
Abstract
Small fish species such as minnows (Phoxinus sp.) are introduced into mountain lakes by anglers that use them as live bait for fishing salmonids that were previously introduced in these naturally fishless habitats. Introduced fish severely impact native biota but minnows are nevertheless released believing to provide forage for game fish, i.e., salmonids. However, our results indicate that minnows negatively interact with game fish, reducing their relative densities. On some occasions, minnows can remain the only present fish following trout extinction. We believe that this information should be urgently disseminated among anglers as a pragmatic argument to also convince anglers less sensitive to conservation issues that carrying minnows to the mountains is not a good idea whatever the main interest is, e.g., nature conservation or outdoor recreation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
though fish introductions severely impact native biota of originally fishless mountain lakes (Knapp et al. 2001; Ventura et al. 2017; Miró et al. 2018), salmonids are commonly introduced to sustain or promote recreational angling. Such game fish are mainly introduced via institutional stocking programmes, but their presence facilitates the collateral introductions of some small fish such as minnows Phoxinus sp. (Fam. Cyprinidae) which are used as live baits by anglers (Museth et al. 2007; Miró and Ventura 2015). As the case has been in northern Europe (Museth et al. 2007), such illegal introductions are spreading minnows in the Pyrenees (Miró and Ventura 2015), Alps (Tiberti et al. 2020; De Santis et al. 2021), and -likely- in other mountain ranges across Europe.
The release of minnows can be either accidental or conscious. In the latter case, the release can be due either to carelessness, or to the common belief that small cyprinids can become food for trout improving their populations, namely “forage fish” (Miró and Ventura 2015). However, the alleged positive effects of minnows on trout are generally not supported by empirical data and neither by food-web theory. On the contrary, scientific literature shows some examples of their negative ecological interactions, mainly from some boreal mountain lakes, where minnows negatively affect trout populations by competition for food and predation on eggs (Lien 1981; Museth et al. 2007, 2010; Borgstrøm et al. 1996, 2010; Global Invasive Species Database 2021). Similar studies are however missing from the lakes of the southern European mountains, where minnow introductions are also widespread (Miró and Ventura 2015; Tiberti et al. 2020).
The aim of the present study was to assess if minnows negatively affect the densities of salmonids inhabiting the mountain lakes in a new geographic area, i.e., southern European mountain ranges (Alps and Pyrenees). Our main study hypothesis is that minnows negatively interact with salmonids, as already observed in northern regions (Museth et al. 2007). Our study hypothesis is verified if the densities of salmonids are lower when coexisting with cyprinids.
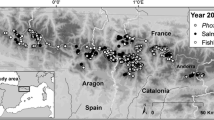
To test our hypothesis, between 2012 and 2019, we sampled 43 originally fishless mountain lakes, 30 with introduced salmonids only and 13 with both introduced salmonids and cyprinids (Fig. 1) in four Sites of Community Importance (SCI) from the Catalan Pyrenees and the Western Italian Alps: the SCIs Aigüestortes (ES0000022), Alt Pallars (ES5130003), Parco Nazionale del Gran Paradiso (IT1201000), and Parco Naturale Mont Avic (IT1202000) (Fig. 1). All study lakes are typical mountain lakes between 0.3 and 24.2 ha (mean ± sd: 4.75 ± 4.72 ha) and 3.0–73.0 m deep (mean ± sd: 15.3 ± 13.6 m), cold and ice-covered for 6–9 months per year, placed close or above the local timberline between 1618 and 2747 m a.s.l. (mean ± sd: 2321 ± 230 m a.s.l.). Introduced fish include several salmonid species and some populations of European minnow species complex, Phoxinus sp. (Fig. 1). Periodic fish stocking could artificially increase the density of salmonids, but, at the time of fish sampling, introductions were already prohibited or suspended since at least 4 years (from 4 to > 40 years) in all lakes.

Study area and lakes with presence of salmonids and minnows; a SCIs Aigüestortes and Alt Pallars in the Pyrenees (20 km grid); b SCIs Parco Nazionale Gran Paradiso and Parco Naturale Mont Avic in the Alps (20 km grid); c position of the SCIs in Europe (1000 km grid)
We sampled salmonids with multi-mesh gillnets and cyprinids with fyke-nets. In lakes with salmonids, we estimated their relative densities as Captures Per Unit Effort (CPUE; No. of salmonids × m−2 of gillnet × Day−1), deploying 1–4 multi-mesh gillnets in the central part of the lakes depending on the lake size. To this end, we used two kinds of gillnets: standard nets in the Pyrenean lakes (30 m long × 1.5 m high, divided into 12 panels with variable mesh size; CEN 2015) and some different nets in the Alpine lakes (36 m long × 1.8 m high, divided into six panels with increasing mesh sizes: 10.0, 12.5, 18.5, 25.0, 33.0, 38.0 mm). In lakes with cyprinids, their CPUE was estimated as No. of cyprinids × trap−1 × day−1 deploying 2–6 fish traps (i.e., fyke nets; 0.5 m wide × 0.4 m high D-shaped mouth, 4 mm mesh size with a 2 m long central wing or lead) in the littoral area of the lakes (with their mouths facing the shoreline). Because gillnetting and fyke netting sessions were sometimes repeated in the same lakes, in such cases we calculated an average CPUE weighting by the duration of the fishing sessions.
