
Abstract
Soil fertility depletion is a major constraint to agricultural production in the Congo Basin and is aggravated by climate change. Therefore, agroforestry systems (AFS) are used to improve soil fertility and crop productivity. Indeed, Pentaclethra macrophylla and Acacia auriculiformis are among the most agroforestry species used in this basin. However, smallholders are limited in their choice of agroforestry species and the spacing. Hence, this study comparatively evaluated the effects of P. macrophylla (native) and A. auriculiformis (exotic) legume tree species on soil macronutrients and food crops’ yield during four cropping seasons. To do this, a multifactor trial design was implemented in Lobilo catchment, Congo Basin, with two tree species, four tree planting densities (T1: 2500 trees × ha−1, T2: 625 trees × ha−1, T3: 278 trees × ha−1; and T0: crop monoculture), and three intercrops (cassava, maize, and peanut). The results revealed that both agroforestry species did not significantly differ regarding their impact on soil macronutrients. The total nitrogen and the pH in soil have increased with the cropping seasons. With regard to the food crops, cassava and maize yield were greater under P. macrophylla than under A. auriculiformis. However, the 2500 trees × ha−1 density negatively affected food crops’ yield and did not allow food crop production after the second cropping season. To maximize the positive effects of these AFS, it is essential to plant or co-plant these two agroforestry species at 625 trees × ha−1 density. Future research could investigate other native species and include water and light interactions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Land-use change in the Congo Basin is mainly driven by shifting agriculture in contrast to the commercial logging and plantation-driven deforestation in South America or South-East Asian (Wasseige et al. 2012; Curtis et al. 2018). The Congo Basin’s rapidly growing population still largely depends on shifting cultivation for food security and incomes (Bauters et al. 2021). This nexus of population growth and ecological impact are exacerbating the landscape degradation in this basin. This relates to the direct dependency of the local population on wood as an energy source and land clearing for food security and income generation. In fact, the shifting cultivation without the use of organic or inorganic fertilizers results in the decline of soil fertility, which is likely to affect primary soil macronutrients (Fonton et al. 2002; Bauters et al. 2021). Furthermore, ferralitic soils in the tropics are generally acidic and deficient in phosphorus (P) and nitrogen (N) (Roose et al. 1993). Thus, producing more food for a growing population without degrading the environment, and at the same time fighting poverty and hunger, seems a huge challenge for African agriculture (Verchot et al. 2007; Garrity et al. 2010).
On the other hand, AFS are recognized as sustainable production mode and nature-based solution for enhancing soils properties, mitigating the climate change effects and diversifying households income (Pretty 2008; Crous-Duran et al. 2019). Moreover, this production system has been known for its capacity to reconcile biodiversity conservation and enhancement of food production than conventional agriculture (Mbow et al. 2014). In addition, previous research had established that legumes are efficient in improving soil fertility by fixing atmospheric N2, which is one of the limiting production factors in the tropics (Fonton et al. 2002; Muyayabantu et al. 2019). However, numerous smallholder farmers in sub-Saharan Africa are constrained by the insufficient knowledge for the choice of agroforestry species and their spacing (Zebene & Agren 2007; Tokede et al. 2020; Katayi et al. 2023). Nowadays, choosing agroforestry species can’t be limited to its capacity to restore soil fertility, but must also take into account the production of wood, rate of carbon sequestration, as well as domestic utilities for local community (Lorenz and Lal 2014; Cardinael et al. 2015; Muyayabantu et al. 2019; Katayi et al. 2023). Despite the advantages of agroforestry species, tree spacing could significantly affect farmers’ adoption (Lékadou et al. 2009). Therefore, for early adoption, the understanding of tree-spacing effect on crop production, especially in the Congo Basin is needed (François et al. 2022).
In the Democratic Republic of the Congo (DRC), particularly in Yangambi landscape, some agroforestry schemes based on A. auriculiformis have been popularized in resolving the challenges described above and conservation of its exceptional resources (Schure et al. 2013; Peltier et al. 2010). While farmers are encouraged to adopt this exotic species for its rapid growth, they are often deliberately left with some useful species in their fields for livelihood diversification (Batsi et al. 2020; Katayi et al. 2023). However, the development agencies and the local community have different views on this exotic species. From the agencies’ view, this species meets the high demand for wood energy, while for farmers, it is not suitable for forest ecosystems and their livelihoods (Udawatta et al. 2017). Nevertheless, some farmers have already started to plant this species on their farms in association with crops such as cassava, maize, etc. in this landscape and Congo basin (Bisiaux et al. 2009).
