
Abstract
Fountain grass (Pennisetum setaceum) is a globally pervasive invasive species and a prime example of an escaped horticultural ornamental. In areas where it is not naturally found, it displaces native plant communities and disrupts ecological systems and processes. Cost-effective removal efforts that protect the native plant community are needed for its control. We conducted an experiment from March 2018 to March 2021 in 5 m × 5 m plots to test the efficacy and record costs for common removal techniques (cut and herbicide, herbicide one or two times per year, manual removal) in the Sonoran Desert, Arizona, United States. Each treatment took 2.5 years to achieve control in the plots, and treatments did not negatively affect the native plant community. The response of native plants was mediated by year, such that native cover in treatment plots recovered to similar levels as uninvaded control plots with sufficient rainfall. Plots that received the manual removal treatment had almost five more native plant species than the invaded control treatment (22.7 ± 1.63 compared to 18.1 ± 1.61). Herbicide applied in spring and fall increased efficacy of removals in the first year but was not significantly different from the other treatments averaged over year. Herbicide once per year was most cost effective across different sized areas. Manual removal was also cost effective in small areas (< 0.06 hectares) but was more expensive than herbicide twice a year in larger areas. Our results provide a toolset that enables managers to select removal treatments based on a balance of convenience, resources, and scale of the infestation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Invasive species are a leading threat to native plant species (Wilcove et al. 1998) and natural systems (Simberloff 2015; Pyšek et al. 2020). Invasive plants displace native plant species by competing for resources (Huenneke and Thomson 1995; Gioria and Osborne 2014), alter nutrient cycling (Vitousek et al. 1987), and increase the intensity and frequency of fire cycles (D’Antonio and Vitousek 1992; Bock and Bock 1992), among other problems. The economic cost of biological invasions in the United States is estimated at $21 billion per year, with $143 million per year spent on management (Fantle-Lepczyk et al. 2002). Horticulture is one primary pathway for invasive species introductions (Reichard and White 2001), but species have been accidentally introduced through crop seed contaminated with weed seed (Baker 1986) or from ship ballasts (Baker 1986) or intentionally introduced for soil stabilization, forage, or other purposes (Belnap et al. 2012). The invasive, non-native perennial C4 bunchgrass species fountain grass (Pennisetum setaceum (Forssk.) Chiov. or Cenchrus setaceus (Forssk.) Morrone) is an example of a horticultural species that has escaped landscaped areas and thrives across dry tropical and subtropical bioclimates worldwide. Patterns of invasion in this species are driven by anthropogenic factors and annual mean temperature (Albuquerque et al. 2020). It is regulated as a noxious weed in many countries, including several states in the U.S. (CABI 2019).
The success of P. setaceum in displacing native plants and spreading globally can partially be explained by functional traits that confer ecophysiological advantages (Gonzalez-Rodriguez et al. 2010). These traits include lower leaf area, which reduces transpiration and increases water use efficiency; lower leaf nitrogen, indicating higher nitrogen use efficiency; a higher and longer lasting photosystem (PSII, measured by the ratio of variable-to-maximal fluorescence, Gonzalez-Rodriguez et al. 2010); high reproductive output (seeds/plant) and potential (ovules/plant); fast seed germination; and rapid recovery under disturbance (Goergen and Daehler 2002). P. setaceum exhibits substantial phenotypic plasticity in response to drought (Williams and Black 1994), additional water and nutrient resources (Poulin et al. 2007; Rahlao et al. 2010), and across climatic and elevational gradients (Williams et al. 1995; Rahlao et al. 2014). In grasslands dominated by native C3 grasses, P. setaceum, with its C4 photosynthetic pathway, can compete through temporal niche partitioning by establishing later in the season once competition from C3 grasses are reduced, a dynamic that will likely be reinforced with climate change (Sweet and Holt 2015). P. setaceum even outcompetes native trees through access to shallow water resources (Cordell and Sandquist 2008).
Through the abovementioned mechanisms, P. setaceum can rapidly establish and spread within an area, leading to the accumulation of fuel loads and increase in the frequency and severity of wildfire (CABI 2019). The phenomenon of invasive grasses promoting increased fire cycles and severity is a global problem (D’Antonio and Vitousek 1992; Brooks et al. 2004). P. setaceum seeds buried at depths of 2.5–5 cm can escape the high heat of a fire and establish because they do not require light for germination (Adkins et al. 2011). In contrast, native plants may not readily return after fire, especially in systems that are not fire adapted, leading to invasive species dominated systems (D’Antonio and Vitousek 1992; Rahlao et al. 2009).
Because of its rapid spread and regrowth, control of P. setaceum can be difficult. Common control practices include manual removal and herbicide application. In Hawaii, a common practice is to cut P. setaceum during the active growth phase and wait for the grass to resprout before applying herbicide (Cordell et al. 2002). This cut and herbicide approach has been compared with bulldozing and cutting alone but not to manual removal or herbicide without cutting (Cabin et al. 2002; Thaxton et al. 2012). We were interested in comparing the costs, efficacy, and native plant community response of common control practices, including herbicide only compared to cut and herbicide, different herbicide application frequencies, and manual removal with a pick. We asked the following research questions: (1) Which technique is the most effective for controlling P. setaceum and reducing fuel load over the long-term: manual removal, cut and spray, herbicide once a year, or herbicide twice per year? (2) Which treatment is most cost effective over the long term, considering the number of years until control and size of invaded area? (3) Which treatments promote native plant community richness and cover?
