
Abstract
Microarthropod communities in the soil and on the bark of trees were investigated along an elevation gradient (1,850, 2,000, 2,150, 2,300 m) in a tropical montane rain forest in southern Ecuador. We hypothesised that the density of microarthropods declines with depth in soil and increases with increasing altitude mainly due to the availability of resources, i.e. organic matter. In addition, we expected bark and soil communities to differ strongly, since the bark of trees is more exposed to harsher factors. In contrast to our hypothesis, the density of major microarthropod groups (Collembola, Oribatida, Gamasina, Uropodina) was generally low and decreased with altitude. However, as we predicted the density of each of the groups decreased with soil depth. Density of microarthropods on tree bark was lower than in soil. Overall, 43 species of oribatid mites were found, with the most abundant higher taxa being Poronota, pycnonotic Apheredermata, Mixonomata and Eupheredermata. The oribatid mite community on bark did not differ significantly from that in soil. The number of oribatid mite species declined with altitude (24, 23, 17 and 13 species at 1,850, 2,000, 2,150 and 2,300 m, respectively). Rarefaction curves indicate that overall about 50 oribatid mite species are to be expected along the studied altitudinal gradient. Results of this study indicate (1) that microarthropods may be limited by the quality of resources at high altitudes and by the amount of resources at deeper soil layers, and (2) that the bark of trees and the soil are habitats of similar quality for oribatid mites.
Similar content being viewed by others


Explore related subjects
Find the latest articles, discoveries, and news in related topics.Avoid common mistakes on your manuscript.
Introduction
Soil microarthropods, such as Oribatida and Collembola, are major decomposer taxa in terrestrial ecosystems, especially in those where macrodecomposers, such as earthworms and diplopods, are rare or lacking (Maraun and Scheu 2000). In tropical montane rain forests macrodecomposers are usually rare (Maraun et al. 2008) and soil microarthropods comprise the most important decomposers (Beck 1971; Plowman 1981; Maraun et al. 2007). Soil microarthropods affect decomposition processes directly and indirectly, e.g. by comminution of litter, feeding on microorganisms and dispersal of microbial propagules (Behan and Hill 1978; Maraun et al. 1998; Renker et al. 2005). However, there is increasing evidence that typical decomposer taxa such as oribatid mites and collembolans do not consume exclusively dead plant material but also (or mainly) feed on fungi and even on dead or living animals (Schneider et al. 2004; K. Heidemann, unpublished data).
Most studies of soil microarthropods in tropical rain forests have been carried out in lowland forests (e.g. Wunderle 1992; Palacios-Vargas et al. 2007). By contrast, few have investigated density, diversity and community structure of soil microarthropods in tropical montane regions (e.g. Holt 1985). In temperate forests the density of soil microarthropods is strongly correlated with the thickness of organic layers and with the pH (Maraun and Scheu 2000). Tropical montane rain forests are rather old and acidic ecosystems. Parallel to the decrease of soil pH the amount of organic material (especially dead and living roots; Röderstein et al. 2005) increases with altitude in montane rain forests in southern Ecuador (Wilcke et al. 2008). To relate these changes to changes in the decomposer community we investigated soil microarthropods along an altitude gradient between 1,800 and 2,300 m.
The density of most soil microarthropods in temperate ecosystems decreases with soil depth (Mitchell 1978; Plowman 1981), which likely is due to a lower amount of organic food resources and reduced habitat complexity in deeper soil layers (Usher 1970; Pande and Berthet 1975). To investigate if this also applies to soil microarthropods of tropical montane rain forests we studied changes in soil microarthropod density with soil depth.
Tropical montane rain forests are hotspots of arthropod diversity (Myers et al. 2000), however, this has mainly been proven for above-ground taxa such as Lepidoptera (Janzen et al. 1976; Brehm and Fiedler 2003, 2004; Brehm et al. 2005) and ants (Andersen 1992; Brühl et al. 1999; Longino et al. 2002). In contrast, knowledge of the diversity of the below-ground fauna is very limited. Soil animal diversity typically declines with altitude (Olson 1994; Rahbek 2005) but diversity may also peak at intermediate altitudes (Bachmann et al. 2004; Krashevska et al. 2007).
One of the microhabitats of microarthropods that has a distinct species composition is the bark of trees, e.g. bark-living oribatid mite species rarely also live in soil (Behan-Pelletier and Winchester 1998; Proctor et al. 2002; Prinzing and Woas 2003; Erdmann et al. 2006; Lindo and Winchester 2006) indicating strong niche differentiation (Erdmann et al. 2007). This niche differentiation contributes substantially to the diversity of microarthropods in temperate systems; the same may also apply for tropical microhabiats.
In the present study we investigated (1) if densities of four soil microarthropod groups—Collembola, Oribatida, Gamasina, Uropodina—change with altitude (1,850, 2,000, 2,150 and 2,300 m) and with horizon (litter, 0–5 and 5–10 cm soil depth) and (2) if the density of soil microarthropods and the community structure of oribatid mites differ between bark and soil. We expected microarthropod densities to be rather high since large macrodecomposer taxa are lacking in the studied forest. We also expected the density of soil microarthropods to increase with altitude since lower temperatures at higher altitudes likely reduce the decomposition rates resulting in larger amounts of organic material, which in turn might result in increased numbers of organisms.