However, smallholder farmers need to strengthen their technical capacities in managing the ecological interactions of this exotic species with food crops in this landscape. Indeed, the interactions of woody plant and food crop are complex in AFS; therefore, it requires the ability of farmers to maximize benefits interaction and upgrade the production, while the negative interactions are minimized (Nair et al. 1995; Kasongo and Van Ranst 2010). On other hand, there is a scientific deficiency of information on ecological interactions between native tree species as Pentaclethra macrophylla and food crops. In spite of the fact that they are better adapted to environmental conditions, research is needed to better understand their ecological interactions with other components of AFS (Nair et al. 1995; Dhanya et al. 2014). Indeed, local species are more likely to be adopted by farmers than exotic ones (Katayi et al. 2023).
Hence, to fill these knowledge gaps, this research was initiated to compare the effects of A. auriculiformis and P. macrophylla on soil macronutrients and yields of high consumption food crops (cassava, maize, and peanut). The research questions included: (a) which agroforestry species have more beneficial interaction with primary macronutrients and yield of maize, peanut, and cassava; and (b) which tree spacing enables smallholder farmers to produce more cassava, maize, and peanut after two years of intercropping?
Material and methods
Description of study area
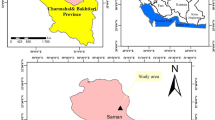
The Lobilo catchment (Fig. 1) is located 100 km from Kisangani town of Tshopo province situated in North-East DRC (Kipute et al. 2021). Its geographic location lies between 24°16′95″ and 25°08′48″ East longitude and 0°38′77″ and 1°10′20″ North latitude, with varying altitudes between 400 and 500 m (Kyale et al. 2019). The catchment is estimated at approximately 1213 km2 and characterized by a plateau area dissected by wide, flat-bottomed valleys, which are occupied by rivers, the main ones of which form tributaries of the Congo River (Kombele 2004). It contains the Yangambi Biosphere Reserve (YBR) that has a high faunal and floral diversity (Kyale et al. 2019).

Location of Lobilo catchment in Democratic Republic of Congo
This region is characterized by an average of 1837 mm of rainfall per year. Its climate is humid equatorial, (Af) according to Köppen's classification, and has high temperature regimes with an annual average of 25.1 °C (Luambua et al. 2021). This climate allows diversity of vegetation ranging from grasses to high canopy trees (Kombele 2004).
Research and experimental design
The experiment design was a multifactorial trial (Fig. 2) and composed of two blocks, (A. auriculiformis and P. macrophylla). For each block, the agroforestry species were combined with three food crops: cassava (Manihot esculenta var. Obama2); maize (Zea mays var. Samaru); and peanut (Arachis hypogea var. G17) replicated three times by food crop. Each replication had three tree-planting densities: 2500 (T1), 625 (T2), and 278 trees (T3) per ha−1, respectively, with T0 as control plot for crop monoculture. This multifactorial trial had a total of 72 plots (2 trees species × 3 food crops × 4 planting densities × 3 repetitions) and each plot size was 400 m2, 3.2 ha at total. It was implemented between March 2020 and March 2022 corresponding to four cropping seasons. For instance, in Yangambi landscape, there are two cropping seasons by cultural year: (1) Season A, which starts from March to June; and (2) Season B, which begins in September and ends in February (Mangaza et al. 2021).

Multifactorial trial design conducted in Yangambi experimental site
The food crops were planted at a spacing of 1 m × 1 m for cassava, 1 m × 0.5 m for maize, and 0.2 m × 0.2 m for peanut. Their vegetative cycle is annual – the maize and peanut being sown twice per year (from March to July, and from September to January), and cassava being planted only once a year. With regards to yield, grain yield was considered for maize and peanut, and root yield for cassava. The selection of agroforestry species was based on charcoal yield, rapid growth, and atmospheric N2 fixation, whereas the food crops selection was based on local food preferences. Also, precocity, disease resistance, and crop yield motivated the choice of varieties (Fonton et al. 2002; Lékadou et al. 2009).
Data collection
Soil parameters
The soil sample was collected using a composite sample per treatment. The composite sample was obtained after mixing the soil collected at four corners and center of each plot. A soil-sampling probe was used to collect up to 30 cm depth. Thus, these composite samples were collected before and after each cropping season for both years. Soil analysis was made following the methods presented by Faithfull (2002) at the Soil Laboratory of the University of Kisangani to assess the macronutrients (N, P, and K) and the pH, which mostly determine the soil fertility level and plant growth status through this catchment.
