
Abstract
In recent years, research on Physarum polycephalum has become more popular after Nakagaki (AIR 407: 6803-470, 2000) performed their famous experiment showing that Physarum was able to find the shortest route through a maze. Subsequent researches have confirmed the ability of Physarum-inspired algorithms to solve a wide range of real-world applications. In contrast to previous reviews that either focus on biological aspects or bio-inspired applications, here we present a comprehensive review that highlights recent Physarum polycephalum biological aspects, mathematical models, and Physarum bio-inspired algorithms and their applications. The novelty of this review stems from our exploration of Physarum intelligent behaviour in competition settings. Further, we have presented our new model to simulate Physarum in competition, where multiple Physarum interact with each other and with their environments. The bio-inspired Physarum in competition algorithms proved to have great potentials for future research.
Similar content being viewed by others

Explore related subjects
Find the latest articles, discoveries, and news in related topics.Avoid common mistakes on your manuscript.
1 Introduction
Bio-inspired computing focuses on extracting computational models for problem solving from in-depth understanding of behaviour and mechanisms of biological systems. In recent years, cellular computational models based on the structure and the processes of living cells, such as bacterial colonies (Kim et al. 2007) and viral models (Cortés et al. 2008) have become an important line of research in bio-inspired computing. Physarum-computing, as an example of cellular computing model, has attracted the attention of many researchers (Tero et al. 2010). Physarum polycephalum (Physarum for short) is an example of plasmodial slime moulds that are classified as a fungus "Myxomycetes" Cavender (1995). In recent years, research on Physarum-inspired computing has become more popular since (Nakagaki et al. (2000)) performed their well-known experiments showing that Physarum was able to find the shortest route through a maze (Nakagaki et al. 2000). Recent research has confirmed the ability of Physarum-inspired algorithms to solve a wide range of problems (Zhang et al. 2016b; Sun 2017).
Physarum can be modelled as a reaction-diffusion system (cytoplasmic liquid) encapsulated in an elastic growing membrane of actin-myosin cytoskeleton (Adamatzky 2009a). In the early stages of growth (i.e., the exploration phase), the Physarum foraging behaviour results in the generation of a branching pattern. In the second phase (i.e., the exploitation phase), it spans the sources of nutrients with a dynamic proximity graph and forms a pattern similar to Voronoi diagram (Werner 2019). This characteristic of continuous change in Physarum protoplasmic flux with the change of environment allows Physarum-inspired algorithms to have great potentials in dealing with graph-optimisation problems (Zhang et al. 2016b).
Physarum can be considered as one of the biological models of unconventional computation. Computer scientists are investigating the potential of Physarum as programmable biological machine (Adamatzky 2010). Physarum is capable of decision-making and information processing that can lead to the emergence of complex social behaviour (Latty and Beekman 2010; Dussutour et al. 2010; Reid et al. 2016). It compares the relative qualities of multiple options and combines the information on reward in order to make correct and adaptive decisions. Physarum is also capable of memorising and anticipating repeated events, and displays both short and long term habituation, as a simple form of learning (Beekman and Latty 2015; Boussard et al. 2019).
In deed, Physarum can be considered as one of the biological example of unconventional computation capable of creating a programmable Physarum machine (Adamatzky 2010). It has been studied in the project "Physarum chip: growing computers from slime mould" (Adamatzky et al. 2012) that ran between 2013 and 2016. The Physarum chip is expected to solve a wide range of computation tasks, including graph optimisation, logic and arithmetical computing (Andrew and Krzysztof 2016). The EU-funded project “Physarum Sensor: Biosensor for Citizen Scientists” is an extension of the PhyChip project (PhySense 2019). This project showed that Physarum is an ideal biological substrate that could be used as biosensors that convert a biological response into an electrical signal. These low-cost biosensors can be used for various applications, including environmental monitoring and health (CORDIS 2019).
Several reviews on Physarum have been published, however, they either focus on biological aspects (Cavender 1995; Reid and Latty 2016) or mathematical models and bio-inspired applications (Zhang et al. 2016b; Sun 2017; Adamatzky 2010; Gao et al. 2019). The novelty of this review stems from a comprehensive survey that summarises the latest published literature on Physarum covering biological behaviour, reflection on modelling, and computing aspects. Further, we have covered Physarum-inspired applications from several aspects. In contrast to other Physarum review papers that focus mainly on Physarum-inspired algorithms to solve graph optimisation problems, we have discussed a second aspect of application which is taking advantage of Physarum characteristics, such as morphological diversity (Gunji et al. 2008) and positive feedback loop (Tero et al. 2005), that will lead to the development of hybrid algorithms that optimise evolutionary algorithms to improve its efficiency and robustness (Zhang et al. 2014a; Gao et al. 2018). In the third aspect of Physarum applications, we demonstrated Physarum as a method of biological computing that has been extensively studied in "Physarum chip: growing computers from slime mould" (Adamatzky et al. 2012), and the PhySense project "Physarum Sensor: Biosensor for Citizen Scientists" (CORDIS 2019). Finally, we presented our Physarum-inspired competition algorithms and explored its applications in solving wireless sensor networks (WSN), and discrete multi-objective optimisation problems(DMOO).
In this review we have presented our novel model that simulate Physarum in competition settings. To the best of our knowledge, we are the first to explore Physarum intelligent behaviour in competition settings, unlike the other models based on a single Physarum. Multiple Physarum with autonomous behaviours react to each other and with their environment, this has allowed the efficient exploration of the whole system evolving to an optimal global network and each Physarum to move to a better position. Further, it has allowed us to deal with the increasingly proposed networks scenarios with multiple sources and multiple sinks. The bio-inspired Physarum competition algorithms proved to have great potentials in dealing with graph-optimisation problems in a dynamic environment as in Mobile Wireless Sensor Networks (Awad et al. 2019b), and Discrete Multi-Objective Optimisation problems (Awad et al. 2019c).
We will start by giving a short overview of bio-inspired computing (Sect. 2). For deep understanding of Physarum biological foraging behaviour we will review the biological aspects of Physarum including its intelligent foraging behaviour, collective swarm behaviour, and competitive behaviour (Sects. 3, 4, 5, and 6). Then we will present some of the most well-known real biological experiments and mathematical models (Sect. 7 and Sect. 8). Furthermore, we will present some of the real-world applications that have been solved by Physarum-inspired algorithms (Sect. 9). Finally, a conclusion is given in Sect. 10.
2 Bio-inspired computing
The inspiration from biology and nature has always been one of the most important and exhaustless sources for researchers and engineers to develop novel algorithms and innovative techniques during the past decades. Earlier works on bio-inspired computing focus on extracting the computational models from complex high-level biological systems of cognition and understanding. Under this umbrella of computational intelligence, there are many paradigms such as artificial neural networks, genetic algorithm, and artificial immune system. These models are based on imitating the behaviour of central nervous system, chromosomal reproduction, and immunity against infection, respectively (Tan and Shi 2017). In recent years, simple cellular computational models based on the structure and the processes of living cells became an essential branch of bio-inspired computing, such as bacterial colonies (Kim et al. 2007) and viral models Cortés et al. (2008). Physarum is an example of a cellular computing model attracting researchers’ attention (Tero et al. 2010).
Two typical categories of bio-inspired algorithms are evolutionary algorithms and swarm intelligence algorithms, which are inspired by the natural evolution and collective behaviour in swarms of animals, respectively (Tan and Shi 2017). However, there are several existing limitations of these optimisation methodologies (Wong 2015). Evolutionary algorithms use iterative progresses in a population in response to environmental pressure that causes natural selection, and this causes an increase in the fitness of the population. Genetic Algorithm (GA) (Holland 1973) is an example of evolutionary algorithms. Swarm intelligence is one of the most exciting topics dealing with the collective behaviour of decentralised and self-organised biological systems. It consists of a population of simple agents which can communicate locally with each other and their local environment. These interactions can lead to the emergence of hugely complicated global behaviour (Tan and Shi 2017). A variety of swarm intelligence algorithms for optimisation problems, such as particle swarm optimisation (Eberhart and Kennedy 1995), ant colony optimisation (Dorigo et al. 1996), and Artificial Bee Colony (ABC) (Karaboga and Basturk 2007), have been developed with increasingly wide applications in the real world.
3 Slime mould (Physarum polycephalum) biology and foraging behaviour
Slime mould was classified as a fungus, a class of Myxomecetes, but now it is considered to be part of kingdom Protista. There are two main types of slime moulds: the cellular slime moulds, and the plasmodial slime moulds. The cellular slime moulds are formed of multiple cells, whereas the plasmodial slime moulds are formed of a large multi-nucleated single cell with thousands of nuclei without any membrane between them (Reid and Latty 2016; Cavender 1995). Physarum polycephalum (Physarum for simplicity) is an example of plasmodial slime moulds; it consists of a single cell amoeba-like organism and has a simple structure which can be easily modelled (compared to others like ants or bees). Physarum strains are not related to fungi and form a genuine branch in the evolution tree of life, other than fungi. More than 800 slime mould species exist worldwide (Adamatzky et al. 2012).
