuttallii herbarium specimens were collected (Fig. 6). Until 1980, canals were the most frequent habitat for E. canadensis recorded in the herbarium dataset. However, from 1990 onwards the number of E. canadensis occurrence records from rivers and lakes increased considerably, with river records exceeding canal records in 2000–2010. E. nuttallii occurred almost exclusively in lakes until 2000, but in the subsequent decade it was most frequently recorded in canals and rivers. In the last 10 years, E. canadensis was mostly found in canals, whereas E. nuttalli in rivers, however differences in the number of occurrences among canals, rivers and lakes became less evident in both species.

Most frequent habitats of Elodea canadensis (A) and E. nuttallii B recorded in herbarium specimen labels. Abbreviations: CAN: canals, RIV: rivers, LAK: lakes
Natura 2000 habitats (sensu Habitats Directive 92/43/EEC) that are more frequent within Elodea occurrence cells are 91E0* (Alluvial forests with Alnus glutinosa and Fraxinus excelsior—Alno-Padion, Alnion incanae, Salicion albae), 3260 (Water courses of plain to montane levels with the Ranunculion fluitantis and Callitricho-Batrachion vegetation) and 3150 (Natural eutrophic lakes with Magnopotamion or Hydrocharition-type vegetation), both in the Continental and Alpine biogeographic regions (Fig. 7). Peat bogs (habitat 7230: Alkaline fens) and petrifying springs (habitat 7220*: Petrifying springs with tufa formation—Cratoneurion) are also frequent in Elodea-invaded cells in the Alpine region.

Natura 2000 habitats in Elodea canadensis and E. nuttallii occurrence cells in the Alpine, Continental and Mediterranean biogeographic regions. Biogeographical regions: ALP: alpine, CON: continental, MED: Mediterranean. Explanation of habitat codes: 1150*: coastal lagoons, 1310: Salicornia and other annuals colonizing mud and sand, 1410: Mediterranean salt meadows (Juncetalia maritimi), 3130: Oligotrophic to mesotrophic standing waters with vegetation of the Littorelletea uniflorae and/or of the Isoëto-Nanojuncetea, 3140: Hard oligo-mesotrophic waters with benthic vegetation of Chara spp., 3150: Natural eutrophic lakes with Magnopotamion or Hydrocharition-type vegetation, 3160: Natural dystrophic lakes and ponds, 3170*: Mediterranean temporary ponds, 3220: Alpine rivers and the herbaceous vegetation along their banks, 3230: Alpine rivers and their ligneous vegetation with Myricaria germanica, 3240: Alpine rivers and their ligneous vegetation with Salix eleagnos, 3250: Constantly flowing Mediterranean rivers with Glaucium flavum, 3260: Water courses of plain to montane levels with the Ranunculion fluitantis and Callitricho-Batrachion vegetation, 3270: Rivers with muddy banks with Chenopodion rubri p.p. and Bidention p.p. vegetation, 3280: Constantly flowing Mediterranean rivers with Paspalo-Agrostidion species and hanging curtains of Salix and Populus alba, 3290: Intermittently flowing Mediterranean rivers of the Paspalo-Agrostidion, 6410: Molinia meadows on calcareous, peaty or clayey-siltladen soils (Molinion caeruleae), 6420: Mediterranean tall humid herb grasslands of the Molinio-Holoschoenion, 6430: Hydrophilous tall herb fringe communities of plains and of the montane to alpine levels, 7110*: Active raised bogs, 7120: Degraded raised bogs still capable of natural regeneration, 7140: Transition mires and quaking bogs, 7150: Depressions on peat substrates of the Rhynchosporion, 7210*: Calcareous fens with Cladium mariscus and species of the Caricion davallianae, 7220*: Petrifying springs with tufa formation (Cratoneurion), 7230: Alkaline fens, 91D0*: Bog woodland, 91E0*: Alluvial forests with Alnus glutinosa and Fraxinus excelsior (Alno-Padion, Alnion incanae, Salicion albae), 91F0: Riparian mixed forests of Quercus robur, Ulmus laevis and Ulmus minor, Fraxinus excelsior or Fraxinus angustifolia, along the great rivers (Ulmenion minoris)
In the Mediterranean regions, the most frequent habitats in Elodea-invaded grid cells are 3140 (Hard oligo-mesotrophic waters with benthic vegetation of Chara spp.), 3150 (Natural eutrophic lakes with Magnopotamion or Hydrocharition-type vegetation) and 3170* (Mediterranean temporary ponds).
Current land use in the grid cells where the two species occur is primarily agriculture (in over 50% of each grid cell) for both species. Urbanisation accounts for only about 8% in each grid cell for both species, nevertheless urbanisation percentage is significantly higher (about 14%, P < 0.001) in cells where both species co-occur rather than in those where they occur alone (Fig. 8).

Land use in Elodea canadensis and E. nuttallii occurrence cells: urbanisation and agriculture percentage per cell
Discussion
Invasion history from occurrence records and historical literature
The first record of E. canadensis in Europe dates back to 1836 in the British Isles (Simpson 1984), whereas in Italy the species appeared 30 years later in parks and botanic gardens. The published records document the cultivation in the Botanic Garden of Mantova in 1866, in that of Padova in 1867, later in that of Pavia (Bozzi 1888; Banfi and Galasso 2010). In the same period, E. canadensis was already widespread in many parts of central Europe and was becoming so invasive that it was the most common alien aquatic species in Europe for a while (Erhard and Gross 2006; Zehnsdorf et al. 2015). The first ascertained Italian herbarium samples were collected in 1888 from northern Italy, that is the Italian area of first introduction and most impacted by the invasion.
