
Abstract
In agroforestry systems (AFS), where environmental conditions are highly variable at small spatial scales, the use of uniform genetic material of a single cultivar commonly grown in monoculture cropping might not be optimal. However, the use of composite cross populations (CCPs) that contain an inherent genetic variability might be a promising approach under the environmental variability created by trees in AFS. In this experimental trial, the performance of a CCP (‘CC-2 k’) of winter wheat was compared to a commercial variety (‘Wiwa’) in a split-plot design at two AFS (Feusisberg and Wollerau) in Central Switzerland. Yield of CC-2k (1.9 ± 0.7 Mg ha−1) was higher than yield of Wiwa (0.7 ± 0.4 Mg ha−1) in Wollerau, but yields did not differ between CCP and variety in Feusisberg (1.9 ± 0.7 Mg ha−1 and 2.0 ± 0.8 Mg ha−1, respectively). The interaction of site and variety was significant (p < 0.05). Wiwa had a higher protein, Fe and Ca content than CC-2k. Therefore, while the CC-2k outperformed Wiwa in terms of yield in one of the two AFS, Wiwa outperformed CC-2k in terms of quality. In this one-year field experiment, the composite cross population might have been better adapted to the heterogenous environment of agroforestry systems (found in one out of two sites) but failed to reach the high-quality product of modern cultivars. These initial results must be seen as first insights which need to be complemented by larger field experiments for generalisation. The findings of this study may be interpreted as an indication that further improvements in terms of quality might make CCPs a viable option for diversified agricultural systems with larger environmental heterogeneity than common monoculture cropping systems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the face of climate change and the ongoing biodiversity crisis – requiring agriculture to adapt to a shifting climate and to be more environmentally-friendly – the development of sustainable and resilient food production systems with adaptive cultivars is highly relevant to meet the Sustainable Development Goals (Bourke et al. 2021; Obrecht et al. 2021; Rist et al. 2020). Agroforestry – agriculture with trees – has gained renewed economic and environmental interest in the past decade as numerous scientists have suggested agroforestry to be a promising approach to regenerative agriculture, defined as farming practices which conserve and restore soils and biodiversity, potentially offering greater resilience to climate change stressors (e.g., Kohli et al. 2007; Jose 2009; Torralba et al. 2016). In addition to the inherent interspecific diversity in agroforestry, intraspecific diversity in form of genetic diversity in tree and crop varieties can further contribute to agrobiodiversity and thus potentially to food security (Lin 2011), health benefits (Fanzo et al. 2013), preservation of agricultural knowledge (Morton 2019) and protection of ecosystem services (Duru et al. 2015).
Since the “Green Revolution” in the 1950s, local crop varieties have been replaced by standardized, commercial varieties worldwide. For instance, 90% of 10,000 wheat varieties cultivated in China in 1940 were lost in 1970 (Rist et al. 2020). Thus, the question is: “How can plant breeders regain intraspecific crop diversity?” One approach are composite cross populations (CCPs). Composite crosses are populations obtained by the reciprocal inter-crossing of genotypes and mixing of the progeny (Suneson 1956; Knapp et al. 2020). A CCP is therefore a particularly plastic and adaptable culture where each plant constitutes a unique genotype. Hence, the use of CCPs might be particularly helpful in the development of novel agroforestry systems. Their dynamic gene pool provides the potential of adaption to locally heterogeneous environmental conditions found in agroforestry systems (AFS) (Smith et al. 2012b; Bourke et al. 2021). Genotypes which are better adapted to local conditions should have more progeny and thus, over time, increase in frequency (Döring et al. 2011).
The spatial heterogeneity in AFS is created by various tree-crop-interactions dependent on the proximity to a single tree or the tree row and the overall-design of the AFS. Aboveground interactions include microclimatic modification (shade, decrease in temperature and wind speed, increased relative humidity) and increased insect density and diversity; belowground interactions include competition for soil water and nutrients, niche differentiation for nutrient capture and sharing, hydraulic lift and allelopathy (Jose et al. 2004). Optimal crop growth in the understorey of an AFS might theoretically require different cultivars that vary in light, water and nutrient requirements, but precision farming at such a small scale is practically challenging. Hence, previous studies have suggested that CCPs can overcome this variability and thereby provide similar yields and quality as pure varieties with increased yield stability (Döring et al. 2015; Vollenweider et al. 2020). The latter is desirable because although there is good scientific evidence that AFS increase land use efficiency in many cases (e.g., Dupraz et al. 2018; Graves et al. 2007), harvests of the individual crops in AFS tend to show lower yields (in particular close to the tree rows) in temperate climate (Dufour et al. 2013; Pardon et al. 2018; Carrier et al. 2019; Swieter et al. 2019). Hence, the use of CCPs in AFS, as suggested by Smith et al. (2012b) seems a promising approach to better match the heterogenous and varying environmental conditions in AFS than varieties bred for high-input and mono-cropping agriculture. Breeding and effective selection of alleles connected to maximum and/or stable yield, respectively, in competitive, heterogeneous and low-input environments as often found in AFS should ideally be undertaken under exactly such conditions. The best varieties specifically targeted for those environments are likely to be produced in breeding programs undertaken in such conditions (Atlin and Frey 1989).
In this study, we set out to test the potential of CCPs for improved yield and quality of wheat under AFS conditions. More specifically, we tested the hypothesis that a wheat CCP will outperform a commercial cultivar monoculture in terms of yield and quality in particular in proximity to the tree row. To do this, wheat was grown at two agroforestry sites and its yield and quality parameters were collected to compare the performance of the CCP and the commercial variety in a field trial under AFS conditions. It was assumed that the plant density, yield and thousand grain weight as well as various quality traits (such as protein and mineral content) of CCP would perform better, especially near the tree.
Materials and methods
Plant material
A common winter wheat variety (‘Wiwa’) and a winter wheat CCP (‘CC-2k’) were used in the field experiment. The variety Wiwa is an obtention of the plant breeder Getreidezüchtung Peter Kunz (gzpk) in Hombrechtikon, Switzerland. Released in 2005, Wiwa has been ranked to Swiss baking class TOP (organic class 1) because of its elevated baking properties (Dierauer 2020) and is recommended for cultivation in organic agriculture. CC-2k is a composite cross obtained as a diallel cross of 20 wheat varieties and breeding lines from Switzerland and from Europe (A. Schori and V. Michel, personal communication). The composite cross population was developed in 2000 and has been cultivated on 10 × 10 m plots (100 m2) surrounded with a 5 m triticale buffer, during 8 years to obtain the F10 generation (F1012600900), at the Agroscope site in Changins (Nyon, Switzerland). For the present experiments, seed material has been produced with seeds from generation 8 on a 150 m2 plot to obtain generation 11 (F1112600900).