Methods
Study site
The McDowell Sonoran Preserve (henceforth Preserve) encompasses 12,375 ha of Sonoran Desert Upland habitat (Brown et al. 1979) at the northeastern edge of the Phoenix metropolitan area in Scottsdale, Arizona, U.S.A. Our study site was in Quartz Wash, in the southern region of the Preserve (between 33.63120, −111.80801 and 33.63319, −111.80692) at elevations between 760 and 766 m in recent (< 10,000 years old) alluvial deposits (Skotnicki 2016). The soil type is Schist Hills 10″–13″ p.z. (R040XA119AZ), part of the Dixileta–Rock Outcrop Complex (AZ645), which is a well-drained, gravelly sandy loam, with a relatively shallow soil depth (USDA NRCS 2021). The plant community is a mixture of Simmondsia chinensis—mixed scrub association (154.123) and Ambrosia deltoidea—Parkinsonia microphylla—mixed scrub association (154.121) (Jones and Hull 2014).
During this study, March 2018–March 2021, the lowest monthly mean temperature was 4.3 °C in February 2019 and a maximum monthly mean temperature of 40.5 °C in August 2020, and the annual mean temperatures ranged from 20.8 to 22.2 °C (PRISM Climate Group 2021). Rainfall varied over the course of the study with significantly below-average rainfall in 2018 and above-average rainfall in 2019 and the winter of 2020 (Online Resource 1). The Sonoran Desert’s rainy seasons occur in the winter (October–March) and in the summer (June–September) with winter rains typically delivering more overall precipitation (but see 2018, Online Resource 1).
Study design and treatments
Using a randomized complete block design, we established 36 5 m × 5 m plots in an area that was heavily invaded with P. setaceum. Thirty of these plots had a minimum of 30% P. setaceum cover (ranged from 33 to 79%) and were used as treatment and invaded control plots. The remaining six were not invaded with P. setaceum and were used as uninvaded controls. We grouped the plots into six blocks of five invaded plots and one uninvaded plot based on relative positions within the wash, such that five invaded and one uninvaded plots located lowest in the wash were one block, the second set of five invaded and one uninvaded plots were in a second block, etc. We randomly assigned treatments to the invaded plots within each block, such that each block contained one of each type of the four removal treatments and one uninvaded and one invaded control plot for comparison purposes.
The four removal treatments were manual removal, cut and herbicide, and herbicide application either one or two times per year. Manual removal treatments were applied by digging up the plant with as much of the roots as possible using a Pick Mattock and laying the plant over the dug-up area. This method of thatching plants rather than removing plants from the site can suppress seedling growth (Jernigan et al. 2016). For the cut treatment, we used a gas-powered weed whacker (Stihl FS56RC) with 10.5 cm plastic blades to cut the grasses as close to the ground as possible given the uneven terrain, between 20 and 30 cm in height, in Spring 2018. Grass cuttings were left in place. The goal of the cut treatment was to remove inert plant material so that herbicide could be applied to the re-growth later in the same year (Cordell et al. 2002). We did not apply the cut treatment in subsequent years because there were few individual plants left in the plots. For herbicide treatments, a certified herbicide applicator (Pest Management Division, Arizona Department of Agriculture) applied a mixture of 3% glyphosate, N-(phosphonomethyl) glycine on an active ingredient basis, in the form of its isopropylamine salt with 13% surfactant (Roundup Pro, Monsanto) with a blue dye (except where noted), directly to target plant leaves using a low volume, single nozzle sprayer (Matabi Berry 7). We chose glyphosate due to its proven effectiveness on P. setaceum (Voigt and Reicher 2009; Thomas and Taylor 2021), lower cost (Cordell et al. 2002), and relatively fewer negative environmental impacts (Duke 2020). We applied herbicide once plants were at least 50% green either in the spring only (herbicide 1x) or in spring and fall (herbicide 2x). However, in Spring 2018, plants were < 10% green due to drought, so we only treated the herbicide 2x plots in the spring and both herbicide 1x and 2x in the fall. Herbicide treatments for the cut and herbicide treatment were also applied in fall the first year and once in the spring, thereafter.
To reduce edge effects, we established 1 m buffers around each plot in which P. setaceum was removed using the same treatment as the plot. We removed P. setaceum in areas outside of the buffers to reduce the propagule pressure on the plots. Initially, the invaded and uninvaded control plots received no treatments. However, in the third year, we recognized that seed rain from P. setaceum in the invaded control plots was likely re-invading the treatment plots. Because this could confound results as to which technique was the most effective for controlling P. setaceum, especially in plots nearest the invaded controls, we elected to treat the invaded control plots. In 2019, we removed seed heads from invaded control plots to minimize seed spread and re-invasion of the plots, but the seed heads grew back within the same season. In 2020, after spring sampling, we applied either the manual removal or herbicide treatment to invaded control plots, such that half of the plots received each treatment. We intended to monitor these plots over time, but the plot markers were washed away with a flooding event in Summer 2021.