The N was determined by the Kjeldahl method that consisted of mineralization with sulfuric acid in the presence of a catalyst, followed by distillation by entrainment steam and titration with 0.05 N hydrochloric acid with ammonium (NH4+). The P was measured using an atomic absorption spectrophotometer after extraction via the Olsen method in a ratio of 1:10. The K was determined by atomic absorption spectrophotometer using Mehlich extract 3. The pH was measured with water in a ratio of 1:2 (weight/volume).
Crop yield
The gross yield was obtained after harvesting crops at physiological maturity. The observed area was harvested for each plot. After drying the harvest up to 14 percent of relative humidity (electronic moisture test), it was then weighted with a precision scale. The harvest was done between the first and third weeks of July and between first and third weeks of January (for maize and peanuts), and during the third week of February for cassava.
Data analysis
Mixed effect regression analysis was carried out to assess the link between soil elements and the treatment combinations, as well for the crops’ yield (Dodge 2008; Kuznetsova et al. 2017). Where needed, the Box-Cox data transformation was applied to fulfill criteria for regression and analysis of variance (ANOVA), applied on the transformed data, and back-transformed to normal for interpretation (Box and Cox 1964; Fox and Weisberg 2019). Hence:
For the soil macronutrients and pH, mixed effect regression model (Eq. 1) with the factors tree species, tree spacings, food crops, and cropping seasons (no interaction for the latter) were applied, as follows:
where \(Soi{l}_{element}=\{N, P,K or pH\}\), and \({\beta }_{n}\) are the regression coefficients.
For yield data, the annual crops were analyzed by using mixed effect regression model (Eq. 2) with the tree species, the tree spacing, and the cropping seasons (no interaction for the latter) as factors; then submitted to multifactor ANOVA to check the differences in means of treatments.
where \(i=\{\mathrm{cassava},\mathrm{ maize or peanut}\}\), and \({\beta }_{n}\) the regression coeffic0ients.
Table 1 presents the characteristics of explanatory variables used in the regression models.
The model parameters were estimated using the maximum likelihood method; only significant regression parameters are presented. The multi-model inference was used for model selection and for checking the accuracy (Kitamura et al. 2012). For the ANOVA, the estimated marginal means were computed for the significant sources of variation and treatments comparisons adjusted to identity homogeneous groups (Dodge 2008; Lenth 2021). All analyses and visualization was performed using R 4.2.1 version (R Core Team 2022).
Results
Soil macronutrients variation
Results showed that soil N varies with the cropping season. There was an increase of soil N rate in the third cropping season (0.02 ± 0.04 units, p = 0.043) and in the fourth cropping season (0.34 ± 0.04 units, p < 0.001), both compared to the first cropping season.
The P rate varied with cropping season, tree spacing, tree species, and food crops. This nutrient decreased by 1.76 ± 0.34 units (p < 0.001) for the third cropping season, while it increased by 4.18 ± 0.34 units (p < 0.001) for the fourth cropping season compared to the first one. The plots with tree-spacing of 2500 trees × ha−1 and 625 trees × ha−1 had an increase of soil P rate of 2.68 ± 0.92 units (p = 0.005) and 2.20 ± 0.92 units (p = 0.020), respectively, both compared to the monoculture plots. Moreover, for the plot with A. auriculiformis, P rate decreased by 4.09 ± 1.60 units (p = 0.014) and 3.06 ± 1.30 units (p = 0.02) at 2500 trees × ha−1 density and 625 trees × ha−1 density, respectively, compared to P. macrophylla. For the plots with maize intercropped with A. auriculiformis at 625 trees × ha−1 density, there was an increase of 3.51 ± 1.72 (p = 0.046) units in soil P rate compared to cassava cropping (Table 2).
There was a decrease of soil K rate in the fourth cropping season (0.15 ± 0.02 units, p = 1.6e-12) compared to the first cropping season. All other parameters were not significant.
The soil pH varied with cropping season and food crops. There was an increase of 0.35 ± 0.13 units (p = 0.035) in the soil pH rate for the fourth cropping season compared to the first one, as well as an increase in soil pH rate for maize intercropped plots (0.58 ± 0.33 units, p = 0.024) compared to the cassava ones.
Crop yield variation
Regression analysis results
Table 3 highlights that the cropping seasons and tree spacing influenced food crop yield.