Materials and methods
Study side and sampling
This study was conducted at the Reserva Biológica San Francisco (RBSF) forest (3°58′S, 79°5′W) in southern Ecuador on the eastern slopes of the Andes. The mean annual temperature at 1,950 m is ~16°C (Wilcke et al. 2002). The forest has a stem density of 1,000 stems ha−1 with a medium diameter at breast height (DBH) of ~0.1 m and of 6,000 stems ha−1 with a DBH of ~0.05 m. The vegetation of the gentle lower slopes is dominated by Euphorbiaceae, Solanaceae, Cecropiaceae and Lauraceae; the higher slopes by Melastomataceae, Lauraceae, Euphorbiaceae and Rubiaceae. The most abundant tree species at 1,800–2,200 m is Graffenrieda emarginata (Melastomataceae). The ground flora is dominated by large ferns, e.g. Dryopteridaceae, and some large herbs (mainly Lobeliaceae). The thickness of the organic soil layer ranges from 5 to 16 cm (Wilcke et al. 2002), and increases with altitude resulting in Histosols (mainly Folic Histosols) above 2,100 m, whereas Cambisols (mainly Dystric and Humic Cambisols) dominate below 2,100 m (Wilcke et al. 2002).
In January 2004, samples were taken at 1,850, 2,000, 2,150 and 2,300 m. Three replicate litter and soil samples (~20 m apart from each other) were taken with a soil corer (Ø 5 cm) from each altitude to a depth of 10 cm and divided into three horizons [L, F/H (~5 cm) and Ah (~5 cm)]. Additionally, in the immediate vicinity of each soil core, a living tree was randomly selected on which bark material (with associated epiphyte cover) was sampled at about 1.5 m height to a depth of 2–4 mm (Ø 5 cm). This is a design with pseudoreplicates since we sampled microarthropods from a single slope. However, since we only aimed at investigating the microarthropods from this specific altitudinal gradient we think it is appropriate to discuss our findings and their relevance for this specific study site. Samples were transferred to the laboratory and microarthropods were immediately extracted by heat using a modified high gradient extractor (Kempson et al. 1963). Microarthropods were quantified and determined to group level (Collembola, Oribatida, Gamasina, Uropodina), and adult Oribatida were identified to family and to species level if possible. Higher-level classification followed Grandjean (1953, 1965, 1969).
Statistical analysis
First, the effects of horizon and altitude on the densities of the soil microarthropod taxa in soil were analysed using MANOVA with the within-subject effects Horizon (litter layer, 0–5, 5–10 cm) and Taxon (Collembola, Oribatida, Gamasina, Uropodina) since (a) densities of animal taxa in the three horizons are interdependent and (b) densities of the animal taxa may also depend on each other. The effect of altitude on microarthropod density was investigated using altitude (1,850, 2,000, 2,150, 2,300 m) as the fixed factor (between-subject effect). Prior to the analyses densities of microarthropods were log-transformed to improve homogeneity of variances. For graphical representation data were back-transformed and multiplied by 509 (to calculate densities per square meter). Standard errors were also back-transformed, which resulted in uneven errors about the mean. The analyses were carried out in SAS 9.13 (SAS Institute Inc., Cary, USA).
Subsequently, using the same data set the community structure of Oribatida at the four altitudes was compared using non-metric multidimensional scaling (NMDS) followed by discriminant function analysis (DFA). To control for different densities at the four altitudes animal densities were calibrated to the mean density of Oribatida at the study sites before the analysis. NMDS served to reduce the number of variables (species) of the dataset. NMDS searches a non-parametric relationship between the dissimilarities in the data matrix and the Euclidian distance between them (Legendre and Legendre 1998). Then, the objects are re-arranged in lower dimensional space to arrive at a condition that best approximates the observed distance. The quality of the NMDS is indicated by stress values. Stress values indicate how strongly the objects in the compressed matrix differ from the originally calculated distances. By comparing the actual stress values with a theoretical exponential function of stress a meaningful number of dimensions was evaluated. In our case NMDS reduced the number of meaningful dimensions to three. The coordinates of the samples in the 3-dimensional space were subsequently used in DFA with Altitude as grouping variable. Squared Mahalanobis distances between group centroids and the reliability of the sample classifications were determined. Two significant canonical roots were derived and the DFA was graphically presented in 2-dimensional space (Fig. S1).
Principal component analysis (PCA) was used to analyse (and to present graphically) the response of Oribatida at family, genus or species level to different altitudes; Altitude (1,850, 2,000, 2,150, 2,300 m) was coded as a supplementary variable and included in the analysis using the passive analysis procedure in CANOCO (Jongman et al. 1995; Ter Braak and Šmilauer 1998), i.e. the variable did not contribute to the ordination. Only species that were found in at least two separate samples were included in the analyses. Prior to the analyses data were log-transformed. The data analysis using NMDS and DFA was done with the STATISTICA 7.1 software package (Statsoft, Tulsa, USA).