Plant sampling
To assess how treatments affected the native plant community, we collected data on plant and litter cover in each plot in March of each year during peak spring annual biomass. Plant sampling occurred prior to treatment application, such that the 2018 sampling established a pre-treatment baseline and the 2019 sampling followed our first year of treatments. We used a line point intercept method along five parallel 5 m transects placed at 0.5 m, 1.5 m, 2.5 m, 3.5 m, and 4.5 m at 1.2 m height across the plot. On each transect, 20 intercept points were sampled 25 cm apart for a total of 100 points per plot. At each intercept, using a dropped pin or laser, we recorded each plant species above or below the line, including standing dead P. setaceum (still rooted). To document species richness, we then searched within the plot and documented all plant species in addition to those recorded on the line intercepts. After the plot was sampled, we pulled non-native species listed on the Preserve invasive species list (Oncosiphon piluliferum and Brassica tournefortii), which composed 2.5% of the overall nonfocal non-native cover.
Treatment application and cost calculations
Spring treatment applications occurred in March–April, following plant sampling, and fall treatments occurred in September–October after monsoon rains ended. To ensure that plants were not missed, at least two observers first surveyed the plot and marked all P. setaceum plants with pin flags. Plots were surveyed again immediately after treatment and any missed plants were treated. We documented how long it took to apply the treatment and amount of herbicide solution used within each plot. In 2019, we also began counting the number of plants treated within each plot. Within three weeks of treatment, we checked the efficacy of removal treatments and documented any new growth or missed plants. In Spring 2019, we re-treated all treatment plots due to the high occurrence of living P. setaceum observed during the post-treatment check; we are not sure if these plants were missed during the initial treatments or were new growth from above-average rainfall.
We calculated costs of each treatment based on measured costs (time to apply the treatment in plots and supplies used) and estimated costs (preparation time). For measured costs, we summed the timed per-person treatment application effort in each plot. Supplies included herbicide and weed whacker blades. For herbicide plots, we measured the herbicide quantity used by marking lines on the sprayer tank before and after the treatment and calculating the volume (in liters) of solution used. We then multiplied that number by 3% to obtain the amount of actual herbicide used and multiplied the result by the cost of the herbicide ($11.66 per liter). We calculated the per plot cost of blades for the weed whacker in 2018 by dividing the total number of blades used by the number of cut and herbicide plots and multiplying by the blade cost [(29 blades/6 plots) * $0.72/blade = $3.48/plot]. We did not include the cost of equipment such as the sprayer tanks, personal protective equipment, or picks.
In addition to the measured costs, we added estimated costs of “per plot” and “per day” preparation time as they are important considerations for managers when comparing treatments. Per plot costs were dependent on number of treatments applied and number of plants treated in each plot and included changing blades for the cut treatment (estimated at 0.15 h/plot) and time to fill water bottles and refill the sprayer for herbicide treatments (estimated at 0.01 h for < 10 plants per plot; 0.02 h for 10–20 plants per plot; and 0.03 h for > 20 plants per plot). The “per day” costs were fixed costs that did not change with number of treatments or plants and included the preparation time in gathering supplies, cleaning the sprayer, and putting on PPE (0.42 h/day) and round-trip travel time (2 h/day). All measured and estimated time was converted to cost by multiplying by the national average herbicide applicators rate of $17.60/hour (Payscale 2021).
To calculate the overall costs per treatment, we added together calculated costs and in-plot estimated costs of all plots within the same treatment for each season within each year. In this calculation, if there were no measured costs (no plants were treated) in given plot, then per plot estimates were also not included for that plot. Per day estimated costs were multiplied by the number of treatments per season/year and added to the calculated and in-plot estimates total.
Analysis
To compare treatment efficacy, we analyzed P. setaceum cover and sum of plants counted prior to every removal in each plot within season. We analyzed the P. setaceum cover data using a mixed model in a full factorial with treatment and year as fixed factors (6 levels: cut and herbicide, herbicide 1x, herbicide 2x, manual removal, uninvaded control, and invaded control) and block and plot nested in block as the random factors. The 2021 P. setaceum cover (post removal) for the invaded control treatment was not included in analysis. Random effects were added to the model to account for the randomized complete block design (block) and repeated measures (plot nested in block). The count data showed a skewed distribution; therefore, we used a poisson model with a link “log” function with treatment (5 levels: cut and herbicide, herbicide 1x, herbicide 2x, manual removal, and uninvaded control) as the fixed factor, number of plants as the response variable, and block and plot nested in block as the random factors. We only counted the number of individuals in the invaded control plots in 2020, before removal; therefore, this treatment was not included in the analysis. Thus, the cover data provided the overall analysis including changes from baseline and comparisons with the invaded control, and the count data served to compare efficacy among treatments. We used the glmer function in the lme4 package (Bates et al. 2015) in R (R Core Team 2021). For a posthoc comparison of means, we used the sidak adjustment using the emmeans package in R (Lenth 2021). All figures were created in JMP version 16.0.
We analyzed effects of the removal treatments on the native plant community cover and richness using a mixed model in a full factorial with treatment and year as fixed factors (6 levels: cut and herbicide, herbicide 1x, herbicide 2x, manual removal, uninvaded control, and invaded control) and block and plot nested in block as the random factors. Response variables were native total, perennial, and annual richness and cover, and nonfocal non-native richness and cover (without P. setaceum). We analyzed the effect of removal treatments on fuel load (litter and standing dead) using the same model with five levels of treatment (cut and herbicide, herbicide 1x, herbicide 2x, manual removal, and uninvaded control). We used the lmer function in the lme4 package (Bates et al. 2015) and the emmeans package (Lenth 2021) in R (R Core Team 2021). Residuals were checked and native annual cover was log + 1 transformed and total native, P. setaceum cover, and non-native cover were square root transformed to meet assumptions of normality.