The tree spacing negatively influences the cassava yields. Indeed, T1 (2500 trees × ha−1) planting density led to a decrease of 8.62 ± 3.59 t × ha−1 (p = 0.022) of cassava compared to monoculture plots. Meanwhile, the cropping season was a main factor that negatively affected the maize yield. This factor decreased maize yield by 0.24 ± 0.08 t × ha−1 (p = 0.004) for the second cropping season, 0.27 ± 0.09 t × ha−1 (p = 0.002) for the third one, and 0.92 ± 0.09 t × ha−1 (p < 0.001) for the fourth one, all compared to the first cropping season.
Peanut yield varied with the cropping season and tree spacing. We found that the density of 2500 trees × ha−1 (T1) decreased the peanut yield by 0.23 ± 0.10 t × ha−1 (p = 0.023) compared to monoculture. With regards to the cropping seasons, peanut yield decreased by 2.48 ± 0.07 t × ha−1 (p < 0.001) for the second cropping season, 2.37 ± 0.08 t × ha−1 (p < 0.001) for the third one, and 3.11 ± 0.08 t × ha−1 (p < 0.001) for the fourth, all compared to the first cropping season.
Means comparison for ANOVA significant sources of variation
There is a significant difference in cassava root yield when intercropping with the tree species (p-value = 0.043) as shown in the Fig. 3. Cassava root yield was greater when intercropped with P. macrophylla (25.7 ± 1.27 t × ha−1) than with A. auriculiformis (21.7 ± 21.7 t × ha−1). Neither the tree spacing nor cropping seasons brought any changes.

Cassava yield regarding the tree species (p = 0.043*)
The maize grain yield varied with the intercropped agroforestry species (p = 0.003) and cropping seasons (p < 0.001). Its yield in intercropping with P. macrophylla (1.30 ± 0.08 t × ha−1) was significantly greater than the one with A. auriculiformis (1.13 ± 0.09 t × ha−1). With regards to the cropping seasons, maize yield had significantly decreased for the last three seasons compared to the first one. However, maize yield did not significantly vary with tree spacing (Fig. 4).

Maize yield linked to agroforestry species (at left, p = 0.003 **) and cropping seasons (at right, p < 0.001***)
Peanut is intolerant to shading. Therefore, tree spacing and cropping seasons had significantly influenced its grain yield. Figure 5 highlighted interactions between these factors and the peanut yield. The shading created by the intercropped trees, as well as the nutrients taken by intercropping, may have significantly affected peanut yield. This could be because the monoculture plots had produced more (1.52 ± 0.06 t × ha−1) than the agroforestry plots (for both A. auriculiformis and P. macrophylla plots). In addition, the average peanut yield for T3 plots (1.33 ± 0.07 t × ha−1) was intermediary between the monoculture plots and other tree spacing (1.33 ± 0.06 t × ha−1 and 1.30 ± 0.06 t × ha−1), respectively, for T2 and T1.

Peanut yield regarding the tree density (p = 0.014*) and the cropping seasons (p < 0.001***)
The peanut yield obtained in the first cropping season (3.36 ± 0.06 t × ha−1) was higher than the ones in the second and third seasons (0.99 ± 0.07 and 0.88 ± 0.06 t × ha−1, respectively), and all of them higher than the peanut yield at the fourth season (0.25 ± 0.07 t × ha−1). In fact, agroforestry species seems more competitive for the peanut production after the first cropping season. In addition, this food crop requires rotation to avoid the N effect from legumes (peanut and agroforestry species).
Discussion of the results
Soil macronutrients variation
The rate of N, P, and K found in this experiment were below and at the edge of the recommended availability ranges. According to Wolf (2000), the quantity of primary macronutrients for agriculture are as follows: N (0.2–0.5%), P (30–100 ppm), and K (~ 120 ppm). However, after two years (Table 2), N values ranged between 0.14 and 0.21 percent for various spacing of P. macrophylla and 0.15 and 0.25 percent for various spacing of A. auriculiformis. Regarding P values, these ranged between 16.24 and 17.83 ppm for various spacing of P. macrophylla, and between 16.35 and 16.72 ppm for A. auriculiformis. For K values, these ranged between 1.19 and 1.12 ppm for P. macrophylla, and between 1.12 and 1.11 ppm for A. auriculiformis. These values proved that the macronutrient available in the catchment are below or at the edge of the recommended availability ranges. These results corroborate the findings of Makelele et al. (2022), which revealed that the flux of N is generally decreased in agricultural systems and young fallows. In addition, Li et al. (2019) had affirmed that tropical soils are deficit in soil macronutrients. The value of pH ranged between four and five, confirming the acidity of tropical soil (Roose et al. 1993).
