
Abstract
Many native South African grass species have become invasive elsewhere in the world. The application of biological control to invasive grasses has been approached with trepidation in the past, primarily due to concerns of a perceived lack of host specific herbivores. This has changed in recent times, and grasses are now considered suitable candidates. The Tetramesa Walker genus (Hymenoptera: Eurytomidae) has been found to contain species that are largely host specific to a particular grass species, or complex of closely related congeners. Very little taxonomic work exists for Tetramesa in the southern hemisphere, and the lack of morphological variability between many Tetramesa species has made identification difficult. This limits the ability to assess the genus for potential biological control agents. Species delimitation analyses indicated 16 putative novel southern African Tetramesa taxa. Ten of these were putative Tetramesa associated with Eragrostis curvula (Schrad.) Nees and Sporobolus pyramidalis Beauv. and S. natalensis Steud., which are alien invasive weeds in Australia. Of these ten Tetramesa taxa, eight were only found on a single host plant, while two taxa were associated with multiple species in a single grass genus. The Tetramesa spp. on S. pyramidalis and S. africanus were deemed suitably host-specific to be used as biological control agents. Field host range data for the Tetramesa species on E. curvula revealed that the wasp may not be suitably host specific for use as a biological control agent. However, further host specificity testing on non-target native Australian species is required.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
South Africa has been a major donor of invasive grasses to other parts of the world due to the perception that African C4 grasses were more palatable to livestock and resistant to grazing than native grasses (Visser et al. 2016). Many African grasses have subsequently become invasive and highly damaging due to the combination of favourable traits, multiple introduction events (D’Antonio and Vitousek 1992; Visser et al. 2016), and high propagule pressure (Firn 2009). Historically, very few grasses have been targeted using biological control, mainly due to concerns of potential non-target attacks on economically valuable crops, tolerance to herbivory, and the belief that there were few or no specialist herbivores on grasses (Sutton et al. 2021a). These perceptions are, however, changing with the discovery of host-specific taxa (Goolsby et al. 2020), particularly eurytomid wasps in the Tetramesa Walker (Hymenoptera: Eurytomidae) genus (Witt and McConnachie 2004; Sutton et al. 2021a,b).
At the time of the review of grass biological control by Sutton et al. (2021a), 23 invasive grass species were currently, or had already been, investigated as targets for biological control using herbivorous arthropods and fungal pathogens. The global weed biological control catalogue listed 14 releases of seven biological control agent species onto five invasive grass species between 1997 and 2019 (Winston et al. 2021; Supplementary Table S1). To date, T. romana Walker is the only Tetramesa species that has been used as a biological control agent for an alien invasive grass, namely Arundo donax L. (giant reed or carrizo cane) in the USA (Goolsby and Moran 2009; Moran and Goolsby 2009). Tetramesa romana is considered successful in the USA, particularly in the Rio Grande, Texas, where A. donax biomass was estimated to be reduced by 32% in 2016, equating to 4.4 million $US a year in water savings (Goolsby et al. 2016; Moran et al. 2017). Following the success of this programme, there has been an increasing interest in the use of Tetramesa as potential biological control agents for other alien invasive grasses (Lotfalizadeh et al. 2020; Sutton et al. 2021b).
The Tetramesa genus currently comprises over 200 described species (Al-Barrak 2006; Lotfalizadeh et al. 2020; Natural History Museum 2021). They are highly host-specific, and typically specialise on only one species or genus, or on closely related genera, in the Poaceae (Phillips 1936; Claridge 1961). The larvae are endophagous feeders, and are either stem borers or gallers (Claridge 1961). Almost all of the sampling effort in collecting and describing Tetramesa species has taken place in the Northern Hemisphere (Al-Barrak 2006) (Supplementary Fig. S1). Only four African species have been described, namely T. aristidae Risbec from Senegal, T. decaryi Risbec and T. tananarivense Risbec from Madagascar, and T. macalusoi De Stefani from Somalia (van Noort 2020). No Tetramesa species have been described yet from South Africa (van Noort 2020), despite the existence of novel taxa in the region (Witt and McConnachie 2004; Sutton et al. 2021b).
In addition to T. romana, a number of other Tetramesa species are currently being considered as potential grass biological control agents. Witt and McConnachie (2004) reported the presence of an unidentified Tetramesa sp. on Sporobolus africanus (Poir.) Robyns and Tournay, S. natalensis (Steud.) T. Durand and Schinz, and S. pyramidalis P. Beauv. in southern Africa. A second unidentified Tetramesa species was found on S. natalensis and S. pyramidalis by Sutton et al. (2021b) during native range surveys, where the wasp was found on only these two target weeds and not on any native congeners or other close relatives. Sutton et al. (2021b) found that both Tetramesa species significantly decreased tiller survival and had a deleterious impact on tiller reproduction. There are several other grasses that are indigenous to South Africa that are potential targets for biological control that may have other Tetramesa species associated with them, particularly Eragrostis curvula (Schrad.) Nees and Andropogon gayanus Kunth (Olckers et al. 2021; Sutton et al. 2021b).