We analyzed measured costs using a mixed model with treatment as a fixed factor (4 levels: cut and herbicide, herbicide 1x, herbicide 2x, and manual removal) and block as a random factor using the same statistical packages as in the native plant community models described above. We used two response factors: (1) measured per plot application time summed by treatment over the duration of the experiment and (2) the measured per plot application time multiplied by the national average herbicide applicators rate of $17.60/h (Payscale 2021) plus the cost of supplies summed by treatment. Residuals were checked and measured costs were square root transformed to meet assumptions of normality.
In order to estimate realistic costs associated with each treatment, we added the estimated costs to the measured costs and then scaled the total costs for treating different sized areas to compare how the treatment costs compare at different scales. These were not compared statistically, due to the inclusion of estimates (without standard errors).
Results
Treatment efficacy
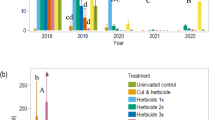
By Fall 2020, two and a half years after treatments began, no living P. setaceum plants were observed in any of the treatment plots. We verified this trend continued in Spring 2021. The P. setaceum cover had a year by treatment interaction effect, such that the removal treatments reduced P. setaceum in the first year, but the invaded control remained high until after plant sampling in 2020 (Table 1, Fig. 1a).The average number of P. setaceum plants was greater in the cut and herbicide, herbicide 1x, and manual removal plots than in the uninvaded control plots, whereas there was not a significant difference between the herbicide 2x and uninvaded control plots (χ2 = 18.255, df = 4, p = 0.001, Fig. 1b).

Effects of removal treatments on P. setaceum a plant cover and b number of plants removed. Counts were taken prior to subsequent treatment applications each year starting one year after removals began. Cover data for invaded control is not included for 2021 because P. setaceum was treated in the plots after sampling in 2020. Bars represent one standard error of the mean. Asterisks indicate a year*treatment interaction effect at this p level: ***p < 0.001. Lower case letters represent significant differences among treatments, averaged over season, using sidak post hoc comparisons of means. Plots are 5 m × 5 m
Effect on plant community
Native cover
Total and perennial native plant cover had a significant year by treatment interaction in which treatment differences were mediated by year (Table 1, Fig. 2). The interaction can be seen in 2018 when, prior to initial treatments, total and perennial native cover were higher in the uninvaded control plots compared with the invaded control plots, whereas these treatment differences were reduced in subsequent years with the removal of P. setaceum and more rainfall (Fig. 2). For annual native cover there were no significant interactions or treatment differences, only differences in year (Table 1, Fig. 2), likely driven by precipitation patterns (Table S1).

Response of native and nonfocal non-native plant cover to removal treatments over time. Upper case letters represent differences among year, averaged over treatment. Treatment differences were calculated using sidak post hoc comparisons of means. Asterisks indicate a year * treatment interaction effect at these p levels: *p < 0.05. and **p < 0.01. Bars represent one standard error of the mean
Native richness
Total native plant richness had significant annual differences (Table 1, Fig. 3). Averaged over years, the manual removal treatment had higher total native richness (22.7 ± 1.63) than the invaded control (18.1 ± 1.61), but no difference was observed between the other treatments and the invaded or uninvaded controls (Fig. 3). Treatment differences for perennial native richness depended on year (Table 1). The interaction can be seen in 2020, in which the invaded control had lower perennial native richness than the treatments, compared with the following year when the perennial native richness was depressed in the herbicide 2x treatment compared with the manual removal treatment (Fig. 3). Annual native richness also had significant annual differences, reflecting the same trend as total native richness, and had a significant main treatment effect (Table 1), but these differences were not significantly separated in posthoc analysis (Fig. 3).

Response of native and nonfocal non-native plant richness to removal treatments over time. Upper case letters represent differences among year, averaged over treatment. Treatment differences were calculated using sidak post hoc comparisons of means. An asterisk indicates a year * treatment interaction effect at p < 0.05. Bars represent one standard error of the mean
Non-native cover and richness
Nonfocal non-native cover and richness differed by year but not treatment (Table 1, Figs. 3, 4). Cover increased in the two years after the first P. setaceum removal, then decreased in 2021, a drier year (Table 1 and Fig. 3). Nonfocal non-native species present included annual grasses (Bromus rubens, Schismus barbatus, Hordeum murinum) and annual forbs (Oncosiphon piluliferum, Erodium cicutarium, Herniaria hirsute, Sonchus oleraceus, Sisymbrium irio, Brassica tournefortii, Malva parviflora, Lactuca serriola). Non-native species richness was higher in the two wetter years, as seen with native plant cover and richness. P. setaceum standing dead had a significant interaction in which there were no differences among treatments in 2018, prior to removals, but in 2019 the herbicide treatments (herbicide 1x, herbicide 2x, cut and herbicide) were higher than manual removal and the uninvaded control (Table 1, Fig. 4). Litter cover was different among years but not treatments (Table 1, Fig. 4).