Soil N increased with cropping seasons. This might be explained by the fact that these tree species are leguminous and are able to fix the N2 by the symbiosis with rhizobium (Bauters et al. 2016; Ishizuka et al. 2021). Indeed, their intercropping increases N rate in the soil. Additionally, leaf fall, litter decomposition, and nutrient mineralization in an agroforestry system can increase this macronutrient in soil. These results corroborate those obtained by Kasongo and Van Ranst (2010) who had found that A. auriculiformis had increased the N in sandy soil of Bateke plateau. They found that in fallows of four years, based on Acacia sp., the amounts of topsoil N increased up to 0.280 percent and were more than six times than those measured under the savannah (0.045%).
Soil P decreased in the third cropping season but increased in the fourth cropping season compared to the first one. This fluctuation can be explained by the P need of food crops and agroforestry species and the P supplying from the agroforestry species litter mineralization. Dollinger and Jose (2018) reported that the litter of Faidherbia albida could supply more than 18 kg P ha−1 × year−1. The P decreased more in plots with A. auriculiformis than plots with P. macrophylla, especially in the second cropping season. This might be linked to the fast-growing nature of A. auriculiformis that needs more P compared to P. macrophylla, which has a moderate growing rate (Ishizuka et al. 2021).
The maize absorbs 80–90 kg × ha−1of P (P2O5) over its cycle. However, the findings of this study revealed that P rate in soil was higher in maize plots compared to cassava ones. This might be explained by the fact that maize could not facilitate rapid litter decomposition than cassava due to the greatest cassava area coverage rate, providing enough shade and favorable conditions for litter mineralization. Else, Okalebo, and Woomer (2003) in Western Kenya found that cassava does need much P input for its development than maize.
The K did not vary with either the tree species nor the tree spacing or food crops. However, the pH increased with cropping seasons and under maize compared to cassava. This might be explained by the growing of trees over the fourth cropping season. During the first three seasons, tree litter was not important. However, after two years, agroforestry trees seem to gain important litter; thus, influence the increase of pH in the soil. Pinho et al. (2012) reported that the pH changes could be explained as a function of the concentration of the organic matter. Soil acidification can be attributed to degradation of organic matter in soils. Increased pH shows soil improvement by decomposition of tree litter.
Crop yield variation
Results revealed that T1 (2500 trees × ha−1) decreased cassava root yield compared to the monoculture plots. This can be explained by the high competition between these agroforestry species and cassava food crop. In fact, the cassava and trees were planted during the same period, but the competition for light led to the decreasing of cassava roots. Mboka (2021) also observed this phenomenon in hinterland of Kinshasa (DRC). Streck et al. (2014) found that cassava yield per plant and per root is higher at lower densities in Brazil. However, ANOVA tests results revealed that cassava yielded more when intercropped with P. macrophylla than with A. auriculiformis. In fact, A. auriculiformis grows faster than P. macrophylla; hence, may be taking more resources and in strong competition with the cassava (Fonton et al. 2002; Lékadou et al. 2009).
The main factor affecting maize yield was the cropping seasons. There was a general decrease of maize yield from the first season up to the fourth. This decrease was significant from the second cropping season and onward compared to the first one. This supports the agronomic theory that successive crop repetition on the same soil leads to a continuous decline in physical yield (Giller 2009). Although agroforestry tree-species perform well, crop rotation remains essential to maintain gross yield. In fact, Bertomeu (2012) also found out that in the Philippines, maize yield, in association with Gmelina arborea (a fast-growing species), decreased up to 50 percent in the second cropping season.
Peanut yield is negatively affected by cropping seasons and tree spacing. Comparative analysis highlights the yield decrease between the first and second cropping seasons up to 70 percent of production. This can also be explained by the successive crop repetition on the same soil. These results corroborate with those obtained by Wang et al. (2021) who showed that peanut yield was affected by year and biochar application factors. Moreover, ANOVA revealed that peanut yield under P. macrophylla is greater compared to A. auriculiformis. Peanut demands a lot of sunlight (Li et al. 2018); hence the yield of peanut was negatively affected by the density of 2500 trees × ha−1. This situation is explained by the shading created by trees when they are growing (Van Noordwijk and Luisiana 2004). In line of this study, Rowell et al. (1999) affirmed that the light factor influences foliage, pod, and seed yield of peanut during his experimentation in Georgia.