Subsequently, the densities of the four soil microarthropod taxa in soil and bark at the four altitudes were compared. For this, the densities of the animals in the horizons were summed to obtain area-based abundance data that can be compared to those of bark. Hence, a two factorial MANOVA with the fixed factors Habitat (soil/bark) and Altitude (1,850, 2,000, 2,150 and 2,300 m) and the dependent variables Collembola, Oribatida, Gamasina and Uropodina was performed. In case of significant results, protected ANOVAs for the four microarthropod taxa were carried out. Prior to the analyses, densities of microarthropods were log-transformed to improve homogeneity of variances. All analyses were carried out using the animal densities per soil sample, i.e. 20 cm². For graphical representation data were back-transformed and converted to individuals per square meter. Standard errors were also back-transformed, which resulted in uneven errors about the mean. Statistical analyses were performed using SAS 9.13 (SAS Institute, Cary, NC, USA).
Additionally, the community structure of Oribatida from the bark of trees was compared with that of the soil. To control for different densities on bark and in soil the data were calibrated to the mean density of Oribatida at both habitats before the analysis. Since a large number of Oribatida taxa were analysed we performed a NMDS and subsequent DFA (for details see above) using STATISTICA 7.1 (Statsoft, Tulsa, USA).
Finally, to study the effect of the number of samples on species richness we used the richness estimates Michaelis–Menten, ICE (incidence-based coverage estimator of species richness) and Jackknife implemented in EstimateS 8.0 (Colwell 1994–2004; URL http://viceroy.eeb.uconn.edu/estimates). Sample base rarefaction curves were produced for each data set. For all calculations involving multiple random orderings of the samples, 50 randomizations were used. The ICE calculations were carried out using the default option “cut-point” of 10 (Lee and Chao 1994). Michaelis–Menten estimates were calculated using the smoothed species accumulation curve (MMMeans; Longino et al. 2002).
Results
Soil and litter
Mean densities of the four microarthropod taxa (Collembola, Oribatida, Gamasina, Uropodina) declined with Altitude (Repeated MANOVA; between subject effect: F 3,8 = 9.08; P = 0.006) and with Horizon (within subject effect; F 2,16 = 77.58; P = 0.0001); however, the decline with altitude differed among the three horizons (within subject interaction: horizon × altitude: F 6,16 = 13.24; P = 0.0001). At 1,850 m the density of the soil microarthropod taxa was much higher in litter compared with the two soil horizons, whereas at the higher altitudes (2,000, 2,150, 2,300 m) these differences were less pronounced (Fig. 1a). Density of Oribatida at the study site was highest followed by that of Collembola, Gamasina and Uropodina (F 3,24 = 36.92; P = 0.0001). Densities of all microarthropod taxa declined with horizon but the magnitude differed among the four taxa (within subject interaction: F 6,48 = 6.8; P = 0.0001). Oribatida and Collembola densities strongly declined from the litter layer to the soil layers whereas densities of Gamasina and Uropodina were only slightly higher in litter compared with the soil layers (Fig. 1b).

Microarthropod densities in the three soil layers at the four different altitudes (a), and density of the four microarthropod taxa in the three soil horizons in a tropical montane rain forest in Southern Ecuador (b). For Fig. 1a the densities of all microarthropod taxa were summed up; for Fig. 1b the mean of the four altitudes in given. Bars and uneven standard errors represent back-transformed means of log-transformed data (for “Statistical analysis” see text)
Soil as compared to the bark of trees
Microarthropod densities were on average higher in soil than on the bark of trees (Roy’s greatest root F 4,13 = 4.52; P = 0.017) and declined with Altitude (Roy’s greatest root F 4,15 = 4.22; P = 0.018), however, the differences in densities between bark and soil tended to depend on Altitude (Roy’s greatest root: bark × altitude interaction; F 4,15 = 2.7; P = 0.07). Densities of Collembola, Oribatida, Gamasina and Uropodina were lower on bark than in soil and declined with altitude (Fig. 2); these differences were significant for all microarthropod taxa except for Gamasina (Table 1). Gamasina had higher densities in soil compared with bark at 1,850 and 2,200 m whereas their densities in soil and bark at 2,150 and 2,300 were similar (significant interaction of habitat and altitude; Table 1; Fig. 2c). The community structure of Oribatida (calibrated for density, i.e. the densities of soil and bark living taxa were equalized, which eliminates the factor density for the analysis of the community structure) did not differ significantly between bark and soil (discriminant function analysis; F 3,16 = 0.97; P = 0.43).

Density of a Oribatida, b Collembola, c Gamasina and d Uropodina in soil and on the bark of trees at four altitudes (1,850, 2,000, 2,150, 2,300 m) in a tropical montane rain forest in southern Ecuador. Bars and uneven standard errors represent back-transformed means of log-transformed data (for “Statistical analysis” see text). Note the different scales
Community structure and diversity of Oribatida in soil and on bark
The number of Oribatida species in soil and on bark declined with increasing altitude with a total number of 24, 23, 17 and 13 species at 1,850, 2,000, 2,150 and 2,300 m, respectively. Overall, the most abundant Oribatida groups were Poronota (44% of all individuals), followed by pycnonotic Apheredermata (37%), Mixonomata (13%), Eupheredermata (4%) and Desmonomata (1%). Enarthronota, Opsiopheredermata and dorsodeficient Apheredermata occurred only sporadically.