The Centre for Biological Control (CBC) at Rhodes University, South Africa, has been conducting field surveys for natural enemies on native grasses since 2017 across more than 200 sites and 70 grass species. The morphological uniformity of the adult and larval stages of the Tetramesa makes it extremely challenging to identify different species (Dawah 1987; Ghajarieh et al. 2006). Even genus-level identifications delineating between the Tetramesa and Eurytoma have proven difficult (Henneicke et al. 1992), and reliable synapomorphies at even the family-level are still lacking (Lotfalizadeh et al. 2007; Gates 2008). The lack of a taxonomic backbone for Tetramesa in South Africa and suitable morphological tools to distinguish between difference species has impeded our ability to assess the host specificity of field collected specimens and to establish pure laboratory cultures. The aims of the present work were therefore to use genetic techniques to distinguish between the Tetramesa species, and from this make inferences about their predicted host ranges. Delineating species in this morphologically cryptic genus will advance efforts to identify potential biological control agents for the African grasses that have become invasive elsewhere.
Materials and methods
Sample collection
Wasp samples for DNA analysis were collected from six South African provinces and 19 grass host plants (Supplementary Fig. S2, Fig. S3, Table S2, Table S3). Surveys were performed through either stem dissections of collected grasses or emergence chambers in the laboratory to ensure that the host-plant record was a true interaction and not due to chance. Voucher specimens were identified to the lowest taxonomic level by experts at the South African National Insect Collection in Pretoria (ARC-PPRI).
Host grasses were selected for sampling based on their invasive status and prioritisation in biological control programmes in Australia and the USA. The main targets were Andropogon gayanus, Eragrostis curvula, Megathyrsus maximus (= Panicum maximum) (Jacq.) B.K. Simon and S.W.L. Jacobs, Sporobolus natalensis, and S. pyramidalis, all of which are serious alien invasive pests in Australia and the USA. Hyparrhenia hirta (L.) Stapf was included because this grass is also a problem species in Australia and might be considered for a future biological control programme (Sutton et al. 2019).
Sample preservation and DNA extraction
All insect specimens were stored in 95% ethanol at − 20 °C. Reference specimens were photographed under a ZEISS Stemi 508 stereo microscope using ZEN imaging software [ZEN Digital Imaging for Light Microscopy (RRID:SCR 013672)], and CombineZP software for z-stacking. A full database of photographs is housed on the associated GitHub project repository (https://github.com/clarkevansteenderen/PhD_files/tree/main/Microscope%20Images). Three legs from each specimen were removed under a dissecting microscope so that the remaining whole specimens could be used for direct comparisons after sequencing. Genomic DNA extractions were performed using a PureLink™ Genomic DNA Mini Kit (ThermoFisher Scientific, catalogue number K182002) according to the manufacturer’s protocols. Supplementary File S1 provides details of the polymerase chain reaction (PCR) protocols, nucleotide sequencing and alignments, phylogenetic analyses, and Generalised Mixed Yule Coalescent (GMYC) species delimitation analysis.
Results
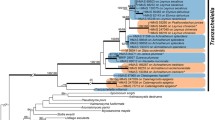
Both the COI (log-likelihood of the ML consensus tree = − 10200.798, BI run 1 = − 10641.18, BI run 2 = − 10661.60) and 28S (log-likelihood of the ML consensus tree = − 3840.301, BI run 1 = − 4097.81, BI run 2 = − 4115.02) phylogenies (see clade name abbreviations in Supplementary Table S4) showed a distinct separation between pronotal spot (PNS) and non-pronotal spot (NPNS) eurytomids, but the NPNS groups were unresolved (Figs. 1, 2, Supplementary Fig. S5, Fig. S6, Fig. S7).

Condensed Bayesian inference (BI) phylogeny for the 28S nuclear gene region. Posterior probability support values are shown as filled circles on the branches (see the legend). Supplementary Fig. S5 shows the full phylogeny with all the sample names displayed, and GMYC species delimitation results included in the figure. See the list of phylogenetic clade abbreviations in Supplementary Table S2

Condensed Bayesian inference (BI) phylogeny for the COI mitochondrial gene region. Posterior probability support values are shown as filled circles on the branches (see the legend). Supplementary Fig. S6 shows the full phylogeny with all the sample names displayed, and GMYC species delimitation results included in the figure. See the list of phylogenetic clade abbreviations in Supplementary Table S2
28S data
Overall inter-and intra-specific p-distances (excluding the outgroup) ranged from 3 to 9.7% and 0 to 2.7%, respectively. Sequence divergences within the southern African PNS groups ranged from 0.6 to 1.9% (Supplementary Table S5 and Fig. S8). The only three well-defined clades within the PNS group were the wasps collected from Eragrostis rigidior (PNS.ERIG.1; clade 1, posterior probability (pp) = 0.71), Sporobolus spp. (PNS.SPOR.1; clade 2, pp = 0.84, bootstrap support (bt) = 91), and Arundo donax (PNS.ADON.1, clade 3, pp = 1, bt = 100) (Fig. 1, Supplementary Fig. S5). The nuclear marker showed that both the Eurytominae subfamily and the proposed Tetramesa formed well-supported monophyletic groups (Fig. 1, Supplementary Fig. S5, pp = 1). These results suggest that a sequence divergence of ∼ 3 to 3.5% and above, likely does not belong to the Tetramesa genus (Supplementary Table S5), and that major groups within the PNS clade can be delineated at a threshold of below 0.4%. The only NPNS wasps that fell within the proposed Tetramesa group were those collected from H. hirta (NPNS.HHIR.1) and A. gayanus (NPNS.AGAYA.1) (Fig. 1, Supplementary Fig. S5). The GMYC species delimitation method estimated 36 entities (95% CI = 20 – 43) and 23 clusters (95% CI = 15 – 28) (log-likelihood of the null model = 957.25, ML of the GMYC model = 963.6845, p < 0.01, singletons = 13). Guided by the GMYC results, we estimated the presence of six host-specific taxa, namely PNS.ERIG.1, PNS.SPOR.1, PNS.ADON.1, NPNS.HHIR.1, NPNS.AGAYA.1, and NPNS.HHIR.2 (Fig. 1, Supplementary Fig. S5, Table S6).
