
Abstract
A large knowledge gap exists regarding the disease profile and pathologic condition of the invasive, slipper limpet Crepidula fornicata. To help address this, we performed a year-long health survey at two sites in South Wales, UK – subtidal Swansea Bay and intertidal Milford Haven. In total, 1,800 limpets were screened systematically for haemolymph bacterial burdens using both general and vibrio-selective growth media (TSA + 2% NaCl and TCBS, respectively), haemolymph (blood) inspection using microscopy, a PCR-based assay targeting Vibrio spp., and multi-tissue histology. Over 99% of haemolymph samples contained cultivable bacterial colony-forming units, and 83% of limpets tested positive for the presence of vibrios via PCR (confirmed via Sanger sequencing). Vibrio presence did not vary greatly between sites, yet a strong seasonal effect was observed with significantly higher bacterial loads during the summer. Binomial logistic regression models revealed larger (older) limpets were more likely to harbour vibrios, and the growth of bacteria on TCBS was a key predictor for PCR-based vibrio detection. Histological assessment of > 340 animals revealed little evidence of inflammation, sepsis or immune reactivity despite the gross bacterial numbers. We contend that slipper limpets are not highly susceptible to bacteriosis at either site, and do not harbour vibrios known to be pathogenic to humans. The lack of susceptibility to local pathogenic bacteria may explain, in part, the invasion success of C. fornicata across this region.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Molluscs form important economic and ecological components of marine ecosystems (FAO 2017; Sowa et al. 2019). They aid in determining benthic community structure, nutrient cycling, act as food sources for higher trophic levels, stabilize shorelines, and maintain water quality, in addition to being exploited for commercial aquaculture and wild-harvest industries (Goss-Custard et al. 2004; Jansen et al. 2012; Kellogg et al. 2014; Walles et al. 2015; Tomatsuri and Kon 2017). As many aquatic molluscs filter-feed, this can lead to the accumulation of diverse bacterial consortia, notably Vibrio spp., with some hosts acting as key reservoirs/sources of pathogenic ecotypes. Various species of vibrios are pathogenic to aquatic molluscs including those of commercial importance (Lynch et al. 2022).
The Atlantic slipper limpet, Crepidula fornicata, is one such filter feeder, and represents a prolific invasive, non-native species (INNS) introduced to Europe in the 1870s with shipments of American oysters (Crassostrea virginica; Blanchard 1997). Slipper limpets tend to form stacks, which involves a large female settling on a substrate, followed by several smaller males adhering on top. For over a century, C. fornicata has occupied European coastlines reaching densities of as many as 4,700 limpets m− 2 (in the bay of Marennes-Oléron, France; de Montaudouin and Sauriau 1999).
These limpets have gained notoriety as oyster pests because of their ability to modulate benthic community composition, sedimentation and pseudofaeces accumulation, and food competition with native and Pacific oysters (Ostrea edulis and Crassostrea gigas, respectively), queen scallops (Aequipecten opercularis) and blue mussels (Mytilus edulis) (Thieltges et al. 2003, 2006; Thieltges 2005; Richard et al. 2006; Kriefall et al. 2018; Preston et al. 2020). There are, however, conflicting reports on the nature of C. fornicata’s adverse influence. De Montaudouin et al. (1999) reported that C. gigas growth, macrozoobenthic density and diversity was not negatively impacted by the presence of C. fornicata. Hayer et al. (2019) reconstructed the changes in distribution and diversity of oysters over the last 200 years in the North Sea, concluding that the decline of O. edulis was near completion before the introduction of C. fornicata.
There is much interest in C. fornicata as an ecosystem engineer and a driver of ecological phase shift outside its native range, yet there is a paucity of knowledge with respect to its pathogen/parasite profile and epizootiology (Quinn et al. 2020). Such data are key to considering the risk of disease transfer to other co-located commercially important oysters (C. gigas) and mussels (M. edulis). Crepidula fornicata can act as a sink of trematode cercariae that use mytilids as intermediate hosts (Pechenik et al. 2001; Thieltges et al. 2006), although infections or tissue damage are rare in limpets themselves (Quinn et al. 2022). Le Cam and Viard (2011) performed a single, systematic study of disease in French populations of C. fornicata, revealing that ~ 43% were infested by a sponge Cliona celata between 2004 and 2007. Regarding bacterial presence, Choquet (2004) isolated a strain of Vibrio tapetis, the cause of brown ring disease in clams, from C. fornicata.
Members of the bacterial genus Vibrio can infect marine invertebrates. Highly virulent strains are associated with mass die-off events of commercial bivalves and crustaceans, e.g., Vibrio aestuarianus in Pacific oysters (Labreuche et al. 2006; Lupo et al. 2019) and V. parahaemolyticus in penaeid shrimp (Kumar et al. 2020). Vibrios are ubiquitous in aquatic environments, found naturally in marine, estuarine and freshwater situations. Many Vibrio species are harmless, yet some cause disease in fish (Froelich and Noble 2016) and ~ 12 species cause illness in humans (e.g., seafood-associated bacterial gastroenteritis) (Liu et al. 2004; Scallan et al. 2011).