The primitive intelligence of Physarum is mostly demonstrated during its vegetative stage when it turns into plasmodium. In this stage, it forms a yellowish vascular network which expands up to tens of centimetres in search of food to connect the food source (e.g., oat flakes) with the Physarum body (Jones and Adamatzky 2014). Physarum can be considered as a parametric bio-blob that presents itself as a geometrically smart adaptive graph structure (Werner 2019). It is formed of a mycelial tubular network through which the chemical and physical signals, the nutrients, and the body mass are transported throughout the organism. The tubes of plasmodium Physarum are made of a gel-like outer membrane of actin-myosin cytoskeleton that generates periodic contractions of the tube walls. Inside this membrane, the cytoplasmic liquid is pumped back and forth in a rhythmically oscillating manner. The contraction amplitude and the frequency generally increase or decrease when encountering an attractant or repellent, respectively (Durham and Ridgway 1976; Ueda et al. 1976a).
The Physarum senses gradients of chemoattractants and repellents and forms a yellowish vascular network in search of nutrition (Durham and Ridgway 1976; Ueda et al. 1976b). It responds to stimulation by changing patterns of electrical potential oscillations, and it is made of hundreds to thousands of biochemical oscillators (Durham and Ridgway 1976). A stimulus triggers the release of a signalling molecule cyclic adenosine monophosphate (cAMP) (Dormann et al. 2000) that starts cytoplasmic streaming. This stimulus gives rise to propagating waves resulting in increased cytoplasmic streaming (shuttling) through that vein (Durham and Ridgway 1976; Ueda et al. 1976a). This generates a positive feedback loop; the higher the rate of cytoplasmic streaming is, the thicker the vein becomes (Alim et al. 2017).
The Physarum foraging behaviour consists of two simultaneous self-organised processes: expansion (exploration) and contraction (exploitation). Physarum structure reveals two distinct geometric patterns: (a) Physarum develops thin branches, searching in their environment for food, (b) the bulging droplet-like blobs enlargement at the tips of the branches (Werner 2019). In the early stages (exploration phase) the organism grows, and the branches with the bulging blobs at their tips become longer through the foraging process, and they divide into further branches and link up like veins. In the second phase (exploitation phase) the tubes that transport the nutrients will grow bigger while the tubes which do not transport enough nutrients will vanish and disappear (Nakagaki et al. 2001; Werner 2019).
4 Physarum intelligent behaviour
Physarum may not have a brain, but computer scientists are investigating its potential as novel, unconventional computers (Adamatzky 2010). Physarum is capable of making complex foraging decisions based on trade-offs between risks, hunger level and food patch quality (Latty and Beekman 2010, 2011; Dussutour et al. 2010; Saigusa et al. 2008; West et al. 2007). The primitive intelligence of Physarum Polycephalum (slime mould) is mostly demonstrated during its plasmodium stage (a large multi-nucleated single cell). The underlying mechanisms of Physarum intelligence and cognition are based on the way with which the organism perceives the environment, integrates this information and makes decisions (Reid et al. 2016). This has motivated many researchers to take inspiration from their biological phenomena to come up with a novel, biologically inspired models for unconventional computational methods capable of solving many NP-hard problems (Adamatzky 2010). In what follows we will summarise the Physarum intelligent behaviours.
4.1 Finding shortest path
This intelligent behaviour was first observed by Nakagaki et al. (2000) Nakagaki et al. (2000). Physarum was able to find the shortest path between two selected points (source node, and sink node) in a maze-solving problem. Bonifaci et al. (2012) proved that under this model, the mass of the Physarum will eventually converge to the shortest path of the network that Physarum lies on, independently of the network topology or of the initial mass distribution (Bonifaci et al. 2012). Other examples of the shortest path approach may include the towers of Hanoi problem (Reid and Beekman 2013).
4.2 Building high-quality networks
Physarum’s network design ability has attracted the attention of many researchers as it showed excellent ability in network construction without central consciousness during foraging process (Adamatzky 2012b; Beekman and Latty 2015; Reid et al. 2016). In the early stages (exploration phase) the organism’s branches grow and the bulging blobs divide into further branches and link up like veins. In the second phase (exploitation phase) the organism eventually spans the sources of nutrients with a dynamic proximity graph (Voronoi pattern), where the links (edges) connect the corresponding nodes (vertices). This network architecture is highly dynamic with flexible rearrangement of its junctions, and once the Physarum moves, the location, size of the vertices and the edges changes, disappear, or new links and vertices (nodes) develop (Werner 2019). One of the most well-known real experiments that showed the intelligence of Physarum for network design was the reconstruction of Tokyo railway network designed by Physarum (Tero et al. 2010).
4.3 Adapting to changing environments
Many biological experiments have shown that Physarum networks disassemble and reassemble within a period of a few hours in response to the change of external conditions (e.g., chemotaxis, phototaxis and thermotaxis) (Ito et al. 2010). Moreover, (Adamatzky (2009c)) has shown that Plasmodium-based computing devices can be precisely controlled and shaped by illumination (Adamatzky 2009c). Jones et al. (2017) have demonstrated how a growth parameter in the model can be used to transit between Convex and Concave Hulls (Jones et al. 2017). These results demonstrated how Physarum can approximate the external and internal shape of a set of points using chemo-attractant stimuli and masking by light illumination (repellent).
5 Physarum collective behaviour and swarm intelligence
Physarum exhibits swarm intelligence and social behaviour as social insects and animals. It shares with these insects and animals many common features of collective behaviour, such as synchronisation, communication, positive feedback, distributed intelligence, and spatial memory (Reid and Latty 2016). Physarum’s collective behaviour is the result of communication and interactions among its individual units. Being a single-cell organism, Physarum individual units do not have a ‘choice’ to behave selfishly; rather, they communicate together via chemical transmitter namely cyclic adenosine monophosphate (cAMP) signals/oscillators which coordinate and synchronise Physarum’s slug behaviour. A stimulus triggers the release of a signalling molecule cAMP that results in changing patterns of electrical potential oscillations which starts cytoplasmic streaming. This is distinct from other social animals such as bees or ants which use other types of communication (e.g., pheromone for ants) (Reid and Latty 2016).
The following points will summarise the Physarum collective behaviour and swarm intelligence.
5.1 Synchronisation and communication
The plasmodium (Physarum) shows synchronous oscillation of cytoplasm throughout all its parts that behave cooperatively for exploring the space, searching for nutrients, and optimising the network of streaming protoplasm. Each tiny oscillator is a segment of a tubule network, which is actively expanding and contracting as a form of distributed, collective behaviour that allows Physarum to make complex decisions when exploring its environment. This response causes the cytoplasm to flow in the direction of the attractant and away from repellent (Ueda et al. 1976b).
5.2 Feedback mechanism
Physarum protoplasm migrates towards the area of the highest cAMP concentration and at the same time starts secreting cAMP. This behaviour creates a positive feedback loop, which will cause protoplasmic tubes with high cAMP levels to grow bigger and those with low cAMP levels to disappear gradually due to lack of flow (Nakagaki et al. 2001). The tubes that are more suitable for transporting the nutrients will grow bigger and will be of less resistance. On the other hand, the tubes which do not transport enough nutrients will eventually vanish and disappear. This feedback mechanism makes Physarum intelligent enough to maximise the number of nutrient sources and minimise transportation costs (Nakagaki et al. 2001; Werner 2019). However, such positive feedback in Physarum is weaker than the ant colonies in the same maze problem, this will allow Physarum to discover and utilise new solutions and prevent the convergence on a single best solution (Reid et al. 2013).
5.3 Distributed intelligence
Physarum may not have a central information processing unit like a brain, but rather a collection of similar parts of protoplasm. Physarum has recently emerged as a model system for studying information processing and problem-solving in non-neuronal organisms (Oettmeier et al. 2017). Physarum is a system describing the characteristics of a liquid geometry computer in conversation with its environment to survive (Werner 2019). This type of intelligence is now considered as a part of the theme "Liquid brains: How distributed cognitive architectures process information" (Boussard et al. 2019). Thus, Physarum is an excellent candidate for research on autonomous distributed network optimisation (Seki et al. 1998).
5.4 Memorising and learning
Both learning and memory are essential features for animals to survive, and information on past experiences is used for optimal decision-making in a dynamic environment. Physarum is capable of memorising and anticipating repeated events. This intelligent behaviour was first revealed by Saigusa et al. (2008) Saigusa et al. (2008). Moreover, Shirakawa et al. (2011) used an associate learning experiment to test this ability further (Shirakawa et al. 2011). Physarum secretes a trail of slime following movement, which acts as an extra-cellular spatial memory. This increases foraging efficiency of Physarum by avoiding previously explored areas (Liu et al. 2015a; Reid et al. 2012). Physarum displays both short and long-term habituation as a simple form of learning. The information acquired during the habituation, even to chemical repellents, is via constrained absorption of these chemicals to be used as a "circulating memory" (Boussard et al. 2019).
6 Physarum competitive foraging behaviour
6.1 Competition
Competition is generally considered as negative effects caused by the presence of competitors, usually leading to the reduction of available resources. However, competition can also yield lower overall costs, better quality, more choices and varieties, more innovation, greater efficiency, and productivity (Fischer 2008). Competition can be classified into exploitation competition and exclusion competition based on the interactions of the competitors (Verhoef and Morin 2010). Exploitation competition happens when a resource that is in short supply is reduced by other competitors. This will negatively affect another competitors using the same resource. Only the more powerful competitors can obtain this limited opportunity. Exclusion competition regulates population density by slowing down the population increase if the population density is high and vice versa. Competition is very important in driving natural selection as a superior competitor can eliminate inferior ones from the area, resulting in competitive exclusion (Mohseni et al. 2014).