Up until 1890, in Italy E. canadensis was regarded as a useful plant for its ability to recover marshes from malaria, assist aquaculture and serve as a fertiliser (Quaglia and Soave 1886, 1889; Gasperini 1890); some authors even suggested its use to support the colonisation of Africa, where malaria fevers prevented Italian colonisers’ success (Gasperini 1890). Even though it was already considered a water pest in Europe (Quaglia and Soave 1886), only in the last years of the nineteenth century the signs of an invasion began to be reported by the Italian botanists (Pasquale 1894, 1896; Fiori 1895), associated with the first local extinctions of rare native aquatic plant species (mainly in the families Haloragaceae, Hydrocharitaceae and Najadaceae; Cavara 1894). We can therefore assume that the species needed about 20–30 years to establish before starting invading new territories, as also shown by the invasion curve.
The initial spread affected principally northern Italy, with isolated occurrences in the centre and south. Escapes from botanic gardens probably played an important role in the spreading of E. canadensis, as already proposed by Fiori (1895), and as documented by our temporal distribution maps in the areas surrounding the botanic gardens of Padova, Pisa and Rome (Leg. U. Ugolini, 1892, in PAD; Leg. Adr. Fiori, 6–1894, in FI; Leg. E. Chiovenda, 17-9-1901, in BOLO; Leg. M. Savelli, 16-10-1913, in FI). The Park of the Royal Palace in Caserta was probably another centre of spread for the invasion in southern Italy. E. canadensis was often introduced as an ornamental plant (Zehnsdorf et al. 2015) for fishponds or accidentally with exotic fishes (Thomson 1922) and quite likely it was also the case in the park of a Royal Palace.
Through the years, the expansion continued primarily in northern Italy. Occurrence records in central and southern regions between 1980 and 2019 (Fig. 3d) were, as expected, centred around the older records associated with the botanic gardens and the Royal Palace of Caserta, but also included occasional new occurrences in regions that previously were not affected by the invasion, suggesting invasion corridors from North to South Italy or new introductions.
Elodea nuttallii invasion took place in the last 40 years (1980–2020), without any records in central and southern Italy so far. As its ecological requirements are very similar to those of E. canadensis (Kočić et al. 2014; Zehnsdorf et al. 2015), its more limited distribution compared to that of E. canadensis might be related to time rather than ecology. Since the first record of E. nuttallii dates back to 1989, the species has spread for only 33 years in Italy, compared to the 160 years of E. canadensis invasion. The central part of northern Italy is the most invaded region; the occurrence in the rest of northern Italy is sporadic.
Like E. canadensis, E. nuttallii is a cold-temperate species that found its climate niche within the European continental biogeographic region, that in Italy roughly extends up to the northern Apennine watershed and follows the Apennines parallel to the Adriatic coasts for half of the peninsula (European Environment Agency 2017).
Invasion curves
Elodea canadensis has been in Italy at least since 1866, and showed two invasion phases: the first in the period 1890–1930 and the second, more aggressive, from around 1980 onwards. An unprecedented effort in the recording of biodiversity and invasive species after 1980 might have skewed the curve with an artifactual recent expansion phase (+ 33.3% accessions for E. canadensis, + 894.7% for E. nuttallii compared to previous period; see Fig. 5). From the 1930s up to the end of the century, in Italy there was a general decrease in systematics studies, with a consequent decrease in herbarium accessions dating to that period. In addition, the Second World War imposed a forced stop to nearly all field activities, at least in the years 1940–1945, and the contemporary destructions caused by bombings led to the loss of some herbarium collections (see e.g. Taffetani 2012, pp. 734 and 743). All of this could have contributed to a further underestimation of the real presence of E. canadensis in Italy in the period 1930–1980. However, it should be noted that the second invasion round is accompanied by a switch in the habitat. In addition to canals, that were E. canadensis preferential habitat until 1970 (registered in the herbarium specimen labels), also rivers and lakes became important for the species after this period: in 2000–2010 the occurrences in rivers even exceeded those in canals (Fig. 6a). The colonisation of new habitats could be due to evolution which occurred post-introduction in the new range or to a cryptic invasion of a second genotype of E. canadensis. The trade of this species has never ceased and introductions of new, still undiscovered genetic lineages may have been overlooked. The new genotype, better adapted to eutrophication and the increasing temperatures of the early twenty-first century, would preferably have occupied eutrophic rivers and lakes, a hypothesis that can be confirmed by genetic analysis. In addition, the 1970s witnessed a massive use of pesticides and herbicides that depleted biodiversity in agricultural areas (Santini and Buldrini 2012): this phenomenon could have accounted for a reduced expansion of E. canadensis in canals in agricultural areas until the 1980s and the subsequent colonization of other water bodies (cfr. Bowmer et al. 1995; Glomski et al. 2005).