COI data
Overall inter-group and intra-group p-distances (excluding the outgroup) ranged from 1.6 to 26.3% and 0 to 3.2% respectively. Chen et al. (2004) reported interspecific distances ranging from 0.2 to 25.8%. Most of the clades in the COI phylogeny were unresolved, rendering taxonomic inferences based on this marker very challenging (Fig. 2, Supplementary Fig. S6). Sequence divergences within the southern African PNS groups ranged from 2.4 to 13.1% (Supplementary Fig. S9). A sequence divergence of 2.8% was found between the PNS.ECUR.1 (PNS wasps on E. curvula) clade and its sister group PNS.ETRICH.1 (PNS wasps on E. trichophora) (Fig. 2, Supplementary Fig. S6), where the latter forms unique galls on E. trichophora that have not been observed on other grasses (Supplementary Fig. S10).
There were four separate clades associated with E. curvula within the PNS group (PNS.ECUR.1 PNS.ECUR.4), although only PNS.ECUR.1 (pp = 0.68, bt = 95) and PNS.ECUR.4 (pp = 1, bt = 100) were strongly supported (Fig. 2, Supplementary Fig. S6). The sequence divergence between PNS.ECUR.1 (Eastern Cape) and PNS.ECUR.2 (Free State) was 4.5% (Fig. 2, Supplementary Fig. S6, Fig. S9). The two E. rigidior clades, PNS.ERIG.1 and PNS.ERIG.2, were collected in the same geographic region, and had a sequence divergence of 7.1% (Supplementary Fig. S9).
The wasps collected on Sporobolus pyramidalis and S. africanus formed two distinct sister clades, with a sequence divergence of 3.9% (Fig. 2, Supplementary Fig. S6; PNS.SPYR.1; pp = 1, bt = 89 and PNS.SAFR.1; pp = 0.73, bt = 99). Additionally, the S. pyramidalis wasps showed some evidence of geographic substructuring between the KwaZulu-Natal (pp = 0.9, bt = 97) and Eastern Cape (pp = 0.88, bt = 95) specimens. The GMYC results estimated 71 entities (95% CI = 62–77) and 42 clusters (95% CI 36–43) (log-likelihood of the null model = 1026.2, ML of the GMYC model = 1082.666, p < 0.001, singletons = 29). Based on the phylogenetic and GMYC analyses, we estimated the presence of thirteen host-specific taxa, namely PNS.ETRICH.1, PNS.EPLAN.1, PNS.ECUR.3, PNS.ERIG.2, PNS.ECUR.4, PNS.SPYR.1, PNS.SAFR.1, NPNS.ECUR.2, NPNS.HHIR.1, NPNS.HHIR.2, NPNS.AGAYA.1, NPNS.HHIR.3, and PNS.ADON.1 (Fig. 2, Supplementary Fig. S6, Table S6).
Discussion
In general, very little taxonomic work has focused on Afro-tropical insects, particularly micro-Hymenoptera such as the Tetramesa (van Noort et al. 2015; Berry and van Noort 2016; Hopkins et al. 2019). This investigation has provided new insights into the Tetramesa assemblages on native African grasses by investigating their diversity, phylogenetic relationships, and host specificity. The work presented here has identified multiple Tetramesa taxa that hold potential as biological control agents of invasive grasses, but some groups may be more suitably host-specific than others, and care must be taken to use an integrative taxonomic approach rather than relying solely on one line of evidence for identification.