6.2 Physarum foraging behaviour in competition settings
There is increasing evidence that a simple organism like Physarum has complex social behaviours including cooperation and competition (Latty and Beekman 2011; Masui et al. 2018; Schumann et al. 2015; Senturk et al. 2012). Physarum is capable of making complex foraging decisions based on trade-offs between risks, hunger level and food patch quality (Latty and Beekman 2011). The skills of individual competitors are effective methods for inspiration to develop intelligent systems and to provide solutions for decision-making problems. Competitions between multiple Physarum is based on Physarum power (genotype), mass, and the availability of nearby food resources (Schumann et al. 2015). Physarum always initiates foraging behaviour quicker in the presence of competitors (Stirrup and Lusseau 2019).
A recent study by Masui et al. (2018) Masui et al. (2018) has provided an answer to a crucial question: can Physarum identify allogeneic individuals? The answer is yes, allorecognition implicitly promotes the Physarum’s ability of to distinguish its own tissues from those of another, when encountering different individuals. In early research, people adopted the hypothesis that Plasmodium allorecognition was based on the premise of contact, and the slime sheath is just regarded as a simple repellent (Reid et al. 2012; Tsuda et al. 2004). However, the recent study by Masui et al. (2018) Masui et al. (2018) has indicated that the slime sheath is a substance that disperses allorecognition information about itself into the environment. This view led to a new self-extension model, in which the mechanism of non-contact allorecognition using a slime sheath expands the plasmodium opportunities for decision-making, which frequently enables early and safe avoidance rather than fusion (Masui et al. 2018).
7 Physarum real biological experiments
Many experiments have been made to reveal Physarum intelligence. From a computer science point of view, the objective of creating such experiments is to build a mathematical model inspired by real biological experiments to solve real-world optimisation problems. We have summarised some of these biological experiments in Table 1.
7.1 Physarum solving maze experiment
Nakagaki et al. (2000) designed a biological experiment where a Physarum was capable of solving a maze (Nakagaki et al. 2000). The goal of the experiment was to demonstrate the intelligent behaviour of a single Physarum capable of finding the shortest path between two points. In this experiment, there was only one Physarum and one food resource (i.e., solving the shortest path problem).
7.2 Physarum network construction experiment
Tero et al. (2010) designed a biological experiment to simulate the Physarum network formation for the Tokyo railway network and other cities (Tero et al. 2010). The goal of the experiment was to demonstrate the intelligent behaviour of a single Physarum (as a representation of Tokyo) capable of finding the minimum spanning tree that covers all points of multiple food resources (as a representation to other Japanese cities).
7.3 Physarum shape representation experiment
The behaviour of the plasmodium is mediated by environmental stimuli. Jones and Adamatzky (2014) demonstrated how a growth parameter in the model can be used to achieve transition between convex and concave hulls (Jones and Adamatzky 2014). These results suggested novel mechanisms of morphological computation mediated by environmental stimuli and demonstrated how Physarum polycephalum can approximate the external and internal shape of a set of points using chemo-attractant stimuli and masking by light illumination.
7.4 Physarum living cellular automata experiment
The majority of these experiments have focused on Physarum behaviour in an open space (Petri dish). However, investigating the Physarum behaviour in a closed space will help us to understand how the organism makes its decision in a stepwise transition. To accomplish this goal, Shirakawa et al. (2015) have developed an experimental setup to discretise the motility of the plasmodium, and the motility was forced to be a stepwise one transition (Shirakawa et al. 2015). In this way the behaviour of the plasmodium was similar to that of a two-dimensional cellular automaton. They analysed the motility of only a single Physarum with no source of attraction (food source). They postulated several models (transition rules) of Physarum movement based on the statistical results of several experiment runs.
7.5 Physarum electrical activity experiment
Traversa et al. (2013); Whiting et al. (2014) designed a real biological experiment where they measured the electrical activity of Physarum in the presence of stimuli (one food source) (Traversa et al. 2013; Whiting et al. 2014). The goal of the experiment was to show how the Physarum changes patterns of its electrical activity when exposed to attractants and repellents. Based on the fact that Physarum learn and adapt to periodic changes in its environment (Liu et al. 2015a; Reid et al. 2012). Gale et al. (2014) demonstrated that the protoplasmic tubes of the Physarum showed current versus voltage characteristics that is consistent with ideal memristor-systems (Gale et al. 2014). Memristor has been previously used in oscillation-based circuits and systems able to solve computationally hard problems (Ntinas et al. 2017b, a) and emerge neuron-like behaviour (Ntinas et al. 2018). For reviewing the foundations of memristor theory and applications, refer to Erokhin (2014) (Erokhin 2014).
Ntinas et al. (2017b) designed a real experiment similar to that of Whiting et al. (2014); where they presented a bio-inspired memristor-based circuit maze-solving approach. They measured the oscillatory electrical activity of Physarum in a maze structure, the goal of the experiment is to develop a memristor-based circuit models based on the fact that Physarum learn and adapt to periodic changes in its environment. Based on the wave-propagation information processing in the reaction-diffusion media, Fyrigos et al. (2019) Fyrigos et al. (2019) utilised the nano-electronic element memristor to design innovative circuits of electronic excitable medium to perform both classical (Boolean) calculations and to model neuromorphic computations in the same Memristor-RLC (M-RLC) re-configurable network (Fyrigos et al. 2019).
7.6 Physarum solving the two-armed bandit problem experiment
The two-armed bandit problem has previously only been used to study organisms with brains. Yet Physarum, a brainless unicellular organism, showed the ability of decision-making and solved the two-armed bandit problem. In this experiment, Physarum was challenged with a choice between two deferentially rewarding environments, where the arm with the greater number of food resources or higher quality was designated as the high-quality (HQ) arm, and the other arm with fewer food resources or low quality was designated as the low-quality (LQ) arm (Reid et al. 2016). The outcome of this experiment was to demonstrate the Physarum decision-making abilities. Physarum always chose the high-quality arm, and it can make multi-objective foraging decisions. It compares the relative qualities of multiple options and combines the information on reward (frequency and magnitude) in order to make correct and adaptive decisions. This experiment provides insight into the fundamental principles of Physarum decision-making and information processing.
7.7 Physarum foraging behaviour in competition settings experiment
In a biological experiment by Schumann et al. (2015), two strains were cultured in the same petri dish, the first was the usual Physarum Polycephalum plasmodium, and the second was another species called a Badhamiautricularis. Physarum Polycephalum definitely grows faster than Badhamiautricularis and overtakes more food resources, and could even grow into the branches of Badhamiautricularis, only if the Physarum inoculum was fatter (Schumann et al. 2015). Furthermore, if the invasive growth in front of Badhamiautricularis is well nourished by oat, it would easily overgrow the opposing tube system of Physarum Polycephalum. Thus, competitions between Physarum Polycephalum and Badhamiautricularis is based on Physarum power (type), mass, and the availability of nearby food resources.
A recent study by Masui et al. (2018), five geographical strains of Physarum with different genotypes were collected. In each experiment, two individual plasmodia on oat flakes were placed on \(2\%\) agar in a round petri dish and were allowed to behave freely. Whether the individuals avoided or fused was recorded for all encounter cases. Allorecognition was defined as the time when the plasmodium came into contact with the other individual. Completion of allorecognition was defined as a change in behaviour (continuing straight, changing direction, or starting to fuse at the point of contact). The study has revealed that Physarum strictly identifies allogeneic individuals when encountering different individuals. The Allorecognition system in Physarum prioritises the avoidance and severely restricts fusion when encountering different individuals (Masui et al. 2018).
Another biological experiment by Stirrup and Lusseau (2019), they have designed a biological experiment to study the behaviour of Physarum under competition settings (Stirrup and Lusseau 2019). The experiment intercalated two Physarum in a common environment (petri dish) where there was only one food resource available. The experimental results showed that the time taken by Physarum to find food depends on their hunger motivation. However, the time taken for a Physarum to start looking for food depended on its motivation and the motivation of its competitor. Physarum always initiates foraging behaviour quicker in the presence of competitors.
8 Mathematical models for simulating Physarum foraging behaviour
Physarum biological experiments are extremely slow and time-consuming to be applied in real-world network design problems. It is rather better to use the meta-heuristic algorithms inspired by Physarum intelligent behaviour (as conducted in real biological experiments) to construct mathematical models. However, rare work has been done to theoretically explain why these models work well experimentally. It is interesting to understand the theoretical analyses of Physarum models. Nevertheless, it is suggested that the protoplasmic flow through tubular veins plays an important role in explaining the observed Physarum’s intelligence. The tubes that transport the nutrients will grow bigger while the tubes which do not transport enough nutrients will vanish and disappear (Nakagaki et al. 2001; Tero et al. 2005; Werner 2019). The simulation and reproduction of this protoplasmic flow is the basis of many successful Physarum-inspired networking models (Sun 2017). For more details revise Section 3.
The existing models are simulating the intelligent behaviour of single Physarum, and have overlooked foraging behaviour of multiple Physarum under competitive settings. For this reason, we have presented our new model to simulate Physarum in competition, where multiple Physarum interact with each other and with their environments. We have summarised some of these existing mathematical models in Table 2.