Other hypotheses concern the ability of Elodea spp. to thrive in stressed environments, that might have facilitated the expansion to new areas, and, not last, the initial misidentification of E. nuttallii with E. canadensis when it first appeared in Italy. The two species are morphologically very similar (Walters 1980; Vanderpoorten et al. 2000; Banfi and Galasso 2010) and the still imperfect knowledge of the newly arrived E. nuttalli could have induced various botanists to erroneously identify it as E. canadensis, causing an overestimation of the presence and spreading ability of the latter at the end of the twentieth century. Another still largely unexplored factor that might have had a role in the colonization of new habitats could be the co-occurrence of other invasive species, as documented for E. nuttalli in Ireland where patch extension was found to be positively correlated with the presence of Dreissena polymorpha Pallas, 1771 (Crane et al. 2022).
Biogeography and habitats
For both species, the invasion range in Italy is centred in the northern part, i.e. in the continental and Alpine biogeographical regions, with isolated occurrences of E. canadensis in central and southern Italy, i.e. in the Mediterranean region. Both species have never been recorded in Sardinia and Sicily and in the small islands around Italy (Celesti-Grapow et al. 2016). This fact is not surprising since the two species are native to northern America (Simpson 1984) and are typical of cold-temperate climates. Indeed, the trophic and thermic seasonal fluctuations in inland waterbodies could be the reason for the lower invasion pressure of Elodea species in the Mediterranean region compared to temperate ecosystems (Guarino et al. 2021). However, the complete absence (at least at the current state of the floristic knowledge) of E. nuttallii in the Mediterranean region suggests that the two species, even if regarded as ecological redundants (Hérault et al. 2008), have some different climatic requirements, with E. nuttallii being less thermophilous than E. canadensis. This fact was already reported by Pignatti et al. (2017–2019) and can also be observed in the habitats recorded for the first occurrences of the two species: mostly canals (i.e. shallow waters, with tendency to summer heating) for E. canadensis, from the beginning up to 1980; mostly lakes (i.e. deep, cold waters, with scarce or negligible summer heating) for E. nuttallii, from the introduction up to 2000. Variation in temperature-associated conditions could be more marked in waters at the margin of the temperate range. Irrespective of the species, a shift from the original habitat to other types of waterbodies has been evident in the last decades: this could be due to adaptation to the new environmental conditions (cfr. Allard 1988), or selection of the most resistant genotypes (Lambertini et al. 2010; Riis et al. 2010; Johnson and Munshi-South 2017), or simply the species had physical access to other habitats. Therefore, the distribution of E. canadensis and E. nuttallii might still change in coming years, especially in central and southern parts of Italy where the distribution range of these species is disjunct, and some areas are poorly investigated (Conti et al. 2016; Stinca et al. 2017; Rosati et al. 2020).
For the time being, the largest alluvial plain of Italy, i.e. the Po Plain, is the area with the highest number of records. The area is quite rich in permanent water bodies and one of the most impacted by human activities in Italy (Bolpagni et al. 2020). Here the probability of finding both species increases with increasing urbanisation and water exploitation independently of taxa (Fig. 8).
River Po crosses northern Italy from west to east and appears as an important barrier for the spread of Elodea species in the peninsula. This is likely because the Apennine catchments, south of River Po, have a torrential regime and undergo a long lean period (and even drought) during the summer months that reduces survival capacity and migration of aquatic species to the Mediterranean region. The Alpine catchments, on the contrary, are supplied by Alpine glaciers all year around, and although they may have lean periods in the summer, they never, or only exceptionally, experience drought. Despite the drought barrier, the risk of spreading of the two species south of River Po is high, since the artificial network of canals that brings irrigation water from River Po to the agricultural lands south of the river (Montanari et al. 2020, 2022) can provide dispersal corridors in the dry season. During summer, in fact, these canals are maintained full of water (Dallai et al. 2015; Montanari et al. 2020), and provide a suitable environment for the survival of both Elodea species exactly when natural watercourses undergo lean or drought. The risk, in the near future, of a mass spreading of the two species to the southern parts of the Po Plain, that at present are still not extensively invaded, is therefore very high, also considering the intensive management of the canal networks (mowing of the vegetation and dredging of the canals) and the more and more frequent flooding events that occur in this area. In addition to the obvious ecosystemic impact, an invasion in this area would have negative consequences for canal maintenance and effectiveness in draining rainwaters (hydrogeological risk is high in this region) and supplying irrigation water (cfr. Dallai et al. 2015). Monitoring Elodea and other invasive aquatic species in this area is therefore strategic to prevent invasions in the peninsula, as well as to plan eradication measures in advance that, in the case of E. nuttallii, are mandatory (EU Reg. 1143/2014). An option that should be considered is that of repristing occasional drought barriers that can interrupt the migration flow and have proved successful in containing these invasions for the past 100 years. In any case, given that eradication and control are obligatory for E. nuttallii, and were successful in other parts of Europe (Hoffmann et al. 2013; Garland et al. 2022), it is also possible (and desirable) that containment actions have a positive effect on the distribution of both species that largely occur in the same areas, thereby leading to a contraction, rather than an expansion, in the ranges of both species in the coming years.