Pennisetum setaceum standing dead and litter cover by treatment over time. Upper case letters represent differences among year, averaged over treatment. Three asterisks indicates a year * treatment interaction effect at p < 0.001. Bars represent one standard error of the mean
Cost effectiveness
Manual removal treatments took significantly more time to apply than the other treatments, and cut and herbicide took more time to apply than the herbicide 1x and 2x treatments (χ2 = 65.014, df = 3, p < 0.001, Fig. 5a). Manual removal and cut and herbicide treatments had significantly higher measured costs than the herbicide only plots (χ2 = 107.01, df = 3, p < 0.001, Fig. 5b). However, when accounting for travel and preparation time prior to and in field and supply costs, at the plot scale (25 m2), manual removal was cheapest and herbicide 2x was the most expensive (Fig. 5c). Herbicide 1x and manual removal had comparable costs at small scales (< 0.1 ha), but herbicide 1x was the least expensive treatment for larger areas. Manual removal was less expensive than herbicide 2x for areas < 0.6 ha but was more expensive in larger areas (Fig. 5c).

Costs of removal treatments by a mean time conducting treatments in 25 m2 plot, b mean measured cost of labor and supplies in 25 m2 plot, and c total estimated costs. Total estimated costs include measured costs (based on cost of labor and supplies in plot) plus estimated preparation time (labor per plot and per day) and two-hour round-trip travel to the site. The x and y axes for plot c are on a logarithmic scale. Area of plots was 25 m2. Lower case letters represent differences among treatments averaged over year using sidak post hoc comparisons of means. Bars represent one standard error of the mean
Discussion
Our study is the first to provide a comprehensive comparison of common control techniques for P. setaceum that also addresses responses of the native plant community and cost effectiveness. All treatments we compared effectively controlled P. setaceum within three years, and none had a negative influence on the native plant community.
Overall, applying herbicide once per year was the most cost-effective treatment. However, it is important to note that in very small areas (< 0.04 hectare) manual removal was the most cost effective; even though it took the most amount of time to apply, no preparation time was required. Conversely, herbicide twice a year required substantial preparation time but less application time; therefore, in larger areas where more time is spent in application vs preparation, it replaced manual removal as the second most cost-effective treatment. Applying herbicide twice a year also appeared to increase efficacy of P. setaceum removal in the first year, which might argue for applying herbicide multiple times at first and then switching to less intensive approaches in subsequent years. Other studies have also had success from applying multiple treatments per year and implementing different types of treatments. In Hawaii, Cabin et al. (2000) removed P. setaceum using a weedwhacker and multiple applications of herbicide in the first year and subsequent herbicide applications and manual removal in following years with rodent control and ungulate exclosures to promote native plant establishment. Cabin et al. (2002) also found that all their treatments (bulldoze, plastic mulch, and cut and herbicide) helped control P. setaceum better than cutting alone.
In our study, the cut and herbicide treatment was the costliest treatment at scales larger than 0.05 hectare due to the supplies used (weedwhacker blades), plus the amount of time required to replace the blades due to breakage. In our first year, we found that weedwhacker string was ineffective as it broke and had to be re-threaded more often. Additionally, we found no evidence that cutting improved the effectiveness of the cutting treatment compared to herbicide alone. This finding was surprising given that previous experiments performed in Hawaii had high success with this treatment, although cut and herbicide was not directly compared to herbicide alone in those studies (Cabin et al. 2000, 2002; Cordell et al. 2002). The rationale for cutting the P. setaceum prior to herbicide at the Hawaiian sites was likely to reduce the abundance of inert plant material from previous years to focus herbicide use only on new growth. In Hawaii, rainfall occurs throughout the year and P. setaceum is rarely dormant and likely occurs in denser stands than in Arizona, thus the cut, re-growth, and herbicide can be applied on a continuous cycle, instead of a seasonal one. These differences may make this treatment more useful in Hawaii; however, concentrating the herbicide spray on the green parts of the grasses met with success in our experiment.
In addition to cost effectiveness, managers need to consider logistical constraints of workforce, material transport to the site, timing, environmental costs, and remaining fuel load (Table 2). Herbicide treatments tend to leave more flammable material in place with standing dead P. setaceum. However, we did not see a comparable increase in litter in the manual removal plots as expected, perhaps as a result of heavy rain events that moved the manually removed plants off the plots. Also, although glyphosate is relatively benign compared with other herbicides, due to strong soil binding and water solubility (Duke 2020), it should not be applied directly to water or where there is high risk of drift to water due to reported impacts on amphibians (Relyea 2005, 2012) and, if applied broadly, can have non-target effects on native plants because it is a non-selective herbicide. Herbicide also requires a substantial amount of water, which can be difficult to transport long distances to inaccessible areas (Table 2).
Although we found that time until control could be achieved within three years, this might fluctuate to some degree depending on precipitation during the spanning years. Precipitation levels can present a trade-off in terms of invasive species spread and control. While a rainy year will nurture more P. setaceum seedlings, growth, and seed (Goergen and Daehler 2002; Poulin et al. 2007), it also provides more opportunity for herbicide application because glyphosate and many other postemergence herbicides rely on phloem transport to the roots and are not effective on dormant plants (Hanson 2015). In drought conditions, herbicide can be less effective due to slowed plant processes or physical changes to the plant leaf surfaces; however, these obstacles can be mitigated through use of surfactants (Hanson 2015). In seasons with limited moisture or during drought, manual removal might be the more effective option for managers, as dormant plants can be effectively removed.