Biological control outlook
The NPNS clades were found largely to be oligophagous feeders, and had a wider host range compared to the PNS groups. Biological control efforts should therefore prioritise the latter, and those that form part of the NPNS H. hirta and Andropogon gayanus clade, as well as PNS T. romana. This is based on the host-use patterns seen particularly in the COI phylogeny, where, with a few minor exceptions, each PNS clade was associated with a single host grass. The prospects for the biological control of the various target weeds are discussed below in the context of the phylogenies and inferences of host ranges. A promising species for M. maximus was found, but preliminary host rearing trials have indicated that this wasp can reproduce on Setaria sphacelata (Schumach.) Stapf and C.E. Hubb (unpublished data, Guy Sutton).
Eragrostis
The PNS E. curvula wasps (morphospecies ‘Tetramesa sp. 4’) belonged to the Tetramesa genus, but are likely not good candidates for biological control due to their broader host range on conspecific Eragrostis species. These could include non-target Australian and North American natives, that are either more closely related to the target weed than to the congeners from which they were collected, or very closely related to the congeners.
The nine PNS clades associated with Eragrostis hosts identified in the present COI phylogeny (spanning E. curvula, E. biflora, E. rigidior, E. trichophora, and E. plana) were delineated into eight GMYC taxa, and may represent unique species. Further taxonomic confirmation is, however, required to validate this observation. The low sequence divergence between the wasps on E. curvula clades PNS.ECUR.1 – PNS.ECUR.3 and E. trichophora PNS. ETRICH.1 (ranging from p-distances of 2.4 – 3.9%) suggests that host-specificity testing of these two wasps is important, as each taxon may be able to utilise both host plants, which could preclude their use as biological control agents. No-choice host-specificity testing should be completed, as it seems unlikely, although not impossible, that the same species of wasp would have such different feeding modes on two different host plant species, namely galling E. trichophora and mining the stems of E. curvula. Tetramesa sp. 26 on E. rigidior (PNS.ERIG.1) may not be an appropriate biological control agent, as it was also recorded on E. gummiflua in the field and therefore has a relatively broad host range. The second Tetramesa sp. 26 clade (PNS.ERIG.2), however, may be useful as it was specific to only E. rigidior. Further sampling focusing on these species is required.
Further work should focus on a population genetics analysis of both the PNS E. curvula Tetramesa wasps and their hosts from a wider geographic range to test whether host plant form may be driving genetic divergence. E. curvula has been developed as a pasture grass and has had numerous varieties developed. Such genetic breeding has led to distinct forms of the grass that vary morphologically and chemically (e.g.? phosphate and fibre concentrations) (Leigh 1961). In the case of different E. curvula haplotypes in the native range, it will be vital to confirm matches to haplotypes in the invaded range to ascertain points of origin and select the most appropriate Tetramesa population(s) as control agents. For example, Harms et al. (2021) identified a novel haplotype of the alien invasive water weed Hydrilla verticillata (L.f.) Royle, and found that the source population was from north-eastern China. This allowed the researchers to conduct targeted herbivore guild collections from this region.
The NPNS E. curvula wasp that was assigned the morphospecies name ‘Tetramesa sp. 5’ does not appear to belong to the Tetramesa genus, based on the present nuclear 28S results. It is possible that the Tetramesa as it currently stands should be divided into two genera: one comprising the predominantly PNS groups, and the other the NPNS clades. This would need to be achieved using an integrative approach, where genetic methods are used in conjunction with expert taxonomists. Both Tetramesa sp. 4 and Tetramesa sp. 5 are currently being used in host-specificity tests on native Australian Eragrostis species, as this will be the ultimate deciding factor in their utility as biological control agents. Tetramesa sp. 4 has already been found in Australia on field-collected E. curuvla (Sutton et al. 2023), making it vital to assess its distribution and possible non-target effects.
Sporobolus pyramidalis
The PNS Sporobolus wasps revealed two potential taxa, namely on S. pyramidalis (PNS.SPYR.1) and S. africanus (PNS.SAFR.1). The GMYC analysis suggested a split of the PNS.SPYR.1 clade into two taxa, but this was an artefact of geographic substructuring (these were from Tetramesa collections in the KwaZulu-Natal and Eastern Cape provinces). Both field-based and no-choice tests have revealed that Tetramesa sp. 1 (PNS.SPYR.1) is not able to utilise S. africanus (Sutton et al. 2021b), but the host specificity of the S. africanus wasps should be investigated further. The Tetramesa in clade PNS.SAFR.1 is a novel group that requires host specificity testing mirroring the work conducted on Tetramesa sp. 1 by Sutton et al. (2021b).
Sutton et al. (2021b) have already found that Tetramesa sp. 1 is suitably host specific and damaging to S. pyramidalis and S. natalensis in the field. Additionally, this wasp has been recently imported into quarantine in Australia for further host specificity testing.
Both the PNS and NPNS Sporobolus wasps (named morphospecies ‘Tetramesa sp. 1’ and ‘Tetramesa sp. 2’) have been shown to be host-specific in the field, where sp. 1 was found to be the more damaging of the two (Sutton et al. 2021b). The present phylogenetic results suggest that the NPNS Tetramesa sp. 2 wasp does not belong to the Tetramesa genus. Tetramesa sp. 2 was identified morphologically as belonging to the Tetramesa genus, but our genetic analyses suggest that they are sufficiently different to be considered a new or different genus. This highlights the need to update the taxonomy of the group.