Natura 2000 habitats that occur within Elodea occurrence records cells are the ones that are most threatened by the invasion of these species. Based on our field work, we can confirm that in the Po Plain both species frequently occur in habitats 3150 (Natural euthrophic lakes with Magnopotamion or Hydrocharition-type vegetation; see also Bolpagni 2013) and 3260 (Water courses of plain to montane levels with the Ranunculion fluitantis and Callitricho-Batrachion vegetation), as well as in temporary alluvial forests formed by the swollen rivers of the Alpine catchment, as detected by our study, often together with other invasive elodeids like Egeria densa (Planch.) Casp. (native to South America) and Lagarosiphon major (Ridl.) Moss (native to South Africa). Elodeas and alien species are one of the major threats for the conservation of Natura 2000 habitats in Italy, both aquatic and terrestrial (Lazzaro et al. 2020; Viciani et al. 2020).
Mediterranean habitats appear more resilient to Elodea invasion because of summer drought, but also because many rivers are dammed for the creation of water reservoirs, and artificial networks for drainage and irrigation are not as extended as in northern Italy. Nevertheless, there are various lakes (both natural and artificial) that provide suitable establishment sites to E. canadensis and E. nuttallii, in case of introduction in these zones. As a matter of fact, we have recently become aware of a few more lakes in internal areas of the peninsula where E. canadensis is currently present and could have occurred for a long time, like Lake Trasimeno in central Italy, where it has been known since the end of 1800 and whose presence could be linked to the various experiments of aquaculture that were discussed and performed in the 1880s (Ministero di Agricoltura, Industria e Commercio 1886, 1887). The distribution of Elodea species could therefore be wider in the Mediterranean region than documented by this study, and the invasion risk should not be underestimated even in this area.
Conclusions
This study reconstructed the invasion history of E. canadensis and E. nuttallii in Italy, two aquatic pests that occur in large parts of Eurasia. Herbarium specimens, occurrence records and historical literature provided insight into the introduction history and dynamics of two invasions by two closely related species. With this information we could identify a strategic area where to address monitoring and management, for the prevention of further spreading. The study also provides new perspectives on the invasion process of plant species. To the best of our knowledge, this is the first study that resolves the initial invasion phase of a plant species with such a detail, especially for E. canadensis. Interestingly, in the nineteenth century the introduction pathways were not very different from those of today, i.e. deliberate human introduction (fish farming, botanic garden collections and applied research then, as documented by this study, aquarium and ornamental plants trade, living collections, cultivation, scientific research and phytoremediation today—Kay and Hoyle 2001; Hulme 2011; Brundu 2015; van Kleunen et al. 2018a, 2018b).
Compared to the invasion curve of E. nuttalli introduced 100 years later in the same area, E. canadensis had a longer lag phase and a less steep expansion in the first invasion round between 1890 and 1930. The steepness (i.e. spread rate) of the second expansion phase was, instead, more similar to that of E. nuttallii invasion occurring in the same period 1990–2005. E. canadensis invasion curve also shows that invasions can have more than one expansion phase as previously suggested (Pyšek and Prach 1993). More research is necessary to corroborate this new finding, i.e. to assess the evolutionary change that was recorded by these occurrence records.
Data availability
All data generated or analysed during this study are included in this published article (and its supplementary information files).
References
Agrawal AA (2001) Phenotypic plasticity in the interactions and evolution of species. Science 294:321–326. https://doi.org/10.1126/science.1060701
Allard RW (1988) Genetic changes associated with the evolution of adaptedness in cultivated plants and their wild progenitors. J Hered 79(4):225–238. https://doi.org/10.1093/oxfordjournals.jhered.a110503
Antunes PM, Schamp BS (2017) Constructing standard invasion curves from herbarium data—towards increased predictability of plant invasions. Invasive Plant Sci Manage 10(4):293–303. https://doi.org/10.1017/inp.2017.38
Banfi E, Galasso G (2010) La flora esotica lombarda. Museo di Storia Naturale di Milano, Milano, pp 1–273
Blackburn TM, Pyšek P, Bacher S, Carlton JT, Duncan RP, Jarošík V et al (2011) A proposed unified framework for biological invasions. Trends Ecol Evol 26(7):333–339. https://doi.org/10.1016/j.tree.2011.03.023
