
Abstract
Although once popular prior to the last century, the aquaculture of crucian carp Carassius carassius (L. 1758) in Sweden gradually fell from favour. This is the first genetic comparison of crucian carp from historic man-made ponds in the Scandinavian Peninsula. The aim was to identify old populations without admixture and to compare the relationship of pond populations from different provinces in Sweden. In total, nine microsatellite loci from 234 individuals from 20 locations in varied parts of Sweden were analysed. The genetic distances of crucian carp populations indicated that the populations in the southernmost province of Sweden, Scania, shared a common history. A pond population in the province Småland also showed a common inheritance with this group. In the province Uppland, further north in Sweden, the population genetic distances suggested a much more complex history of crucian carp distributions in the ponds. The data showed that there are some ponds with potentially old populations without admixture, but also that several ponds might have been stocked with fish from many sources.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The crucian carp Carassius carassius (L. 1758) is a member of the Cyprinidae family. It has a widespread distribution in northern parts of Eurasia from France and England in the west towards Kazakhstan and Siberia in the east. Due to extensive releases, feral populations exist in many places (Kottelat and Freyhof 2007). While its popularity as food source has decreased in Sweden and most other western European countries, it is still an appreciated food item in Eastern European cuisine (Szczerbowski and Szczerbowski 2002). Crucian carp is regarded as a native species in Sweden, widespread in lakes and in brackish waters along the Baltic Sea coast (Kullander 2012). Furthermore, crucian carp was one of the most common fish used in aquaculture in Sweden. There are numerous reports about crucian carp aquaculture in Sweden until the early nineteenth century (Bonow and Svanberg 2011). The pond keeping in Sweden dates back to the late Middle Ages. It was extensive during centuries, present at vicarages, manors and castles (Bonow and Svanberg 2012). Documents show that Carl Linnaeus was enticed by this type of aquaculture after visiting castles in the southernmost province of Sweden, Scania, and he therefore encouraged pond keeping in other regions of Sweden (Svanberg et al. 2012). The prime goal for Linnaeus was to obtain an additional source of food for the Swedish population that was on the rim of starvation. Even though some ponds still remain, this kind of aquaculture has in large been abandoned in Sweden.
In Sweden, crucian carp is found in high densities in small shallow water bodies, such as in historic eighteenth century ponds. Crucian carp have the extraordinary ability to survive under the ice in small ponds for up to several months in the absence of oxygen, enabling them to survive the harsh Swedish winters (Holopainen et al. 1997). Due to this extraordinary ability to survive in such adverse conditions, one may surmise that crucian carp in old ponds may well be the same as the original stock.
Crucian carp can display different body morphologies, apart from displaying a wide colour range from golden/green to reddish/brown; it will also become more deep-bodied with relatively smaller head-size in lakes (Poléo et al. 1995). The difference in morphology in the two habitats has previously led to the classification of crucian carp into different variants distinguishing between lake and pond fish (Holopainen et al. 1997). Experimental research has pointed out biotic factors as the determinant factor of body morphology (e.g. Brönmark and Miner 1992; Domenici et al. 2008). There is, however, no genetic comparison between fish from ponds and lakes that address this question.
In this study, microsatellites developed for gibel carp C. gibelio and goldfish C. auratus were used to investigate the relationship of population structure between fish from historic ponds, lakes and coastal waters in Sweden. These species are closely related to each other and appear as a sister group to crucian carp in phylogenetic analyses (Rylková et al. 2013). The aim was primarily to identify ponds with old populations and secondly to compare pond populations within and between regions of Sweden. This information could give insights on where the fish in the ponds originate from and how they were distributed between estates. One additional aim was to compare pond fish with wild/feral fish. The impact of aquaculture of crucian carp on populations in lakes has never been investigated. The populations in lakes are potentially threatened by invasive species such as gibel carp that has recently been found in Swedish waters (Wouters et al. 2012). Old populations from ponds could therefore serve as reservoirs for the original Swedish crucian carp to be used in future aquaculture, or restoration projects, instead of using recently imported stocks.
Materials and methods
Fish collection and study sites
Specimens from ponds were collected from the selected sampling sites by fish-traps using commercial bait for cyprinids. Typically, the traps were left overnight and harvested the following morning. Fish were anaesthetised with 2-phenoxy-ethanol before sampling. Fin clips were persevered in 95 % ethanol until DNA extraction. A total of 14 old ponds from six provinces were selected along with five wild populations from three provinces. We defined old ponds as being at least 100 years old. The sampling locations for pond fish were selected by examining the oldest historical maps where the pond first appear and confirming present-day existence with Google earth (Bonow and Svanberg 2011), thus determining the earliest historical record of the pond. The historical record of the pond at Glimmingehus (SK5P) could not be defined by this method. This study was approved by the ethical committee of Animal Research (Huddinge City Court; dno. S97-10).