The present genetic results suggest that the NPNS Tetramesa sp. 2 wasp may be able to feed on E. curvula (two wasp samples out of the nine that were collected on S. pyramidalis), but field host range surveys by Sutton et al. (2021b) have found that the wasp was specific to only S. pyramidalis and S. natalensis. Laboratory-based host specificity tests have also been conducted on over 20 non-target species, including eight native South African Sporobolus species, with no non-target feeding recorded to date (Guy Sutton, unpublished data).
This scenario is an example of why it is imperative to have correctly identified each prospective agent in a biological control programme, and to have a solid understanding of their life histories and interspecific interactions in order to achieve the greatest level of damage. Since Tetramesa sp. 2 may be able to use E. curvula as a marginal host in the field, and that the wasp is less damaging than Tetramesa sp. 1, it might be prudent to only use Tetramesa sp. 1 for biological control.
Hyparrhenia hirta and Andropogon gayanus
The NPNS Tetramesa collected on Hyparrhenia hirta and Andropogon gayanus that formed a sister group to the PNS clade were host-specific and formed a monophyletic clade in both gene trees. This is not surprising, since H. hirta and A. gayanus are phylogenetically closely related, both being in the tribe Andropogoneae (Skendzic et al. 2007), and structurally similar (i.e., both are tall-statured grasses) (Canavan et al. 2019b).
The COI GMYC species delimitation results suggested that this group may comprise three cryptic species, and could be considered as potential biological control agents because they are unique and host specific. Hyparrhenia hirta has naturalised in the USA and many parts of Europe, but it is particularly problematic in Australia, where it has become highly invasive (Chejara et al. 2010). It has also been reported as a problematic weed in wheat fields in Pakistan (Hussain et al. 2004). Similarly, A. gayanus has invaded the northern regions of Australia’s tropical savannas (Rossiter-Rachor et al. 2009). Invaded regions such as these could benefit from the Tetramesa biological control agents identified here, particularly in mitigating intense bushfires and devalued land. Although there were only two specimens in this group, it would be worth conducting additional surveys on A. gayanus in Zimbabwe, as there is likely to be a large degree of species diversity on the grass. Additional field surveys on other close relatives within the Andropogoneae are also required to assess their potential as biocontrol agents.
Southern African Tetramesa phylogenetics
The nuclear 28S rRNA marker was effective in broadly separating what are likely true Tetramesa from other described or undescribed genera, where a suggested sequence divergence of ∼ 3 to 3.5% can be used for genus-level delimitation. Our interspecific p-distances were comparable to Chen et al. (2004), who reported interspecific distances of 1.69–13.5% for their eurytomid data set. Of the four 28S sequences identified as Tetramesa gleaned from GenBank, two fell within the proposed Tetramesa clade (sourced from Chen et al. (2004) and Munro et al. (2011)) while the other two (Gillespie et al. (2005), and a sequence from unpublished material (https://www.ncbi.nlm.nih.gov/nuccore/DQ080114), fell in a NPNS polytomy that was more similar to some Eurytoma sequences, and with a divergence of as much as 7.1% from the T. romana group. This high nuclear divergence suggests that these NPNS GenBank specimens were either misidentified, or that the Tetramesa genus needs to be split using morphological and molecular tools. Similarly, all the COI sequences deposited on BOLD that were identified as Tetramesa (collected in Canada and Germany) showed sequence divergences of nearly 20% compared to T. romana and T. bambusae, which is a clear indication that these specimens likely belong to a different genus. The lack of genetic data across the entire genus is an impediment, and more sequence data is need to revise the genus.
In agreement with Chen et al. (2004), the nuclear 28S rRNA marker is preferred as an initial identification guide, because it yielded well-supported basal nodes and broader-scale taxonomic relationships that will be useful for genus-level taxonomic revisions. The COI marker should be used as second filter to focus on specific target groups identified in the nuclear gene tree, as it provided greater resolution within the PNS group and revealed some potential cryptic species and/or genetically distinct populations. Numerous polytomies, however, were a confounding factor in inferring how these groups are evolutionarily related. Additionally, the COI phylogeny did not produce the same monophyletic Tetramesa group as was seen using the 28S marker, and yielded surprisingly high sequence divergence values between the Northern Hemisphere T. romana and T. bambusae groups (10.8%). It was unexpected that the NPNS H. hirta and A. gayanus and the T. romana clade in particular did not cluster with the PNS group in the COI phylogeny as it did in the 28S phylogeny, and that the COI genetic divergences were unusually high for these groups.
