
Abstract
Objectives
A novel β-carotene-9,10′-oxygenase (ScBCO2) has been characterized from Saccharomyces cerevisiae ULI3 to convert β-carotene to β-apo-10′-carotenal, which is a precursor of the plant hormone strigolactone.
Results
The ScBCO2 enzyme was purified to homogeneity by ammonium sulfate precipitation, Q sepharose and Superdex-200 chromatography. The molecular mass of the enzyme was ~50 kDa by SDS-PAGE. The purified ScBCO2 enzyme displayed optimal activity at 45 °C and pH 8. Tween 20 (1%, w/v), Trition X-100 (1%, w/v), Mg2+ (5 mM), Zn2+ (5 mM), Cu2+ (5 mM), Ca2+ (5 mM) or DTT (5 mM) increased in the activity by 3, 7, 14, 17, 23, 26 and 27%, respectively. ScBCO2 only exhibited cleavage activity towards carotenoid substrates containing two β-ionone rings and its catalytic efficiency (kcat/Km) followed the order β-carotene > α-carotene > lutein.
Conclusion
ScBCO2 could be used as a potential candidate for the enzymatic biotransformation of β-carotene to β-apo-10′-carotenal in biotechnological applications.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
β-Carotene is an isoprenoid pigment occurring naturally in plants and fruits, and serves as the major dietary source of provitamin A. The in vivo cleavage of β-carotene can occur via two different biochemical pathways (Leuenberger et al. 2001; During and Harrison 2004). The first of these pathways, which is catalyzed by β-carotene 15,15′-monooxygenases (BCO1) in mammals (Lindqvist and Andersson 2002; Kim et al. 2011) and marine bacteria (Kim et al. 2009), involves the symmetrical cleavage of β-carotene into two molecules of retinal. The second pathway is an unusual and unsymmetrical cleavage process that occurs at the double bonds of β-carotene to produce long-chain (>C20) apocarotenoids, which are subsequently converted to retinal (Kim et al. 2009). Apocarotenoids function as antioxidants, hormones, volatile aromas and flavors, provitamin A and retinoid regulators (Walter and Strack 2011).
β-Apo-10′-carotenal (C27), which is one product of the asymmetric cleavage of β-carotene (apocarotenoids), is a precursor of strigolactone that acts as a novel plant hormone by promoting the seed germination and stimulating colonization of roots by symbiotic arbuscular mycorrhizal fungi (Akiyama et al. 2005). β-Apo-10′-carotenal could be converted from β-carotene by β-carotene-9,10′-oxygenase (BCO2) (Scherzinger and Al-Babili 2008), which cleave the C9-C10 double bond. Although BCO2 activities have been found in several higher organisms, including humans (Lindqvist and Andersson 2002), mice (Kiefer et al. 2001), ferrets (Hu et al. 2006) and cattle (Tian et al. 2009), there have been no reports in the literature to date pertaining to the isolation of an enzyme from a microorganism that is capable of converting β-carotene to β-apo-10′-carotenal. In this study, we have isolated and biochemically characterized a novel β-carotene-9,10′-oxygenase (ScBCO2) from Saccharomyces cerevisiae ULI3 and demonstrated that this enzyme could potentially be used for the enzymatic biotransformation of β-carotene to β-apo-10′-carotenal.
Materials and methods
Chemicals
α-Carotene, β-carotene, γ-carotene, zeaxanthin, lutein, lycopene and lutein were purchased from Pioneer Biotech Co., Ltd. (Xian, China). Unless otherwise stated, all of the chemicals used in the current study were purchased as the analytical grade.
Organism and culture conditions
Saccharomyces cerevisiae ULI3 was isolated from fresh tobacco leaves, which were collected from Kunming, China. ScBCO2 was produced in complex medium (CM) containing (g/l): 1.5 g β-carotene, 3 g NaNO3, 0.5 g MgSO4, 1 g K2HPO4, 0.01 g FeSO4, 0.5 g KCl, 30 g sucrose and 3 g yeast extract power. The ULI3 strain was grown in a 500 ml flask containing 125 ml medium at 28 °C for 72 h with shaking at 150 rpm. The culture broth was collected and centrifuged at 6000×g for 20 min, and the supernatant was used as the crude enzyme for subsequent experiments.