Experimental sites
Field experiments were carried out between autumn 2021 and summer 2022 at two organically managed AFS in Feusisberg and Wollerau in Switzerland. Both farms are located in the Kanton Schwyz in Central Switzerland. A silvoarable AFS, with standard fruit trees (apple, pear, plum) and walnut trees arranged in west–east oriented tree rows, is located in Wollerau (N 47°10′53.928″, E 8°43′56.188″) on a farm with 4.5 ha. The crop rotation at the experimental field includes lupin, barley, oat, wheat triticale with clover undersown and wheat. The soil is a medium-heavy, slightly acidic loam. The experimental field lies 620 m above sea level on the Northwest side of 16 years old standard apple trees. Standard plum trees are also arranged in west–east oriented tree rows in a natural meadow which was ploughed for the purpose of the experiment in Feusisberg (N 47°10′53.928″, E 8°43′56.188″). The whole farm size amounts to 14 ha. The soil is a sandy, slightly alkaline loam rich in humus. The experimental field lies 705 m above sea level on the North side of a 30-year-old plum tree row. Both farms’ climate are classified as warm and temperate (“Cfb” in Köppen and Geiger classification) with a mean annual temperature of 8.1 °C and a mean annual precipitation of 1618 mm, with high amounts of precipitation even in the driest month (February, 12 rainy days) (https://de.climate-data.org, 2022). May has the highest average number of rainy days per month (18 days). Soil carbon, nitrogen and total phosphorous content amounted to 29.9 ± 5.0 g C kg‑1, 3.4 ± 3.4 g N kg‑1 and 1000 ± 104 mg P kg−1 for Wollerau and 58.3 ± 10.6 g C kg‑1, 5.8 ± 0.9 g N kg‑1 and 935 ± 129 mg P kg−1 for Feusisberg, respectively; these concentrations correspond to averaged C, N and P concentrations found in agricultural soils in Switzerland (C: 31.3 g kg‑1, N: 2.9 g kg‑1, P: 932 mg kg−1, source: NABO, personal communication), with the exception of higher C and N levels in Feusisberg. The C:N ratio for Wollerau was 8.8, for Feusisberg: 10.2. More information on the experimental sites and their management can be found in the supplementary information SI Tab. 1 and Tab. 2.
Site management
The field was tilled with a plough and a harrow in the first week of October 2021. Wiwa and CC-2 k were both sown on 15 October 2021 with 350 grains per m2 using a small drag coulter seed drill, available at the farm.
Fertilisation was carried out according to farmer’s practice with compost from farmyard manure and green waste (SI Tab. 2). At the Wollerau site, additional fertilisation with liquid manure (cattle slurry with low fecal matter and urine manure diluted 1:2, 400 hl ha−1) was necessary as plants showed yellowing and chlorophyll measurements showed a deficient nutrient status.
Experimental design
Wiwa and CC-2k were planted in two 1.8 × 9 m strips (“long plot”) in a perpendicular manner on the North side of a tree row (see Fig. 1) to include the effect of shade cast by the 4 to 6 m high trees at a distance of 1 to 2 m from the field. The Wiwa and CC-2k long plots were planted at 2.5 m from the trunk of a reference tree in a “split-plot” design with three replicates per site. These long plots were surrounded by either Wiwa or CCP plants. Within each long plot, four observation and sampling plots of 1 m2 for measurements on and harvest of individual plants were defined at 1, 2.4, 3.8 and 7 m distance from the tree-row border (Fig. 1).

Experimental design at two Swiss agroforestry systems in Feusisberg and Wollerau, respectively. Each replicate consisted of a strip (“long plot”) with the composite cross population “CC-2k” and commercial variety “Wiwa”. Physiological and yield parameters were collected from 1 m2 plots at four distances (1, 2.4, 3.8 and 7 m) from the tree row
Plant physiological measurements
Plant physiological measurements were carried out in the sampling plots at 1.0, 2.4, 3.8 and 7.0 m distance from the tree row. Leaf chlorophyll content and stomatal conductance were measured to allow a comparison of potential shade-related increases in chlorophyll content and gas exchange and transpiration rates as indicators for photosynthetic activity. Leaf chlorophyll content was assessed indirectly by usage of a chlorophyll meter (SPAD-502Plus Konica Minolta®). Stomatal conductance was determined with a leaf porometer (SC-1 Leaf Porometer from METER Group®). Chlorophyll and stomatal conductance measurements were conducted once per site in April, May and June on two subsequent sunny days around midday for the SPAD measurements and during the morning (9–11 am) and afternoon (3–5 pm) hours for the porometer measurements on three individual plants within each plot.
Yield and quality parameters
At full maturity (BBCH 89), four individual plants in each plot were randomly collected for individual phenotypic trait measurements (plant height, total grain weight, grain mass, number of grains) and the remaining plants of the 1 m2 fields were counted and manually harvested (22–24 July 2022). The harvested material (ears with short stalks) was threshed with a plot harvester machine “HEGE 150”. Grains were stored in paper bags under dark and dry conditions at room temperature. Thousand grain weight (TGW) and hectolitre weight were measured with a MARVIN optical grain counter (Digital Seed Analyser, GTA Sensorik GmbH, Neubrandenburg, Germany) and a balance (Mettler PM2000, Mettler-Toledo, Greifensee, Switzerland). Protein (%) and mineral contents (% or mg kg−1) were analysed by near-infrared reflectance spectroscopy (NIRS) using a NIRFlex N-500 (Büchi Labortechnik AG). The protein calibration of the NIRFlex was annually adjusted with 50–100 wheat samples from different varieties and origins. Reference analyses were made with the Kjeldahl method, according to ICC standard method No. 105/2. The coefficient of confidence of the calibration was R2 = 0.93 (Cécile Brabant, personal communication). The protein content of the herein examined samples fitted into the range of the NIRS calibration.
Environmental parameters
Photosynthetically active radiation (PAR) sensors (Apogee QSO-S PAR Photon Flux, METER Environment) were installed on 27 February 2022 in three replicates (one in Feusisberg, two in Wollerau) at three positions per replicate, i.e., 0, 3.8 and 7 m from the tree row (Fig. 1). Sensors were placed at 1.3 m height and connected to ECHO Em50 Dataloggers. PAR was measured every 10 min as Photosynthetically Active Photon Flux Density (PPFD) in µmol m−2 s−1 until harvest. Volumetric soil water content measurements were carried out with a ML3 ThetaProbe Soil Moisture Sensor (Delta-T, Cambridge) at two time points (21 April, 26 May). Measurements were taken in the centre of the plots and a standard calibration for mineral soil was used.