It is known that the COI marker could yield unexpected results when delimiting species that have undergone a rapid, recent radiation, as the lack of recombination in the mitochondrial genome can lead to an overestimation of sequence divergences due to the accumulation and retention of mutations (Hupalo et al. 2023). Comparatively, due to recombination, nuclear markers will thus more readily indicate renewed gene flow between previously isolated populations (e.g., due to climatic cycles and habitat changes), which can result in conflicting phylogenies (Eyer et al. 2017; Després 2019). Additionally, it is likely that these wasps can reproduce facultatively via thelytokous parthenogenesis (i.e., a form of asexual reproduction where diploid daughters are produced from unfertilised eggs) (Moran and Goolsby 2009). Compared to the default reproductive mode of arrhenotoky in the Hymenoptera (i.e., unfertilised eggs develop into haploid males and fertilized eggs develop into diploid females), thelytoky can lead to increased levels of homozygosity over time (Mateo Leach et al. 2009). It is even possible that infection by different Wolbachia strains can cause reproductive isolation between intraspecific populations (e.g., in Nasonia wasps (Bordenstein et al. 2001)). Future phylogenetic studies could investigate a wider range of nuclear markers—both ribosomal and protein-coding—and also the relationship between Wolbachia and their Tetramesa hosts in order to determine the degree to which these endosymbionts might be affecting genetic diversity, and under what conditions the wasps change their mode of reproduction.
This study has presented the first phylogenetic analysis of southern African Tetramesa, and has revealed multiple taxa that are new to science. Combined with field host range data, we have identified at least five Tetramesa taxa that show promise as biological control agents (these were namely clades PNS.SPYR.1, PNS.SAFR.1, NPNS.HHIR.2, NPNS.HHIR.3, and NPNS.AGAYA.1 in Fig. 2 and Supplementary Fig. S6). These wasps appear to be host-specific to S. pyramidalis, S. africanus, H. hirta, and A. gayanus. Based on field host range data, Tetramesa sp. 4 on E. curvula showed evidence of oligophagy across congenerics, and is unlikely to be suitably host-specific for release in Australia where there is a very large diversity of Eragrostis species. Recommendations for future research include: (1) incorporating multiple other nuclear regions and ddRADseq techniques into phylogenetic analyses to refine divergence thresholds for delimiting true Tetramesa from other eurytomid genera, as well as to resolve the high incidence of polytomies observed, (2) using phylogenetic methods in conjunction with morphological and ecological data to revise the taxonomy of the group (with the possibility of creating subgenera), and (3) conducting further host-specificity tests, impact assessments, and potential hybridisation trials with the different Tetramesa taxa to determine their efficacy as biological control agents.
References
Al-Barrak M (2006) Host choice in Tetramesa petiolata (Walker) (Hymenoptera: Eurytomidae). J Entomol 3:55–60
Berry TR, van Noort S (2016) Review of Afrotropical Cryptopimpla Taschenberg (Hymenoptera, Ichneumonidae, Banchinae), with description of nine new species. ZooKeys 13(640):103–137
Bordenstein SR, O’Hara FP, Werren JH (2001) Wolbachia-induced incompatibility precedes other hybrid incompatibilities in Nasonia. Nature 409:707–710
Canavan K, Paterson I, Hill M, Dudley T (2019a) Testing the Enemy Release Hypothesis on tall-statured grasses in South Africa, using Arundo donax, Phragmites australis, and Phragmites mauritianus as models. Bull Entomol Res 109(3):287–299
Canavan S, Meyerson LA, Packer JG, Pyšek P, Maurel N, Lozano V, Richardson DM, Brundu G, Canavan K, Cicatelli A, Čuda J (2019b) Tall-statured grasses: a useful functional group for invasion science. Biol Invasions 21:37–58
Chejara KV, Kriticos D, Kristiansen P, Sindel BM, Whalley R, Nadolny C (2010) The current and future potential geographical distribution of Hyparrhenia hirta. Weed Res 50:174–184
Chen Y, Xiao H, Fu J, Huang DW (2004) A molecular phylogeny of eurytomid wasps inferred from DNA sequence data of 28S, 18S, 16S, and COI genes. Mol Phylogenet Evol 31:300–307
Claridge M (1961) A contribution to the biology and taxonomy of some Palaearctic species of Tetramesa Walker (= Isosoma Walk.; = Harmolita Motsch.) (Hymenoptera: Eurytomidae), with particular reference to the British fauna. Proc R Entomol Soc Lond 113:175–217
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu Rev Ecol Evol Syst 23:63–87
Dawah H (1987) Biological species problems in some Tetramesa (Hymenoptera, Eurytomidae). Biol J Linn 32:237–245
Després L (2019) One, two or more species? Mitonuclear discordance and species delimitation. Mol Ecol 28:3845–3847
Eyer P, Seltzer R, Reiner-Brodetzki T, Hefetz A (2017) An integrative approach to untangling species delimitation in the Cataglyphis bicolor desert ant complex in Israel. Mol Phylogenet Evol 115:128–139
Firn J (2009) African lovegrass in Australia: a valuable pasture species or embarrassing invader? Trop Grassl 43:86
Gates MW (2008) Species revision and generic systematics of world Rileyinae (Hymenoptera: Eurytomidae), vol 127. University of California Press, California
Ghajarieh H, Bruford M, Dawah HA, Fernandes C, Dodd CS (2006) Mitochondrial phylogenetics of UK eurytomids. J Entomol 3:167–179