8.1 The flow-conductivity model
The flow conductivity model is the most successful model that has been proposed to simulate the protoplasmic flow through Physarum’s tubular veins This model was first proposed by Tero et al. (2006) to solve the shortest path problem (Tero et al. 2006). Experiments on Physarum led by Nakagaki and Guy (2008) have proposed the mechanism of protoplasmic flow through Physarum’s tubular veins, which is believed to account for Physarum’s intelligence (Nakagaki and Guy 2008). The model illustrates the feedback between the flux and the thick of protoplasmic tubes; first, open-ended tubes, which are not connected between the two food sources, are likely to disappear. Second, when two or more tubes connect the same two food sources, the longer tube is likely to disappear. The flow-conductivity model is based on Hagen-Poiseuille Law and Kirchhoff Law to describe the adaptive feature of path finding and the feedback between flux and conductivity of the protoplasm tubes (Tero et al. 2007; Nakagaki and Guy 2008; Tero et al. 2008).
In this model, two terminals are representing Physarum (source/node), and the other terminal is food resource (sink/node). The protoplasm flows in every edge from the source node to the sink node. There is a pressure at each vertex, and the quantity of flux in each edge is proportional to the pressure difference between the two ends of these edges. Specifically, the flux \(Q_{ij}\) in edge (i, j) is given by the Hagen-Poiseuille equation below.
where \(D_{ij}\) is the edge conductivity, \(c_{ij}\) is the edge length, \(p_i\) and \(p_j\) are pressures at vertices i and j, \(r_{ij}\) is the edge radius, and \(\xi\) is the viscosity coefficient.
Physarum solver based on the flow-conductivity model (Tero et al. 2006) is probably the most famous one. Many of the Physarum-inspired algorithms suffer from a low convergence speed. Recent researches have proposed methods to accelerate the original path-finding model. Zhang et al. (2014b) accelerated the search by reducing the number of iterations by combining the original Physarum-solver model with a new parameter, called energy Zhang et al. (2014b). Huang et al. (2021) innovatively proposed the concept of the dominant path that could distinguish the shortest path from other paths, and the exact transition point when the shortest path can be identified (Huang et al. 2021). These two novel termination criteria led to a fast algorithm outperforming other baseline algorithms. Still more theoretical work is needed to improve the performance of these models and to prove its ability in solving real world applications.
8.2 Reaction-diffusion model
Adamatzky (2009a, 2007) regards the Physarum as an encapsulated reaction-diffusion computer, and utilises a two-variable Oregonator equation to simulate the Physarum spanning tree construction (Adamatzky 2009a, 2007). In this model, the wavefront is used to simulate the motion of Physarum, whose trajectory is steered by the gradient of chemo-attractants. It was treated as a bio-realised unconventional computer called "Physarum Machine" to solve maze problems, graph problems and design logical gates (Adamatzky 2010).
8.3 The cellular model
The cellular model was proposed by Gunji et al. (2008). Given a planar lattice, and every lattice site has various states: the inside (state 1) is surrounded by a boundary (state 2) in a lattice outside (state 0). In the foraging phase, there is cell invasion of the outside with softening of the membrane of Physarum. The protoplasmic flow toward the softened area, which leads to a re-organisation of the distribution of the cytoskeleton. This model was applied to simulate the amoebic motion and solve the classical Steiner tree problem in planes (Gunji et al. 2008, 2011). Moreover, other researchers have developed a cellular automata model based on reaction diffusion to simulate the behaviour of Physarum (Tsompanas and Sirakoulis 2012; Tsompanas et al. 2016; Liu et al. 2013).
8.4 The multi-agent model
Jones (2009) has been proposed a multi-agent, where Physarum is thought to consist of a population of particle-like agents (Jones 2009). Each agent senses and deposits trails as it moves towards the nearby stimulus within a 2D diffusive lattice. In this model, the structure of the Physarum network is indicated by the collective pattern of the positions of agents, and the protoplasmic flow is represented by the collective movement of agents. Furthermore, Wu et al. (2012) improved the initial multi-agent model by adding a memory module to each agent (Wu et al. 2012). This improved model is more flexible and adaptive, and it approximates the behaviours of Physarum more closely.
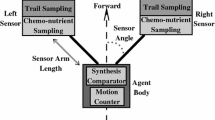
Liu et al. (2017b) proposed a self-organised system modelling approach in which two types of agents are used for simulating both the search (exploration) and the contraction (exploitation) of Physarum in foraging behaviours (Liu et al. 2017b). In this model, the body comprises a synthesis module and a motion module, and each sensor is armed with a trail sampling module and a chemo-nutrient sampling module.
8.5 Physarum competition model
Awad et al. (2019b) proposed a novel model to imitate the complex patterns observed in Physarum polycephalum generated in competition settings (Awad et al. 2019b). This new model is based on hexagonal Cellular Automata (CA) and Reaction-Diffusion (RD) systems. This is the first time Physarum has been simulated in a 2-D hexagonal grid that is more applicable to Physarum natural diffusion in a circular pattern to equidistant cells (Figs. 1, 2). All other models considered either Von-Neumann (4 adjacent neighbours) (Shirakawa et al. 2015) or Moore neighbourhoods (8 adjacent neighbours) (Tsompanas et al. 2016). However, in Von-Neumann model, diagonal diffusion of Physarum can still occur, while in Moore model the neighbourhoods are not equidistant. In this competition model multiple Physarum interact with each other and with their environment, each Physarum has its autonomous behaviour: it compares information on reward determined by food resources’mass and quality, negative effects of competing neighbours according to their mass, and hunger motivation in order to make correct and adaptive decisions. They believe that competition among different Physarum individuals can lead to the emergence of a complex global behaviour, far beyond the capability of individual Physarum. The individual skills of competition are more efficient to achieve an optimal balance between exploration and exploitation and maintain population diversity.

The structure of the cellular automaton and how diffusion process is executed over time

An example of a grid of four Physarum competing for two food resources, some obstacles and empty cells
9 Physarum-inspired applications
In this section, we will address the most important question "What Physarum can offer to computing?". Many Physarum-inspired algorithms have been developed and proved to have great potential to solve various optimisation problems using simple heuristics. In this context we will not be restricted to graph optimisation problems as previous reviews (Zhang et al. 2016b; Sun 2017; Adamatzky 2010; Gao et al. 2019), we will open the horizon and through light to more recent applications. We will address this issue by briefly reviewing some of the existing researches on these Physarum-inspired applications.
9.1 Physarum-inspired algorithms for graph-optimisation problems
Physarum protoplasmic flux is changing continuously with the change of environment in its foraging process. This characteristic allows Physarum to have great potentials in dealing with graph-optimisation problems which are considered the main application. The Physarum solver constructs networks by making some nodes in the network "sources" and cytoplasmic streaming as edges to others "sinks". Physarum network design has attracted the attention of many researchers as it demonstrated excellent performance in network construction without central consciousness during the process of foraging. For a detailed discussion on the existing methods and applications refer to (Zhang et al. 2016b; Sun 2017).
As the previous work by Sun (2017) has indicated, these network graph-optimisation problems are typically based on the following four strategies:
-
One source node and one sink node: It was first proposed by Nakagaki et al. (2000) after performing his famous experiments showing that Physarum was able to find the shortest route through a maze (Nakagaki et al. 2000). Qian et al. (2013) solved the travelling salesman problem Qian et al. (2013). Zhang et al. (2016b) accelerated its optimisation process by intentionally removing the edges with a stable decreasing flow Zhang et al. (2016b).
-
Multiple source nodes and one sink node: this strategy is to select one terminal to be the sink node and then select the other terminals to be source nodes. It has been applied by Liu et al. (2015b) to solve the classical Steiner tree problem in graphs (Liu et al. 2015b). It has also been used to solve the prize-collecting Steiner tree problem and the node-weighted Steiner tree problem (Sun and Halgamuge 2016).
-
One source node and multiple sink nodes: this strategy is to select one terminal to be the source node and then select the other terminals to be sink nodes. It was first used by Watanabe and Takamatsu (2014) to design transportation networks with fluctuating traffic distributions (Watanabe and Takamatsu 2014).
-
Multiple source nodes and multiple sink nodes: this strategy is to select multiple terminals to be the source nodes and multiple terminals to be the sink nodes. It was recently proposed by Zhang et al. (2016b) to solve the supply chain network design problem (Zhang et al. 2016b).
The Physarum-inspired Graph-Optimisation applications can be categorised as follows:
9.1.1 Finding the shortest path in directed and undirected networks
Nakagaki et al. (2000) were the first to show how this simple organism has the ability to find the shortest path between two points in a labyrinth (Nakagaki et al. 2000). Subsequent research has confirmed and broadened the range of its computation abilities to spatial representations of various graph problems (Zhang et al. 2016b). It showed that the Physarum’s network geometry met the requirements of a smart network: short tubes, close connections among all the branches, and tolerance to dynamic changes. A lot of Physarum-inspired algorithms (PAs) have been proposed to solve challenging network optimisation problems, such as the travelling salesman problem (Masi and Vasile 2014), spanning tree approximation (Adamatzky 2009b), vehicle routing problems Liu et al. (2016), designing supply chain networks (Zhang et al. 2016a), and community detection (Gao et al. 2018).