Our aim was to perform a large-scale temporal survey for vibrio-like bacteria in the alien invader C. fornicata at two locations in South Wales with established populations – Swansea Bay, in a native oyster restoration zone, and Milford Haven, in an area of key fisheries/culture industries (Fig. 1). We sampled limpets at each site for one year (January–December 2019) and used a multi-resource screen for determining vibrio presence, i.e. immune cell (haemocyte) numbers, bacterial load quantification, histological examination of tissues and PCR for Vibrio identification.

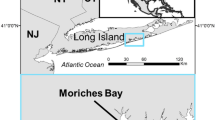
Locations of sample sites in South Wales, UK. Inset, geographic location of Swansea Bay in the United Kingdom (red box). Maps were created in QGIS 3.10.3-A Coruña
Methods
Sampling
We collected live adult limpets every month from January to December 2019 at each site. At Swansea Bay (51.570345, -3.974591) limpets were sampled subtidally by dredging (< 2.9 m below chart datum), whereas at Milford Haven (Hazelbeach; 51.7042, -4.971295) limpets were handpicked intertidally during low tide. Sample size calculations using an α-value of 0.05 and desired power > 80% indicated that a minimum of 57 (1-sided test) up to 73 (2-sided test) limpets were required based on an a priori prediction of 15–35% prevalence of infected animals (in line with observations of Le Cam and Viard (2011) when screening limpets for the sponge Cliona celata). We therefore collected 75 limpets each month from each site, yielding 1800 in total.
Biometric and population data
Wet weight (g), shell length (mm), shell width (mm) and position in a limpet stack were recorded for each specimen. Sex was determined and assigned as either male (presence of developed penis), female (absence of penis), or transitionary (reduced penis) as described by Coe (1936).
Haemocyte counts and bacterial loads
Haemolymph was collected from each limpet by removing the flesh from the shell using a bunt-ended probe, allowing the haemolymph to pool in the shell cavity. Haemolymph was then aspirated using a needle (23G) fitted onto a 1 mL sterile syringe. Total haemocyte (immune cell) counts were recorded using an improved Neubauer haemocytometer. Bacterial colony-forming units (CFUs) were determined by spreading 100 µL of haemolymph diluted 1:100 with 3% NaCl solution on tryptone soya agar (TSA) plates supplemented with 2% NaCl (two technical replicates (plates) per sample), and tryptone thiosulfate-citrate-bile salts-sucrose (TCBS) agar. Plates were incubated at 25 °C for 24 h prior to counting CFUs. Bacterial loads are expressed as CFUs per mL haemolymph.
Molecular detection and identification of Vibrio spp.
Sigma Aldrich GenElute Blood Genomic DNA kits were used for DNA extraction from C. fornicata haemolymph (100 µL) following the supplier’s guidelines. For 29 of the 1800 specimens collected it was not possible to obtain 100 µL of haemolymph because of their small size; instead, 20 mg of solid foot tissue (largely muscle) was used. DNA yields were quantified using the Invitrogen Qubit Fluorometer in combination with the Invitrogen dsDNA High Sensitivity Assay Kit.
All PCR assays were carried out in 25 µL total reaction volumes using 2x Mastermix (New England Biolabs Inc., Ipswich, USA), 1.25 µL oligonucleotide primers (10 µM) synthesized by Eurofins (Ebersberg, Germany), 2 µL of genomic DNA (1–3 µg), and performed in a PCR thermocycler (BioRad Laboratories Inc., Hemel Hempstead, UK). For amplification of vibrio DNA, the Vib1-F (GGCGTAAAGCGCATGCAGG) and Vib2-R (GAAATTCTACCCCCCTCTACAG) universal primer set (Vezzulli et al. 2012) was used, following conditions described by Thompson et al. (2004) and modified by Vezzulli et al. (2012). The expected amplicon size was 113 bp. Negative controls consisted of DEPC-treated Molecular Biology Grade Water (Sigma Aldrich) in the absence of DNA template to avoid false positives due to contamination. Positive controls consisted of 1 µL (~ 200 ng) DNA purified from the haemolymph of an infected crab (Carcinus maenas) (Davies et al. 2022). PCR products were visualised using 2% (w/v) agarose/TBE gels.
In preparation for sequencing of material from gel bands, amplicons were purified using HT ExoSAP-IT Fast high-throughput PCR product clean-up (Thermo Fisher Scientific, Altrincham, UK). Direct Sanger sequencing was carried out by Eurofins. In total, 72 representative samples were chosen from each month/site sampled and 22 of these returned usable data. Chromatograms were checked for mis-called bases. The sequences generated were trimmed of primer regions manually and matched against the National Center for Biotechnology Information (NCBI) nucleotide database using BLASTn (Basic Local Alignment Search Tool). Query Coverage (QC), Maximum Identity (MI) and E-value data were recorded for the top three returns per sample. Sequences were submitted to the NCBI short read archive (SRA) under accession numbers SRR13165025–SRR13165046.
Retrieved sequences were screened against GenBank using default BLASTn settings followed by restricting the search to the genus Vibrio. Bacterial nucleotide sequences derived from C. fornicata were added to a selection of known, geographically distributed Vibrio reference sequences to make up a dataset. A complete sequence alignment was achieved using the Clustal tool in MEGA X. Evolutionary analyses and reconstructions were carried out in MEGA X (Kumar et al. 2018) using the maximum likelihood routine based on the Kimura 2-parameter model and an independent Neighbour Joining routine. A consensus tree with the highest log likelihood value (− 150.41) from 1,000 bootstrap re-samplings was formatted using iTOL (Letunic and Bork 2019).