Genetic analysis
DNA extraction was performed as described previously (Wouters et al. 2012). Seven primer pairs for microsatellites loci from gibel carp Carassius gibelio (Bloch 1782) YJ10, YJ20, YJ22 (Guo and Gui 2008), J58, J60, J68 and J69 (Yue and Orban 2002) were used in this study. In addition, the primer pairs MFW7 (Crooijmans et al. 1997) and GF29 (Zheng et al. 1995) previously used on crucian carp (Hänfling et al. 2005) were also used. Primers were labelled with fluorescence marker 6-FAM or HEX, respectively. The PCR was programmed for one cycle of 2 min denaturing at 94 °C, 40 cycles of 15 s at 94 °C, 20 s annealing at variable temperatures and 30 s at 72 °C, with a 10 min final extension at 72 °C. The annealing temperature was 52 °C for all primer pairs except for loci: MFW7 (55 °C), J68 (50 °C) and GF29 (45 °C).
Allele size determination of PCR products was performed by Genescan analysis at Macrogen Inc. The length of the DNA fragments was analysed in Peak Scanner Software v1.0 (Applied Biosystems; www.appliedbiosystems.com). Fragment size was reduced by one base to compensate for Taq-polymerase-mediated addition of adenosine residues.
Data analysis
The microsatellite data were analysed for the presence of null alleles using MICRO-CHECKER (Van Oosterhout et al. 2004). General F-statistics were calculated using the MSA software package (Dieringer and Schlötterer 2003), and allelic richness was determined with HP-RARE (Kalinowski 2005). Conversions of data formats used in different software were performed using PGD SPIDER (Lischer and Excoffier 2012). The presence of outlier among the nine loci was tested with LOSITAN (Antao et al. 2008). This software uses an Fst-outlier method to detect candidate loci for directional, neutral or balancing selection.
To examine the relationship among population sites, Nei’s genetic chord distances (Da) (Nei et al. 1983) and Cavalli-Sforza and Edwards’ (1967) chord distances (Dc) were calculated from the allelic frequency data with the MSA program and used to generate a bootstrapped dataset with 1,000 replicates. The genetic trees were constructed using PHYLIP 3.63 (Felsenstein 2005). Neighbour-joining trees were inferred using NEIGHBOR, with 1,000 bootstrapped distances as input. The consensus tree of the 1,000 trees was produced in CONSENSE. The population from the Czech Republic was used as an outgroup as it has been shown by mitochondrial DNA analysis to be distinct from the Swedish populations (Wouters et al. 2012).
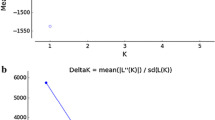
In order to further evaluate distinctive populations, the program STRUCTURE was used. STRUCTURE uses a Markov chain Monte Carlo model independent of assumptions about mutation process within a Bayesian framework (Pritchard et al. 2000; Falush et al. 2003). Hardy–Weinberg equilibrium and linkage equilibrium within each population is assumed in STRUCTURE. Population-assignment was tested by varying the number of groups (K) in independent runs of the algorithm, ranging from K = 1 to K = 21. The program then assigns individuals to one or more groups, with their relative frequency of predicted membership in colour-coded clusters. Burn-ins of 100,000 and 100,000 replicates were used, and the log-likelihood and bar-plot were recorded for each K, and 10–100 repeats at each K. The most likely value of K according to STRUCTURE was determined by plotting the number of K against the likelihood scores and reading the K value where the curve levels out. This was verified by determining the highest delta K value with STRUCTURE HARVESTER (Earl and vonHoldt 2012). The population structures were then simulated using the determined value of K and the default settings with an admixture model and linked allele frequencies. In the admixture model, both a fixed value and varied value of alpha were tested.
Results
From 246 individuals, DNA was successfully extracted and amplified using nine microsatellite primer pairs. General information along with allelic richness and likelihood of heterozygosity excess are summarised in Table 1. The preponderance of allelic richness was found in the wild population U7W, but the pond populations SK3P and U1P scored a higher value than some of the wild populations. Private alleles were found in the populations U6W (YJ10; 153), U7W (J58; 143), ÖG1P (212; GF29) and SK3P (J68; 265). In the data set, the presence of null alleles was recorded with MICRO-CHECKER. The analysis indicated null alleles at locus J60 in some of the populations. Removing J60 from the analysis did not significantly alter the results. Moreover, it had the greatest number of alleles, which could be of interest when looking for differences between populations. The overall F-statistics of each locus showed that locus J60 had a much higher Fis-value than the other loci (Table 2). On the other hand, the outlier analysis performed with LOSITAN did not suggest that any of the loci were candidates for directional selection (data not shown).