Data analyses
Statistical analyses were carried out with R version 3.6.1. The data was tested for normality and homogeneity of variance by a visual inspection of residuals (normal quantile–quantile plots, standardised residuals versus fit plots) and revision of coefficients of determination (R2). When necessary, logarithmic or square root transformations were applied to achieve normality and homogeneity of variance. The split-plot design was accounted for by including “longplot” in the random term and a spatial correlation with coordinates of distances from the tree row (“d_cor”), where the lack of randomization in terms of distance was accounted for by using a Gaussian correlation structure (“corGaus”) (Piepho et al. 2004). The random term further included a factorial term with site and replicate (“siterep”). The model explaining yield, and plant density, at the plot level was thus: lme(sqrt(yield) ~ site*distance*variety, and lme(plantnumber ~ site*distance*variety, random = ~ 1|siterep/longplot, correlation = corGaus(form = ~ d_cor)), where “distance” was a factor with four levels and “variety” referred to the variety ‘Wiwa’ and the ‘CC-2k’ population. For better readability, the term “variety” is used in the remainder of this document to refer to both variety and population. At the individual level, “plot” was nested additionally in the random term and a second coordinate (which was randomly assigned to each individual sample) perpendicular to the distance-coordinate had to be given not to create zero distances in the model. Differences in group means among groups was analysed by multifactorial ANOVA (type I, sequential sum of squares). Significances of each factor were assessed by means of the F-test. For post-hoc analysis, the means of treatment groups were compared with a Tukey test (HSD.test()-function within the R agricolae package (de Mendiburu 2020)) with a significance threshold of α = 0.05.
To calculate the relative amount of PAR at different distances from the tree row, the total amount of PAR per day (PPFD in µmol m-2/day) was calculated for each sensor with the formula \(PPFD= \sum_{0}^{n}{m}_{n}*600+ \frac{{m}_{n}- {m}_{n-1}*600}{2}\), where “m” was the measurement and “n” the number of measurements per day. All measurements were exactly 10 min apart (600 s) apart. Finally, the sensor placed at 7 m distance to the tree row was taken as the reference and the percental PPFD for the 0 and 3.8 m distance were calculated with the formula \(\% PPFD= \frac{{PPFD}_{d 1/d2}}{{PPFD}_{d 3}}* 100\%\). Statistical analyses of chlorophyll content, stomatal conductance and soil moisture followed the model outlined above for analyses on the individual level, with the addition of “month” as fixed factor.
Results
Plant Density, Yield and Thousand Grain Weight
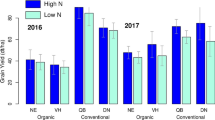
Plant density was significantly influenced by variety (p < 0.05) and interactions of site and distance (p < 0.001) and site and variety (p < 0.01). In Feusisberg, plant density at 3.8 m from the tree row (41 plants m−2) was significantly higher than plant density in Wollerau at 3.8 m (20 plants m−2), all other distances had intermediate plant densities (27–40 plants m−2). Plant densities of CC-2k and Wiwa were similar in Feusisberg, but statistically different in Wollerau (CC-2k: 45 ± 14 SD plants m−2, Wiwa: 16 ± 8 plants m−2) (Fig. 2). The correlation coefficient between yield and plant number was 0.70.
Variety (p < 0.05) and the interactions of site and distance (p < 0.05) and site and variety (p < 0.05) significantly affected wheat yield at the plot level. Yield of CC-2k (1.9 ± 0.8 Mg ha−1) was significantly higher than yield of Wiwa (1.4 ± 0.9 Mg ha−1) across both sites and all distances. However, CC-2k and Wiwa yield (across all distances) amounted to 1.9 ± 0.7 Mg ha−1 and 2.0 ± 0.8 Mg ha−1, respectively, in Feusisberg and to 1.9 ± 0.9 Mg ha−1 and 0.7 ± 0.4 Mg ha−1 in Wollerau, i.e. mean CC-2k yield was identical for Feusisberg and Wollerau while mean Wiwa yield in Wollerau was only 35% of mean Wiwa yield in Feusisberg (Fig. 2). In Feusisberg, yield was highest at 3.8 m (2.3 ± 0.4 Mg ha−1) and similarly low at 1.0, 2.4 and 7.0 m (1.8 ± 0.8, 1.7 ± 0.7 and 1.9 ± 0.9 Mg ha−1, respectively). In Wollerau, yield at 1.0, 2.4 and 7.0 m was similar (1.3 ± 1.1, 1.4 ± 0.9 and 1.5 ± 0.9 Mg ha−1, respectively), at 3.8 m it was lowest (1.1 ± 0.9 Mg ha−1). In the post-hoc Tukey test, Wiwa yield in Wollerau was significantly lower than both Wiwa yield in Feusisberg and CC-2k yield in Feusisberg and Wollerau. The interaction of population/variety and distance varied with CC-2k having higher yields in proximity to the tree row in Feusisberg but lower ones in Wollerau compared to the reference yield at 7.0 m distance. For Wiwa, a decreased yield in proximity to the tree row was observed in Feusisberg for the 1.0 and 2.4 m distance, while in Wollerau Wiwa yield was in general very low, amounting to 0.7 to 0.8 Mg ha−1 at all distances (Fig. 2). To compare yield variability, the coefficient of variation, also known as relative standard deviation, as the ratio of the standard deviation to the mean was calculated for each variety, being 40% for CC-2 k and 64% for Wiwa across the two sites.

Plant density (A) and yield (B) of the winter wheat composite cross population ‘CC-2k’ and the common organic variety ‘Wiwa’ at two agroforestry systems in Switzerland. Site-variety-distance effects are shown. Different lowercase letters indicate significant differences at p < 0.05 probability level. The box plots range from the first to the third quartile where the horizontal line shows the median. The vertical lines go from each quartile to the minimum or maximum, respectively
Thousand grain weight (TGW) derived from subsamples analysed with MARVIN was significantly affected by distance (p < 0.05). Across both sites, mean TGW was highest at 1.0 and 7.0 m (39.4 ± 3.6 and 39.4 ± 2.9 g, respectively) and lowest at 3.8 m (35.7 ± 5.7 g). The difference between Wiwa (39.0 ± 4.1 g) and CC-2k (37.5 ± 4.1 mg) was not significant. TGW was higher in Feusisberg (39.8 ± 3.1 mg) than Wollerau (36.7 ± 4.5 mg). Hectolitre weight differed between varieties (p < 0.05) with significantly higher weights for Wiwa (77.1 ± 3.4 kg hl−1) than CC-2k (73.6 ± 2.6 kg hl−1).