Gillespie JJ, Munro JB, Heraty JM, Yoder MJ, Owen AK, Carmichael AE (2005) A secondary structural model of the 28S rRNA expansion segments D2 and D3 for chalcidoid wasps (Hymenoptera: Chalcidoidea). Mol Biol Evol 22:1593–1608
Goolsby J, Hathcock C, Vacek A, Kariyat R, Moran P, Martinez Jimenez M (2020) No evidence of non-target use of native or economic grasses and broadleaf plants by Arundo donax biological control agents. Biocontrol Sci Technol 30:795–805
Goolsby JA, Moran P (2009) Host range of Tetramesa romana Walker (Hymenoptera: Eurytomidae), a potential biological control of giant reed, Arundo donax L. in North America. Biol Control 49:160–168
Goolsby JA, Moran PJ, Racelis AE, Summy KR, Jimenez MM, Lacewell RD, Perez de Leon A, Kirk AA (2016) Impact of the biological control agent Tetramesa romana (Hymenoptera: Eurytomidae) on Arundo donax (Poaceae: Arundinoideae) along the Rio Grande River in Texas. Biocontrol Sci Technol 26:47–60
Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W, Gascuel O (2010) New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol 59(3):307–321
Harms NE, Williams DA, Purcell MF (2021) The role of overseas genetic surveys to potentially accelerate biological control development for a new Hydrilla verticillata introduction in the USA. BioControl 66:271–280
Henneicke K, Dawah H, Jervis M (1992) Taxonomy and biology of final-instar larvae of some Eurytomidae (Hymenoptera: Chalcidoidea) associated with grasses in the UK. J Nat Hist 26:1047–1087
Hopkins T, Roininen H, van Noort S, Broad GR, Kaunisto K, Sääksjärvi IE (2019) Extensive sampling and thorough taxonomic assessment of Afrotropical Rhyssinae (Hymenoptera, Ichneumonidae) reveals two new species and demonstrates the limitations of previous sampling efforts. ZooKeys 878:33
Hupalo K, Copilas-Ciocianu D, Leese F, Weiss M (2023) Morphology, nuclear SNPs and mate selection reveal that COI barcoding overestimates species diversity in a Mediterranean freshwater amphipod by an order of magnitude. Cladistics 39:129–143
Hussain F, Murad A, Durrani M (2004) Weed communities in the wheat fields of Mastuj, District Chitral, Pakistan. Pak J Weed Sci Res 10:101–108
Leigh J (1961) The relative palatability of various varieties of weeping lovegrass (Eragrostis curvula (schrad) nees). Grass Forage Sci 16:135–140
Lotfalizadeh H, Delvare G, Rasplus JY (2007) Phylogenetic analysis of Eurytominae (Chalcidoidea: Eurytomidae) based on morphological characters. Zool J Linn Soc 151:441–510
Lotfalizadeh H, Rasplus JY, Cristofaro M, Marini F (2020) Tetramesa amica and its parasitoid Eurytoma amicophaga (Hymenoptera, Eurytomidae): two new species associated with medusahead, Taeniatherum caput-medusae (Poaceae). ZooKeys 1005:133
Mateo Leach I, Hesseling A, Huibers W, Witsenboer H, Beukeboom L, Van De Zande L (2009) Transcriptome and proteome analysis of ovaries of arrhenotokous and thelytokous Venturia canescens. Insect Mol Biol 18:477–482
Moran PJ, Goolsby JA (2009) Biology of the galling wasp Tetramesa romana, a biological control agent of giant reed. Biol Control 49:169–179
Moran PJ, Vacek AT, Racelis AE, Pratt PD, Goolsby JA (2017) Impact of the Arundo wasp, Tetramesa romana (Hymenoptera: Eurytomidae), on biomass of the invasive weed, Arundo donax (Poaceae: Arundinoideae), and on revegetation of riparian habitat along the Rio Grande in Texas. Biocontrol Sci Technol 27:96–114
Munro JB, Heraty JM, Burks RA, Hawks D, Mottern J, Cruaud A, Rasplus JY, Jansta P (2011) A molecular phylogeny of the Chalcidoidea (Hymenoptera). PLoS ONE 6(11):e27023
Natural History Museum (2021) Universal Chalcidoidea Database: Species in genus Tetramesa. https://www.nhm.ac.uk/our-science/data/chalcidoids/database/. Accessed 11 Jan 2021
Olckers T, Coetzee J, Egli D, Martin G, Paterson I, Sutton G, Wood A (2021) Biological control of South African plants that are invasive elsewhere in the world: a review of earlier and current programmes. Afr Entomol 29:1005–1029
Phillips WJ (1936) A second revision of the chalcid flies of the genus Harmolita (Isosoma) of America north of Mexico, with descriptions of 20 new species. United States Department of Agriculture
Rossiter-Rachor NA, Setterfield SA, Douglas MM, Hutley LB, Cook GD, Schmidt S (2009) Invasive Andropogon gayanus (gamba grass) is an ecosystem transformer of nitrogen relations in Australian savanna. Ecol Appl 19:1546–1560
Skendzic EM, Columbus JT, Cerros-Tlatilpa R (2007) Phylogenetics of Andropogoneae (Poaceae: Panicoideae) based on nuclear ribosomal internal transcribed spacer and chloroplast trnL–F sequences. Aliso 23:530–544
Sutton GF, van Steenderen CJM, Yell LD, Canavan K, McConnachie A, Paterson ID (2023) First record of an African grass-feeding wasp (Tetramesa; Eurytomidae) on the invasive grass Eragrostis curvula (African lovegrass; Poaceae) in Australia. BioInvasions Rec 12(3):673–680
Sutton G, Bownes A, Visser V, Mapaura A, Canavan K (2021a) Progress and prospects for the biological control of invasive alien grasses (Poaceae) in South Africa. Afr Entomol 29:837–858
Sutton GF, Canavan K, Day MD, Paterson ID (2021b) Field-based ecological studies to assess prospective biological control agents for invasive alien plants: an example from giant rat’s tail grass. J Appl Ecol 58:1043–1054
Sutton G, Canavan K, Day M, Den Breeyen A, Goolsby J, Cristofaro M, McConnachie A, Paterson I (2019) Grasses as suitable targets for classical weed biological control. BioControl 64:605–622
van Noort, S., 2020. WaspWeb: Hymenoptera of the Afrotropical region. https://www.waspweb.org. Accessed 11–01–2021.