Tissue histology
A subset of 343 animals was screened via whole tissue histology to visualise potential immune responses to vibrio-like bacteria, e.g., haemocyte infiltration or tissue damage. Whole tissue histology of C. fornicata was performed as described by Quinn et al. (2020). Briefly, samples were submerged in Davidson’s seawater fixative for 24 h, rinsed in water and stored in 70% ethanol prior to processing. Samples were dehydrated using an ethanol series, transferred to Histoclear/Histochoice and infiltrated with molten wax prior to embedding. Sections were cut at 5–7 μm using a rotary microtome and stained with Cole’s haematoxylin and eosin. Images were adjusted for colour balance and contrast/brightness only.
Models and statistical analyses
To determine if specific predictor variables had any significant effect on the probability of finding vibrio-positive limpets, binomial logistic regression models were generated using the Logit link functions in the MASS library following Bernoulli distributions in RStudio v1.2.5033 and R v4.0.2. An information theoretical approach was used for both model selection and assessment of model performance (Richards, 2005). Initial models will from herein be referred to as full models. Using the drop1 function in R, each non-significant predictor variable from the full model was removed sequentially to enhance the predictive power of the final model. The function of drop1 is to compare the initial full model with the same model, but with the least significant predictor variable removed. If the difference between the reduced model and initial full model is significant, the removed predictor variable is kept out of the new, reduced model. A Chi-square test is used for comparison of the residual sum of squares in both models in the case of binomial response variables. Variables included in the full model were: vibrio (PCR negative/positive, 0 or 1), location (Milford Haven or Swansea Bay), season (Spring (March, April, May), Summer (June, July, August), Autumn (September, October, November), Winter (November, December, January)), sex (male, female, transitionary), wet weight (continuous number), and position in stack (1st, 2nd, 3rd, etc.). All other statistics (tests of normality, ANOVA, Kruskal-Wallis, Mann-Whitney, Chi-square) and figure preparation were performed in GraphPad Prism v.8/9 (GraphPad Software, La Jolla California USA).
Results
Biometric and population data
Wet weight, shell length and shell width of limpets dredged from Swansea Bay were significantly greater than at Milford Haven (Table 1; P < 0.0001 in all comparisons, Mann-Whitney). The proportions of male, female and transitionary limpets differed within a site but not when comparing the two sites. Female proportions were significantly greater than either males or transitionary limpets (Χ2 (2, n = 1800) = 8.852, P = 0.012). At Milford Haven, females accounted for 48%, males 25%, and transitionary limpets 27%. At Swansea Bay, females made up 49%, males 20%, and transitionary limpets 31%. Not only were there fewer males in general, but female and transitionary limpets were significantly heavier (P < 0.01, Kruskal-Wallis). This trend was also reflected in the average shell lengths and shell widths (Table 1) among the sexes: females and transitionary limpets were significantly longer and wider when compared to males (P < 0.001, Kruskal-Wallis).
Crepidula fornicata form stacks (Fig. 2a) on hard surfaces or basibionts (e.g., oyster or mussel shells). Stack sizes of limpets in Swansea Bay (up to 11 individuals) were consistently, and significantly, greater than those in Milford Haven (up to five individuals; Χ2(10, N = 1800) = 95.0, P < 0.0001; Chi-square test). At the latter site, 50% of the stacks were individual limpets, 35% consisted of two individuals, 12% of three individuals, and the remaining 3% of four or five individuals. At Swansea Bay, 40% of the stacks were individual limpets, 24% of two, 14% of three, 8% of four, and the remaining 14% of five to 11 limpets. Generally, weight, length and width of limpets decreased the further up the stack (Fig. 2b and c). Position within a stack was correlated with the wet weight of an individual, with those at the bottom being significantly larger than those above (P < 0.0001, Kruskal-Wallis).

Differential stack formation of C. fornicata across two sites in South Wales, UK. a Limpets formed stacks up to 11 individuals at Swansea Bay (photo: January 2019). Morphometric data (weight, length, width) categorised according to position within stacks for Milford Haven (MH) b and Swansea Bay (SB) c. Sample sizes are shown in the weight panels for each position within the stack
Haemocyte counts and bacterial loads
Freely circulating, homogeneous immune cell (haemocyte) populations were present in the haemolymph of C. fornicata (Fig. 3) and were quantified from 1,771 individuals (Milford Haven n = 883, Swansea Bay n = 888). Overall, significantly fewer haemocytes were present in limpets from Milford Haven (mean ± standard deviation 2.7 × 106 ± 9.3 × 104 mL− 1, range 1.3 × 105 – 3.3 × 107 mL− 1) than Swansea Bay (3.3 × 106 ± 1.1 × 105 mL− 1, 3 × 105 – 3.1 × 107 mL− 1) (P < 0.01). Significant fluctuations in haemocyte numbers were apparent with season (F(3, 1761) = 21.56, P < 0.001) and site (F(1, 1761) = 17.96, P < 0.001). At each site, haemocyte levels peaked in the summer/autumn, ranging from ~ 2.8 × 105 to 3.3 × 107 mL− 1, and were at their nadir in spring at Swansea Bay and winter at Milford Haven, ranging from ~ 1.3 × 105 to 1.6 × 107 mL− 1 (Fig. 3).