We had anticipated that the manual removal treatment, given the amount of soil disturbance caused by digging out roots, might negatively affect the plant community but, by the final year, the manual removal treatment had higher native perennial plant species richness than the herbicide 2x treatment (interaction effect). It is possible that perennials might have benefitted from the effects of soil disturbance. Researchers have found that manually removing invasive grasses can change the soil microbial community (Williams et al. 2021), and the digging action can increase water infiltration, incorporate organic matter into the soil, and increase plant establishment and growth (Kinyua et al. 2010; Ruthrof et al. 2013; Rowe et al. 2021). Other studies have seen similar effects from bulldozing areas to remove P. setaceum, which improved soil moisture and native seedling performance compared to cut and herbicide treatments (Cabin et al. 2002; Thaxton et al. 2012), although disturbing areas can result in higher occurrence of other non-native species (Hobbs and Huenneke 1992; Rowe et al. 2021).
Although we had expected to see negative treatment effects on the native plant community, any that occurred may have been balanced by removing competition from P. setaceum. In this study, the native plant community cover and richness increased in 2019, the year after treatments, but 2019 was also a relatively wet year compared to 2018. Other studies observed positive influences on native species from removal of P. setaceum, particularly for establishment of desired species. Cabin et al. (2000) found that, combined with exclusion of rodents, controlling P. setaceum allowed for the establishment of Diospyros seedlings, a key native dry forest species, as well as herbaceous seedlings. In a subsequent study, Cabin et al. (2002) tested seeding and planting target species in combination with P. setaceum control and found species-specific responses. Thaxton et al. (2012) compared the effects of removing fountain grass, shade (60% of full sun, full sun), and water (supplemental vs ambient) on transplanting seedling survival, growth and physiology of five native trees, three native shrubs, and two vines. They found that each treatment independently increased survival and growth of the seedlings (Thaxton et al. 2012).
As with the native plants, nonfocal non-native cover and richness also increased after the removals began. This is a fairly common phenomenon; in a survey of studies encompassing 35 national parks, 44% of the 16 studies that evaluated nonfocal exotic species found an increase (Abella 2014). Fortunately, the nonfocal non-native species in our study generally have lower biomass and, thus, the overall fire load and risk is lower with P. setaceum removal even with the associated influx of non-native species.
Conclusion
Manually removing, cutting followed by herbicide treatment, and herbicide treatment alone were equally effective in controlling P. setaceum while protecting the native plant community. A sustained, multi-year effort is required, and managers should expect to apply removal treatments for at least three years. In our study, removals were undertaken with a level of effort unlikely to be realized in a regular field situation (e.g., flagging plants, checking and re-treating after removal, and re-treating in spring 2019 to account for missed plants or new sprouts); therefore, this estimated timeframe likely represents the best-case scenario and it may take longer to achieve control. Continued monitoring and retreatments in subsequent years are recommended. In areas of small infestation, managers can consider manual removal as a cost-effective technique, replacing that with herbicide for larger infestations. Depending on environmental conditions and green-up of the plants, multiple herbicide treatments in the first year could be considered to help speed up control efforts. These findings are especially useful for helping managers to set expectations and plan for their overall efforts. The appropriate treatment(s) to use will depend on each individual scenario, including the location and extent of infestation, available resources, staffing and volunteer workforce, and environmental conditions. Managers can use our findings as a toolset to determine what will work best for them.
Data availability
The data generated and analyzed during the study are available on GitHub (https://HelenIvy.github.io/FountainGrass/).
References
Abella SR (2014) Effectiveness of exotic plant treatments on National Park Service lands in the United States. Invasive Plant Sci Manag 7:147–163. https://doi.org/10.1614/IPSM-D-13-00058.1
Adkins E, Cordell S, Drake DR (2011) Role of fire in the germination ecology of fountain grass (Pennisetum setaceum), an invasive African bunchgrass in Hawaii. Pac Sci 65:17–25. https://doi.org/10.2984/65.1.017
Albuquerque F, Macías-Rodríguez MÁ, Búrquez A, Rowe HI (2020) Toward an understanding of broad-scale patterns of the habitat suitability of fountain grass (Cenchrus setaceus (Forssk.) Morrone, Poaceae). Plant Ecol 221:1029–1043. https://doi.org/10.1007/s11258-020-01060-x