The interaction of site and distance was significant (p < 0.0001) for total seed weight per plant and total seed number. Differences in total seed weight and seed number per plant were statistically not significant between any distances in Feusisberg. Total seed weight was significantly different between 3.8 m (7.9 ± 4.9 g) and 1.0 m (4.7 ± 4.3 g) and total seed number was significantly different between 3.8 m (181 ± 96 seeds) and 1.0 m (98 ± 85) in Wollerau. Distance was marginally significant (p = 0.06) for total seed number with the highest average seed number at 3.8 (151 ± 85), followed by distance 1.0 (131 ± 94), 7.0 (122 ± 63) and 2.4 m (113 ± 58), respectively (across both sites). No effects of site, distance and variety were observed on tillering.
Quality parameters and plant height
Grain hardness was significantly affected by variety (p < 0.01) and the interaction of variety and distance (p < 0.01) where CC-2k had a higher mean grain hardness (26.3 ± 1.2%) compared to Wiwa (25.0 ± 5.0%). While grain hardness was generally more homogenous in CC-2k and only slightly decreased at 1.0 m distance, it was highest at 1.0 m distance in Wiwa (Fig. 3).

Protein content, grain hardness and content of insoluble and soluble fibers of the winter wheat composite cross population ‘CC-2k’ and the common organic variety ‘Wiwa’ at two agroforestry systems in Switzerland. The distances refer to the distance to the tree row. Different lowercase letters indicate significant differences at p < 0.05 probability level. The box plots range from the first to the third quartile where the horizontal line shows the median. The vertical lines go from each quartile to the minimum or maximum, respectively
Protein content was significantly affected by variety (p < 0.01) and marginally significantly by site (p = 0.06). Protein content was significantly higher in Wiwa (12.7 ± 1.0%) than in CC-2k (11.1 ± 0.6%) and in Wollerau (12.2 ± 1.3%) than Feusisberg (11.5 ± 0.9%) (Fig. 3).
Potassium (K), phosphorus (P), calcium (Ca), magnesium (Mg), iron (Fe) and zinc (Zn) content depended on variety (p = 0.07, p < 0.01, p = 0.06, p < 0.01, p = 0.01, p < 0.01, respectively) (Fig. 4, see also SI Tab. 10). Some nutrients showed effects by site (Mg), distance (K, P) and interactions of site and variety (Ca, Zn), site and distance (P) and distance and variety (P). Mg content was higher in Wollerau than Feusisberg across both varieties and all distances. For Zn, contents were equal in CC-2k at both sites (Feusisberg: 28.2 ± 1.3 mg kg−1, Wollerau: 28.3 ± 1.7 mg kg−1) but significantly different for Wiwa between Wollerau (33.6 ± 0.9 mg kg−1) and Feusisberg (30.3 ± 1.9 mg kg−1). Wiwa had a significantly higher content of Fe (46.1 ± 4.0 mg kg−1) compared to CC-2k (38.5.1 ± 3.3 mg kg−1). For Ca, the difference in content between Wiwa and CC-2k was significant in Wollerau (Wiwa: 460 ± 30 mg kg−1, CC-2k: 420 ± 30 mg kg−1) but similar for both varieties in Feusisberg (Wiwa: 430 ± 20 mg kg−1, CC-2k: 430 ± 30). K content showed an interaction of site and distance: In Wollerau no significant difference between distances was observed, whereas K content in Feusisberg was significantly higher at 7.0 m (3,950 ± 230 mg kg−1) than at 1.0 m (3,730 ± 100 mg kg−1). Concerning the interaction of distance and variety, no difference between distances were observed for CC-2k while P content was significantly lower for Wiwa at 1.0 m (3,870 ± 150 mg kg−1) than at 3.8 and 7 m (4,070 ± 160 and 4,080 ± 120 mg kg−1).

Mineral contents of the winter wheat composite cross population ‘CC-2k’ and the common organic variety ‘Wiwa’ at two agroforestry systems in Switzerland. The distances refer to the distance to the tree row. The box plots range from the first to the third quartile where the horizontal line shows the median. The vertical lines go from each quartile to the minimum or maximum, respectively
With respect to insoluble fibres, CC-2k had the highest soluble fibre content at 1.0 (2.33 ± 0.18%) and the lowest at 3.8 m (2.13 ± 0.033%) while it was the opposite for Wiwa (lowest content at 1.0 m with 2.12 ± 0.21% and highest content at 3.8 m with 2.38 ± 0.17%). For insoluble fibres, site (p < 0.05) and variety (p < 0.05) were significant with higher contents in Wollerau (7.09 ± 1.16%) compared to Feusisberg (6.17 ± 1.05%) and higher in Wiwa (7.17 ± 1.23%) compared to CC-2k (6.08 ± 0.88%).
Distance also influenced plant height significantly (p < 0.001), with plants being higher close to the tree (distance 1–4: 90 ± 10 cm, 88 ± 8 cm, 87 ± 9 cm and 84 ± 10 cm, respectively). Wiwa was marginally significantly (p = 0.05) higher (89 ± 8 cm) than CC-2k (85 ± 10 cm) across both sites. No effects of site, distance and variety were observed on tillering.
Chlorophyll content and stomatal conductance
Chlorophyll content was significantly affected by month (p < 0.01), site (p < 0.05) and distance (p < 0.01). The interactions of site and distance (p = 0.07) and site and variety (p = 0.07) were statistically not significant but relevant to notice. Chlorophyll content increased with time and reached highest values in June (Fig. 5). Plants in Feusisberg had significantly higher chlorophyll contents (43.3 ± 4.6) than in Wollerau (40.4 ± 4.4). In terms of distance to the tree row, chlorophyll content was higher at 2.4 and 3.8 m (42.9 ± 4.8 and 42.7 ± 4.1, respectively) than at 1.0 and 7.0 m (41.6 ± 5.2 and 40.3 ± 4.3, respectively) across both sites. In Feusisberg, it was highest at 2.4 m, followed by 1.0, 3.8 and 7.0 m. In Wollerau, it was highest at 3.8 m, followed by distance 2.4, 1.0 and 7.0. In Feusisberg, chlorophyll content of CC-2k (43.7 ± 5.0) and Wiwa (43.0 ± 4.2) were the same, but in Wollerau, Wiwa had a significantly higher chlorophyll content (41.1 ± 4.0) than CC-2k (39.6 ± 4.6).