Total haemocyte counts observed in the haemolymph of C. fornicata. Temporal profiles of haemocyte numbers according to site and season (Milford Haven (MH) = 883; Swansea Bay (SB) n = 888). Unshared letters denote significant differences (P < 0.05)
Sufficient volume of haemolymph was retrieved from 1,780 individuals and plated on the generalist TSA (+ 2% NaCl) growth medium, and on the vibrio selective TCBS agar (Fig. 4). Of these samples, only 8 (TSA) and 12 (TCBS) lacked any sign of cultivable bacteria. No significant differences in bacterial loads were detected between Milford Haven and Swansea Bay samples for either TSA or TCBS (P = 0.487 and P = 0.665 (Mann-Whitney), respectively; Table 2). A clear and similar temporal trend was detected among haemolymph CFU densities from the two locations across the 12-month survey (Fig. 4a and b). When data were combined from both sites, CFU numbers were at their highest during the summer and lowest in winter (Table 2). There were some differences in monthly CFU highs/lows when comparing each site and growth medium (Supplementary Fig. 1).

Spatiotemporal patterns of cultivable vibrio-like bacteria from the haemolymph of C. fornicata. Average number of bacterial CFUs per site, per season on TSA a and TCBS b media. Individuals were sampled from Swansea Bay (SB) and Milford Haven (MH). Unshared letters represent significant differences (P < 0.05) determined from Tukey’s post-hoc tests (n = 1780)
Stack position of a limpet was not significantly related to CFU values on either TSA (P = 0.32) or TCBS agar (P = 0.063; Supplementary Fig. 2). The bacterial burden (CFUs mL1) of the haemolymph and haemocyte numbers in Milford Haven limpets were not significantly correlated on either medium (TSA, P = 0.978; TCBS, P = 0.157) (Supplementary Fig. 3a) but were strongly positively correlated on both media in Swansea Bay limpets (P < 0.001 for both TSA and TCBS, Spearman correlation; Supplementary Fig. 3b).
Molecular detection and identification of Vibrio spp.
All 1,800 limpets were screened for the presence of Vibrio spp. using end-point PCR, with 85% (763/900) and 81% (729/900) testing positive (single amplicons of the expected size; e.g., Supplementary Fig. 4 for Milford Haven) for Swansea Bay and Milford Haven, respectively. August had the highest number of PCR-positive samples (150/150) and February the lowest 84/150 (56%) when data from the two sites were combined (Fig. 5).

Prevalence of vibrio-like bacteria among C. fornicata determined via PCR-based screening. Limpets detected positive for Vibrio spp. according to site and month
Twenty two high quality sequences were generated with seven proving clear matches to known Vibrio species and the remainder matching unclassified/uncultured vibrio-like bacteria (Supplementary Table 1). BLASTn searches revealed consistent matches (85–100% coverage/identity) with members of the Splendidus clade (V. tasmaniensis, V. celticus, V. crassostreae, V. gallaecicus, V. kanaloae and V. toranzoniae) and Mediterranean clade (V. mediterranei). Phylogenetic trees of the partial 16 S ribosomal RNA sequences displayed consistent topology when reconstructed using maximum likelihood and neighbour-joining routines – indicating the presence of some unique vibrio sequence types in C. fornicata (Fig. 6). Trees included vibrios isolated from diseased and healthy molluscs, as well as environmental isolates. A distinct Splendidus clade was formed with previously characterised vibrios, as well as sequences amplified from Milford Haven and Swansea Bay C. fornicata. A second, distinct well supported (99%) cluster of vibrio-like sequences included samples from Milford Haven and Swansea Bay only, thereby suggesting at least two vibrio ecotypes are present in these limpets. Sequences were distributed among benign and potentially pathogenic bacteria. Because of the small size of some final reads (~ 60 bp), caution is needed when interpreting the potential species identity of these bacteria; however, they do represent members of the Vibrio genus and the Splendidus clade.

Consensus phylogram (unrooted) of the partial 16 S rRNA region recovered from slipper limpets Crepidula fornicata (SRR13165025–SRR13165046). Reference sequences represent the top BLASTn-search results from NCBI plus a broad list of known, geographically distributed Vibrio species. Bootstrap support values are depicted as spheres (ML | NJ). The scale represents nucleotide substitutions per site (maximum likelihood (ML) estimation, 1000 bootstrap replicates). All reference sequences used were ≥ 90% (coverage and identity) matches for the sequences retrieved from limpets. Stars represent sampling sites (blue, Milford Haven; yellow, Swansea Bay). The red box highlights a putative vibrio ecotype within the Splendidus clade
Models
Biometric data recorded (size, sex, and position in stack), along with collection site, bacterial CFU counts and haemocyte levels were incorporated into Binomial Logistic Regression models to determine whether any of these factors contributed to the occurrence/detection of Vibrio spp. by PCR. Limpet wet weight, season and TCBS counts were significant predictor variables (Table 3). Significant decreases in the number of vibrio-positive limpets were observed during Spring (P = 0.0053) and Winter (P = 0.0064) (Fig. 7). Heavier limpets (13.8 ± 0.24 g) were significantly (P < 0.001) more likely to test positive for Vibrio spp. than limpets testing negative (11.4 ± 0.44 g). On selective TCBS medium, individuals with more bacterial colonies were more likely to test positive (n = 1480) for vibrio-like species than those that tested negative (n = 299), P < 0.001; ~2 × 105 ± 7.7 × 103 CFU/mL versus ~ 9 × 104 ± 8.7 × 103 CFU/mL, respectively; there was no such trend on TSA.