The community structure of Oribatida changed along the altitudinal gradient (DFA: Wilks’ Lambda 0.0097; F 9,14 = 9.38; P = 0.001), and differed significantly between each of the four altitudes as indicated by Mahalanobis distances (MD). Differences were most pronounced between 2,150 and 2,300 m (MD = 74.11; P = 0.0006), and least pronounced between 1,850 and 2,300 m (MD = 13.37; P = 0.045). PCA with the calibrated densities of Oribatida also showed that the community structure did not gradually change from the lower to the higher altitudes; rather the community structure of Oribatida at lower altitudes was similar to that at the highest altitude (Fig. 3). Galumnidae dominated at 1,850 m, parthenogenetic taxa such as (most) Suctobelbidae, Oppiella nova and Rostrozetes sp. were dominant at 2,000 m; Rhynchoribates ecuadoriensis, Epidamaeus sp. and Brachyoppiella periculosa had their highest relative densities at 2,150 m, whereas the parthenogenetic species Malaconothrus angulatus was the only Oribatida species that had its highest relative abundance at 2,300 m (Fig. 3).

Principal component analysis (PCA) of oribatid mites at four different altitudes (1,850, 2,000, 2,150, 2,300 m) in soil. PCA was carried out with log-transformed data that were calibrated for density. The altitude was included as a passive variable. Eigenvalues of axes one and two were 0.38 and 0.19 respectively
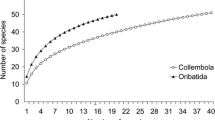
The incidence-based coverage estimator (ICE) of the sample based rarefaction curve stabilized at about 45 species (Fig. 4); the Michaelis–Menten estimate converged to the observed richness of 49 species. The number of expected species given the empirical data (Sobs) ended at 38 species but was not yet saturated. The singleton and doubleton curves were flat and stabilized after about 20 samples.

Sample-based rarefaction curves and corresponding estimators of soil and bark living oribatid mites. ICE, incidence-based coverage estimator; MMMean, asymptote of Michaelis–Menten curve estimated from the sample-based rarefaction curve; Jack 2 Mean, second-order Jackknife richness estimator. Sobs, observed species richness; Singletons, number of species present in only one samples; Doubletons, number of species present in exactly two samples
Discussion
Microarthropod densities in soil
Densities of all microarthropod taxa at the study site were low when compared with those of temperate forests (Maraun and Scheu 2000) indicating that resource limitations are a major factor for the soil microarthropod community. Oribatida were the most abundant microarthropods in the soil and on the bark of the studied tropical montane rain forest, consistent with the general idea that Oribatida commonly are the most abundant microarthropods in forest ecosystems (Petersen and Luxton 1982; Wallwork 1983; Paoletti et al. 1991). Density of Oribatida at the study site (~25,000 ind./m² at 1,850 m) was similar to that in other rain forests (Plowman 1979; Holt 1985; Olson 1994) but also base rich European forests (Luxton 1981; Maraun and Scheu 2000). Collembolan densities were slightly lower than those of oribatid mites and predatory taxa such as Gamasina and Uropodina had even lower densities. Lower densities of predators are in agreement with food web theory since energy is lost during transfer to higher trophic levels.
Density changes with altitude
The density of microarthropods at the study site declined mostly according to altitude, in the order 1,850, 2,000, 2,300 and 2,150 m. A similar decline in microarthropod density along a wider altitudinal gradient (from 1,000 to 3,100 m) at our study site was found by Maraun et al. (2008); in a litter bag field experiment at the study site, densities of microarthropods also declined with altitude (Illig et al. 2008). Declining numbers of litter invertebrates along Neotropical altitudinal gradients were also found in previous studies (Olson 1994; Richardson et al. 2005). Four factors that may be responsible for the low microarthropod densities at higher altitudes are most obvious: (1) higher soil acidity, (2) harsher abiotic conditions (low temperatures, high precipitation, waterlogging), (3) high densities of macroarthropods, which reduce the habitable space and the amount of resources, and (4) lower quantity and quality of resources. At the study site the soil pH indeed decreases from 4.4 to 3.9 at 1,850 and 2,250 m, respectively (Wilcke et al. 2008). However, since Oribatida density generally increases with soil acidity in temperate regions (Maraun and Scheu 2000)—reaching maximum densities of up to 400,000 ind./m2 in acidic boreal forests—the decrease in microarthropod densities with altitude is unlikely to be caused by soil acidity. Low temperatures also are unlikely to be responsible for low density of microarthropods at higher altitudes since temperatures in boreal forests, where densities of microarthropods are high, are considerably lower. Similarly, high humidity also is unlikely to be responsible for low density of microarthropods at higher altitudes since the water content of boreal forest soils also is high. Macroarthropods are also unlikely to negatively affect the densities of soil microarthropods since their density is generally low and decreases with increasing altitude (Illig et al. unpublished data). Organic material accumulates at higher altitudes of our study site (Wilcke et al. 2002), indicating that the amount of resources also does not limit soil microarthropods. Therefore, the most important factor for the low densities of soil microarthropods at high altitudes likely is the low quality of the available resources. Indeed, the C-to-N ratio of the organic material is at a maximum at higher altitudes indicating low resource quality. The low resource quality also results in low decomposition rates at high altitudes (Leuschner et al. 2007; Illig et al. 2008).