van Noort S, Buffington ML, Forshage M (2015) Afrotropical Cynipoidea (Hymenoptera). ZooKeys 493:1–176
Visser V, Wilson JR, Fish L, Brown C, Cook GD, Richardson DM (2016) Much more give than take: South Africa as a major donor but infrequent recipient of invasive non-native grasses. Glob Ecol Biogeogr 25:679–692
Winston R, Schwarzländer M, Hinz HL, Day MD, Cock MJ, Julien M (2021) Biological control of weeds: A world catalogue of agents and their target weeds. Based on FHTET-2014-04, USDA Forest Service, forest health technology enterprise team. Available online at https://www.ibiocontrol.org/catalog/
Witt A, McConnachie A (2004) The potential for classical biological control of invasive grass species with special reference to invasive Sporobolus spp. (Poaceae) in Australia, in: XI international symposium on biological control of weeds, p 198
Yu G, Smith DK, Zhu H, Guan Y, Lam TTY (2017) ggtree: an R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol Evol 8:28–36
Acknowledgements
The African lovegrass biocontrol project is supported by funding from the Australian Government Department of Agriculture, Fisheries and Forestry as part of its Rural Research and Development for Profit program and the AgriFutures Australia Programme. This project has been assisted by the New South Wales (NSW) Government through its Environmental Trust. The NSW Environmental Trust and the NSW Weed Biocontrol Taskforce are two of the cash contributors for African Love grass research under the national project `Underpinning agricultural productivity and biosecurity by weed biological control’, supported by the Australian Government programme Rural Research and Development for Profit (RRnD4P) (Round 4). Additional funding was provided by the Department of Forestry, Fisheries and the Environment: Natural Resource Management programme (DFFE: NRM) and the South African National Research Foundation (NRF) Research Chairs Initiative of the Department of Science and Technology. Any opinions, findings, conclusions and recommendations are those of the authors and the funders do not accept liability in this regard. We also wish to thank Lenin D. Chari for collecting specimens, the Albany museum for the use of their microscope and photographic software, Shelley Edwards for the use of the Zoology and Entomology Molecular Laboratory (ZEML) facilities, and Abigail Kirkaldy for assistance in photographing specimens.
Funding
Open access funding provided by Rhodes University.
Author information
Authors and Affiliations
Contributions
CJMvS: data curation, formal analysis, investigation, methodology, software, validation, visualisation, writing—original draft, writing—review and editing; GFS: conceptualization, data curation, investigation, project administration, supervision, writing—original draft, writing—review and editing; LDY: data curation, writing—review and editing; KC: data curation, writing—review and editing; AMcC: funding acquisition, resources, writing—review and editing; MDD: funding acquisition, resources, writing—review and editing; IDP: funding acquisition, project administration, resources, supervision, writing—original draft, writing—review and editing.
Corresponding author
Ethics declarations
Competing interests
We have no competing interests to declare.
Ethical approval
The Centre for Biological Control has a general ethical clearance number for all 2023 projects: 2022-3842-7223.
Informed consent
Informed consent was not applicable for this work.
Research involving human participants or animals
A statement on the welfare of humans and/or animals is not applicable.
Additional information
Handling Editor: Michelle Rafter
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
van Steenderen, C.J.M., Sutton, G.F., Yell, L.D. et al. Phylogenetic analyses reveal multiple new stem-boring Tetramesa taxa (Hymenoptera: Eurytomidae): implications for the biological control of invasive African grasses. BioControl 68, 697–708 (2023). https://doi.org/10.1007/s10526-023-10231-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-023-10231-4