Leaf chlorophyll content (A) and stomatal conductance (B) of the winter wheat composite cross population ‘CC-2k’ and the common organic variety ‘Wiwa’ at two agroforestry systems in Switzerland. Measurements were taken on two subsequent sunny days from April to June. Different lowercase letters indicate significant differences at p < 0.05 probability level. The box plots range from the first to the third quartile where the horizontal line shows the median. The vertical lines go from each quartile to the minimum or maximum, respectively
Stomatal conductance was significantly determined by month (p < 0.01), distance (p < 0.01) and the interaction of site and distance (p < 0.01). It increased significantly with time (April: 133 ± 70 mmol m−2 s−1, May: 214 ± 89 mmol m−2 s−1, June: 314 ± 101 mmol m−2 s−1). Regarding the site-distance interaction, stomatal conductance was not significantly different between any distances in Wollerau; in Feusisberg stomatal conductance at 7.0 m was significantly higher than at 2.4 and 1.0 m (183 ± 91 and 173 ± 97 mmol m−2 s−1, respectively) (Fig. 5, see also SI Fig. 2).
Photosynthetically active photon flux density and volumetric soil moisture
The PAR sensors placed at 7 m distance had a higher PPFD than the sensors on the South side of the tree row intended as control. Thus, the relative minima, maxima and means of PAR were calculated from division by the sensors placed in 7 m distance to the tree row within each replicate. Across both sites, relative mean was 71 ± 18% PAR at the edge of the tree row and 99 ± 9% PAR at 3.8 m distance (Fig. 6). At the edge of the tree row, relative maximum was equal to the reference sensor, but relative minimum amounted to 51 ± 23% of it. At 3.8 m distance, relative minimum amounted to 88 ± 18%.

Percentage of Photosynthetically Active Photon Flux Density (PPFD) at the tree row border (0 m distance) and at 3.8 m distance to it at the two experimental sites in Switzerland in comparison to the reference PAR sensor placed at 7 m distance. There was one replicate (R2) with three sensors in Feusisberg and two replicates (R1, R3) with three sensors in Wollerau
Volumetric soil moisture was significantly affected by month (p < 0.0001) and the interaction of distance and variety (p < 0.05). Distance (p = 0.057) was marginally significant. Soil moisture was significantly higher in May (33.3 ± 5.6%) than April (20.2 ± 8.5%) and at 2.4 m distance to the tree row (31.5 ± 9.6%) than at 1 m (28.3 ± 8.6%). Soil moisture at 3.8 and 7 m distance lay in between. In the CC-2k plots, soil moisture was similar across all distances, except for significantly higher soil moisture at 2.4 m distance (33.9 ± 9.6%) than at 1 m distance (27.1 ± 8.8%).
Discussion
The main hypothesis of this field experiment was that a genetically diverse cereal population would outperform a high-yielding commercial variety in a heterogenous environment of an AFS. The underlying assumption is that breeding efforts have created high-yielding modern varieties which are designed for high-input sole crop agriculture, have specific environmental requirements and are not particularly suitable for low-input mixed cropping systems like silvoarable agroforestry (Atlin and Frey 1989; Smith et al. 2012a; Bourke et al. 2021). According to Lammerts van Bueren et al. (2011), 95% of organic production is based on crop varieties bred for conventional agriculture, which has substantially different input levels than organic farming (see also Le Campion et al. 2020). Given that Wiwa is a long established (admission in 2005) and the most commonly grown organic winter wheat variety in Switzerland today (Klaiss and Dierauer 2019), its selection as reference variety was solid. In our field experiment at two agroforestry sites in temperate climate, yield of the CCP CC-2k was more stable across the two sites than yield of the commercial organic variety Wiwa; in the sense that CC-2k showed similar yields in Feusisberg and Wollerau while yield of Wiwa was significantly less in Wollerau than Feusisberg. Also, the coefficient of variation across both sites was lower for CC-2k (40% compared to 64% for Wiwa). More sites and seasons are obviously needed for a statistically sound assessment (see the section about limitations of this study at the end of this discussion), but our results are a first indicator.
In terms of yield, CC-2k outperformed Wiwa in Wollerau and a yield-related interaction of variety and distance was not observed. In comparison to average winter wheat yields from organic production in Switzerland of 4.5 ± 1.3 Mg ha−1 in 2022, the achieved yields of 1.9 ± 0.8 Mg ha−1 (CC-2k) and 1.4 ± 0.9 Mg ha−1 (Wiwa) across all distances and both sites are low (Agristat 2022). Even the higher yields at 7 m distance to the tree row (2.2 ± 0.3 Mg ha−1 for CC-2k and 2.3 ± 0.7 Mg ha−1 for Wiwa) were only half of the average yield in Switzerland. We attribute this low yield to various reasons: relatively high losses due to the use of old machinery equipment (seeder and thresher) with small plot sizes, weed pressure and bird predation. This may have mitigated differences between plot distances near the tree row and the reference plot at seven metres which would have been expected (as they occurred, e.g., in Pardon et al. 2018). At the same time, tree height was maximum 6 m and trees were positioned 1.3–2.3 m from the border of the tree row. Plots at 7 m distance to the tree row could have been shaded only for a very short period of time at dawn and dusk. PAR sensor data further showed that even 4 m from the tree row light conditions were very similar to the reference sensor at the 7 m distance position. The results should therefore not be interpreted to mean that shading in AFS was the cause of the lower yields. Indeed, the small magnitude of the differences between the distances in all parameters suggests that other factors were of greater importance than the influence of the tree row. Plant density is of major importance to maximise yield (Bastos et al. 2020) and final plant density in our study was clearly below the initial seeding density of approx. 350 seeds per m2. Though in high-yielding environments, the agronomic optimum planting density is considerably lower than in low- or medium-yielding environments, still its estimates range from 140 to 400 plants per m2 (Bastos et al. 2020), being well above the attained plant densities in our trial.
Most wheat parameters showed no effects of distance to the tree row. In contrast, reductions of light and soil moisture were observed for the 1 m distance. However, shading and competition for water were not really reflected in the wheat parameters. There was also no difference in wheat parameters at the 2.4 m distance, where soil moisture was apparently increased by shading, or at the area outside the influence of the tree. The fact that both soil moisture and light changed with distance, but the wheat parameters hardly changed at all, is interesting and can be interpreted as a physiological plasticity to cope with heterogeneous conditions in wheat. As shown in a 40-year long experimental trial, varieties which performed well at high-input levels were also the best at low input levels, suggesting that interactions between the genotype and environmental conditions were not strong enough to inverse the performance ranking (Büchi et al. 2016). Overall, CC-2k and Wiwa demonstrated similar plasticity in terms of distance-effects in this trial.