Seasonal variation in limpets testing positive for Vibrio spp. via PCR. Data for both sites (Swansea Bay and Milford Haven) were combined. Significant differences are denoted by asterisks (** P < 0.01). The number of positive individuals from each season is placed above each column
Histological assessment of limpet tissues
There was no gross evidence of bacterial infection, tissue-wide damage and necrosis, or inflammation in the slipper limpets examined, regardless of their haemolymph bacterial load. In very few individuals, there were discrete signs of immune cell (haemocyte) infiltration, nodule formation, and varied states of what appeared to be compromised tissue; however, these could not be attributed to vibriosis.
Discussion
To date, few parasites or diseases have been reported in C. fornicata other than the shell boring sponge C. celata (Le Cam and Viard 2011) and interactions with trematodes (Pechenik et al. 2001; Thieltges et al. 2006, 2008; Quinn et al. 2022). We determined the bacterial loads of C. fornicata, focusing on a key aetiological agent in commercial shellfish, namely Vibrio spp. Slipper limpets from two survey sites contained high levels of vibrio-like bacteria in the haemolymph (~ 83 to > 99% depending on the detection method), but these were not pathogenic to C. fornicata (as determined via histology). Instead, the limpets seemed to accumulate bacteria passively as a result of their filter feeding nature, thereby acting as a sink for these microbes autochthonous to the area. Such observations have been made previously for marine invertebrates (Froelich and Oliver 2013). Bivalves, such as clams, can be found with levels of vibrios in their tissue of 105 CFU/g or higher, this being up to 100-fold greater than in the surrounding waters (Froelich et al. 2017). Larger limpets were statistically more likely to contain vibrios, and at higher levels. This could be a consequence of age or immune-senescence as older limpets are positioned toward the bottom of a stack onto which younger/smaller limpets pile on and remain in situ for extended periods of time, thereby increasing the likelihood of bacterial accumulation.
The genus Vibrio includes free-living bacteria, opportunists, pathobionts and symbionts (e.g., McFall-Ngai 2014; Wendling et al. 2014; Destoumieux-Garzón et al. 2020). A great diversity of Vibrio species, strains and ecotypes can be found among the microflora of marine invertebrates, e.g., representing ~ 21 to 77% of bacterial isolates of spider crab (Maja brachydactyla) haemolymph sampled along the coastal waters of north-west Spain and the Canary Islands (Gomez-Gil et al. 2010). Known pathogenic species of vibrios that affect molluscs include strains of V. aestuarianus, V. alginolyticus, V. bivalvicida, V. harveyi, V. spendidus, V. parahaemolyticus, V. pectenicida and V. tapetis but none of these were found in the limpets screened in this study. For vibrios to colonise a host, they need to be able to attach and penetrate intact tissues. Non-pathogenic strains tend to not have the capacity to produce necessary adhesions or enzymes/factors that permit their invasion (Lynch et al. 2022). Much work has been performed on the environmental drivers of microbial disease in bivalve molluscs, like the Pacific oyster (C. gigas), linked to vibriosis and mass mortality events that can devastate populations (Destoumieux-Garzón et al. 2020; Lynch et al. 2022) but little information exists on vibriosis in gastropod molluscs probably because of their limited commercial importance. Despite some morphometrical differences between C. fornicata collected at Swansea Bay and Milford Haven, site was not a significant factor regarding the probability of a limpet testing positive for vibrios. Instead, seasonality was a key contributing factor. The highest prevalence of vibrio-like bacteria occurred during summer and late autumn. This is not entirely surprising as higher temperatures, especially those exceeding 15–17 °C, enable vibrios to thrive and are linked to disease outbreaks and the expression of virulence factors (Stauder et al. 2010; Vezzulli et al. 2012, 2013). In addition to season, both weight and CFU growth on TCBS medium, were key predictors of bacterial presence. Criticism has been levelled at the use of TCBS medium for monitoring vibrios other than key pathogens such as V. cholerae and V. parahaemolyticus (Donovan and van Netten 1995; Cavallo and Stabili 2002; Gyraite et al. 2019). This medium was originally developed to enumerate vibrios from human and environmental samples (Kobayashi et al. 1963); however, it is apparent that not all vibrios grow on this medium and that some other taxa, such as aeromonads, can grow on it (Lotz et al. 1983). Even considering the putative drawbacks of using TCBS agar for vibrio enumeration in haemolymph, our results found that CFU growth on TCBS aligned with PCR outputs, thereby representing a validated approach for monitoring vibrio-limpet epizootiology as the coastal waters become warmer (or limpets move further outside of their native range). The frequency of vibrio infections has been increasing worldwide: between 1996 and 2006, the average annual incidence rose by 78%, and between 2008 and 2018 by 272% (Froelich and Daines 2020). In summer 2015, Canada experienced the highest number of cases of human V. parahaemolyticus infections ever reported from the consumption of raw oysters (Taylor et al. 2018). This outbreak was associated with higher-than-normal sea surface temperatures (SST > 15 °C), and the incidence rates decreased once SST dropped below 15 °C (Taylor et al. 2018).