Bolpagni R (2013) Macrophyte richness and aquatic vegetation complexity of the Lake Idro (Northern Italy). Ann Bot (Roma) 3:35–43. https://doi.org/10.4462/annbotrm-10207
Bolpagni R (2021) Towards global dominance of invasive alien plants in freshwater ecosystems: The dawn of the Exocene? Hydrobiologia 848:2259–2279. https://doi.org/10.1007/s10750-020-04490-w
Bolpagni R, Laini A, Soana E, Tomaselli M, Nascimbene J (2015) Growth performance of Vallisneria spiralis under oligotrophic conditions supports its potential invasiveness in mid-elevation freshwaters. Weed Res 55(2):185–194. https://doi.org/10.1111/wre.12128
Bolpagni R, Laini A, Buldrini F, Ziccardi G, Soana E, Pezzi G, Chiarucci A, Lipreri E, Armiraglio S, Nascimbene J (2020) Habitat morphology and connectivity better predict hydrophyte and wetland plant richness than land-use intensity in overexploited watersheds: evidence from the Po plain (northern Italy). Landsc Ecol 35:1827–1839. https://doi.org/10.1007/s10980-020-01060-2
Boscutti F, Lami F, Pellegrini E, Buccheri M, Busato F, Martini F, Sibella R, Siura M, Marini L (2022) Urban sprawl facilitates invasions of exotic plants across multiple spatial scales. Biol Invasions 24:1497–1510. https://doi.org/10.1007/s10530-022-02733-6
Bowmer KH, Jacobs SWL, Sainty GR (1995) Identification, biology and management of Elodea canadensis, Hydrocharitaceae. J Aquat Plant Manage 33:13–19
Bozzi L (1888) Sopra alcune piante americane naturalizzate nei dintorni di Pavia. Atti Soc It Sci Nat 31:281–288
Brundu G (2015) Plant invaders in European and Mediterranean inland waters: profiles, distribution, and threats. Hydrobiologia 746:61–79. https://doi.org/10.1007/s10750-014-1910-9
Brundu G, Stinca A, Angius L, Bonanomi G, Celesti-Grapow L, D’Auria G, Griffo R, Migliozzi A, Motti R, Spigno P (2012) Pistia stratiotes L. and Eichhornia crassipes (Mart.) Solms.: emerging invasive alien hydrophytes in Campania and Sardinia (Italy). EPPO Bull 42(3):568–579. https://doi.org/10.1111/epp.12004
Bubíková K, Svitková I, Svitok M, Hrivnák R (2021) Invasive elodeas in Slovakia (Central Europe): distribution, ecology and effect on native macrophyte assemblages. Aquat Inv 16(4):617–636. https://doi.org/10.3391/ai.2021.16.4.03
Buccheri M, Boscutti F, Pellegrini E, Martini F (2019) La flora aliena nel Friuli Venezia Giulia. Gortania (Botanica, Zoologia) 40:7–78
Castello M, Poldini L, Altobelli A (2021) The aquatic and wetland vegetation of Lake Doberdò: an analysis for conservation value assessment of a disappearing lake of the Classical Karst (North East Italy). Plant Sociol 58(1):75–106. https://doi.org/10.3897/pls2020581/05
Cavara F (1894) Nuova stazione della Solidago serotina Ait. Malpighia 8:94–95
Celesti-Grapow L, Bassi L, Brundu G, Camarda I, Carli E, D’Auria G et al (2016) Plant invasions on small Mediterranean islands: an overview. Plant Biosyst 150(5):1119–1133. https://doi.org/10.1080/11263504.2016.1218974
Cervellini M, Zannini P, Di Musciano M, Fattorini S, Jiménez-Alfaro B, Rocchini D et al (2020) A grid-based map for the Biogeographical Regions of Europe. Biodivers Data J 8:e53720. https://doi.org/10.3897/BDJ.8.e53720
Ceschin S, Abati S, Ellwood NTW, Zuccarello V (2018) Riding invasion waves: spatial and temporal patterns of the invasive Lemna minuta from its arrival to its spread across Europe. Aquat Bot 150:1–8. https://doi.org/10.1016/j.aquabot.2018.06.002
European Commission (2017) Invasive alien species of Union concern. European Union, Publications Office of the European Union, Luxembourg
European Commission (2019) Commission implementing Regulation (EU) 2019/1262 of 25 July 2019 amending Implementing Regulation (EU) 2016/1141 to update the list of invasive alien species of Union concern. OJL 199(1):1–4
Conti F, Abbate G, Alessandrini A, Blasi C (2005) An annotated checklist of the Italian vascular flora. Palombi Editori, Roma
Conti F, Bartolucci F, Manzi A, Paolucci M, Santucci B, Petriccione B, Miglio M, Ciaschetti G, Stinca A (2016) Integrazioni alla flora vascolare dell’Italia Centrale. Atti Soc Tosc Sci Nat Mem Ser B 122:33–42. https://doi.org/10.2424/ASTSN.M.2015.05
Crane K, Kregting L, Coughlan NE, Cuthbert RN, Ricciardi A, MacIsaac HJ, Dick JT, Reid N (2022) Abiotic and biotic correlates of the occurrence, extent and cover of invasive aquatic Elodea nuttallii. Freshw Biol 67(9):1559–1570. https://doi.org/10.1111/fwb.13960
Crawford PHC, Hoagland BW (2009) Can herbarium records be used to map alien species invasion and native species expansion over the past 100 years? J Biogeogr 36(4):651–661. https://doi.org/10.1111/j.1365-2699.2008.02043.x