9.1.2 Designing transport networks
Adamatzky (2012a) presented a bioevaluation of world transport networks (Adamatzky 2012a). This work showed that man-made transport networks and their design are closely related to the shortest path problem. Tero et al. (2010) carried their designed well-known real experiment to design a Physarum bio-inspired networks similar to the Tokyo rail system (Tero et al. 2010). The resulting networks are both efficient and robust. Some other real-world transportation networks have also been approximated by Physarum since then, such as Mexican highway (Adamatzky et al. 2011), Iberian highway (Adamatzky and Alonso-Sanz 2011), Route 20 in USA Adamatzky (2014) and Autobahn 7 in Germany (Adamatzky 2014). Tsompanas et al. (2015) presented a cellular automaton-based, Physarum-inspired, transport network designing model (Tsompanas et al. 2015).
9.1.3 Solving steiner tree problem and applications
The classical geometric Steiner tree problem is to find the shortest network to connect multiple terminals. Several algorithms have been proposed to solve this problem such as flow-conductivity based model (Tero et al. 2006) and cellular model (Gunji et al. 2008). Inspired by the path-finding and network formation capability of Physarum, Liu et al. (2015b) explored a cellular computing model to solve the Steiner tree problem, especially in network design (Liu et al. 2015b).
In the prize-collecting Steiner tree problem, we not only have edge costs, but also have none-negative node weights. Recently, Sun et al. (2016) proposed a flow-conductivity model to solve this problem for drug re-positioning (Sun et al. 2016). The node-weighted Steiner tree problem is a more general version of the prize-collecting Steiner tree problem, and the node weight can be positive, negative, or zero. It has been applied to various areas, such as communication network design (Sun and Halgamuge 2016) and biomedical data analytics (Sun et al. 2017). These Physarum algorithms compute Steiner trees by iteratively calculating pressures, updating edge conductivities, and cutting edges.
Recently Sun et al. (2020) have conducted theoretical analyses on some Physarum-inspired algorithms to reveal their potential of computing Steiner trees (Sun et al. 2020). It was concluded that Physarum-inspired algorithms can find low-cost networks to connect multiple compulsory vertices together, and have the potential to compute Steiner trees. However, it is observed that using such iterations are computationally too expensive for large graphs. To overcome this point they have proposed a hybrid algorithm by incorporating Steiner tree techniques into the Physarum-inspired optimisation process. They formulated the Node-Weighted Partial Terminal Steiner Tree Problem for minimum-cost relay node placement in two-tiered wireless sensor networks. The objective is to minimise the sum of heterogeneous production and placement costs of relay nodes and the sum of outage probabilities of transmission routes in a routing tree simultaneously.
9.2 Evolutionary algorithm optimisation (Hybrid Models)
Prior knowledge plays a vital role in the computational efficiency of evolutionary algorithms (e.g., Genetic Algorithm, and Ant Colony). Taking advantage of Physarum powerful computational capabilities, such as morphological diversity (Gunji et al. 2008) and positive feedback loop (Tero et al. 2005), these characteristics have been used to optimise some evolutionary algorithms to improve its efficiency and robustness (Zhang et al. 2014a; Gao et al. 2018).
Ant colony optimisation (ACO) algorithms have been shown to provide an approximate solution for NP-hard problems existing in many real-world applications. However, premature convergence has significantly reduced the performance of these algorithms. Zhang et al. (2014a) proposed an optimisation strategy for updating the pheromone matrix in ant colony algorithms based on a Physarum mathematical model (Zhang et al. 2014a). This strategy has accelerated the positive feedback process in ACO, for solving NP-hard problems such as travelling salesman problem (TSP) and 0/1 knapsack problem, which contributed to the quick convergence of the optimal solution (Liu et al. 2017a). Later on Gao et al. (2018) has incorporated Physarum-inspired initialisation to optimise the genetic algorithm, ant colony optimisation algorithm and Markov clustering algorithm for solving community detection problems (Gao et al. 2018).
9.3 Biological computing and physarum logic gates
Boolean logic which describes binary arithmetic is fundamental to computer science as electronic logic gates form the basis of digital operations in computers. Organism based Bio-Logic gates have been attempted using cell constituent (bacteria) as transducers (Wang et al. 2005). Bacteria have many drawbacks, mainly due to the fragility, short life, limited temperature, and pH conditions. Also, bacteria will often not grow on specific substrates which would be ideal for the cell-transducer interface. Yeast and wild fungi are offering the advantage of high growth rate and the ability to grow on a broad range of surface substrates used for cell-transducer interface (Baronian 2004). Moreover, yeast can survive for over a long time after dehydration and could be re-hydrated when required.
Like other fungi and yeast, Physarum is accessible to culture on moist filter paper or agar and resist dehydration for a long time. This is why it can be considered as a prospective experimental prototype of biological computers which does not require sophisticated support. In standard electric devices, we deal with electrical signals to code information. However, in a Physarum biological device instead of electrical signals, the calculation process is performed by using the Physarum chemotaxis to food (Adamatzky 2010; Umedachi et al. 2011).
Physarum as a method of biological computing has been extensively studied in the PhyChip project that ran between 2013 and 2016 "Physarum chip: growing computers from slime mould" (Adamatzky et al. 2012). A Physarum chip is formed of a living network of protoplasmic tubes that acts as an active non-linear transducer of information, while templates of tubes coated with conductor act as fast information channels. The symbolic-logical, mathematical and programming aspects of the Physarum chip have been studied by Schumann et al. (2015) Schumann et al. (2015). Schumann (2019) studied the logics of swarm behaviours, where each unicellular organism can be regarded as a logic gate, and all behaviours can be controlled by stimuli in the environment: attractants (motivational reinforcement) and repellents (motivational punishment). The reactions to stimuli can be regarded as: sensing stage (perceiving input signals) and motoring (appropriate direct reactions output signals) Schumann (2019). Physarum was also used as a Boolean gate, where the presence and absence of Physarum in a given locus of space is equivalent to logic values 1 and 0, respectively (Schumann and Adamatzky 2011). The Physarum chip is expected to solve a wide range of computation tasks, including graph optimisation, logic and arithmetical computing (Andrew and Krzysztof 2016).
The EU-funded PhySense project "Physarum Sensor: Biosensor for Citizen Scientists" is an extension of the PhyChip project. This project showed that Physarum is an ideal biological substrate that could be used as a biosensor that converts a biological response into an electrical signal, providing a unique fusion of living and digital technology. The PhySense software calculates any changes in the frequency and amplitude of oscillations in the tubular structures of Physarum. The aim of this project is developing marketable low-cost biosensors for various applications, including environmental monitoring and health (CORDIS 2019)Footnote 1.
9.4 Physarum competition algorithm applications
The Physarum competition model (Awad et al. 2019b) has a great potentials to be customised to formulate new algorithms to solve the real-world problems that have heuristic similarity to Physarum foraging behaviour in competition settings. Thus we are inspiring from biology to advance computational solution. Graph-optimisation problems in a dynamic environment as in Mobile Wireless Sensor Networks, and Discrete Multi-Objective Optimisation problems are examples of these applications.
9.4.1 Physarum competition algorithm for wireless sensor networks
The popular Physarum-inspired Algorithms (PAs) have proven its potential in solving challenging network optimisation problems (Reid and Beekman 2013; Zhang et al. 2016b). However, some network optimisation problems remain unsolved. New techniques are required to address the large scale of the next-generation networks, where centralised control of communication becomes impractical. Physarum distributed intelligence may inform the design of an adaptive, robust and spatial infrastructure networks with decentralised control systems (Sun 2017). We have proposed a Physarum competition model (Awad et al. 2019b), where multiple Physarum with autonomous behaviours react to each other and with their environment without central control to achieve efficient exploration of the whole system evolving to an optimal global network, this has allowed us to deal with the increasingly proposed networks scenarios with multiple sources and multiple sinks. In our previous work, we have presented a Physarum-inspired competition algorithms for mobile wireless sensor networks, where multiple Physarum (as represented by sensors) will sense the surrounding environment, and compete over multiple food resources (as represented by interest points). These algorithms have demonstrated their promising performance in solving node deployment (Awad et al. 2018) and connectivity restoration even in harsh environment (Awad et al. 2019a).
9.4.2 Physarum competition Algorithm (PCA) for discrete multiobjective optimisation problems
Many real-world problems can be naturally formulated as discrete multi-objective optimisation problems. Physarum competition algorithm has been proposed to solve this problem (Awad et al. 2019c). This algorithm is based on Physarum motility over hexagonal cellular automaton (problem search space). Each cell is considered a food resource (objective functions) to attract Physarum. The PCA has achieved similar performance results compared to the state-of-the-art multi-objective evolutionary algorithms and even outperformed other well-established algorithms.