Purification of ScBCO2 from strain ULI3
The crude enzyme was fractionated by (NH4)2SO4 precipitation, where 60–80 % saturation with (NH4)2SO4 gave a precipitate, which was allowed to stand overnight. The precipitate was then collected by centrifugation at 12,000×g and suspended in 20 mM phosphate buffer (pH 8.0) before being loaded onto a Q-Sepharose column (1 × 20 cm2), which had been equilibrated with 20 mM phosphate buffer (pH 8.0). The proteins were eluted with a 100 ml of a linear gradient of NaCl from 0 to 1 M at 1 ml/min. Fractions were collected and tested for β-carotene degradation. Active fractions were pooled, filter concentrated, dialyzed in 50 mM phosphate buffer (pH 8) containing 200 mM NaCl and loaded onto a Superdex-200 (16/60) column, which had been equilibrated with 50 mM phosphate buffer (pH 8). All of the purification steps were carried out at 4 °C. The purified protein was analyzed by SDS-PAGE and the protein concentration was determined according to the Bradford method. Protein samples were stored in 20 mM phosphate buffer (pH 8.0) containing 25 % (v/v) glycerol at −80 °C.
Enzyme assay
The reaction solution contained 200 mM NaCl, 12.5 μM Fe2SO4, 10 mM Tris (2-carboxylethyl) phosphine hydrochloride and 1.5 % (w/v) 1-S-octyl-β-D-thioglucopyranoside, as described previously (Kim et al. 2008). The enzyme and substrate solutions were mixed at 3:1 (v/v) and the reaction mixture was held at 37 °C for 60 min in 100 mM Tricine/KOH buffer (pH 8.0) containing 60 mM β-carotene, 1.5 % (w/v) Tween 40 and 0.3 U enzyme/ml. One unit of enzymatic activity was defined as the amount of enzyme required to liberate 1 mol β-apo-10′-carotenal per min under the standard conditions. The measurements were corrected for background hydrolysis in the absence of the enzyme.
Analysis of HPLC and LC–MS
The ScBCO2-catalyzed conversion of β-carotene to β-apo-10′-carotenal was followed by HPLC using a Zorbax RX-C18 column (4.6 × 250 mm, 5 μm particle size, Agilent Technologies). The column was eluted with methanol/water (7:3 v/v) containing 0.1 % ammonium acetate (solvent B) and methanol (solvent A). The gradients were as follows: 100 % B to 0 % B over 12 min; 0 % solvent B for 8 min; and then 100 % A for 8 min at a flow rate of 0.5 ml/min. The reaction products were characterized by LC–MS. For quantification, a curve correlating peak area to moles of the reaction product was attained with β-apo-10′-carotenal as standard sample. A concentrated stock was prepared by dissolving β-apo-10′-carotenal in chloroform at 100 mg/l. This was diluted in chloroform to generate standards at 1, 2, 3, and 4 mg/l. The stocks were stored at −20 °C. Sample and standards were analyzed using an LTQ XL ultra-HPLC-ion trap MS equipped with an atmospheric pressure chemical ionization ion source (Thermo Electron Corp, USA). Mass fragmentation spectra were monitored for masses in the range of 50–800 atomic mass units on the LC–MS system. The chromatography conditions used for the LC–MS analysis were identical to those described above for the HPLC analysis.
Characterization of ScBCO2
The effects of different temperatures, pH values, metal ions, organic solvents and reagents on the ScBCO2 activity were examined. The kinetic parameters were determined by assaying the purified enzyme at a variety of different substrate concentrations (10–800 μM) in three independent trials, and the corresponding Km, Vmax and kcat values were computed using Hanes–Woolf plots and the Michaelis–Menten equation.
Analysis of purified enzyme
Protein bands were excised from SDS-PAGE, digested by trypsin and the sample was subjected to MALDI-TOF MS using an Applied Biosystems 4700 proteomics analyzer (Applied Biosystems, CA, USA). Data analysis was performed using the GPS Explorer software and MASCOT with the NCBI database.
Results and discussion
Purification and identification of ScBCO2
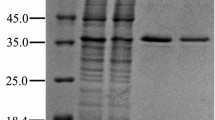
The ScBCO2 enzyme with β-carotene cleavage activity was purified to homogeneity from the culture supernatant of the ULI3 strain. The purification procedure is shown in Table 1. The enzyme was purified 9.5-fold to give a yield of 10.8 % and a specific activity of 0.38 U/mg. The molecular mass of the purified enzyme was estimated to be 50 kDa by SDS-PAGE (Fig. 1), which is therefore less than those of the BCO2 enzymes isolated from mice (Kiefer et al. 2001), humans (Lindqvist and Andersson 2002), ferrets (Hu et al. 2006) and cattle (Tian et al. 2009). After being separated by SDS-PAGE, the ScBCO2 enzyme was analyzed by MALDI-TOF-MS, and the results showed that the peptide mass fingerprint of this enzyme did not fit with any of the other peptides in the database (data not shown). These results therefore indicated that the ScBCO2 enzyme from S. cerevisiae ULI3 could be a novel BCO2 oxygenase.