The neighbour-joining tree shown in Fig. 2 depicts the relationship between sampled population sites for crucian carp (as described in Table 1), with the Czech Republic population as outgroup. The tree that was calculated from chord distances (Cavalli-Sforza and Edwards 1967) was similar in topology and resolution and is thus not shown. There were two clear cases where population genetic branch support and geographic proximity coincided. One was the samples from Scania and one location in Småland (S3P). The neighbour-joining trees showed these populations as a separate cluster with a bootstrap support of 74 % (Fig. 2). In the second case, the populations from Ö1P and the creek S2W were grouped with relatively high bootstrap support (69 %).
Two sampled populations from two different provinces (ÖG2P and U7W) were clustered with a high bootstrap support of 76 %. These localities were not close to each other (Fig. 1). Other branches in the tree showed low bootstrap support, clustered in smaller groups without reflecting geographic proximity (Figs. 1, 2). The populations U5P and G1P were clustered together with a low bootstrap value.

The map shows the southern part of Sweden where the 20 sampling sites are located as indicated by dots

A neighbour-joining tree showing relationships among pond populations and wild fish from different areas in Sweden for C. carassius based on Nei’s genetic distance. A wild population from the Czech Republic was used as an “outgroup”. Values at nodes denote the support of re-sampling from 1000 bootstrap in percentage for value over 50 %. The scale bar indicates the sequence divergence in the tree
Using Bayesian analyses in the software STRUCTURE, the number of K was determined to 15, indicating that there were 15 distinct populations among the samples from Sweden. In Fig. 3, the results for K = 15 are illustrated along with the amount each individual contributed to the clusters. The information indicated how homogeneous each individual was by one or several colours. Two populations with the same colour therefore indicate a close relationship. A uniform colour within one sampling site indicates that there has been no admixture event in that population.

The figure shows the population structure for Swedish crucian carp from ponds, lakes and Baltic coast, and the number of populations set to 15. Individuals are represented by vertical lines. Each line is either uni-coloured or segmented into different colours, where each colour represents one cluster and the y-axis shows the probability scale. Black horizontal lines define the different sampling sites as indicated above each line. (Color figure online)
The pond population U1P had a great deal of admixture (Fig. 3). This population was comparable to wild populations from the same province U6W and U7W. The other four pond populations from this province were rather homogenous in genetic structure with smaller contributions from other clusters. The same was true for the two pond populations from Östergötland, Ö1P and Ö2P, although a few individuals indicated relationships with more than one cluster. On examination, the one pond population from Öland showed a uniform structure with individuals belonging to one cluster. This cluster was also represented in two individuals from the wild population S2W. The other wild population from this province S1W had individuals that were related to clusters found in Uppland, but also from a cluster represented in S2W.
The pond populations with the most uniform individuals with a low probability of admixture were U5P from Bogesund in Uppland, G1P from Vall Bryungs on the island Gotland and Ö1P from Ottenby on the island Öland (Fig. 3). The pond populations in U5P and G1P shared the same population cluster when alpha was set to vary between populations in STRUCTURE. Apart from that, there were no major changes in the other populations (data not shown).
Discussion
This is the first analysis of the genetic population structure of crucian carp from Sweden, the aim being to discover whether historic ponds still hold populations of crucian carp, and secondly to study their genetic constitution, not only as a measure of relatedness between pond populations but also in comparison to wild populations. The sampling strategy was therefore not optimised for a rigorous and detailed analysis of the population genetics in each of these ponds and lakes.
The analysis of the population structure revealed that three pond populations, U5P, G1P and Ö1P, comprised members that were closest to be completely without detectable admixture events. This indicates that they have been isolated from other populations for a long time period and presumably genetic drift has led to unique allele frequencies not shared by the other populations in this study. In those ponds, having little or no admixture, there is the potential to become sources of future stocking and reservoirs of ancient/native crucian carp populations. In contrast, the populations in U1P, SK1P and SK3P were conspicuously mixed in terms of population structure and are therefore not likely to comprise only one, original, population. They may have been re-stocked several times with fish caught from different populations, or with fish from other ponds. Indeed, historic notes of medieval aquaculture suggest that wild fish were caught and put into ponds for later use (Bonow and Svanberg 2012).