Conclusion
To sustain food security for a growing population without significantly degrading the environment, AFS are reputed to improve soil fertility and crop productivity and they provide ecological services. However, household famers are uninformed about the interactions of agroforestry species (exotic or native species) and the effectiveness of spacing to practice. Therefore, this research addresses the influence of P. macrophylla and A. auriculiformis on food crop (cassava, maize and peanut) production in the Congo Basin to respond to these gaps.
Results revealed that both agroforestry-species did not significantly differ regarding their effects on soil nutrients. Their total N and the pH increased over the growing of cropping season. With regards to the food crops’ yield variation, the yields of cassava and maize were greater under P. macrophylla than under A. auriculiformis. Furthermore, the tree spacing of 2500 tress × ha−1 of these two tree species negatively affect food crops’ yield.
Hence, to maximize the positive effect of these system, it is essential to plant or co-plant these two tree species at 625 trees × ha-1 density. Furthermore, research could assess the general context of the system, including water and light interactions and explore other native species like Milletia sp. Thus, the popularization of these agroforestry species could be adopted in Lobilo catchment for promoting the livelihood of the riparian communities, maximizing agroecosystem services, and biodiversity conservation through the reduction of deforestation rate in Congo Basin.
References
Batsi G, Sonwa DJ, Mangaza L et al (2020) Biodiversity of the cocoa agroforests of the Bengamisa-Yangambi forest landscape in the Democratic Republic of the Congo (DRC). Forests. https://doi.org/10.3390/f11101096
Bauters M, Mapenzi N, Kearsley E et al (2016) Facultative nitrogen fixation by legumes in the central Congo basin is downregulated during late successional stages. Biotropica. https://doi.org/10.1111/btp.12312
Bauters M, Moonen P, Summerauer L et al (2021) Soil nutrient depletion and tree functional composition shift following repeated clearing in secondary forests of the Congo Basin. Ecosystems. https://doi.org/10.1007/s10021-020-00593-6
Bertomeu M (2012) Growth and yield of maize and timber trees in smallholder agroforestry systems in Claveria, Northern Mindanao, Philippines. Agrofor Syst. https://doi.org/10.1007/s10457-011-9444-x
Bisiaux F, Peltier R, Muliele J-C (2009) Plantations industrielles et agroforesterie au service des populations des plateaux Batéké, Mampu, en République Démocratique du Congo. Bois & Forets des Tropiques 301(301):21. https://doi.org/10.19182/bft2009.301.a20404
Box GEP, Cox DR (1964) An analysis of transformations. J R Stat Soc Ser B 26:211–243
Cardinael R, Chevallier T, Barthès BG et al (2015) Impact of alley cropping agroforestry on stocks, forms, and spatial distribution of soil organic carbon: a case study in a Mediterranean context. Geoderma. https://doi.org/10.1016/j.geoderma.2015.06.015
CIFOR (2018) Agro-ecology and agroforestry for sustainable forest management
Crous-Duran J, Graves AR, Paulo JA et al (2019) Modelling tree density effects on provisioning ecosystem services in Europe. Agroforest Syst 93:1985–2007. https://doi.org/10.1007/s10457-018-0297-4
Curtis PG, Slay CM, Harris NL et al (2018) Classifying drivers of global forest loss. Science 80:361. https://doi.org/10.1126/science.aau3445
Dhanya B, Sathish BN, Viswanath S et al (2014) Ecosystem services of native trees: experiences from two traditional agroforestry systems in Karnataka Southern India. Int J Biodiv Sci Ecosyst Serv Manage 10(2):101–111. https://doi.org/10.1080/21513732.2014.918057
Dodge Y (2008) The concise encyclopedia of statistics.