10 Conclusion
By studying Physarum foraging behaviour and translating that behaviour into mathematical models, we increase our understanding of how to inspire from biology to develop Physarum bio-inspired algorithms can solve many challenging real world problems. Physarum polycephalum is an example of plasmodial slime moulds. The primitive intelligence of Physarum polycephalum is mostly demonstrated during its plasmodium stage (a large multi-nucleated single cell) that consists of a single cell amoeba-like organism. Physarum senses gradients of chemo-attractants and repellents and forms a yellowish vascular network in search of nutrition. A stimulus triggers the release of a signalling molecule cyclic adenosine monophosphate (cAMP) which starts cytoplasmic streaming. This generates a feedback loop; the higher the rate of cytoplasmic streaming is, the thicker the vein becomes. The Physarum foraging behaviour consists of two simultaneous self-organised processes of expansion (exploration) and shrinkage (exploitation). Just like social insects and animals, Physarum too exhibits swarm intelligence; it shares many features of collective behaviour such as synchronisation, communication, positive feedback, leadership, and response thresholds. There is increasing evidence that a simple organism like Physarum has complex social behaviours including cooperation and competition. Physarum is capable of making complex foraging decisions based on trade-offs between risks, hunger level and food patch quality. The skills of individual competitors are effective methods for inspiration to develop intelligent systems and to provide solutions for decision-making problems.
Physarum may not have brains, but the advantages of Physarum unconventional computational capabilities, as morphological diversity and positive feedback loop, have great potentials for solving many real-world applications. Physarum, as a simple organism, has the ability to find the minimum-length between two points in solving the maze problem and discover the shortest path in real-world networks such as the Tokyo railway network using simple heuristics. Much research has confirmed and broadened the range of its computation abilities to spatial representations of various graph optimisation problems. Physarum-inspired initialisation of other bio-inspired techniques has the ability to accelerate convergence and improve the searching capability of evolutionary algorithms (e.g., Genetic Algorithm, and Ant Colony) in terms of accuracy and computational cost. Physarum as a biological model has been studied in the PhyChip and PhySense projects to develop marketable and low-cost biosensors for various applications including environmental monitoring and health.Physarum can be considered one of the biological models of unconventional computation capable of making a programmable Physarum machine.
Notes
More information: PhySense project website: www.physense.eu/.
References
Adamatzky A (2007) Physarum machines: encapsulating reaction-diffusion to compute spanning tree. Naturwissenschaften 94(12):975–980
Adamatzky A (2009) From reaction-diffusion to physarum computing. Nat. Comput 8(3):431–447
Adamatzky A (2009) If BZ medium did spanning trees these would be the same trees as Physarum built. Phys Lett, Sect A: General, At Solid State Phys 373(10):952–956
Andrew A (2009) Steering plasmodium with light: Dynamical programming of Physarum machine. arXiv preprint ( 2009). arXiv: 0908.0850
Adamatzky A (2010) Physarum machines: computers from slime mould. World Sci. https://doi.org/10.1142/7968
Adamatzky A (2012) Bioevaluation of world transport. Networks. https://doi.org/10.1142/8482
Adamatzky A (2012) Slime mold solves maze in one pass, assisted by gradient of chemo-attractants. IEEE Trans Nanobioscience 11(2):131–134. https://doi.org/10.1109/TNB.2011.2181978
Adamatzky A, Alonso-Sanz R (2011) Rebuilding Iberian motorways with slime mould. Biosystems 105(1):89–100. https://doi.org/10.1016/j.biosystems.2011.03.007
Adamatzky A, Erokhin V, Grube M, Schubert T, Schumann A (2012) Physarum chip project: growing computers from slime mould. IJUC 8(4):319–323
Andrew A, Genaro JM, Sergio VC-V, Rene A-P, Stephens CR (2011) Approximating Mexican highways with slime mould. Nat Comput. https://doi.org/10.1007/s11047-011-9255-z
Adamatzky AI (2014) Route 20, autobahn 7, and slime mold: approximating the longest roads in usa and germany with slime mold on 3-d terrains. IEEE Trans Cybernet 44(1):126–136. https://doi.org/10.1109/TCYB.2013.2248359
Alim K, Andrew N, Pringle A, Brenner MP (2017) Mechanism of signal propagation in Physarum polycephalum. Proceed Natl Acad Sci United States of Am 114(20):5136–5141. https://doi.org/10.1073/pnas.1618114114
Schumann A, Pancerz K (2016) Physarum Chip: Growing Computers from Slime Mould. Logical Aspects. University of Information Technology and Management in Rzeszow St. Sucharskiego 2, 35-225 Rzeszów, Poland Scientific Publishing House IVG St. Cyfrowa 6, 71-441 Szczecin, Poland www.wydawnictwoivg.pl email: biuro@wydawnictwoivg.pl, Rzeszów-Szczecin
Awad A, Pang W, Coghill G (2018) Physarum inspired model for mobile sensor nodes deployment in the presence of obstacles. Lect Notes Inst Comput Sci, Soc-Inform Telecommun Eng, LNICST. https://doi.org/10.1007/978-3-319-95450-9_12
Awad A, Pang W, Coghill GM (2019) Physarum inspired connectivity and restoration for wireless sensor and actor networks. Adv Intell Syst Comput 840:327–338. https://doi.org/10.1007/978-3-319-97982-3_27
Abubakr A, Wei P, David L, Coghill GM (2019) A hexagonal cell automaton model to imitate physarum polycephalum competitive behaviour. In: The 2018 Conference on Artificial Life: a Hybrid of the European Conference on Artificial Life (ECAL) and the International Conference on the Synthesis and Simulation of Living Systems (ALIFE)
Abubakr A, Muhammad U, David L, George MC, Wei P (2019) . A physarum-inspired competition algorithm for solving discrete multi-objective optimization problems. In: Genetic and Evolutionary Computation Conference Companion (GECCO’19 Companion) (GECCO’19). ACM, New York, NY, USA. https://doi.org/10.1145/3319619.3322030
Baronian KHR (2004) The use of yeast and moulds as sensing elements in biosensors. Biosens Bioelectron 19(9):953–962. https://doi.org/10.1016/j.bios.2003.09.010
Beekman M, Latty T (2015) Brainless but multi-headed: decision making by the acellular slime mould physarum polycephalum. J Molecular Biol 427(23):3734–3743. https://doi.org/10.1016/j.jmb.2015.07.007
Bonifaci V, Mehlhorn K, Varma G (2012) Physarum can compute shortest paths. J Theor Biol 309(2012):121–133
Boussard A, Delescluse J, Pérez-Escudero A, Dussutour A (2019) Memory inception and preservation in slime moulds: the quest for a common mechanism. Philosophical Trans Royal Soc B 374:1774. https://doi.org/10.1098/rstb.2018.0368
Cavender J (1995) Myxomycetes: a handbook of slime molds. Bioscience 45(11):795–797. https://doi.org/10.2307/1312636
CORDIS. (2019) How to develop affordable sensors using slime mold. https://phys.org/news/2019-06-sensors-slime-mold.html
Cortés Pablo, García José M, Muñuzuri Jesús, Onieva Luis (2008) Viral systems: a new bio-inspired optimisation approach. Comput Op Res 35(9):2840–2860. https://doi.org/10.1016/j.cor.2006.12.018 (Part Special Issue: Bio-inspired Methods in Combinatorial Optimization)
Dorigo M, Maniezzo V, Colorni A (1996) Ant system: optimization by a colony of cooperating agents. IEEE Transactions on Systems, Man, and Cybernetics, Part B (Cybernetics) 26, 1, 29–41. https://doi.org/10.1109/3477.484436
Dirk D, Bakhtier V, Weijer CJ (2000) The control of chemotactic cell movement during Dictyostelium morphogenesis. Philosophical Transactions of the Royal Society of London.Series B: Biological Sciences 355: 1399, 983–991 https://doi.org/10.1098/rstb.2000.0634
Durham ACH, Ridgway EB (1976) Control of chemotaxis in Physarum Polycephalum. J Cell Biol 69(1):218–223. https://doi.org/10.1083/jcb.69.1.218
Dussutour A, Latty T, Beekman M, Simpson SJ (2010) Amoeboid organism solves complex nutritional challenges. Proceed Natl Acad Sci United States of Am 107(10):4607–4611. https://doi.org/10.1073/pnas.0912198107
Eberhart R, Kennedy J (1995). A new optimizer using particle swarm theory. In: MHS’95. Proceedings of the Sixth International Symposium on Micro Machine and Human Science. https://doi.org/10.1109/MHS.1995.494215
Victor E (2014) . Organic Memristive Devices and Neuromorphic Circuits. Springer, pp. 389–411