Dallai D, Buldrini F, Santini C, Fanti E, Tonelli F, Zampighi C (2015) Aspetti di tutela della flora idro-igrofila nei canali della Bonifica Burana. Atti Soc Nat Mat Modena 146:207–232
Daru BH, Park DS, Primack RB, Willis CG, Barrington DS, Whitfield TJS, Seidler TG, Sweeney PW, Foster DR, Ellison AM, Davis CC (2018) Widespread sampling biases in herbaria revealed from large-scale digitization. New Phytol 217:939–955. https://doi.org/10.1111/nph.14855
Desfayes M (1995) Appunti floristici sulle acque del Trentino e territori circostanti. Ann Mus Civ Rovereto Sez Archeol Stor Sci Nat 10:223–248
Domina G, Di Gristina E, Scafidi F, Calvo R, Venturella G, Gargano ML (2020) The urban vascular flora of Palermo (Sicily, Italy). Plant Biosyst 154(5):627–634. https://doi.org/10.1080/11263504.2019.1651787
Erhard D, Gross EM (2006) Allelopathic activity of Elodea canadensis and Elodea nuttallii against epiphytes and phytoplankton. Aquat Bot 85(3):203–211. https://doi.org/10.1016/j.aquabot.2006.04.002
European Environment Agency (2017) Biogeographical regions in Europe. https://www.eea.europa.eu/data-and-maps/figures/biogeographical-regions-in-europe-2
Fiori A (1895) L’Elodea canadensis Mich. nel Veneto e in Italia. Malpighia 9:119–120
Fiori A, Paoletti G (1896) Flora analitica d’Italia: ossia descrizione delle piante vascolari indigene inselvatichite e largamente coltivate in Italia disposte per quadri analitici, vol 4. Tipografia del seminario, Padova
Garland D, Earle W, Caffrey J, Taylor C, Meehan S, Touzet N, Lucy F (2022) Management of the invasive Nuttall’s pondweed (Elodea nuttallii) in Lough Arrow, a Natura 2000 designated lake in Western Ireland. Manag Biol Invasions 13(1):118–130. https://doi.org/10.3391/mbi.2022.13.1.07
Gasperini A (1890) Una pianta benefica. La scienza pratica 25:95-96
Glomski LAM, Skogerboe JG, Getsinger KD (2005) Comparative efficacy of diquat for control of two members of the Hydrocharitaceae: Elodea and Hydrilla. J Aquat Plant Manage 43:103–105
Goiran A (1897) Le piante fanerogame dell’agro veronese. Censimento, vol I. Tipografia Franchini, Verona, p 104
Greulich S, Trémolières M (2006) Present distribution of the genus Elodea in the Alsatian Upper Rhine floodplain (France) with a special focus on the expansion of Elodea nuttallii St. John during recent decades. Hydrobiologia 570(1):249–255. https://doi.org/10.1007/s10750-006-0188-y
Guarino R, Chytrý M, Attorre F, Landucci F, Marcenò C (2021) Alien plant invasions in Mediterranean habitats: an assessment for Sicily. Biol Invasions 23:3091–3107. https://doi.org/10.1007/s10530-021-02561-0
Hammer O, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software for education and data analysis. Palaeontol Electron 4(1):1–9
Hérault B, Bornet A, Trémolières M (2008) Redundancy and niche differentiation among the European invasive Elodea species. Biol Invasions 10:1099–1107. https://doi.org/10.1007/s10530-007-9187-9
Hoffmann MA, Benavent González A, Raeder U, Melzer A (2013) Experimental weed control of Najas marina ssp. intermedia and Elodea nuttallii in lakes using biodegradable jute matting. J Limnol 72(3):485–493. https://doi.org/10.4081/jlimnol.2013.e39
Hulme PE (2011) Addressing the threat to biodiversity from botanic gardens. Trends Ecol Evol 26(4):168–174. https://doi.org/10.1016/j.tree.2011.01.005
Hussner A (2009) Growth and photosynthesis of four invasive aquatic plant species in Europe. Weed Res 49(5):506–515. https://doi.org/10.1111/j.1365-3180.2009.00721.x
Imagery ©2021, TerraMetrics, Map data ©2021, GeoBasis-DE/BKG (©2009), Google, Inst. Geogr. Nacional
Johnson MTJ, Munshi-South J (2017) Evolution of life in urban environments. Science 358:eaam8327. https://doi.org/10.1126/science.aam8327
Kay SH, Hoyle ST (2001) Mail order, the Internet, and invasive aquatic weeds. J Aquat Plant Manage 39(1):88–91
Kočić A, Horvatić J, Jelaska S (2014) Distribution and morphological variations of invasive macrophytes Elodea nuttallii (Planch) H. St. John and Elodea canadensis Michx in Croatia. Acta Bot Croat 73(2):437–446. https://doi.org/10.2478/botcro-2014-0011
Lambertini C, Riis T, Olesen B, Clayton JS, Sorrell BK, Brix H (2010) Genetic diversity in three invasive clonal aquatic species in New Zealand. BMC Genet 11(1):52. https://doi.org/10.1186/1471-2156-11-52
Lang PLM, Willems FM, Scheepens JF, Burbano HA, Bossdorf O (2019) Using herbaria to study global environmental change. New Phytol 221(1):110–122. https://doi.org/10.1111/nph.15401
Lavoie C (2013) Biological collections in an ever changing world: herbaria as tools for biogeographical and environmental studies. Persp Plant Ecol Evol Syst 15(1):68–76. https://doi.org/10.1016/j.ppees.2012.10.002
Lazzaro L, Bolpagni R, Buffa G, Gentili R, Lonati M, Stinca A et al (2020) Impact of invasive alien plants on native plant communities and Natura 2000 habitats: state of the art, gap analysis and perspectives in Italy. J Environ Manage 274:111140. https://doi.org/10.1016/j.jenvman.2020.111140