The populations from two ponds in the province of Östergötland showed very different structures and were not closely related to the population genetic tree. Some individuals in each population belonged to several clusters according to STRUCTURE, indicating that they were admixed. The populations in ÖG2P seemed to be closely related to the coastal population from the province Uppland, U7W. These two populations also shared some population clusters. It is therefore possible that the ÖG2P pond has been stocked with fish from the coast of the Baltic Sea.
The two sampling sites Ö1P and the small creek near Kalmar S2W are separated by Kalmar Strait and could thus have a source population in common. Indeed, the population genetic tree showed a well-supported cluster of these two populations. The other sampled population from Kalmar Strait, S1W, did not belong to this cluster. However, the STRUCTURE analysis indicated that they shared population clusters with each other.
All populations from Scania and from the pond S3P in Småland uniquely belonged to the same neighbour-joining cluster. In all other populations, there was little or no correlation with the geographic location. This suggests that the fish used for stocking ponds came from the same source in Scania and were more diverse in the other provinces. Judging from available sources, the pond farming of crucian carp in Scania may have been established well before it became common in Uppland (Hofmeister 2004; Svanberg et al. 2012). Scania was until 1658 a province of Denmark, and the fish in these ponds might have been of Danish origin. However, there are no genetic data from Danish crucian carp to compare with.
In Scania, the populations from the lake and the ponds were part of the same neighbour-joining cluster, well supported by bootstrap analysis. This suggests that ponds were stocked with wild fish caught in nearby waters or that fish has been released from ponds into lakes. The wild populations on the other hand could be feral fish spread from the first man-made ponds in the late middle ages. This could be indicated by the small differences in allelic richness between wild and pond fish. If the wild fish were naturally colonised, one would expect a higher degree of allelic richness in these populations. The expected heterozygosity should also be higher in the wild population, but this was not the case if compared to some of the pond populations. Whether the fish has been moved to or from the lake requires further investigations with a larger number of populations included.
Even though it was a weak association between populations U5P and G1P in both genetic distance and population structure, it suggests that there has been some contact between these populations. A stronger association was on the other hand seen among the populations in Scania, especially the ponds SK4P and SK5P that are located quite far from each other (close to 70 km) and yet share a common population cluster as suggested by the STRUCTURE analysis. One possibility is that land owners shared their crucian carp populations with each other and therefore are populations spread over larger distances than would be the case if they have been spread naturally.
In conclusion, this is the first study of population genetics of crucian carp in Sweden and it shows that there are candidates for old populations present in historic ponds. These populations can be of great value should the aquaculture of crucian carp be re-introduced in Sweden.
References
Antao T, Lopes A, Lopes RJ, Beja-Pereira A, Luikart G (2008) LOSITAN: a workbench to detect molecular adaptation based on a Fst-outlier method. BMC Bioinformatics 9:323
Bonow M, Svanberg I (2011) » Säj får jag dig bjuda ur sumpen en sprittande ruda « : en bortglömde läckerhet från gångna tiders prästgårdskök. In Bonow M, Rytkönen P (eds) Gastronomins (politiska) geografi. Svenska Sällskapet för Antropologi och Geografi, Stockholm, pp 147–169
Bonow M, Svanberg I (2012) Rudodling i Uppland: ett bidrag till akvakulturens historia. Årsbok, Uppland, pp 123–152
Brönmark C, Miner JG (1992) Predator-induced phenotypical change in body morphology in crucian carp. Science 258:1348–1350
Cavalli-Sforza LL, Edwards AWF (1967) Phylogenetic analysis: models and estimation procedures. Am J Human Genet 19:233–257
Crooijmans A, Vander J, Groene M, Groenen M (1997) Microsatellite markers in common carp (Cyprinus carpio L.). Anim Genet 28:129–134
Dieringer D, Schlötterer C (2003) Microsatellite analyser (MSA): a platform independent analysis tool for large microsatellite data sets. Mol Ecol Notes 3:167–169
Domenici P, Turesson H, Brodersen J, Brönmark C (2008) Predator-induced morphology enhances escape locomotion in crucian carp. Proc R Soc B 275:195–201