Baker H (1986) Patterns of plant invasion in North America. In: Mooney H, Drake J (eds) Ecology of biological invasions in North America and Hawaii. Springer-Verlag, New York, pp 44–57
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Belnap J, Ludwig JA, Wilcox BP et al (2012) Introduced and invasive species in novel rangeland ecosystems: friends or foes? Rangel Ecol Manag 65:569–578. https://doi.org/10.2111/REM-D-11-00157.1
Bock JH, Bock CE (1992) Vegetation responses to wildfire in native versus exotic arizona grassland. J Veg Sci 3:439–446. https://doi.org/10.2307/3235800
Brooks ML, D’Antonio CM, Richardson DM, Grace JB, Keeley JE, DiTomaso JM, Hobbs RJ, Pellant M, Pyke D (2004) Effects of invasive alien plants on fire regimes. Bioscience 54:677–688. https://doi.org/10.1641/0006-3568(2004)054[0677:EOIAPO]2.0.CO;2
Brown D, Love C, Pase C (1979) A digitized classification system for the biotic communities of North America, with community (series) and association examples for the Southwest. Arizona-Nevada Acad Sci 14:1–16
CABI (2019) Pennisetum setaceum (fountain grass, original text by: Sylvan Kaufman). In: Invasive species compendium. CAB International, Wallingford. www.cabi.org/isc
Cabin RJ, Weller SG, Lorence DH, Flynn TW, Sakai AK, Sandquist D, Hadway LJ (2000) Effects of long-term ungulate exclusion and recent alien species control on the preservation and restoration of a Hawaiian tropical dry forest. Conserv Biol 14:439–453. https://doi.org/10.1046/j.1523-1739.2000.99006.x
Cabin RJ, Weller SG, Lorence DH, Cordell S, Hadway LJ, Montgomery R, Goo D, Urakami A (2002) Effects of light, alien grass, and native species additions on Hawaiian dry forest restoration. Ecol Appl 12:1595–1610. https://doi.org/10.1890/1051-0761(2002)012[1595:EOLAGA]2.0.CO;2
Cordell S, Sandquist DRR (2008) The impact of an invasive African bunchgrass (Pennisetum setaceum) on water availability and productivity of canopy trees within a tropical dry forest in Hawaii. Funct Ecol 22:1008–1017. https://doi.org/10.1111/j.1365-2435.2008.01471.x
Cordell S, Cabin RJ, Weller SG, Lorence DH (2002) Simple and cost-effective methods control fountain grass in dry forests (Hawaii). Ecol Restor 20:139–140
D’Antonio CM, Vitousek PM (1992) Biological invasion by invasive grasses, the grass/fire cycle, and global change. Annu Rev Ecol Syst 23:63–87
Duke SO (2020) Glyphosate: environmental fate and impact. Weed Sci 68:201–207. https://doi.org/10.1017/wsc.2019.28
Fantle-Lepczyk JE, Haubrock PJ, Kramer AM et al (2022) Economic costs of biological invasions in the United States. Sci Total Environ 806:151318. https://doi.org/10.1016/J.SCITOTENV.2021.151318
Gioria M, Osborne BA (2014) Resource competition in plant invasions: emerging patterns and research needs. Front Plant Sci 5
Goergen E, Daehler CC (2002) Factors affecting seedling recruitment in an invasive grass (Pennisetum setaceum) and a native grass (Heteropogon contortus) in the Hawaiian Islands. Plant Ecol 161:147–156. https://doi.org/10.1016/J.SCITOTENV.2021.151318
Gonzalez-Rodriguez AM, Baruch Z, Palomo D, Cruz-Trujillo G, Soledad Jimenez M, Morales D (2010) Ecophysiology of the invader Pennisetum setaceum and three native grasses in the Canary Islands. Acta Oecol 36:248–254. https://doi.org/10.1016/j.actao.2010.01.004
Hanson B (2015) Effects of drought conditions on weed control performance and herbicide fate. Cooperative Extension Weed Specialist, Plant Sci Dept, Davis, California
Hobbs RJ, Huenneke LF (1992) Disturbance, diversity, and invasion: implications for conservation. Conserv Biol 6:324–337. https://doi.org/10.1046/j.1523-1739.1992.06030324.x
Huenneke LF, Thomson JK (1995) Potential interference between a threatened endemic thistle and an invasive nonnative plant. Conserv Biol 9:416–425. https://doi.org/10.1046/j.1523-1739.1995.9020416.x
Jernigan MB, McClaran MP, Biedenbender SH, Fehmi JS (2016) Uprooted buffelgrass thatch reduces buffelgrass seedling establishment. Arid Land Res Manag 30:320–329. https://doi.org/10.1080/15324982.2015.1107152
Jones S, Hull C (2014) Vegetation and flora of the McDowell Sonoran Preserve, Maricopa County, Arizona. Canotia 10:1–34
Kinyua D, McGeoch LE, Georgiadis N, Young TP (2010) Short-term and long-term effects of soil ripping, seeding, and fertilization on the restoration of a tropical rangeland. Restor Ecol 18:226–233. https://doi.org/10.1111/j.1526-100X.2009.00594.x
Lenth RV (2021) emmeans: estimated marginal means, aka least-squares means. R package version 1.6.3. https://CRAN.R-project.org/package=emmeans
Payscale (2021) Hourly rate for certification: certified pesticide applicator (TCEQ). https://www.payscale.com/research/US/Certification=Certified_Pesticide_Applicator_(TCEQ)/Hourly_Rate. Accessed 11 Nov 2021
Poulin J, Sakai A, Weller SG, Nguyen T (2007) Phenotypic plasticity, precipitation, and invasiveness in the fire-promoting grass Pennisetum setaceum. Am J Bot 94:533–541. https://doi.org/10.3732/ajb.94.4.533
PRISM Climate Group (2021) PRISM Climate Group. http://prism.oregonstate.edu. Accessed 11 May 2021
Pyšek P, Hulme PE, Simberloff D et al (2020) Scientists’ warning on invasive alien species. Biol Rev 95:1511–1534. https://doi.org/10.1111/brv.12627