Density changes with soil depth
Densities of most microarthropod taxa in lower soil levels remained unaffected by altitude whereas their densities in litter strongly declined with increasing altitude. This finding reflects the soil structure at the study sites. At 1,850 m a litter layer with (comparably) high quality litter exists on top of a mineral soil layer poor in food resources, whereas at higher altitudes dead organic material (litter and roots) accumulates and the upper horizons become more similar.
Densities of Oribatida, Collembola, Gamasina and Uropodina declined with soil depth, which typically is the case in forest and arable soils (Adis et al. 1987; Hijii 1987; Migge et al. 1998). In a rain forest in Queensland 92% of all arthropods sampled to a depth of 16 cm were located in the upper 8 cm (Holt 1985). Fragmentation and mineralisation processes during decomposition of litter material result in homogenisation of soil organic matter with increasing soil depth, reduced habitat complexity, and reduced resource quality. At our study site reduced availability and quantity of resources in deeper soil layers presumably are the most important factor responsible for the low densities of microarthropods in deeper soil layers. Indeed, microbial biomass at the study site decreases with soil depth indicating lower availability of resources (Krashevska et al. 2008).
Densities of oribatid mites and collembolans were highest in the litter and much lower in deeper soil layers, whereas these differences were less pronounced for predatory Uropodina and Gamasina. This indicates that these predatory taxa do not only feed on oribatid mites and collembolans (Peschel et al. 2006; Schneider and Maraun 2009) but also on other taxa such as enchytraeids and nematodes. This conclusion is also supported by recent stable isotope analyses (Pollierer et al. 2009).
Oribatid mite community structure changes with altitude
Similar to density, the community structurce of oribatid mites changed with altitude, but these changes were not linear; rather, community structure at 1,850–2,300 m, i.e. the lowest and highest altitude, were most similar. The similar community structure at 1,850 and 2,300 m was mainly caused by undetermined Oppiidae. Oppiidae are rather euryoekous species, so their occurence at the highest and lowest site is unlikely to indicate specific environmental condition.
Parthenogenetic oribatid mite taxa did not vary consistently along the altitudinal gradient. They occurred at the intermediate altitude (e.g. most Suctobelbidae, Oppiella nova) but also at the highest altitude (e.g. Malaconothrus angulatus). Some of the likely parthenogenetic taxa, e.g. Rostrozetes spp., likely feed on litter and root material, others on fungi, lichens or animal prey (Illig et al. 2005).
Generally, derived groups of Oribatida, such as Poronota and pycnonotic Apheredermata, dominated the Oribatida community. However, species of tropical forests mostly differ from those of temperate and boreals forests (Maraun et al. 2008). One of the main differences between temperate and tropical forest soils is the different type of mycorrhizal fungi (ectomycorrhiza vs. VA mycorrhiza). Potentially, the oribatid species of tropical forests may differ from those of temperate and boreal forests in consuming VA mycorrhizal rather than ectomycorrhizal fungi (Schneider et al. 2005).
Microarthropods in soil as compared to the bark of trees
Generally, the number of soil microarthropods on bark was lower than in soil. This is consistent with results of Karasawa and Hijii (2008) and Proctor et al. (2002), who found lower densities of Oribatida on the bark of trees than in litter of subtropical forests. Low microarthropod abundances on the bark may be due to harsh microclimatic conditions, including variations in temperature, stronger wind, more frequent wet/dry cycles, reduced habitat complexity (Delamare-Deboutteville 1951; Prinzing and Woas 2003), low quantity of resources, or defence mechanisms of resources on the bark (e.g. secondary compounds of lichens). Presumably, a combination of these factors is responsible for the low densities of Oribatida on the bark of trees.
With increasing altitude microarthropod densities on bark did not vary significantly, whereas in soil densities declined. As hypothesized earlier, resource quality declines with altitude in soil suggesting that microarthropod communities are controlled by resource quality whereas abiotic factors such as temperature and humidity are of minor importance.
At our study sites the community structure of oribatid mites on bark did not differ significantly from that in soil, which is in contrast to temperate forests where soil and bark species hardly overlap. This indicates that in the tropics habitat conditions for oribatid mites in soil and on the bark of trees differ little, whereas in temperate regions they differ strongly (Lindo and Winchester 2007a, b). Bark-dwelling oribatid mite taxa and taxa living in tropical soils are mainly sexual whereas parthenogens dominate in temperate soils (Maraun et al. 2008). Presumably, oribatid mites on bark and in tropical soils are more strongly limited by resources. Corticolous oribatids consume fungi and lichens growing on the bark surface, which perhaps can be exploited more easily than resources of soil-dwelling species, which have to locate resources in the litter and soil matrix. This hypothesis is supported by the fact that bark-dwelling oribatid mites also occur on lichens, mosses and algae covering the surface of stones (Meier et al. 2002).
The number of Oribatida species declined with altitude from 24, 23, 17 to 13 at 1,850, 2,000, 2,250 and 2,300 m, respectively. Walter (1985) also found the diversity of oribatid mites to decline with elevation in a forest in Oregon. Presumably, low resource quality at higher altitudes and lower numbers of microhabitats (Anderson 1975) are responsible for the low species number of Oribatida at higher altitudes (Illig et al. 2008).