Limpets at both our survey sites are co-located with high value shellfish, e.g., native oyster (Ostrea edulis) in Swansea Bay. Therefore, part of our study was to determine the potential connectivity of disease, i.e., whether slipper limpets represent an overlooked reservoir for disease-causing vibrios. PCR-based detection of vibrios in limpets using the universal Vib1-F/Vib2-R primer set was associated significantly with culture (TCBS)-based detection of bacteria, indicating either method is reliable for vibrio detection in limpets. However, the amplicons generated were short (~ 60–100 bp), making it difficult to identify vibrios to species. Nonetheless, high quality sequences (although short) were retrieved across sites and months and identified as members of the Splendidus clade (e.g., V. tasmaniensis, V. celticus), and putative members of the Mediterranean clade. The former clade is commonly encountered among the ‘vibriome’ of marine animals (e.g., Romalde et al. 2014). Only a limited number of species in the Vibrionaceae are a threat to shellfish or humans. We did not encounter evidence to suggest that major pathogens of humans (e.g., V. cholerae and V. parahaemolyticus) or shellfish (e.g., V. aestuarianus) are enriched within the bacterial consortia of slipper limpet haemolymph.
Concluding remarks
Our study goes some way to address the knowledge deficit with respect to the disease profile of invasive C. fornicata, alongside identification of key variables (age and seasonality) that influence vibrio presence in limpets at two locations in South Wales, UK. The vibrios present in limpet haemolymph may provide an element of protection through competitive exclusion of other disease-causing agents but this is speculative. We consider that slipper limpets are not highly susceptible to bacteriosis at either site, and do not harbour vibrios known to be pathogenic to humans. The lack of susceptibility to local pathogenic bacteria may explain, in part, the invasion success of C. fornicata across this region.
Data Availability
Sequence data have been deposited into NCBI’s short read archive (SRA) under accession numbers SRR13165025 – SRR13165046. Data for limpet morphometrics, haemocyte numbers, bacterial colony-forming units and PCR-based screening have been included as supplementary material.
Code Availability
Not applicable.
References
Blanchard M (1997) Spread of the slipper limpet Crepidula fornicata (L. 1758) in Europe. Current state and consequences. Sci Mar 61:109–118
Cavallo RA, Stabili L (2002) Presence of vibrios in seawater and Mytilus galloprovincialis (Lam.) from the Mar Piccolo of Taranto (Ionian Sea). Water Res 36(15):3719–3726
Choquet G (2004) Caractérisation et pathogénie des isolats de Vibrio tapetis, bactérie responsable de la maladie de l’anneau brun chez la palourde japonaise. Doctorat Microbiologie Thesis. Brest: Universite de Bretagne occidentale-Brest
Coe W (1936) Sexual Phases in Crepidula. J Exp Zool 83:455–477
Davies CE, Thomas JE, Malkin SH, Batista FM, Rowley AF, Coates CJ (2022) Hematodinium sp. infection does not drive collateral disease contraction in a crustacean host. eLife 11:e70356. DOI: https://doi.org/10.7554/eLife.70356
de Montaudouin X, Sauriau PG (1999) The proliferating Gastropoda Crepidula fornicata may stimulate macrozoobenthic diversity. J Mar Biolog Assoc U K 79(6):1069–1077
Destoumieux-Garzón D, Canesi L, Oyanedel D, Travers MA, Charrière GM, Pruzzo C, Vezzulli L (2020) Vibrio–bivalve interactions in health and disease. Environ Microbiol 22(10):4323–4341
Donovan TJ, Van Netten P (1995) Culture media for the isolation and enumeration of pathogenic Vibrio species in foods and environmental samples. Prog Ind Microbiol 34:203–217
FAO (2017) FAO yearbook of fishery and aquaculture statistics. Fish Aquac Dep 0:26–28
Froelich BA, Daines DA (2020) In hot water: effects of climate change on Vibrio–human interactions. Environ Microbiol 22(10):4101–4111
Froelich BA, Noble RT (2016) Vibrio bacteria in raw oysters: Managing risks to human health. Philos Trans R Soc B Biol Sci 371:e20150209. https://doi.org/10.1098/rstb.2015.0209
Froelich B, Oliver JD (2013) The interactions of Vibrio vulnificus and the oyster Crassostrea virginica. Microb Ecol 65(4):807–816
Froelich BA, Phippen B, Fowler P, Noble RT, Oliver JD (2017) Differences in abundances of total Vibrio spp., V. vulnificus, and V. parahaemolyticus in clams and oysters in North Carolina. Appl Environ Microbiol 83(2): e02265-16