Fischer Justina A.V. (2008) Is competition good for trust? Cross-country Evidence Using Micro-data. Econom Lett 100(1):56–59. https://doi.org/10.1016/j.econlet.2007.11.001
Iosif-AF, Vasileios N, Georgios Ch S, Andrew A, Victor E, Antonio R (2019). Wave computing with passive memristive networks. In: 2019 IEEE International Symposium on Circuits and Systems (ISCAS). IEEE, 1–5
Gale E, Adamatzky A, de Lacy Costello B (2014) Slime mould memristors. BioNanoScience. https://doi.org/10.1007/s12668-014-0156-3
Gao C, Liang M, Li X, Zhang Z, Wang Z, Zhou Z (2018) Network community detection based on the Physarum-inspired computational framework. IEEE/ACM Trans Comput Biol Bioinform 15(6):1916–1928. https://doi.org/10.1109/TCBB.2016.2638824
Gao C, Liu C, Schenz D, Li X, Zhang Z, Jusup M, Wang Z, Beekman M, Nakagaki T (2019) Does being multi-headed make you better at solving problems? A survey of Physarum-based models and computations. Phys Life Rev 29(2019):1–26. https://doi.org/10.1016/j.plrev.2018.05.002
Gunji YP, Shirakawa T, Niizato T, Haruna T (2008) Minimal model of a cell connecting amoebic motion and adaptive transport networks. J Theor Biol 253(4):659–667. https://doi.org/10.1016/j.jtbi.2008.04.017
Gunji YP, Shirakawa T, Niizato T, Yamachiyo M, Tani I (2011) An adaptive and robust biological network based on the vacant-particle transportation model. J Theor Biol 272(1):187–200. https://doi.org/10.1016/j.jtbi.2010.12.013
Holland John H (1973) Genetic algorithms and the optimal allocation of trials. SIAM J Comput 2(2):88–105. https://doi.org/10.1137/0202009
Yusheng H, Dong C, Joel Weijia L, Yong D, Kang HC (2021). When does the Physarum solver distinguish the shortest Path from other Paths: the transition point and its applications. arXiv preprint arXiv: 2101.02913 ( 2021)
Ito K, Sumpter D, Nakagaki T (2010) Risk management in spatio-temporally varying field by true slime mold. Nonlinear Theory Its Appl IEICE 1(1):26–36. https://doi.org/10.1587/nolta.1.26
Jeff J (2009) Approximating the behaviours of Physarum polycephalum for the construction and minimisation of synthetic transport networks. In: International Conference on Unconventional Computation. Springer Berlin Heidelberg, pp. 191–208. https://doi.org/10.1007/978-3-642-03745-0_23
Jones J (2011) Influences on the formation and evolution of Physarum polycephalum inspired emergent transport networks. Nat Comput 10(4):1345–1369. https://doi.org/10.1007/s11047-010-9223-z
Jones J, Adamatzky A (2014) Material approximation of data smoothing and spline curves inspired by slime mould. Bioins Biomimet. https://doi.org/10.1088/1748-3182/9/3/036016
Jones Jeff, Mayne Richard, Adamatzky Andrew (2017) Representation of shape mediated by environmental stimuli in Physarum polycephalum and a multi-agent model. Int J Parallel, Emerg Distribut Syst 32(2):166–184. https://doi.org/10.1080/17445760.2015.1044005
Karaboga D, Basturk B (2007) A powerful and efficient algorithm for numerical function optimization: artificial bee colony (ABC) algorithm. J Glob Optim 39(3):459–471. https://doi.org/10.1007/s10898-007-9149-x
Kim DH, Abraham A, Cho JH (2007) A hybrid genetic algorithm and bacterial foraging approach for global optimization. Inform Sci 177(18):3918–3937. https://doi.org/10.1016/j.ins.2007.04.002
Latty T, Beekman M (2010) Food quality and the risk of light exposure affect patch-choice decisions in the slime mold Physarum polycephalum. Ecology 91(1):22–27. https://doi.org/10.1890/09-0358.1
Latty T, Beekman M (2011) Speed-accuracy trade-offs during foraging decisions in the acellular slime mould Physarum polycephalum. Proceed Royal Soc B: Biol Sci 278(1705):539–545. https://doi.org/10.1098/rspb.2010.1624
Liu L, Song Y, Zhang H, Ma H, Vasilakos AV (2015) Physarum optimization: a biology-inspired algorithm for the steiner tree problem in networks. IEEE Trans Comput 64(3):818–831. https://doi.org/10.1109/TC.2013.229
Liu Y, Gao C, Liang M, Tao L, Zhang Z (2015) A physarum-inspired vacant-particle model with shrinkage for transport network design. Lecture Notes Comput Sci (including subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics) 9140:74–81. https://doi.org/10.1007/978-3-319-20466-6_8
Liu Y, Gao C, Zhang Z, Lu Y, Chen S, Liang M, Tao L (2017) Solving NP-Hard problems with Physarum-based Ant colony system. IEEE/ACM Trans Comput Biol Bioinform 14(1):108–120. https://doi.org/10.1109/TCBB.2015.2462349
Liu Y, Gao C, Zhang Z, Wu Y, Liang M, Tao L, Lu Y (2017) A new multi-agent system to simulate the foraging behaviors of Physarum. Nat Comput 16(1):15–29. https://doi.org/10.1007/s11047-015-9530-5
Liu Y, Khalifa I, El-Kamel A (2016). The multi-period and multi-depot dynamic vehicle routing problem with time windows. In: 2016 3rd International Conference on Logistics Operations Management (GOL). IEEE, pp. 1–6. https://doi.org/10.1109/GOL.2016.7731713
Liu Y,Zhang Z, Gao C, Wu Y, Qian T (2013) . A Physarum network evolution model based on IBTM. In; International Conference in Swarm Intelligence. Springer, pp. 19–26. https://doi.org/10.1007/978-3-642-38715-9_3
Masi L, Vasile M (2014) A multi-directional modified physarum algorithm for optimal multi-objective discrete decision making. Stud Comput Intell 500:195–212. https://doi.org/10.1007/978-3-319-01460-9_9
Masui Mana, Satoh Shinobu, Seto Kensuke (2018) Allorecognition behavior of slime mold plasmodium-Physarum rigidum slime sheath-mediated self-extension model. J Phys D: Appl Phys 51(28):284001. https://doi.org/10.1088/1361-6463/aac985
Mohseni S, Gholami R, Zarei N, Zadeh AR (2014) Competition over resources: a new optimization algorithm based on animals behavioral ecology. In: Proceedings - 2014 International Conference on Intelligent Networking and Collaborative Systems, IEEE INCoS 2014. pp. 311–315. https://doi.org/10.1109/INCoS.2014.55
Nakagaki T, Guy R (2008) Intelligent behaviors of amoeboid movement based on complex dynamics of soft matter. Soft Mat 4(1):57–67
Nakagaki T, Yamada H, Tóth Á (2000) Maze-solving by an amoeboid organism. Nature 407(6803):470. https://doi.org/10.1038/35035159
Nakagaki T, Yamada H, Tóth Á (2001) Path finding by tube morphogenesis in an amoeboid organism. Biophys Chem 92(1–2): 47–52. https://doi.org/10.1016/S0301-4622(01)00179-X
Vasileios N, Ioannis V, Georgios Ch S, Andrew A, Antonio R (2018) Coupled physarum-inspired memristor oscillators for neuron-like operations. In: 2018 IEEE International Symposium on Circuits and Systems (ISCAS)
Ntinas V, Vourkas I, Sirakoulis GC, Adamatzky AI (2017) Modeling Physarum space exploration using memristors. Appl Phys J Phys D. https://doi.org/10.1088/1361-6463/aa614d
Ntinas V, Vourkas I, Sirakoulis GC, Adamatzky AI (2017) Oscillation-based slime mould electronic circuit model for maze-solving computations. IEEE Trans Circuits and Syst I: Regular Papers 64(6):1552–1563. https://doi.org/10.1109/TCSI.2016.2566278
Oettmeier C, Brix K, Döbereiner H (2017) Physarum polycephalum-A new take on a classic model system. J Phys D: Appl Phys 50(41):413001
PhySense. [n. d.] . PhySense. https://www.physense.eu/ [Online; accessed November-2019]
Tao Q, Zili Z, Chao G, Yuheng W, Yuxin L (2013) An ant colony system based on the Physarum network. In: International Conference in Swarm Intelligence. Springer, pp. 297–305. https://doi.org/10.1007/978-3-642-38703-6_35
Reid CR, Beekman M (2013) Solving the Towers of Hanoi - How an amoeboid organism efficiently constructs transport networks. J Exp Biol 216(9):1546–1551. https://doi.org/10.1242/jeb.081158
Reid CR, Beekman M, Latty T, Dussutour A (2013) Amoeboid organism uses extracellular secretions to make smart foraging decisions. Behav Ecol 24(4):812–818. https://doi.org/10.1093/beheco/art032
Reid CR, Latty T (2016) Collective behaviour and swarm intelligence in slime moulds. FEMS Microbiol Rev 40(6):798–806. https://doi.org/10.1093/femsre/fuw033
Reid CR, Latty T, Dussutour A, Beekman M (2012) Slime mold uses an externalized spatial memory to navigate in complex environments. Proceed Natl Acad Sci United States of Am 109(43):17490–17494. https://doi.org/10.1073/pnas.1215037109
Reid CR, MacDonald H, Mann RP, Marshall JA, Latty T, Garnier S (2016) Decision-making without a brain: how an amoeboid organism solves the two-armed bandit. J Royal Soc Interface 13:119. https://doi.org/10.1098/rsif.2016.0030
Saigusa T, Tero A, Nakagaki T, Kuramoto Y (2008) Amoebae anticipate periodic events. Phys Rev Lett 100:1. https://doi.org/10.1103/PhysRevLett.100.018101