Earl DA, vonHoldt BM (2012) STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour 4:359–361
Falush D, Stephens M, Pritchard JK (2003) Inference of population structure: extensions to linked loci and correlated allele frequencies. Genetics 164:1567–1587
Felsenstein J (2005) PHYLIP (Phylogeny Inference Package) version 3.6. Distributed by the author. Department of Genome Sciences, University of Washington, Seattle [http://www.evolution.genetics.washington.edu/phylip.html]
Guo W, Gui J-F (2008) Microsatellite marker isolation and cultured strain identification in Carassius auratus gibelio. Aquacult Int 16:497–510
Hänfling B, Bolton P, Harley M, Carvalho GR (2005) A molecular approach to detect hybridisation between crucian carp (Carassius carassius) and non-indigenous carp species (Carassius spp. and Cyprinus carpio). Freshw Biol 50:403–417
Hofmeister E (2004) Fiskeproduktion ved de ferske vande—fra karper til regnbuger. In: Hofmeister E (ed) De ferske vandes kulturhistorie i Danmark. Aqua, Silkeborg, pp 75–86
Holopainen IJ, Tonn WM, Paszkowski CA (1997) Tales of two fish: the dichotomous biology of crucian carp (Carassius carassius (L.)) in northern Europe. Ann Zool Fenn 34:1–22
Kalinowski ST (2005) HP-rare: a computer program for performing rarefaction on measures of allelic diversity. Mol Ecol Notes 5:187–189
Kottelat M, Freyhof J (2007) Handbook of European freshwater fishes. xiv+ . Berlin: Kottelat, Cornol & Freyhof
Kullander SO (2012) Ryggsträngsdjur: Strålfeniga fiskar: Chordata: Actinopterygii, Nationalnyckeln till Sveriges fauna och flora. Artdatabanken, Sveriges Lantbruksuniversitet, Uppsala
Lischer HEL, Excoffier L (2012) PGDSpider: an automated data conversion tool for connecting population genetics and genomics programs. Bioinformatics 28:298–299
Nei M, Tajima F, Tateno Y (1983) Accuracy of estimated phylogenetic trees from molecular data. J Mol Evol 19:153–170
Poléo ABS, Øxnevad SA, Østbye K, Heibo E, Anderson RA, Vøllestad LA (1995) Body morphology of crucian carp Carassius carassius in lakes with or without piscivorous fish. Ecography 18:225–229
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155:945–959
Rylková K, Kalous L, Bohlen J, Lamatsch DK, Petrtıl M (2013) Phylogeny and biogeographic history of the cyprinid fish genus Carassius (Teleostei: Cyprinidae) with focus on natural and anthropogenic arrivals in Europe. Aquaculture 380–383:13–20
Svanberg I, Bonow M, Olsén H (2012) Fish ponds in Scania, and Linnaeus’s attempt to promote aquaculture in Sweden. Svenska Linnésällskapets Årsskrift 2012:83–98
Szczerbowski JA, Szczerbowski AJ (2002) Carassius carassius (Linnaeus, 1758). In: Bănărescue PM, Paepke H-J (eds) The freshwater fishes of Europe. Vol 5/III. Cyprinidae 2/III and Gasterosteidae, Aula, Wiebelsheim, pp. 43–78
Van Oosterhout C, Hutchinson WF, Wills DPM, Shipley P (2004) MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Mol Ecol Notes 4:535–538
Wouters J, Janson S, Lusková V, Olsén KH (2012) Molecular identification of hybrids of the invasive gibel carp Carassius auratus gibelio and crucian carp Carassius carassius in Swedish waters. J Fish Biol 80:2595–2604
Yue GH, Orban L (2002) Microsatellites from genes show polymorphism in two related Oreochromis species. Mol Ecol Notes 2:99–100
Zheng W, Stacey NE, Coffin J, Strobeck C (1995) Isolation and characterization of microsatellite loci in the goldfish Carassius auratus. Mol Ecol 4:791–792
Acknowledgments
For their assistance in collecting fish we acknowledge: V. Lusková (Institute of Vertebrate Biology of the Academy of Sciences of Czech Republic); P. Koch-Smith, O. Engstedt, P. Larsson, S. Andersson and S. Tobiasson (Department of Natural Sciences, Linnaeus University, Kalmar, Sweden); C. Brönmark and K. Hulthén (Department of Biology, Lund University); O. Sandsröm; B. Tengelin Structor Miljöteknik AB; M. Andersson; N. Hellenberg; A. Olsén-Wannefjord. The landowners are greatly acknowledged for giving their permission to catch fish from the ponds. The group of I. Porsch-Hällström and S. Hallgren (School of Natural Science, Technology and Environmental Studies, Södertörn University) is thanked for assistance in the lab. This study was funded by a grant from The Foundation for Baltic and Eastern European Studies (Östersjöstiftelsen). We are grateful to W. Horn for a critical review of the text.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Janson, S., Wouters, J., Bonow, M. et al. Population genetic structure of crucian carp (Carassius carassius) in man-made ponds and wild populations in Sweden. Aquacult Int 23, 359–368 (2015). https://doi.org/10.1007/s10499-014-9820-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10499-014-9820-4