Rarefaction analysis showed that about 50 species of oribatid mites are to be expected at the study site, which is in the range previously reported from this site (Illig et al. 2008). In general, species numbers are similar to those in temperate systems, in contrast to above-ground taxa. Maraun et al. (2008) suggested that the comparatively low diversity of below-ground animals in tropical ecosystems is due to the fact that they do not interact with living resources, i.e. there is a general absence of co-evolutionary processes. This may not apply to fungal feeding soil animals such as oribatid mites and collembolans, since fungi are living food. However, as there seem to be no oribatid mites or collembolans that restrict feeding to particular fungal species the argument still may apply. Why there are no food specialists in soil microarthropods, however, needs further clarification.
References
Adis J, de Morais JW, de Mesquita HG (1987) Vertical distribution and abundance of arthropods in the soils of a neotropical secondary forest during the rainy season. Stud Neotrop Fauna Environ 22:189–197
Andersen AN (1992) Regulation of momentary diversity by dominant species in exceptionally rich ant communities of the Australian seasonal tropics. Am Nat 140:401–420
Anderson JM (1975) Succession, diversity and trophic relationships of some soil animals in decomposing leaf litter. J Animal Ecol 44:475–495
Bachmann S, Baker WJ, Brummitt N, Dransfield J, Moat J (2004) Elevational gradients, area and tropical island diversity: an example from the palms of New Guinea. Ecography 27:299–310. doi:10.1111/j.0906-7590.2004.03759.x
Beck L (1971) Bodenzoologische Gliederung und Charakterisierung des amazonischen Regenwaldes. Amazoniana 3:69–132
Behan VM, Hill SB (1978) Feeding habits and spore dispersal of oribatid mites in the North American arctic. Rev Ecol Biol Sol 15:497–516
Behan-Pelletier VM, Winchester NN (1998) Arboreal oribatid mite diversity: colonizing the canopy. Appl Soil Ecol 9:45–51
Brehm G, Fiedler K (2003) Faunal composition of geometrid moths changes with altitude in an Andean montane rain forest. J Biogeogr 30:431–440
Brehm G, Fiedler K (2004) Bergmann’s rule does not apply to geometrid moths along an elevational gradient in an Andean montane rain forest. Global Ecol Biogeogr 13:7–14. doi:10.1111/j.1466-882X.2004.00069.x
Brehm G, Pitkin LM, Hilt N, Fiedler K (2005) Montane Andean rain forests are a global diversity hotspot of geometrid moths. J Biogeogr 32:1621–1627. doi:10.1111/j.1365-2699.2005.01304.x
Brühl C, Mohamed M, Linsenmair KE (1999) Altitudinal distribution of leaf litter ants along a transect in primary forests on Mount Kinabalu, Sabah, Malaysia. J Trop Ecol 15:265–277
Colwell RK (1994–2004) Estimates: statistical estimation of species richness and shared species from samples. http://viceroy.eeb.uconn.edu/estimates
Delamare-Deboutteville C (1951) Microfaune du sol des pays tempérés et tropicaux. Vie Milieu, Suppl 1:1–360
Erdmann G, Floren A, Linsenmair KE, Scheu S, Maraun M (2006) Little effect of forest age on oribatid mites on the bark of trees. Pedobiologia 50:433–441. doi:10.1016/j.pedobi.2006.08.004
Erdmann G, Otte V, Langel R, Scheu S, Maraun M (2007) The trophic structure of bark-living oribatid mite communities analysed with stable isotopes (15N, 13C) indicates strong niche differentiation. Exp Appl Acarol 41:1–10. doi:10.1007/s10493-007-9060-7
Grandjean F (1953) Essai de classification des Oribates (Acariens). Bull Soc Zool France 78:421–446
Grandjean F (1965) Complément a mon travail de 1953 sur la classification des oribates. Acarologia 7:713–734
Grandjean F (1969) Considérations sur le classement des Oribates: Leur division en 6 groupes majeurs. Acarologia 11:127–153
Hijii N (1987) Seasonal changes in abundance and spatial distribution of the soil arthropods in a Japanese cedar (Cryptomeria japonica D. Don) plantation, with special reference to Collembola and Acarina. Ecol Res 2:159–173
Holt JA (1985) Acari and Collembola in the litter and soil of three north Queensland rainforests. Austr J Ecol 10:57–65
Illig J, Langel R, Norton RA, Scheu S, Maraun M (2005) Where are the decomposers? Uncovering the soil food web of a tropical montane rain forest in southern Ecuador using stable isotopes (15N). J Trop Ecol 21:589–593. doi:10.1017/S0266467405002646
Illig J, Schatz H, Scheu S, Maraun M (2008) Decomposition and colonization by micro-arthropods of two litter types in a tropical montane rain forest in southern Ecuador. J Trop Ecol 24:157–167. doi:10.1017/S0266467407004750
Janzen DH, Ataroff M, Farinas M, Reyes S, Rincon A, Soler A, Soriano P, Vera M (1976) Changes in the arthropod community along an elevational transect in the Venezuelan Andes. Biotropica 8:193–203