TGW commonly decreases with shade (e.g., Qiao et al. 2019; Artru et al. 2017; Li et al. 2010). Other studies have reported increased (Zhang et al. 2022) or similar (Zhang et al. 2022; Vaccaro et al. 2022) TGWs under moderate shade. In this study, TGW was below average at both sites (gzpk 2022, Agroscope, personal communication), and highest at 1 and 2.4 m distance for CC-2k and Wiwa, respectively, and lowest, for both varieties, at 3.8 m distance. However, PAR measurements showed that the mean reduction in PPFD at 3.8 m distance was minor. Apart from light, temperature, water availability and micronutrients may affect TGW (Kaur and Behl 2010; Arif et al. 2006). Soil moisture measurements did not indicate competition for water at 3.8 m distance, though, and neither P nor micronutrient contents showed reduced values at 3.8 m distance. Total seed weight per plant was also lowest at 3.8 m, while total seed number was highest there. The negative relationship between seed number and weight is well-known (Knott and Talukdar 1971) and was similar for CC-2k and Wiwa.
Protein content, as an important parameter for baking quality, was slightly lower in CC-2k, though still in the range of minimum protein content for white flour which is specified at 10–12% (Kamel and Stauffer 1993). It is well known that the relationship between protein content and bread volume drops above 12% protein content (Gabriel et al. 2017). In a study over a 5-year-period, baking quality of 11 winter wheat varieties were assessed and the influence of year was higher for all evaluated baking quality characteristics compared to the influence of variety – with the influence of year being most important to protein content (Koppel and Ingver 2010). Some quality parameters were slightly higher in Wiwa (protein content, Fe content, insoluble fibres) but also showed interactions with site (e.g., significantly different Ca content between Wiwa and CC-2k in Wollerau but not in Feusisberg). Most mineral nutrient contents were higher in Wiwa than in CC-2k, though.
The significant interaction of site and variety, which was of greater importance than distance-effects in terms of yield and mineral nutrient content, may be explained by the lower soil fertility in Wollerau. Besides the lower C:N ratio (8.8) observed in soil samples, leaf measurements with the chlorophyll meter suggest significantly lower nutrient contents in Wollerau than Feusisberg. The CC-2k as a CCP proved to be more suited to cope with lower nutrient availability in this trial, as proposed by plant breeders (Döring et al. 2011; 2015; Atlin and Frey 1989; Smith et al. 2012b; Bourke et al. 2021).
There is the aspect of natural adaptation over time of CCP grown year after year in the same location and the evolution of facilitative interactions, which should lead to higher and more stable yields (Schöb et al. 2018; Vollenweider and Spieß 2018). Improvements can certainly be expected compared to varieties bred in highly fertile, weed-free, densely seeded environments (Atlin and Frey 1989). Besides that, the importance of maintaining and enhancing agrobiodiversity and diverse genetic varieties is emphasized and encouraged by major agreements and other policy instruments such as the FAO’s International Treaty on Plant Genetic Resources for Food and Agriculture and the Nagoya Protocol on Access to Genetic Resources and the Fair and Equitable Sharing of Benefits Arising from their Utilization.
Lastly, the authors of this study would like to emphasize the limitations of this study. Results from a one-year experiment at two sites must not be generalised, in particular as the study involves a CCP, whose inherent characteristic is high adaptability. The presented findings are limited to the growing season of 2021/2022 and contingent to the climate of this experimental year, i.e., rather wet soil and weather conditions in autumn during sowing and (late) spring, but also warm temperatures in spring. Furthermore, the results should also be seen in context: It was a trial at two locations that were geographically adjacent but differed greatly in terms of microclimate (higher sea level and a higher tree density around the trial field at Feusisberg), soil properties (humus-rich soil at Feusisberg), management (two farmers with different practices, in particular years of spreading high-quality compost produced on the farm and a natural meadow at Feusisberg compared to a long-term crop rotation at Wollerau) and properties at field level (tree species, distance between trees, slope). In this sense, the lack of replication in time and limited replication in space and the limitations of randomisation due to given circumstances at farms (farm- and field-specific conditions) prohibit a generalisation of the results found. Instead, these should be seen as initial insights of a case study, which have to be complemented by further field experiments and future larger research projects.
Conclusion
In summary, the CC-2k outperformed the commercial organic variety Wiwa in terms of yield but not in terms of grain quality at one agroforestry site during one growing year of experimentation. As variety-/population-specific yield was independent from the distance to the tree row, it may be concluded that shade-induced reductions in yield-related characteristics must not have been decisive or might have been outweighed by positive interactions or a beneficial microclimate in the AFS of this experiment. However, these initial results must be seen in their specific context and require further and larger field studies in order to derive general trends.