SDS-PAGE of the purified ScBCO2 enzyme from S. cerevisiae ULI3. Lane 1 molecular weight standards; lane 2 Superdex-200 chromatography product (purified enzyme)
Identification of the reaction product of ScBCO2
LC–MS analysis of the enzyme reaction product revealed m/z values of 193 and 377, which corresponded to the quasimolecular ions of authentic β-ionone and β-apo-10′-carotenal samples, respectively (Supplementary Fig. 1a, b). These results therefore demonstrated that ScBCO2 could convert β-carotene to β-apo-10′carotenal and β-ionone as a consequence of its unusual cleavage activity towards the 9′,10′ double bond of β-carotene (Fig. 2). β-Apo-10′-carotenal is a precursor of the plant hormone strigolactone, which could promote seed germination and stimulate the colonization of plant roots by arbuscular mycorrhizal fungi (Akiyama et al. 2005). β-Ionone is a flavor and fragrance compound with fruity, violet-like characteristics and used in particular by foodstuff and beverage industries (Nacke et al. 2012). Some oxygenases in higher organisms exhibit high levels of unusual cleavage activity towards β-carotene, resulting in the production of β-apo-10′-carotenal (Kiefer et al. 2001; Lindqvist et al. 2005; Hu et al. 2006; Tian et al. 2009). There have, however, been no reports pertaining to the isolation and characterization of an enzyme from a microorganism that can convert β-carotene to β-apo-10′-carotenal. To the best of our knowledge, ScBCO2 therefore represents the first reported oxygenase to have been purified and characterized from a yeast source with unusual cleavage activity towards the production of β-apo-10′-carotenal.

ScBCO2-catalyzed cleavage of β-carotene to give β-apo-10′-carotenal and β-ionone
Effect of temperature and pH on enzyme activity
The standard curve of the β-apo-10′-carotenal to quantify the product of the ScBCO2-catalyzed conversion of β-carotene is shown in Supplementary Fig. 2. The enzyme exhibited maximal activity ~45 °C with more than half of its maximal activity at 30–50 °C (Fig. 3a). It was optimally active at pH8 (Fig. 3b). Over 50 % of the maximal activity was maintained between pH 7 and 9. (Figure 3b).

Temperature (a) and pH (b) optima of the ScBCO2 enzyme. a The optimum temperature for the ScBCO2 enzyme was determined using β-carotene as a substrate in 50 mM Tricine/KOH buffer (pH 8.0) at temperatures in the range of 25–65 °C. The activity at 45 °C (optimum temperature) was taken to be 100 % (0.3 U/mg). b The optimum pH for the ScBCO2 enzyme was determined at pH values in the range of 5.5–10.0 using β-carotene as a substrate for 60 min at 45 °C. The following buffer systems were used: 50 mM sodium acetate (pH 5.5 to 6.0), sodium phosphate (pH 6.5 to 8.0), Tris–HCl (pH 8.5 to 9.0) and N-cyclohexyl-3-aminopropanesulfonic acid (pH 9.5 to 10.0). The activity at pH 8.0 (optimum pH) was taken to be 100 % (0.3 U/mg). The relative activity was calculated by defining original activity as 100 %. The reported values are mean values of three independent experiments
Effects of different metal ions, organic solvents and surfactants on the enzyme activity
Mg2+, Zn2+, Cu2+ or Ca2+ (5 mM) increased in the activity of ScBCO2 by 14, 17, 23 and 26 %, respectively, whereas Mn2+ or Fe2+ decreased the activity by 13 and 37 %, respectively (Table 2). The enzyme was not inhibited by 5 mM EDTA, which indicated that it was not a metalloprotein. The addition of Tween 20, Triton-X100 or DTT led to a slight increase in activity, whereas SDS (1 %) and urea (4 M) strongly inhibited the activity of ScBCO2 to less than 20 % of its optimal activity.
As shown in Table 3, the ScBCO2 enzyme was active in a number of organic solvents. Organic solvents are used in many industrial biocatalysts and therefore the effect of organic solvents on the activity of this enzyme is of special interest. ScBCO2 retained more than 70 % of its activity in 50 % (v/v) toluene, acetone, benzene, DMF or DMSO.
Carotenoid substrate specificity of ScBCO2
Enzymatic activity was detected for α-carotene, β-carotene and lutein, but not for γ-carotene, zeaxanthin and lycopene (Supplementary Table 1). The kcat/km values for ScBCO2 followed the order β-carotene (77 min−1 mM−1) > α-carotene (49 min−1 mM−1) > lutein (10 min−1 mM−1) (Table 4). α-Carotene, β-carotene and lutein have two β-ionone rings in their structure, whereas γ-carotene and zeaxanthin contain only one β-ionone ring. In contrast, lycopene does not contain a β-ionone ring. Taken together, these results suggest that ScBCO2 has a stronger affinity and higher catalytic efficiency for substrates containing two β-ionone rings than any other substrates.