Jongman RHG, ter Braak CJF, van Tongeren OFR (1995) Data analysis in community and landscape ecology. Cambridge University Press, Cambridge
Karasawa S, Hijii N (2008) Vertical stratification of oribatid (Acari: Oribatida) communities in relation to their morphological and life-history traits and tree structures in a subtropical forest in Japan. Ecol Res 23:57–69. doi:10.1007/s11284-007-0337-4
Kempson D, Lloyd M, Ghelardi R (1963) A new extractor for woodland litter. Pedobiologia 3:1–21
Krashevska V, Bonkowski M, Maraun M, Scheu S (2007) Testate amoebae (Protista) of an elevational gradient in the tropical montane rain forest of Ecuador. Pedobiologia 51:319–331. doi:10.1016/j.pedobi.2007.05.005
Krashevska V, Bonkowski M, Maraun M, Ruess L, Kandeler E, Scheu S (2008) Microorganisms as driving factors for the community structure of testate amoebae along an altitudinal gradient in tropical mountain rain forests. Soil Biol Biochem 40:2427–2433. doi:10.1016/j.soilbio.2008.06.004
Lee SM, Chao A (1994) Estimating population-size via sample coverage for closed capture-recapture models. Biometrics 50:88–97
Legendre P, Legendre L (1998) Numerical ecology. Elsevier, Amsterdam
Leuschner C, Moser G, Bertsch C, Röderstein M, Hertel D (2007) Large altitudinal increase in tree root/shoot ratio in tropical montane forests of Ecuador. Basic Appl Ecol 8:219–230. doi:10.1016/j.baae.2006.02.004
Lindo Z, Winchester NN (2006) A comparison of microarthropod assemblages with emphasis on oribatid mites in canopy suspended soils and forest floors associated with ancient western redcedar trees. Pedobiologia 50:31–41. doi:10.1016/j.pedobi.2005.09.002
Lindo Z, Winchester NN (2007a) Resident corticolous oribatid mites (Acari: Oribatida): decay in community similarity with vertical distance from the ground. Ecoscience 14:223–229. doi:10.2980/1195-6860(2007)14[223:RCOMAO]2.0.CO;2
Lindo Z, Winchester NN (2007b) Oribatid mite communities and foliar litter decomposition in canopy suspended soils and forest floor habitats of western redcedar forests, Vancouver Island, Canada. Soil Biol Biochem 39:2957–2966. doi:10.1016/j.soilbio.2007.06.009
Longino JT, Coddington J, Colwell RK (2002) The ant fauna of a tropical rain forest: estimating species richness three different ways. Ecology 83:689–702
Luxton M (1981) Studies on the oribatid mites of a Danish beech wood soil III. Introduction to the field population. Pedobiologia 21:301–311
Maraun M, Scheu S (2000) The structure of oribatid mite communities (Acari, Oribatida): patterns, mechanisms and implications for future research. Ecography 23:374–783
Maraun M, Visser S, Scheu S (1998) Oribatid mites enhance the recovery of the microbial community after a strong disturbance. Appl Soil Ecol 9:175–181
Maraun M, Schatz H, Scheu S (2007) Awesome or ordinary? Global diversity patterns of oribatid mites. Ecography 30:209–216. doi:10.1111/j.2007.0906-7590.04994.x
Maraun M, Illig J, Sandmann D, Krashevska V, Norton RA, Scheu S (2008) Soil Fauna: composition and function. In: Beck E, Bendix J, Kottke I, Makeschin F, Mosandl R (eds) Gradients in a tropical montane ecosystem of Ecuador. Ecological Studies vol 198, Springer, Berlin, pp 181–192
Meier FA, Scherrer S, Honegger R (2002) Faecal pellets of lichenivorous mites contain viable cells of the lichen-forming ascomycete Xanthoria parietina and its green algal photobiont Trebouxia arboricola. Biol J Linn Soc 76:259–268. doi:10.1046/j.1095-8312.2002.00065.x
Migge S, Maraun M, Scheu S, Schaefer M (1998) The oribatid mite community (Acarina) on pure and mixed stands of beech (Fagus sylvatica) and spruce (Picea abies) at different age. Appl Soil Ecol 9:119–126
Mitchell MJ (1978) Vertical and horizontal distributions of oribatid mites (Acari: Cryptostigmata) in an aspen woodland soil. Ecology 59:516–525
Myers N, Mittermeier RA, Mittermeier CG, de Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858. doi:10.1038/35002501
Olson DM (1994) The distribution of leaf litter invertebrates along a Neotropical altitudinal gradient. J Trop Ecol 10:129–150
Palacios-Vargas JG, Castano-Meneses G, Gomez-Anaya JA, Martinez-Yrizar A, Mejia-Recamier BE, Martinez-Sanchez J (2007) Litter and soil arthropod diversity and density in a tropical dry forest ecosystem in Western Mexico. Biodiv Conserv 16:3703–3717. doi:10.1007/s10531-006-9109-7
Pande YD, Berthet P (1975) Observations on the vertical distribution of soil Oribatei in a woodland soil. Trans Roy Entomol Soc Lond 127:259–275
Paoletti MG, Taylor RAJ, Stinner BR, Stinner DH, Benzing DH (1991) Diversity of soil fauna in the canopy and forest floor of a Venezuelan cloud forest. J Trop Ecol 7:373–383