Dollinger J, Jose S (2018) Agroforestry for soil health. Agrofor Syst 92:213–219
Faithfull NT (2002) Methods in agricultural chemical analysis. A Practical Handbook, Cabi
Fonton NH, Kakai RG, Rondeux J (2002) Étude dendrométrique d’Acacia auriculiformis A. Cunn. ex Benth. en mélange sur vertisol au Bénin. Biotechnol Agron Soc Environ 6
Fox J, Weisberg S (2019) An R companion to applied regression, 3rd edn. Sage, Thousand Oaks
François M, Merveillia-Gilfarde MK, Suspense IA (2022) Dynamique du carbone organique du sol et de l’azote dans une chronoséquence de plantation de Acacia auriculiformis A Cunn ex Benth (Fabaceae), à Bambou-Mingali (République du Congo). Eur Sci J ESJ. https://doi.org/10.19044/esj.2022.v18n8p172
Garrity DP, Akinnifesi FK, Ajayi OC et al (2010) Evergreen Agriculture: A robust approach to sustainable food security in Africa. Food Secur. https://doi.org/10.1007/s12571-010-0070-7
Giller KE (2009) Putting nitrogen fixation to work for smallholder farmers in Africa. N2 Africa project document
Ishizuka S, Mori T, Nakayama Y et al (2021) Effects of conversion from leguminous acacia to non-leguminous eucalyptus on soil N2O emissions in tropical monoculture plantations. For Ecol Manage. https://doi.org/10.1016/j.foreco.2020.118702
Kasongo RK, Van Ranst E (2010) Improving the quality of sandy soils of the Batéké plateau (DR Congo) by using local geological materials and industrial organic waste products. Afrika Focus. https://doi.org/10.21825/af.v23i1.18010
Katayi LA, Kafuti C, Kipute DD et al (2023) Factors inciting agroforestry adoption based on trees outside forest in Biosphere Reserve of Yangambi landscape (Democratic Republic of the Congo) in Agrofores. Syst. https://doi.org/10.1007/s10457-023-00854-y
Kipute DD, Mampeta SW, Kahindo J-MM et al (2021) Complexity of zoning management in biosphere reserves: the case of the Yangambi Biosphere Reserve in DRC. Int for Rev 23(1):55–67. https://doi.org/10.1505/146554821832140358
Kitamura J, Nagata N, Nakajima J, Sota T, Bartoń K (2012) MuMIn: Multi-model inference. R package version 1.47.1. 2022. J Evol Biol 25
Kombele FBM (2004) Diagnostic de la fertilité des sols dans la cuvette centrale congolaise. Cas des séries Yangambi et Yakonde (Thèse de doctorat). Université de Liège–Gembloux Agro-Bio Tech
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package: tests in linear mixed effects models. J Stat Softw. https://doi.org/10.18637/JSS.V082.I13
Kyale JK, Maindo AMN, Wardell AD (2019) Réserve de Biosphère de Yangambi à l’épreuve de la cristallisation des pratiques locales de survie : une réponse à la faillite de l’État en République Démocratique du Congo. VertigO. https://doi.org/10.4000/vertigo.24677
Lékadou TT, N’guessan A, Konan JL, et al (2009) Effet de la densite sur les parametres dendrometriques D’Acacia auriculaeformis en association avec le cocotier (Cocos nucifera L.) sur sables quaternaires en Côte d’Ivoire. Agron Africaine 21
Lenth RV (2021) Emmeans: estimated marginal means, aka least-squares means. R packageversion 1.7.1. R Found Stat Comput 34:216–221
Li C, Liu D, Li L et al (2018) Effects of light-emitting diodes on the growth of peanut plants. Agron J 110:2369–2377. https://doi.org/10.2134/agronj2017.11.0674
Li S, Gong Q, Yang S (2019) Analysis of the agricultural economy and agricultural pollution using the decoupling index in Chengdu, China. Int J Environ Res Public Health. https://doi.org/10.3390/ijerph16214233
Lorenz K, Lal R (2014) Soil organic carbon sequestration in agroforestry systems: a review. Agron Sustain Dev 34:443–454
Luambua NK, Hubau W, Salako KV et al (2021) Spatial patterns of light-demanding tree species in the Yangambi rainforest (Democratic Republic of Congo). Ecol Evolut. 11:18691–18707. https://doi.org/10.1002/ece3.8443
Makelele IA, Bauters M, Verheyen K et al (2022) Conservative N cycling despite high atmospheric deposition in early successional African tropical lowland forests. Plant Soil. https://doi.org/10.1007/s11104-022-05473-7
Mangaza L, Sonwa DJ, Batsi G et al (2021) Building a framework towards climate-smart agriculture in the Yangambi landscape Democratic Republic of Congo (DRC). Int J Clim Change Strat Manage. https://doi.org/10.1108/IJCCSM-08-2020-0084
Mboka IJ-C (2021) Culture du manioc au Congo-Kinshasa. Anthropol Sociétés 44:91–108. https://doi.org/10.7202/1078166ar