Gomez-Gil B, Roque A, Lacuesta B, Rotllant G (2010) Diversity of vibrios in the haemolymph of the spider crab Maja brachydactyla. J Appl Microbiol 109(3):918–926
Goss-Custard JD, Stillman RA, West AD, Caldow RWG, Triplet P, Dit Durell SEALV, McGrorty S (2004) When enough is not enough: Shorebirds and shellfishing. Proc R Soc B Biol Sci 271:233–237. https://doi.org/10.1098/rspb.2003.2602
Gyraite G, Katarzyte M, Schernewski G (2019) First findings of potentially human pathogenic bacteria Vibrio in the south-eastern Baltic Sea coastal and transitional bathing waters. Mar Pollut Bull 149:e110546
Hayer S, Bick A, Brandt A, Ewers-Saucedo C, Fiege D, Füting S et al (2019) Coming and going - Historical distributions of the European oyster Ostrea edulis Linnaeus, 1758 and the introduced slipper limpet Crepidula fornicata Linnaeus, 1758 in the North Sea. PLoS ONE 14:e0224249
Jansen HM, Strand Ø, Verdegem M, Smaal A (2012) Accumulation, release and turnover of nutrients (C-N-P-Si) by the blue mussel Mytilus edulis under oligotrophic conditions. J Exp Mar Bio Ecol 416–417:185–195. https://doi.org/10.1016/j.jembe.2011.11.009
Kellogg ML, Smyth AR, Luckenbach MW, Carmichael RH et al (2014) Use of oysters to mitigate eutrophication in coastal waters. Estuar Coast Shelf Sci 151:156–168. https://doi.org/10.1016/j.ecss.2014.09.025
Kobayashi T, Enomoto S, Sakazaki R, Kuwahara S (1963) A new selective medium for pathogenic vibrios, TCBS agar (modified Nakanishi’s agar). Jpn J Bacteriol 18:387–391
Kriefall NG, Pechenik JA, Pires A, Davies SW (2018) Resilience of Atlantic slippersnail Crepidula fornicata larvae in the face of severe coastal acidification. Front Mar Sci 5:e312
Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35:1547–1549. https://doi.org/10.1093/molbev/msy096
Kumar R, Ng TH, Wang HC (2020) Acute hepatopancreatic necrosis disease in penaeid shrimp. Rev Aquac 12:1867–1880. https://doi.org/10.1111/raq.12414
Labreuche Y, Lambert C, Soudant P, Boulo V, Huvet A, Nicolas JL (2006) Cellular and molecular hemocyte responses of the Pacific oyster, Crassostrea gigas, following bacterial infection with Vibrio aestuarianus strain 01/32. Microbes Infect 8:2715–2724. https://doi.org/10.1016/j.micinf.2006.07.020
Le Cam S, Viard F (2011) Infestation of the invasive mollusc Crepidula fornicata by the native shell borer Cliona celata: A case of high parasite load without detrimental effects. Biol Invasions 13:1087–1098. https://doi.org/10.1007/s10530-011-9958-1
Letunic I, Bork P (2019) Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res 47:256–259. https://doi.org/10.1093/nar/gkz239
Liu X, Chen Y, Wang X, Ji R (2004) Foodborne disease outbreaks in China from 1992 to 2001 national foodborne disease surveillance system. J Hyg Res 33:725–727
Lotz MJ, Tamplin ML, Rodrick GE (1983) Thiosulfate-citrate-bile salts- sucrose agar and its selectivity for clinical and marine vibrio organisms. Annals Clin Lab Sci 13:45–48
Lupo C, Travers MA, Tourbiez D, Barthélémy CF, Beaunée G, Ezanno P (2019) Modeling the transmission of vibrio aestuarianus in pacific oysters using experimental infection data. Front Vet Sci 6:1–15. https://doi.org/10.3389/fvets.2019.00142
Lynch SA, Rowley AF, Longshaw M, Malham SK, Culloty SC (2022) Diseases of molluscs. Invertebrate Pathology eds AF Rowley, CJ Coates and MMA Whitten. Oxford University Press, Oxford, pp 171–215
McFall-Ngai M (2014) Divining the essence of symbiosis: insights from the squid-vibrio model. PLoS Biol 12(2):e1001783
Pechenik JA, Fried B, Simpkins HL (2001) Crepidula fornicata is not a first intermediate host for trematodes: who is? J Exp Mar Biol Ecol 261(2):211–224
Preston J, Fabra M, Helmer L, Johnson E, Harris-Scott E, Hendy IW (2020) Interactions of larval dynamics and substrate preference have ecological significance for benthic biodiversity and Ostrea edulis Linnaeus, 1758 in the presence of. Crepidula fornicata Aquat Conserv 30(11):2133–2149
Quinn EA, Malkin SH, Rowley AF, Coates CJ (2020) Laccase and catecholoxidase activities contribute to innate immunity in slipper limpets, Crepidula fornicata. Dev Comp Immunol 110:e103724
Quinn EA, Thomas JE, Malkin SH, Eley MJ, Coates CJ, Rowley AF (2022) Invasive slipper limpets Crepidula fornicata are hosts for sterilizing digenean parasites. Parasitology 149:811–819
Richard J, Huet M, Thouzeau G, Paulet YM (2006) Reproduction of the invasive slipper limpet, Crepidula fornicata, in the Bay of Brest, France. Mar Biol 149(4):789–801
Richards SA (2005) Testing ecological theory using the information-theoretic approach: examples and cautionary results. Ecology 86:2805–2814. https://doi.org/10.1890/05-0074