Schumann A (2019) Behaviourism in studying swarms: logical models of sensing and motoring. Springer
Schumann A, Adamatzky A (2011) Physarum spatial logic. New Math Nat Comput 7(03):483–498. https://doi.org/10.1142/S1793005711002037
Schumann A, Pancerz K, Adamatzky A, Grube M (2015) Bio-inspired game theory: the case of Physarum polycephalum. ICST. https://doi.org/10.4108/icst.bict.2014.257869
Kazuo S, Yoshitsugu K, Yoshifumi Y (1998) . Analysis methods of phase propagation in autonomic oscillation of Physarum polycephalum. In Proceedings of the 20th Annual International Conference of the IEEE Engineering in Medicine and Biology Society. Vol. 20 Biomedical Engineering Towards the Year 2000 and Beyond (Cat. No. 98CH36286), 3. IEEE, pp. 1606–1609. https://doi.org/10.1109/IEMBS.1998.747208
Senturk I, Yilmaz S, Akkaya K (2012) Connectivity restoration in delay-tolerant sensor networks using game theory. Int J Ad Hoc and Ubiquitous Comput 11(2–3):109–124. https://doi.org/10.1504/IJAHUC.2012.050268
Shirakawa T, Gunji Y-P, Miyake Y (2011) An associative learning experiment using the plasmodium of Physarum polycephalum. Nano Commun Netw 2(2–3):99–105. https://doi.org/10.1016/j.nancom.2011.05.002
Shirakawa T, Sato H, Ishiguro S (2015) Construction of living cellular automata using the Physarum plasmodium. Int J Gen Syst 44(3):292–304. https://doi.org/10.1080/03081079.2014.997531
Eilidh S, David L (2019) Getting a head start: the slime mold, Physarum polycephalum, tune foraging decision to motivational asymmetry when faced with competition. arXiv e-prints (2019) arXiv:1905.06534
Yahui S (2017) . Physarum-inspired Network Optimization: A Review. CoRR abs/1712.02910 ( 2017). arXiv: 1712.02910
Yahui S, Saman H (2016) Fast algorithms inspired by physarum polycephalum for node weighted steiner tree problem with multiple terminals. In:2016 IEEE Congress on Evolutionary Computation (CEC). IEEE, 3254–3260. https://doi.org/10.1109/CEC.2016.7744201
Sun Yahui, Hameed Pathima Nusrath, Verspoor Karin, Halgamuge Saman (2016) A physarum-inspired prize-collecting steiner tree approach to identify subnetworks for drug repositioning. BMC Syst Biol 10(5):25–38
Sun Y, Ma C, Halgamuge S (2017) The node-weighted Steiner tree approach to identify elements of cancer-related signaling pathways. BMC Bioinform 18(16):53–65
Sun Y, Rehfeldt D, Brazil M, Thomas D, Halgamuge S (2020) A Physarum-inspired algorithm for minimum-cost relay node placement in wireless sensor networks. IEEE/ACM Trans Netw 28(2):681–694
Tan Y, Shi Y (2017) Editorial: special section on Bio-inspired swarm computing and engineering. IEEE/ACM Trans Comput Biol Bioinform 14(1):1–3. https://doi.org/10.1109/TCBB.2016.2566438
Tero A, Kobayashi R, Nakagaki T (2005) A coupled-oscillator model with a conservation law for the rhythmic amoeboid movements of plasmodial slime molds. Physica D: Nonlinear Phenomena 205(1–4):125–135. https://doi.org/10.1016/j.physd.2005.01.010
Tero A, Kobayashi R, Nakagaki T (2006) Physarum solver: a biologically inspired method of road-network navigation. Physica A: Statistical Mech its Appl 363(1):115–119. https://doi.org/10.1016/j.physa.2006.01.053
Tero Atsushi, Kobayashi Ryo, Nakagaki Toshiyuki (2007) A mathematical model for adaptive transport network in path finding by true slime mold. J Theor Biol 244(4):553–564. https://doi.org/10.1016/j.jtbi.2006.07.015
Tero A, Takagi S, Saigusa T, Ito K, Bebber DP, Fricker MD, Yumiki K, Kobayashi R, Nakagaki T (2010). Rules for biologically inspired adaptive network design. Science (New York, N.Y.)
Tero A, Yumiki K, Kobayashi R, Saigusa T, Nakagaki T (2008) Flow-network adaptation in Physarum amoebae. Theory Biosci 127(2):89–94. https://doi.org/10.1007/s12064-008-0037-9
Traversa FL, Pershin YV, Di Ventra M (2013) Memory models of adaptive behavior. IEEE Trans Neural Netw Learn Syst 24(9):1437–1448. https://doi.org/10.1109/TNNLS.2013.2261545
Tsompanas MAI, Sirakoulis GC (2012) Modeling and hardware implementation of an amoeba-like cellular automaton. Bioinspi Biomimet. https://doi.org/10.1088/1748-3182/7/3/036013
Tsompanas M-AI, Sirakoulis G Ch, Adamatzky A (2016) Cellular automata models simulating slime mould computing. Springer pp. 563–594. https://doi.org/10.1007/s11047-016-9559-0
Tsompanas MAI, Sirakoulis GC, Adamatzky AI (2015) Evolving transport networks with cellular automata models inspired by slime mould. IEEE Trans Cybernet 45(9):1887–1899. https://doi.org/10.1109/TCYB.2014.2361731
Tsuda S, Aono M, Gunji YP (2004) Robust and emergent Physarum logical-computing. BioSystems 73(1):45–55. https://doi.org/10.1016/j.biosystems.2003.08.001
Ueda T, Muratsugu M, Kurihara K, Kobatake Y (1976) Chemotaxis in Physarum polycephalum: Effects of chemicals on isometric tension of the plasmodial strand in relation to chemotactic movement. Exp Cell Res 100(2):337–344. https://doi.org/10.1016/0014-4827(76)90157-9
Ueda T, Muratsugu M, Kurihara K, Kobatake Y (1976) Chemotaxis in Physarum polycephalum. Effects of chemicals on isometric tension of the plasmodial strand in relation to chemotactic movement. Exp Cell Res 100(2):337–344. https://doi.org/10.1016/0014-4827(76)90157-9
Takuya U, Masakazu A, Atsushi T, Akio I (2011) Simulation of a soft-bodied fluid-driven amoeboid robot that exploits thixotropic flow. In; 2011 IEEE International Conference on Robotics and Automation. IEEE, pp. 5123–5128. https://doi.org/10.1109/ICRA.2011.5980150
Verhoef Herman A, Morin Peter J (2010) Community ecology: processes, models, and applications. Oxford Univ Press. https://doi.org/10.1093/acprof:oso/9780199228973.001.0001
Wang Ping, Gaixia Xu, Qin Lifeng, Ying Xu, Li Yan, Li Rong (2005) Cell-based biosensors and its application in biomedicine. Sens Actuators B: Chem 108(1–2):576–584. https://doi.org/10.1016/j.snb.2004.11.056
Shin W, Atsuko T (2014) Transportation network with fluctuating input/output designed by the bio-inspired Physarum algorithm. PloS one 9, 2. https://doi.org/10.1016/j.copbio.2014.08.00
Werner Liss C (2019) Disruptive material intelligence of Physarum: liquid architecture of a biological geometry computer. pp. 227–247. https://doi.org/10.14279/depositonce-8629
West Stuart A, Diggle Stephen P, Buckling Angus, Gardner Andy, Griffin Ashleigh S (2007) The social lives of microbes. Annu Rev Ecol Evol Syst 38(2007):53–77. https://doi.org/10.1146/annurev.ecolsys.38.091206.095740
Whiting JGH, De Lacy Costello BPJ, Adamatzky A (2014) Towards slime mould chemical sensor: mapping chemical inputs onto electrical potential dynamics of Physarum Polycephalum. Sens Actuators, B: Chem 191(2014):844–853. https://doi.org/10.1016/j.snb.2013.10.064
Ka-Chun W (2015) Evolutionary multimodal optimization: a short survey. arXiv preprint. arXiv: 1508.00457
Yuheng W, Zili Z, Yong D, Huan Z, Tao Q (2012) An enhanced multi-agent system with evolution mechanism to approximate Physarum transport networks. In: Australasian Joint Conference on Artificial Intelligence. Springer, pp. 27–38. https://doi.org/10.1007/978-3-642-35101-3_3
Zhang X, Adamatzky A, Yang XS, Yang H, Mahadevan S, Deng Y (2016) A Physarum-inspired approach to supply chain network design. Sci China Inform Sciences 59, 5 https://doi.org/10.1007/s11432-015-5417-4
Zhang X, Gao C, Deng Y, Zhang Z (2016) Slime mould inspired applications on Graph-optimization problems. Springer International Publishing, Cham pp. 519–562. https://doi.org/10.1007/978-3-319-26662-6_26
Zhang X, Wang Q, Adamatzky A, Chan FTS, Mahadevan S,Deng Y (2014) An improved Physarum polycephalum algorithm for the shortest path problem. Sci World J https://doi.org/10.1155/2014/487069
Zili Z, Chao G, Yuxin L, Tao Q (2014) A universal optimization strategy for ant colony optimization algorithms based on the Physarum-inspired mathematical model. Bioinspiration & biomimetics 9, 3. https://doi.org/10.1088/1748-3182/9/3/036006
Acknowledgements
Dr. Abubakr Awad research was supported by Elphinstone PhD Scholarship, University of Aberdeen. Dr. Wei Pang, Prof. David Lusseau, and Prof. George M. Coghill were supported by the Royal Society International Exchange program (Grant Ref IE160806).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Awad, A., Pang, W., Lusseau, D. et al. A survey on physarum polycephalum intelligent foraging behaviour and bio-inspired applications. Artif Intell Rev 56, 1–26 (2023). https://doi.org/10.1007/s10462-021-10112-1
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10462-021-10112-1