Malavasi M, Acosta ATR, Carranza ML, Bartolozzi L, Basset A, Bassignana M et al (2018) Plant invasions in Italy: an integrative approach using the European LifeWatch infrastructure database. Ecol Ind 91:182–188. https://doi.org/10.1016/j.ecolind.2018.03.038
Ministero di Agricoltura, Industria e Commercio (1886) Annali di Agricoltura. Atti della Commissione consultiva per la pesca. Sessione 1886. Tipografia Eredi Botta, Roma, pp 9–14
Ministero di Agricoltura, Industria e Commercio (1887) Annali di Agricoltura. Atti della Commissione consultiva per la pesca. Sessione 1887. Tipografia Eredi Botta, Roma, pp 56–64
Montanari I, Buldrini F, Bolpagni R, Laini A, Dalla Vecchia A, De Bernardini N, Campione L, Castellari I, Gizzi G, Landi S, Chiarucci A (2020) Role of irrigation canal morphology in driving riparian flora in over-exploited catchments. Comm Ecol 21(2):121–132. https://doi.org/10.1007/s42974-020-00024-5
Montanari I, De Bernardini N, Gizzi G, Bolpagni R, Buldrini F, Campione L, Castellari I, Landi S, Spiezia L, Chiarucci A (2022) Flora and plant communities across a complex network of heavily modified water bodies: geographical patterns, land use and hydrochemical drivers in a temperate overexploited floodplain. Landsc Ecol Eng. https://doi.org/10.1007/s11355-022-00504-y
Muller S (2015) The use of herbarium specimens for investigating the spatio-temporal dynamics of biological invasions. Revue d’Ecologie (la Terre et la Vie) 70(Suppl. 12):229–235
Muller S (2016) The use of herbarium specimens for investigating macroecological and environmental changes. In: Rakotoarisoa NR, Blackmore S, Riera B (eds) Botanists of the twenty-first century: roles, challenges and opportunities – Based on the proceedings of the UNESCO International conference «Botanists of the twenty-first century: roles, challenges and opportunities» held in September 2014 in Paris. UNESCO, Paris, pp 78–87
Nualart N, Ibáñez N, Soriano I, López-Pujol J (2017) Assessing the relevance of herbarium collections as tools for conservation biology. Bot Rev 83(3):303–325. https://doi.org/10.1007/s12229-017-9188-z
Paglia E (1879) Saggio di studi naturali sul Mantovano. Mantova
Pasquale F (1894) La Marsilia quadrifoliata nelle provincie meridionali d’Italia e la Elodea canadensis Rich. in Italia. Bull Soc Bot Ital anno 1894(8):265–266
Pasquale F (1896) L’Elodea canadensis Rich. nelle provincie meridionali d’Italia. Bull Soc Bot Ital anno 1896(1):5–6
Pignatti S, Guarino R, La Rosa M (2017) Flora d’Italia, II ed. Edagricole, Bologna
Pyšek P, Prach K (1993) Plant invasions and the role of riparian habitats: a comparison of four species alien to central Europe. J Biogeogr 20(4):413–420. https://doi.org/10.2307/2845589
Pyšek P, Prach K (1994) How important are rivers for supporting plant invasion? In: de Waal LC, Child E, Wade PM, Brock JM (eds) Ecology and management of invasive riverside plants. Wiley, Chichester, pp 19–26
Pyšek P, Prach K, Mandak B (1998) Invasions of alien plants into habitats of central European landscape: an historical pattern. In: Starfinger U, Edwards K, Kowarik I, Williamson M (eds) Plant invasions. Ecological mechanisms and human responses. Backhuys, Leiden, pp 23–32
Pyšek P, Bacher S, Chytrý M, Jarošík V, Wild J, Celesti-Grapow L et al (2010) Contrasting patterns in the invasions of European terrestrial and freshwater habitats by alien plants, insects and vertebrates. Glob Ecol Biogeogr 19(3):317–331. https://doi.org/10.1111/j.1466-8238.2009.00514.x
Pyšek P, Hulme PE, Simberloff D, Bacher S, Blackburn TM, Carlton JT et al (2020) Scientists’ warning on invasive alien species. Biol Rev 95(6):1511–1534. https://doi.org/10.1111/brv.12627
Quaglia G, Soave P (1886) Elodea canadensis, pianta riproducentesi con immensa rapidità. Il Progresso 14:124
Quaglia G, Soave P (1889) Anacharis alsinastrum, pianta febbrifuga. Il Progresso 17:53
Riis T, Lambertini C, Olesen B, Clayton JS, Brix H, Sorrell BK (2010) Invasion strategies in clonal aquatic plants: Are phenotypic differences caused by phenotypic plasticity or local adaptation? Ann Bot 106(5):813–822. https://doi.org/10.1093/aob/mcq176
Rodríguez-Merino A, García-Murillo P, Cirujano S, Fernández-Zamudio R (2018) Predicting the risk of aquatic plant invasions in Europe: how climatic factors and anthropogenic activity influence potential species distributions. J Nat Conserv 45:58–71. https://doi.org/10.1016/j.jnc.2018.08.007
Rosati L, Fascetti S, Romano VA, Potenza G, Lapenna MR, Capano A et al (2020) New chorological data for the Italian vascular flora. Diversity 12(1):22. https://doi.org/10.3390/d12010022
Santini C, Buldrini F (2012) Le siepi del sistema Resega-Bellaria-Foresto (Novi di Modena). Atti Soc Nat Mat Modena 142:153–174