References
Agristat (2022) ‘Berichterstatter-Erhebung [Unpublished Raw Data]’
Arif M, Chohan MA, Ali S, Gul R, Khan S (2006) Response of Wheat to Foliar Application of Nutrients. J Agricult Ural Biol Sci 1(4):30–34
Artru S, Garré S, Dupraz C, Hiel M-P, Blitz-Frayret C, Lassois L (2017) Impact of Spatio-Temporal Shade Dynamics on Wheat Growth and Yield, Perspectives for Temperate Agroforestry. Eur J Agron 82(January):60–70. https://doi.org/10.1016/j.eja.2016.10.004
Atlin GN, Frey KJ (1989) Breeding Crop Varieties for Low-Input Agriculture. Am J Altern Agric 4(2):53–58. https://doi.org/10.1017/S0889189300002721
Bastos LM, Carciochi W, Lollato RP, Jaenisch BR, Rezende CR, Rai Schwalbert PV, Prasad V et al (2020) Winter Wheat Yield Response to Plant Density as a Function of Yield Environment and Tillering Potential: A Review and Field Studies. Front Plant Sci 11(March):54. https://doi.org/10.3389/fpls.2020.00054
Bourke PM, Evers JB, Bijma P, van Apeldoorn DF, Smulders MJM, Kuyper TW, Mommer L, Bonnema G (2021) Breeding Beyond Monoculture: Putting the “Intercrop” Into Crops. Front Plant Sci 12(November):734167. https://doi.org/10.3389/fpls.2021.734167
Büchi L, Charles R, Schneider D, Sinaj S, Maltas A, Fossati D, Mascher F (2016) Performance of Eleven Winter Wheat Varieties in a Long Term Experiment on Mineral Nitrogen and Organic Fertilisation. Field Crop Res 191(May):111–122. https://doi.org/10.1016/j.fcr.2016.02.022
Carrier M, Gonzalez F-A, Cogliastro A, Olivier A, Vanasse A, Rivest D (2019) Light Availability, Weed Cover and Crop Yields in Second Generation of Temperate Tree-Based Intercropping Systems. Field Crop Res 239(June):30–37. https://doi.org/10.1016/j.fcr.2019.05.004
Dierauer H (2020) ‘Bioweizen Mit Guten Erträgen Bei Mittlerer Qualität’. Schweizer Bauer. p 14. https://orgprints.org/id/eprint/39397/1/Bioweizen-Ernte%202020%20mit%20guten%20Ertr%C3%A4gen%20bei%20mittlerer%20Qualit%C3%A4t.pdf. Accessed 12 Sept 2020
Döring TF, Knapp S, Kovacs G, Murphy K, Wolfe MS (2011) Evolutionary Plant Breeding in Cereals—Into a New Era. Sustainability 3(10):1944–1971. https://doi.org/10.3390/su3101944
Döring TF, Annicchiarico P, Clarke S, Haigh Z, Jones HE, Pearce H, Snape J, Zhan J, Wolfe MS (2015) Comparative Analysis of Performance and Stability among Composite Cross Populations, Variety Mixtures and Pure Lines of Winter Wheat in Organic and Conventional Cropping Systems. Field Crop Res 183(November):235–245. https://doi.org/10.1016/j.fcr.2015.08.009
Dufour L, Metay A, Talbot G, Dupraz C (2013) Assessing Light Competition for Cereal Production in Temperate Agroforestry Systems Using Experimentation and Crop Modelling. J Agron Crop Sci 199(3):217–227. https://doi.org/10.1111/jac.12008
Dupraz C, Lawson GJ, Lamersdorf N, Papanastasis VP, Rosati, Ruiz-Mirazo J (2018) ‘Temperate Agroforestry: The European Way.’ In Temperate Agroforestry Systems, edited by A. M. Gordon, S. M. Newman, and B. R. W. Coleman, 2nd ed., 98–152. Wallingford: CABI. https://doi.org/10.1079/9781780644851.0098
Duru M, Therond O, Martin G, Martin-Clouaire R, Magne M-A, Justes E, Journet E-P et al (2015) How to Implement Biodiversity-Based Agriculture to Enhance Ecosystem Services: A Review. Agron Sustain Dev 35(4):1259–1281. https://doi.org/10.1007/s13593-015-0306-1
Fanzo J, Hunter D, Borelli T, Mattei F (eds) (2013) Diversifying Food and Diets: Using Agricultural Biodiversity to Improve Nutrition and Health. 1st edn. Issues in Agricultural Biodiversity. London ; New York: Earthscan from Routledge
Gabriel D, Pfitzner C, Haase NU, Hüsken A, Prüfer H, Greef J-M, Rühl G (2017) New Strategies for a Reliable Assessment of Baking Quality of Wheat – Rethinking the Current Indicator Protein Content. J Cereal Sci 77(September):126–134. https://doi.org/10.1016/j.jcs.2017.08.002
Graves AR, Burgess PJ, Palma JHN, Herzog F, Moreno G, Bertomeu M, Dupraz C et al (2007) Development and Application of Bio-Economic Modelling to Compare Silvoarable, Arable, and Forestry Systems in Three European Countries. Ecol Eng 29(4):434–449. https://doi.org/10.1016/j.ecoleng.2006.09.018
gzpk (2022) ‘Rohdaten Wiwa [Unpublished Raw Data]’
Jose S (2009) Agroforestry for Ecosystem Services and Environmental Benefits: An Overview. Agrofor Syst 76(1):1–10. https://doi.org/10.1007/s10457-009-9229-7
Jose S, Gillespie AR, Pallardy SG (2004) Interspecific Interactions in Temperate Agroforestry. Agrofor Syst 61:237–255
Kaur V, Behl R (2010) Grain Yield in Wheat as Affected by Short Periods of High Temperature, Drought and Their Interaction during Pre- and Post-Anthesis Stages. Cereal Research Communications 38(4):514–520. https://doi.org/10.1556/CRC.38.2010.4.8
Klaiss M, Dierauer H (2019) ‘Ergebnisse Aus Den Weizensorten Streifenversuchen 2019’. FiBL. https://orgprints.org/id/eprint/37235/1/klaiss-dierauer-2019-Bericht-MahlWeizenVersucheKABB-2019_191126_Def.pdf
Knapp S, Döring TF, Jones HE, Snape J, Wingen LU, Wolfe MS, Leverington-Waite M, Griffiths S (2020) Natural Selection Towards Wild-Type in Composite Cross Populations of Winter Wheat. Front Plant Sci 10(February):1757. https://doi.org/10.3389/fpls.2019.01757
Knott DR, Talukdar B (1971) Increasing Seed Weight in Wheat and Its Effect on Yield, Yield Components, and Quality 1. Crop Sci 11(2):280–283. https://doi.org/10.2135/cropsci1971.0011183X001100020034x
Kohli R, Kumar H, Singh P, Batish DR, Jose S (2007) 1 Ecological Interactions in Agroforestry: An Overview. Ecological Basis of Agroforestry 13
Koppel R, Ingver A (2010) ‘Stability and Predictability of Baking Quality of Winter Wheat’. Agronomy Research 8 (Special Issue III): 637–44. https://www.cabidigitallibrary.org/doi/pdf/10.5555/20113271032
Lammerts van Bueren ET, Jones SS, Tamm L, Murphy KM, Myers JR, Leifert C, Messmer MM (2011) ‘The Need to Breed Crop Varieties Suitable for Organic Farming, Using Wheat, Tomato and Broccoli as Examples: A Review’. NJAS: Wageningen Journal of Life Sciences 58 (3–4): 193–205. https://doi.org/10.1016/j.njas.2010.04.001