Romalde JL, Diéguez AL, Lasa A, Balboa S (2014) New Vibrio species associated to molluscan microbiota: A review. Front Microbiol 4:1–11. https://doi.org/10.3389/fmicb.2013.00413
Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV et al (2011) Foodborne illness acquired in the United States-Major pathogens. Emerg Infect Dis 17:7–15. https://doi.org/10.3201/eid1701.P11101
Sowa A, Krodkiewska M, Halabowski D, Lewin I (2019) Response of the mollusc communities to environmental factors along an anthropogenic salinity gradient. Sci Nat 106. https://doi.org/10.1007/s00114-019-1655-4
Stauder M, Vezzulli L, Pezzati E, Repetto B, Pruzzo C (2010) Temperature affects Vibrio cholerae O1 El Tor persistence in the aquatic environment via an enhanced expression of GbpA and MSHA adhesins. Environ Microbiol Rep 2(1):140–144
Taylor M, Cheng J, Sharma D, Bitzikos O, Gustafson R et al (2018) Outbreak of Vibrio parahaemolyticus associated with consumption of raw oysters in Canada, 2015. Foodborne Pathog Dis 15(9):554–559
Thieltges DW (2005) Impact of an invader: epizootic American slipper limpet Crepidula fornicata reduces survival and growth in European mussels. Mar Ecol Progr Ser 286:13–19
Thieltges D, Strasser M, Reise K (2003) The American slipper limpet Crepidula fornicata (L.) in the northern Wadden Sea 70 years after its introduction. Helgol Mar Res 57(1):27–33
Thieltges DW, Strasser M, Reise K (2006) How bad are invaders in coastal waters? The case of the American slipper limpet Crepidula fornicata in western Europe. Biol Invasions 8(8):1673–1680
Thieltges DW, Jensen KT, Poulin R (2008) The role of biotic factors in the transmission of free-living endohelminth stages. Parasitology 135(4):407–426
Thompson FL, Lida T, Swings J (2004) Biodiversity of vibrios. Microbiol Mol Biol Rev 68:403–431. https://doi.org/10.1128/MMBR.68.3.403
Tomatsuri M, Kon K (2017) Effects of dead oyster shells as a habitat for the benthic faunal community along rocky shore regions. Hydrobiologia 790:225–232. https://doi.org/10.1007/s10750-016-3033-y
Vezzulli L, Brettar I, Pezzati E, Reid PC, Colwell RR, Höfle MG, Pruzzo C (2012) Long-term effects of ocean warming on the prokaryotic community: Evidence from the vibrios. ISME J 6:21–30. https://doi.org/10.1038/ismej.2011.89
Vezzulli L, Colwell RR, Pruzzo C (2013) Ocean warming and spread of pathogenic vibrios in the aquatic environment. Microb Ecol 65(4):817–825
Walles B, Salvador de Paiva J, van Prooijen BC, Ysebaert T, Smaal AC (2015) The ecosystem engineer Crassostrea gigas affects tidal flat morphology beyond the boundary of their reef structures. Estuar Coast 38:941–950. https://doi.org/10.1007/s12237-014-9860-z
Wendling CC, Batista FM, Wegner KM (2014) Persistence, seasonal dynamics and pathogenic potential of Vibrio communities from Pacific oyster hemolymph. PLoS ONE 9(4):e94256
Acknowledgements
We thank skippers Mr Keith Naylor and Mr Max Robinson (R.V. Mary Anning, Swansea University) and Mr Barry Thomas for their assistance with sample collection in the Swansea Bay area.
Funding
Operations were part funded by the European Regional Development Fund through the Ireland-Wales Cooperation programme, BLUEFISH, awarded to CC and AR, and Swansea University start-up funds assigned to CC. A BLUEFISH innovation bursary and a College of Science (Swansea University) doctoral training grant supported EQ.
Author information
Authors and Affiliations
Contributions
Conceptualization, CC. Data curation, EQ, SM, JT, AR, CC. Formal analysis, EQ, AR, CC. Funding acquisition, AR, CC. Investigation, EQ, JT, SM, RP, CD, AR, CC. Methodology, EQ, JT, SM, CD, AR, CC. Project administration, SM, AR, CC. Resources, AR, CC. Software, EQ, CD, CC. Supervision, AR, CC. Validation, EQ, AR, CC. Visualisation, EQ, CC. Writing – original draft presentation, reviewing and editing, EQ, AR, CC.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests, financial or otherwise.
Ethics approval
Sampling and experimental work on C. fornicata were approved by the College of Science (Swansea University) research ethics committee, SU-Ethics-111217/446.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Quinn, E.A., Malkin, S.H., Thomas, J.E. et al. Invasive slipper limpets (Crepidula fornicata) act like a sink, rather than source, of Vibrio spp.. Biol Invasions 24, 3647–3659 (2022). https://doi.org/10.1007/s10530-022-02868-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-022-02868-6