Selvaggi A, Dellavedova R (2016) Note floristiche piemontesi: 747. Elodea nuttallii (Planch.) H. St. John (Hydrocharitaceae). In: Selvaggi A, Soldano A, Pascale M, Dellavedova R (eds) Note floristiche piemontesi n. 706–773. Riv Piem Stor Nat 37:349
Simberloff D, Rejmánek M (2012) Encyclopedia of biological invasions. University of California Press, Berkeley
Simpson DA (1984) A short history of the introduction and spread of Elodea Michx. in the British Isles. Watsonia 15:1–9
Simpson DA (1988) Phenotypic plasticity of Elodea nuttallii (Planch.) H. St. John and Elodea canadensis Michx. in the British Isles. Watsonia 17:121–132
Simpson DA (1990) Displacement of Elodea canadensis Michx. by Elodea nuttallii (Planch.) H. St John in the British Isles. Watsonia 18:173–177
Spampinato G, Laface VLA, Posillipo G et al (2022) Alien flora in Calabria (Southern Italy): an updated checklist. Biol Invasions. https://doi.org/10.1007/s10530-022-02800-y
Stinca A, Chianese G, D’Auria G, Del Guacchio E, Fascetti S, Perrino EV, Rosati L, Salerno G, Santangelo A (2017) New alien vascular species for the flora of southern Italy. Webbia 72(2):295–301. https://doi.org/10.1080/00837792.2017.1349236
Stinca A, Musarella CM, Rosati L, Laface VLA, Licht W, Fanfarillo E et al (2021) Italian vascular flora: new findings, updates and exploration of floristic similarities between regions. Diversity 13(11):600. https://doi.org/10.3390/d13110600
Stoppani A (1873) Corso di geologia, II. G Bernardoni e G Brigola editori, Milano
Taffetani F (2012) Herbaria. Il grande libro degli erbari Italiani. Nardini Editore, Firenze
Thiers B (2022) Index herbariorum: a global directory of public herbaria and associated staff. New York Botanical Garden. http://sweetgum.nybg.org/ih/. Accessed 10 Mar 2022
Thomson GM (1922) The naturalisation of animals and plants in New Zealand. Cambridge University Press, Cambridge
Tolomei G (1893) Sopra l’azione della luce del magnesio sullo sviluppo dei vegetali. Le Stazioni Sperimentali Agrarie Italiane 26:377–386
Van Kleunen M, Bossdorf O, Dawson W (2018a) The ecology and evolution of alien plants. Annu Rev Ecol Evol Syst 49:25–47. https://doi.org/10.1146/annurev-ecolsys-110617-062654
Van Kleunen M, Essl F, Pergl J, Brundu G, Carboni M, Dullinger S et al (2018b) The changing role of ornamental horticulture in alien plant invasions. Biol Rev 93(3):1421–1437. https://doi.org/10.1111/brv.12402
Vanderpoorten A, Lambinon J, Tignon M (2000) Morphological and molecular evidence of the confusion between Elodea callitrichoides and E. nuttallii in Belgium and northern France. Belg J Bot 133(1–2):41–52
Verloove F (2006) Catalogue of neophytes in Belgium (1800–2005). Scr Bot Belgica 39:3–89
Viciani D, Vidali M, Gigante D, Bolpagni R, Villani M, Acosta ATR et al (2020) A first checklist of the alien-dominated vegetation in Italy. Plant Sociol 17(1):29–54. https://doi.org/10.3897/pls2020571/04
Walters SM (1980) Hydrocharitaceae. In: Tutin TG, Heywood VH, Burges NA, Moore DM, Valentine DH, Walters SM, Webb DA (eds) Flora Europaea, vol 5, pp 3–5
Wärner C, Welk E, Durka W, Wittig B, Diekmann M (2011) Biological flora of Central Europe: Euphorbia palustris L. Perspect Plant Ecol Evol Syst 13(1):55–69. https://doi.org/10.1016/j.ppees.2011.02.001
Wolff P (1980) Die Hydrilleae (Hydrocharitaceae) in Europa. Gött Flor Rundbr 14:33–56
Zanotti E (2000) Flora della pianura bresciana centro-occidentale. III Aggiornamento. «Natura Bresciana» Ann Mus Civ Sc Nat Brescia 32:225–229
Zehnsdorf A, Hussner A, Eismann F, Rönicke H, Melzer A (2015) Management options of invasive Elodea nuttallii and Elodea canadensis. Limnologica 51:110–117. https://doi.org/10.1016/j.limno.2014.12.010
Acknowledgements
We thank Antonella Albano (Università del Salento) and Carlo Argenti (Belluno) for providing us further occurrence records (both herbaria and field data).
Funding
Open access funding provided by Alma Mater Studiorum - Università di Bologna within the CRUI-CARE Agreement. The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Data collection was performed by all authors and coordinated by FB, material preparation and analysis were performed by MB, CL, GP and FB. The first draft of the manuscript was written by FB, GP and CL and all authors critically read, amended and commented the first version of the manuscript, contributing validly to data interpretation. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Buldrini, F., Pezzi, G., Barbero, M. et al. The invasion history of Elodea canadensis and E. nuttallii (Hydrocharitaceae) in Italy from herbarium accessions, field records and historical literature. Biol Invasions 25, 827–846 (2023). https://doi.org/10.1007/s10530-022-02949-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-022-02949-6