Le Campion A, Oury FX, Heumez E, Rolland B (2020) Conventional versus organic farming systems: dissecting comparisons to improve cereal organic breeding strategies. Organic Agriculture 10(1):63–74. https://doi.org/10.1007/s13165-019-00249-3
Li H, Jiang D, Wollenweber B, Dai T, Cao W (2010) Effects of Shading on Morphology, Physiology and Grain Yield of Winter Wheat. Eur J Agron 33(4):267–275. https://doi.org/10.1016/j.eja.2010.07.002
Lin BB (2011) Resilience in Agriculture through Crop Diversification: Adaptive Management for Environmental Change. Bioscience 61(3):183–193. https://doi.org/10.1525/bio.2011.61.3.4
Mendiburu, Felipe de (2020) ‘Agricolae: Statistical Procedures for Agricultural Research’. https://CRAN.R-project.org/package=agricolae
Morton LW (2019) ‘Farmers’ Knowledge and Adaptation to Climate Change to Ensure Food Security’. In Food Security and Climate Change, edited by S Yadav, R Redden, J Hatfield, A Ebert, and D Hunter, 449–70. Hoboken, NJ, USA, and Chichester, West Sussex, UK: Wiley-Blackwell
Obrecht A, Pham-Truffert M, Spehn E, Payne D, Altermatt F, Fischer M, Passarello C et al (2021) Achieving the SDGs with Biodiversity. Zenodo. https://doi.org/10.5281/ZENODO.4457298
Pardon P, Reubens B, Mertens J, Verheyen K, De Frenne P, De Smet G, Van Waes C, Reheul D (2018) Effects of Temperate Agroforestry on Yield and Quality of Different Arable Intercrops. Agric Syst 166(October):135–151. https://doi.org/10.1016/j.agsy.2018.08.008
Piepho HP, Büchse A, Richter C (2004) A Mixed Modelling Approach for Randomized Experiments with Repeated Measures. J Agron Crop Sci 190(4):230–247. https://doi.org/10.1111/j.1439-037X.2004.00097.x
Qiao Xu, Sai L, Chen X, Xue L, Lei J (2019) Impact of Fruit-Tree Shade Intensity on the Growth, Yield, and Quality of Intercropped Wheat. Edited by Guangyuan He PLOS ONE 14(4):e0203238. https://doi.org/10.1371/journal.pone.0203238
Rist S, Bonanomi EB, Giger M, Hett C, Scharrer B, Jacobi J, Lannen A (2020) Variety Is the Source of Life: Agrobiodiversity Benefits, Challenges, and Needs. Zenodo. https://doi.org/10.5281/ZENODO.3568133
Schöb C, Brooker RW, Zuppinger-Dingley D (2018) Evolution of Facilitation Requires Diverse Communities. Nature Ecology & Evolution 2(9):1381–1385. https://doi.org/10.1038/s41559-018-0623-2
Smith Jo, Pearce BD, Wolfe MS (2012a) Reconciling Productivity with Protection of the Environment: Is Temperate Agroforestry the Answer? Renewable Agric Food Syst 28(1):80–92. https://doi.org/10.1017/S1742170511000585
Smith Jo, Pearce BD, Wolfe MS (2012b) A European Perspective for Developing Modern Multifunctional Agroforestry Systems for Sustainable Intensification. Renewable Agric Food Syst 27(4):323–332. https://doi.org/10.1017/S1742170511000597
Suneson CA (1956) An Evolutionary Plant Breeding Method. Agron J 48(4):188–191. https://doi.org/10.2134/agronj1956.00021962004800040012x
Swieter A, Langhof M, Lamerre J, Greef JM (2019) Long-Term Yields of Oilseed Rape and Winter Wheat in a Short Rotation Alley Cropping Agroforestry System. Agrofor Syst 93(5):1853–1864. https://doi.org/10.1007/s10457-018-0288-5
Torralba M, Fagerholm N, Burgess PJ, Moreno G, Plieninger T (2016) Do European Agroforestry Systems Enhance Biodiversity and Ecosystem Services? A Meta-Analysis. Agr Ecosyst Environ 230(August):150–161. https://doi.org/10.1016/j.agee.2016.06.002
Vaccaro C, Six J, Schöb C (2022) Moderate Shading Did Not Affect Barley Yield in Temperate Silvoarable Agroforestry Systems. Agrofor Syst 96(4):799–810. https://doi.org/10.1007/s10457-022-00740-z
Vollenweider C, Spieß H (2018) ‘Composite Cross Populations: Legal Considerations and Their Value for Plant Breeding. In: Vere’. In , 49–50. Raumberg-Gumpenstein: Vereinigung der Pflanzenzüchter und Saatgutkaufleute Österreichs. https://www.dottenfelderhof.de/fileadmin/images/forschung/Veroeffentlichungen/Zuechtung/Gumpenstein_2017_19_Vollenweider.pdf
Vollenweider C, Haak A, Buhmann K, Locher M, Weyermann V, Schwittek G, Mascher F, Finckh M, Weedon O (2020) ‘Stability of Yield and Baking Quality Parameters of Heterogeneous Wheat Populations’. In 71st Conference. Breeding for Resilience against Biotic and Abiotic Stresses, edited by Vereinigung der Pflanzenzüchter und Saatgutkaufleute Österreichs. Online Conference, Raumberg-Gumpenstein, Österreich. https://www.dottenfelderhof.de/fileadmin/images/forschung/Veroeffentlichungen/Zuechtung/02_03-06_VOLLENWEIDER_v2.pdf
Zhang W, Xie H, Han S-A, Wang M, Ming-Qi Pan Xu, Qiao, and Long Li. (2022) Effect of Tree Form on Wheat Yield via Changing Microenvironment in Almond-Wheat Intercropping. Agrofor Syst 96(2):387–406. https://doi.org/10.1007/s10457-021-00726-3
Acknowledgements
We thank Karl Eggler and Jakob Bürgi von Aarburg for their permission and help to conduct the experiments on their fields. We are particularly grateful for Stef den Hond and his continuous physical and mental support throughout the experiment. We also thank Anita Vaccaro, Helga Palli, Maria Schubert-Kastner, Johannes Blacha, Levin Spiegel and Clemens Proksch for their help during harvest.
Funding
Open access funding provided by Swiss Federal Institute of Technology Zurich. This work was partially supported by the World Food System Center [Call 2 | 2021] and by core funding provided to JS from ETH Zurich.
Author information
Authors and Affiliations
Contributions
C.dH-V. designed the field trial with the help of C.S. and F.M. and carried out the field work and the post-harvest analyses. A part of the post-harvest analyses were also carried out by F.M.. C.dH-V. analysed the data and was thereby assisted by C.S.. C.dH-V. wrote the main manuscript. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
den Hond-Vaccaro, C., Mascher, F., Six, J. et al. Performance of a winter wheat composite cross population in two temperate agroforestry systems – a Swiss case study. Agroforest Syst (2024). https://doi.org/10.1007/s10457-024-00997-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10457-024-00997-6