Conclusion: A novel β-carotene-9,10′-oxygenase from S. cerevisiae ULI3 has been purified and biochemically characterized for the first time. The molecular mass of the purified enzyme was ~50 kDa by SDS-PAGE; ptimum activities were at 45 °C and pH 8.0. The enzyme showed high cleavage activity towards β-carotene to form β-apo-10′-carotenal and β-ionone. These results therefore demonstrate that this enzyme could potentially be used for the synthesis of apocarotenoids in biotechnological applications.
References
Akiyama K, Matsuzaki K, Hayashi H (2005) Plant sesquiterpenes induce hyphal branching in arbuscular mycosrhizal fungi. Nature 435:824–827
During A, Harrison EH (2004) Intestinal absorption and metabolism of carotenoids: insights from cell culture. Arch Biochem Biophys 430:77–88
Hu KQ, Liu C, Ernst H, Krinsky NI, Russell RM, Wang XD (2006) The biochemical characterization of ferret carotene-9′,10′-monooxygenase catalyzing cleavage of carotenoids in vitro and in vivo. J Biol Chem 281:19327–19338
Jia BL, Wei T, Huang S, Jia CX, Yang J, Zhang GH, Bai B, Mao DB (2015) Isolation and identification of a α-carotenes-degradating strain and optimization of fermentation conditions. Food Fermn Ind 41:34–39
Kiefer C, Hessel S, Lampert JM, Vogt K, Lederer MO, Breithaupt DE, von Lintig J (2001) Identification and characterization of a mammalian enzyme catalyzing the asymmetric oxidative cleavage of provitamin A. J Biol Chem 276:14110–14116
Kim NH, Kim YS, Kim HJ, Oh DK (2008) Optimized formation of detergent micelles of β-carotene and retinal production using recombinant human β, β-carotene 15,15′-monooxygenase. Biotechnol Prog 24:227–231
Kim YS, Kim NH, Yeom SJ, Kim SW, Oh DK (2009) In vitro characterization of a recombinant Blh protein from an uncultured marine bacterium as a β-carotene 15,15′-dioxygenase. J Biol Chem 284:15781–15793
Kim YS, Yeom SJ, Oh DK (2011) Production of β-apo-10′-carotenal from β-carotene by human β-carotene-9′,10′-oxygenase expressed in E. coli. Biotechnol Lett 33:1195–1200
Leuenberger MG, Engeloch-Jarret C, Woggon WD (2001) The reaction mechanism of the enzyme-catalyzed central cleavage of β-carotene to retinal. Angew Chem Int Ed 40:2613–2617
Lindqvist A, Andersson S (2002) Biochemical properties of purified recombinant human beta-carotene 15,15′-monooxygenase. J Biol Chem 277:23942–23948
Nacke C, Huttmann S, Etschmann MMW, Schrader J (2012) Enzymatic production and in situ separation of natural beta-ionone from beta-carotene. J Ind Microbiol Biotechnol 39:1771–1778
Scherzinger D, Al-Babili S (2008) In vitro characterization of a carotenoid cleavage dioxygenase from Nostoc sp. PCC 7120 reveals a novel cleavage pattern, cytosolic localization and induction by highlight. Mol Microbiol 69:231–244
Scherzinger D, Ruch S, Kloer DP, Wilde A, Al-Babili S (2006) Retinal is formed from apo-carotenoids in Nostoc sp. PCC7120: in vitro characterization of an apo-carotenoid oxygenase. Biochem J 398:361–369
Tian R, Pitchford WS, Morris CA, Cullen NG, Bottema CD (2009) Genetic variation in the beta, beta-carotene-9′,10′-dioxygenase gene and association with fat colour in bovine adipose tissue and milk. Anim Genet 41:253–259
Walter MH, Strack D (2011) Carotenoids and their cleavage products: biosynthesis and functions. Nat Prod Rep 28:663–692
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (21406210 and 21476216) and the Innovation Scientists and Technicians Troop Construction Projects of the Henan Province (144200510011).
Supporting information
Supplementary Table 1 – Carotenoid substrate specificity of ScBCO2.
Supplementary Figure 1 – LC-MS analysis of the reaction product catalyzed by ScBCO2.
Supplementary Figure 2 – The standard curve of the β-apo-10′-carotenal to quantify the product of the ScBCO2-catalyzed conversion of β-carotene.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Wei, T., Jia, B., Huang, S. et al. Purification and characterization of a novel β-carotene-9′,10′-oxygenase from Saccharomyces cerevisiae ULI3. Biotechnol Lett 37, 1993–1998 (2015). https://doi.org/10.1007/s10529-015-1872-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10529-015-1872-7