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rahlao SJ, Milton SJ, Esler KJ, Van Wilgen BW, Barnard P (2009) Effects of invasion of fire-free arid shrublands by a fire-promoting invasive alien grass (Pennisetum setaceum) in South Africa. Austral Ecol 34:920–928. https://doi.org/10.1111/j.1442-9993.2009.02000.x
Rahlao SJ, Esler KJ, Milton SJ, Barnard P (2010) Nutrient addition and moisture promote the invasiveness of crimson fountaingrass (Pennisetum setaceum). Weed Sci 58:154–159. https://doi.org/10.1614/WS-09-091.1
Rahlao SJ, Milton SJ, Esler KJ, Barnard P (2014) Performance of invasive alien fountain grass (Pennisetum setaceum) along a climatic gradient through three South African biomes. S Afr J Bot 91:43–48. https://doi.org/10.1016/j.sajb.2013.11.013
Reichard SH, White P (2001) Horticulture as a pathway of invasive plant introductions in the United States. Bioscience 51:103–113. https://doi.org/10.1641/0006-3568(2001)051[0103:HAAPOI]2.0.CO;2
Relyea RA (2005) The lethal impact of Roundup on aquatic and terrestrial amphibians. Ecol Appl 15:1118–1124. https://doi.org/10.1890/04-1291
Relyea RA (2012) New effects of Roundup on amphibians: predators reduce herbicide mortality; herbicides induce antipredator morphology. Ecol Appl 22:634–647. https://doi.org/10.1890/11-0189.1
Rowe HI, Sprague TA, Ball B, Langenfeld D, Rivera L (2021) Restoring closed trails in the Sonoran Desert: interactions of seed timing, seed source, and ripping. Restor Ecol. https://doi.org/10.1111/rec.13532
Ruthrof KX, Fontaine JB, Buizer M, Matusick G, McHenry MP, St J Hardy GE (2013) Linking restoration outcomes with mechanism: the role of site preparation, fertilisation and revegetation timing relative to soil density and water content. Plant Ecol 214:987–998. https://doi.org/10.1007/s11258-013-0224-8
Simberloff D (2015) Non-native invasive species and novel ecosystems. F1000Prime Rep 7:1–7. https://doi.org/10.12703/P7-47
Skotnicki SJ (2016) Geology of the McDowell Mountains. Arizona Geological Survey contributed map. http://tinyurl.com/GeoMap-McdowellMtns. Accessed 23 Nov 2021
Sweet LC, Holt JS (2015) Establishment stage competition between exotic crimson fountaingrass (Pennisetum setaceum, C4) and native purple needlegrass (Stipa pulchra, C3). Weed Sci Soc Am 8:139–150. https://doi.org/10.1614/IPSM-D-14-00048.1
Thaxton JM, Cordell S, Cabin RJ, Sandquist DR (2012) Non-native grass removal and shade increase soil moisture and seedling performance during Hawaiian dry forest restoration. Restor Ecol 20:475–482. https://doi.org/10.1111/j.1526-100X.2011.00793.x
Thomas J, Taylor M (2021) Evaluation of chemical control methods of fountain grass. HortTechnology 31:382–384. https://doi.org/10.21273/HORTTECH04789-20
USDA NRCS (2021) Web soil survey. https://websoilsurvey.sc.egov.usda.gov/App/HomePage.htm?TARGET_APP=https://edit.jornada.nmsu.edu/catalogs/esd/040X/R040XA119AZ. Accessed 23 Nov 2021
Vitousek PM, Walker LR, Whiteaker LD et al (1987) Biological invasion by myrica faya alters ecosystem development in Hawaii. Science (80-) 238:802–804. https://doi.org/10.1126/SCIENCE.238.4828.802. Accessed 23 Nov 2021
Voigt TB, Reicher ZJ (2009) Selectively controlling escaped fountain grass in cool-season turf. Appl Turfgrass Sci 6:1–7. https://doi.org/10.1094/ats-2009-0720-02-rs
Wilcove DS, Rothstein D, Dubow J et al (1998) Quantifying threats to imperiled species in the United States
Williams DG, Black RA (1994) Drought response of a native and introduced Hawaiian grass. Oecologia 97:512–519. https://doi.org/10.1007/BF00325890
Williams DG, Mack RN, Black RA (1995) Ecophysiology of introduced pennisetum setaceum on Hawaii: the role of phenotypic plasticity. Ecology 76:1569–1580. https://doi.org/10.2307/1938158
Williams JP, Gornish ES, Barberan A (2021) Effects of buffelgrass removal and nitrogen addition on soil microbial communities during an extreme drought in the Sonoran Desert. Restor Ecol. https://doi.org/10.1111/rec.13570
Acknowledgements
We thank the countless citizen scientists at the McDowell Sonoran Conservancy for their time and effort on this project, including Barb Pringle, Gary Binder, and Jeff Lund and to Melanie Tluczek for early contributions to the project. We also thank Steve Jones, Danika Setaro, JP Solves, Natalie Melkonoff, Steve Blackwell, and Zachary Berry for providing their botany expertise. We are grateful to the editor, Dr. Lieurance, and two anonymous reviewers for their comments that helped improve this manuscript. We acknowledge the City of Scottsdale for funding the project and contributing to applying treatments.
Funding
This work was supported by the City of Scottsdale (#18-RP-009).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design, material preparation, and data collection. TS managed data and protocols. HR analyzed data and wrote the first draft of the manuscript. All authors edited and commented on prior versions of the manuscript and read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rowe, H.I., Sprague, T.A. & Staker, P. Comparing common fountain grass removal techniques: cost efficacy and response of native plant community. Biol Invasions 24, 3817–3830 (2022). https://doi.org/10.1007/s10530-022-02879-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-022-02879-3