Cyprinids were found coexisting with Salmo trutta in 11 out of the 13 lakes with both salmonids and cyprinids, and they were never found in the eleven lakes with Salvelinus fontinalis. Their CPUE varied considerably among lakes (mean ± sd: 137.08 ± 202.14 minnows trap−1 day−1), up to 690.17 minnows trap−1 day−1. Also, the CPUE of salmonids varied a lot (mean ± sd: 0.41 ± 0.68 fish m−2 day−1) depending on the sampled lake and species, being Salvelinus fontinalis the species with the highest average and maximum CPUEs (Fig. 2a). Where salmonid and cyprinid coexist, the relationship between their CPUEs suggest that salmonids can reach higher densities only where minnow density was low and vice-versa (Fig. 3).

Panel a Boxplot comparing the relative densities (Capture Per Unit Effort-CPUE) of different salmonid species (OM: rainbow trout Oncorhynchus mykiss; ST: brown trout Salmo trutta; SA: arctic char Salvelinus alpinus; SF: brook trout Salvelinus fontinalis) in 43 mountain lakes from the Pyrenees and Alps; panel b salmonid CPUE when cyprinids are absent (S) or present (S + C); panel c salmonid CPUE when cyprinids are absent (S) or present (S + C), excluding lakes with brook trout Salvelinus fontinalis; panel d brown trout CPUE when cyprinids are absent (S) or present (S + C)

Relationship between salmonid and cyprinid Capture Per Unit Efforts (CPUEs) in the 13 mountain lakes with both fish families
To cope with non-normal distribution of CPUE data and small sample sizes, we tested the study hypothesis with a non-parametric one-tailed Wilcoxon rank sum tests (or Mann Whitney U Tests). The median CPUE of salmonids was significantly higher when cyprinids were not present (W = 322, n1 = 30, n2 = 13, p < 0.001; Fig. 2a). The same result could be observed also when we excluded the lakes with S. fontinalis (W = 181, n1 = 19, n2 = 13, p < 0.05; Fig. 2c) and when we considered only the lakes with S. trutta (W = 121, n1 = 15, n2 = 11, p < 0.05; Fig. 2d), to avoid possible bias related to the likely existence of different species-specific interactions between salmonid species and minnows.
The initial results obtained show a negative interaction between minnows and salmonids. Where minnows coexist with salmonids, the observed density patterns (Fig. 3) could be the result of a progressive replacement of trout by minnows in some lakes. This could be due to predation (e.g., on trout eggs) and competition, which -in a few lakes where fishing is permitted- sum up to the effects of trout removal by anglers. In this regard it is also interesting to note that there are mountain lakes where salmonid populations -including self-sustaining populations- disappeared after the introduction of cyprinids (Miró and Ventura 2015; Tiberti et al. 2020), which should be considered a further and extreme consequence of minnow introductions on salmonids. In addition, the fact that minnows were not found in any lake with S. fontinalis, suggest that also salmonid populations may exclude/control minnow populations, e.g., preventing their establishment by predation or competition for food resources (see Museth et al. 2003). Better understanding the mechanisms of mutual exclusion between minnows and salmonids could have potential management and conservation implication, for example in the field of biological control (Koenig et al. 2015). We however aim to address such mechanisms of exclusion in a better detail, when a more representative sample of lakes will be available.
In conclusion, our results confirm our study hypothesis showing that releasing minnows generally has negative consequences for salmonids, as already observed in some mountain and lowland lakes from the boreal region (Museth et al. 2007). While the exact nature of the ecological interaction between minnows and salmonids in southern mountain lakes systems deserves further studies, we believe that our results are sufficient to support the idea that minnow introductions should be avoided. The same indication stems from conservation instances, from several studies on the huge ecological impact that minnows generate in mountain lakes, through predation on native amphibians and invertebrates, and trophic cascades (Gacia et al. 2018; Miró et al. 2018; Tiberti et al. 2021). However, providing evidence of the impact of minnows on salmonids could be a highly effective argument for ensuring the collaboration of anglers (i.e., the main vectors of minnow invasion; Miró and Ventura 2015) and their compliance with some necessary restrictions about the use of minnows as live baits.
Data availability
All data are summarized in the paper.
Change history
28 July 2022
Missing Open Access funding information has been added in the Funding Note.