Peschel K, Norton RA, Scheu S, Maraun M (2006) Do oribatid mites live in enemy-free space? Evidence from feeding experiments with the predatory mite Pergamasus septentrionalis. Soil Biol Biochem 38:2985–2989. doi:10.1016/j.soilbio.2006.04.035
Petersen H, Luxton M (1982) A comparative analysis of soil fauna populations and their role in decomposition process. Oikos 39:288–388
Plowman KP (1979) Litter and soil fauna of two Australian subtropical forests. Austr J Ecol 4:87–104
Plowman KP (1981) Distribution of Cryptostigmata and Mesostigmata (Acari) within the litter and soil layers of two subtropical forests. Austr J Ecol 6:365–374
Pollierer M, Langel R, Scheu S, Maraun M (2009) Compartmentalization of the soil animal food web as indicated by dual analysis of stable isotopes (15N and 13C). Soil Biol Biochem 41:1221–1226. doi:10.1016/j.soilbio.2009.03.002
Prinzing A, Woas S (2003) Habitat use and stratification of Collembola and oribatid mites. In: Basset Y, Novotny V, Miller SE, Kitching RL (eds) Arthropods of tropical forests: spatio-temporal dynamics and resource use in the canopy. Cambridge University Press, Cambridge, pp 271–282
Proctor HC, Montgomery KM, Rosen KE, Kitching RL (2002) Are tree trunks habitats or highways? A comparison of oribatid mite assemblages from hoop-pine bark and litter. Austr J Entomol 41:294–299. doi:10.1046/j.1440-6055.2002.00309.x
Rahbek C (2005) The role of spatial scale and the perception of large-scale species-richness patterns. Ecol Lett 8:224–239. doi:10.1111/j.1461-0248.2004.00701.x
Renker C, Otto P, Schneider K, Zimdars B, Maraun M, Buscot F (2005) Oribatid mites as potential vectors for soil microfungi: study of mite-associated fungal species. Microb Ecol 50:518–528. doi:10.1007/s00248-005-5017-8
Richardson BA, Richardson MR, Soto-Adames FN (2005) Separating the effects of forest type and elevation on the diversity of litter invertebrate communities in a humid tropical forest in Puerto Rico. J Animal Ecol 74:926–936. doi:10.1111/j.1365-2656.2005.00990.x
Röderstein M, Hertel D, Leuschner C (2005) Above- and below-ground litter production in three tropical montane forests in southern Ecuador. J Trop Ecol 21:483–492. doi:10.1017/S026646740500249X
Schneider K, Maraun M (2009) Top-down control of soil microarthropods–evidence from a laboratory experiment. Soil Biol Biochem 41:170–175. doi:10.1016/j.soilbio.2008.10.013
Schneider K, Migge S, Norton RA, Scheu S, Langel R, Reineking A, Maraun M (2004) Trophic niche differentiation in soil microarthropods (Oribatida; Acari): evidence from stable isotope ratios (15N/14N). Soil Biol Biochem 36:1769–1774. doi:10.1016/j.soilbio.2004.04.033
Schneider K, Renker C, Maraun M (2005) Oribatid mite (Acari, Oribatida) feeding on ectomycorrhizal fungi. Mycorrhiza 16:67–72. doi:10.1007/s00572-005-0015-8
Ter Braak CJF, Šmilauer P (1998) CANOCO reference manual and user’s guide to CANOCO for Windows: software for canonical community ordination (version 4). Microcomputer Power, Ithaca, New York, USA
Usher MB (1970) Seasonal and vertical distribution of a population of soil arthropods: Collembola. Pedobiologia 10:224–236
Wallwork JA (1983) Oribatids in forest ecosystems. Ann Rev Entomol 28:109–130
Walter DE (1985) The effect of litter type and elevation on colonization of mixed coniferous litterbags by oribatid mites. Pedobiologia 28:383–387
Wilcke W, Yasin S, Abramowski U, Valarezo O, Zech W (2002) Nutrient storage and turnover in organic layers under tropical montane rain forest in Ecuador. Europ J Soil Sci 53:15–27. doi:10.1046/j.1365-2389.2002.00411.x
Wilcke W, Yasin S, Schmitt A, Valarezo C, Zech W (2008) Soils along the altitudinal transect and in catchments. In: Beck E, Bendix J, Kottke I, Makeschin F, Mosandl R (eds) Gradients in a tropical montane ecosystem of Ecuador. Ecological Studies vol 198, Springer, Berlin, pp 75–85. doi: 10.1007/978-3-540-73526-7
Wunderle I (1992) Arboricolous and edaphic Oribatei (Acari) in the lowland rain-forest of Panguana, Peru. Amazoniana 12:119–142
Acknowledgments
Financial support by the German Research Foundation (DFG) is gratefully acknowledged. We thank Heinz Schatz for help in determination of oribatid mite species, Dorothee Sandmann for help during field work, and Melanie M. Pollierer for reading and commenting on an earlier version of the manuscript.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Illig, J., Norton, R.A., Scheu, S. et al. Density and community structure of soil- and bark-dwelling microarthropods along an altitudinal gradient in a tropical montane rainforest. Exp Appl Acarol 52, 49–62 (2010). https://doi.org/10.1007/s10493-010-9348-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-010-9348-x