References
Borgstrøm R, Brittain JE, Hasle K, Skjølås S, Dokk JG (1996) Reduced recruitment in brown trout Salmo trutta, the role of interactions with the minnow Phoxinus phoxinus. Nordic J Freshwat Res 72:30–38
Borgstrøm R, Museth J, Brittain JE (2010) The brown trout (Salmo trutta) in the lake, Øvre Heimdalsvatn: long-term changes in population dynamics due to exploitation and the invasive species, European minnow (Phoxinus phoxinus). In: Brittain JE, Borgstrøm R (eds) The subalpine lake ecosystem, Øvre Heimdalsvatn, and its catchment: local and global changes over the last 50 years. Springer, Dordrecht, pp 81–91
De Santis V, Delmastro GB, Vanetti I, Britton JR, Zaccara S (2021) Species composition of introduced and natural minnow populations of the Phoxinus cryptic complex in the westernmost part of the Po River Basin (north Italy). Biol Inv 23:657–668
Gacia E, Buchaca T, Bernal-Mendoza N, Sabás I, Ballesteros E, Ventura M (2018) Non-native minnows threaten quillwort populations in high mountain shallow lakes. Front Plant Sci 9:329
Knapp RA, Matthews KR, Sarnelle O (2001) Resistance and resilience of alpine lake fauna to fish introductions. Ecol Monogr 71:401–421
Koenig MK, Meyer KA, Kozfkay JR, DuPont JM, Schriever EB (2015) Evaluating the ability of tiger muskellunge to eradicate Brook Trout in Idaho alpine lakes. North Am J Fish Manag 35:659–670
Lien L (1981) Biology of the minnow Phoxinus phoxinus and its interactions with brown trout Salmo trutta in Øvre Heimdalsvatn, Norway. Ecography 4:191–200
Miró A, Ventura M (2015) Evidence of exotic trout mediated minnow invasion in Pyrenean high mountain lakes. Biol Inv 17:791–803
Miró A, Sabás I, Ventura M (2018) Large negative effect of non-native trout and minnows on Pyrenean lake amphibians. Biol Cons 218:144–153
Museth J, Borgstrøm R, Hame T, Holen LÅ (2003) Predation by brown trout: a major mortality factor for sexually mature European minnows. J Fish Biol 62:692–705
Museth J, Hesthagen T, Sandlund OT, Thorstad EB, Ugedal O (2007) The history of the minnow Phoxinus phoxinus (L.) in Norway: from harmless species to pest. J Fish Biol 71:184–195
Museth J, Borgstrøm R, Brittain JE (2010) Diet overlap between introduced European minnow (Phoxinus phoxinus) and young brown trout (Salmo trutta) in the lake, Øvre Heimdalsvatn: a result of abundant resources or forced niche overlap? In: Brittain JE, Borgstrøm R (eds) The subalpine lake ecosystem, Øvre Heimdalsvatn, and its catchment: local and global changes over the last 50 years. Springer, Dordrecht, pp 81–91
Tiberti R, Buscaglia F, Armodi M, Callieri C, Ribelli F, Rogora M, Tartari G, Bocca M (2020) Mountain lakes of Mont Avic Natural Park: ecological features and conservation issues. J Limnol 79:43–58
Tiberti R, Buchaca T, Boiano D, Knapp R, Pou Rovira Q, Tavecchia G, Ventura M, Tenan S (2021) Alien fish eradication from high mountain lakes by multiple removal methods: estimating residual abundance and eradication probability in open populations. J Appl Ecol 58:1055–1068
Ventura M, Tiberti R, Buchaca T, Buñay D, Sabás I, Miró A (2017) Why should we preserve fishless high mountain lakes? In: Catalan J, Ninot JM, Aniz M (eds) High mountain conservation in a changing world. Springer, Cham, pp 181–205
CEN (2015) Water quality sampling of fish with multi-mesh gillnets. European standard. European Committee for Standardization. Ref. No. EN 14757:2015
Global Invasive Species Database 2021. Species profile. Phoxinus phoxinus. http://www.iucngisd.org/gisd/speciesname/Phoxinus+phoxinus Accessed 16 Jun 2021
Acknowledgements
We thank the authorities, the scientific staff and the park wardens of the Gran Paradiso National Park, Mont Avic Natural Park, Aiguestortes and Estany de Sant Maurici National Park, Alt Pireneu Natural Park for providing logistic support and for their commitment for the conservation of mountain lakes. We also thank several field assistants, students, and collaborators for their help during the fieldwork.
Funding
Open access funding provided by Università degli Studi di Pavia within the CRUI-CARE Agreement. Funding was provided by the EU Projects LIFE BIOAQUAE (LIFE11 BIO/IT/000020) and LIFE LIMNOPIRINEUS (LIFE13 NAT/ES/001210), LIFE RESQUE ALPYR (LIFE20 NAT/ES/000369), MONOCLE (H2020-CLIMATE/0357), and Biodiversa FISHME (BiodivRestor-280), and by the Spanish Government projects FUNBIO (RTI2018-096217-B-I00) and BIOOCULT (2413/2017). Annual permits for fish sampling have been delivered by the two parks.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
None of the authors declares a conflict of interest.
Consent to participate
All authors expressed their consent to participate in the paper.
Consent for publication
All authors agree with the paper contents and with its publication in biological invasions.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tiberti, R., Buchaca, T., Cruset Tonietti, E. et al. Minnow introductions in mountain lakes result in lower salmonid densities. Biol Invasions 24, 2285–2289 (2022). https://doi.org/10.1007/s10530-022-02783-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-022-02783-w