Mbow C, Smith P, Skole D et al (2014) Achieving mitigation and adaptation to climate change through sustainable agroforestry practices in Africa. Curr Opin Environ Sustain 6:8–14
Muyayabantu GM, Kabwe NM, Mutombo JM et al (2019) Effect of the Form and Nature of Nitrogen on growth and nodulation of pigeon pea (Cajanus cajan (L.) Millsp) on sandy soil of Mont-Amba, in Democratic Republic of Congo. Am J Plant Sci 10(08):1457. https://doi.org/10.4236/ajps.2019.108103
Nair PKR, Kang BT, Kass DCL (1995) Nutrient cycling and soil-erosion control in agroforestry systems. Agric Environ Bridg Food Prod Environ Prot Dev Ctries 60:117–138
Okalebo JR, Woomer PL (2003) Organic resources for integrated nutrient management in western Kenya. In: Organic resource management in Kenya: perspectives and guidelines. Nairobi, Kenya
Peltier R, Bisiaux F, Dubiez E, et al (2010) De la culture itinérante sur brûlis aux jachères enrichies productrices de charbon de bois, en République Démocratique du Congo
Pinho RC, Miller RP, Alfaia SS (2012) Agroforestry and the improvement of soil fertility: a view from amazonia. Appl Environ Soil Sci
Pretty J (2008) Agricultural sustainability: concepts, principles and evidence. Philos Trans R Soc B Biol Sci 363:447–465
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. R Found Stat Comput
Roose E, Ndayizigiye F, Sekayange L (1993) L’agroforesterie et la GCES au Rwanda. Comment restaurer la productivite des terres acides dans une region tropicale de montagne a forte densite de population? Cah - ORSTOM, Ser Pedol 28
Rowell T, Mortley DG, Loretan PA et al (1999) Continuous daily light period and temperature influence peanut yield in nutrient film technique. Crop Sci. https://doi.org/10.2135/cropsci1999.0011183X003900040026x
Schure J, Ingram V, Assembe-Mvondo S, et al (2013) La filière bois énergie des villes de Kinshasa et Kisangani, eds Marien N. et al. Quand la Ville mange la forêt pp 27–44
Streck NA, Pinheiro DG, Zanon AJ et al (2014) Effect of plant spacing on growth, development and yield of cassava in a subtropical environment. Bragantia. https://doi.org/10.1590/1678-4499.0159
Tokede AM, Banjo AA, Ahmad AO, Fatoki OA, Akanni OF (2020) Farmers’ knowledge and attitude towards the adoption of agroforestry practices in Akinyele Local Government Area, Ibadan, Nigeria. J Appl Sci Environ Manage 24(10):1775–1780. https://doi.org/10.4314/jasem.v24i10.10
Udawatta RP, Gantzer CJ, Jose S (2017) Agroforestry practices and soil ecosystem services. In: Soil health and intensification of agroecosystems
Van Noordwijk M, Luisiana B (2004) WaNuLCAS version 3.1. Backgr a Model water, Nutr Light capture Agrofor Syst ICRAF, Bogor, Indonesia
Verchot L, Van Noordwijk M, Kandji S et al (2007) Climate change: Linking adaptation and mitigation through agroforestry. Mitig Adapt Strateg Glob Chang. https://doi.org/10.1007/s11027-007-9105-6
Wang S, Zuo Q, Cao Q et al (2021) Acceleration of soil N2O flux and nitrogen transformation during tropical secondary forest succession after slash-and-burn agriculture. Soil Tillage Res 208:104868
Wasseige C, Marchen P, Bayol N, et al (2012). Les Forets du bassin du Congo: Etat des Forets 2010
Wolf B (2000) The fertile triangle: the interrelationship of air, water, and nutrients in maximizing soil productivity. Food Products Press, New York
Zebene A, Ågren GI (2007) Farmers’ local knowledge and topsoil properties of agroforestry practices in Sidama, Southern Ethiopia. Agroforest Syst 71:35–48
Acknowledgements
The work leading to this publication has been funded by the European Union's FORETS (Formation, Recherche et Environnement dans la Tshopo) project, implemented by the Center for International Forestry Research (CIFOR). The authors are grateful to the Institut National pour l'Etude et la Recherche Agronomique (INERA) for allowing this research to be conducted on their premises at the Yangambi Research Station, and to all the FORETS project staff for providing a very valuable support. We would like to thank all those who were involved in one way or another in bringing this research to fruition. All usual caveats apply.
Funding
We, the authors, declare that we have no competing financial interests or personal relationships that could have prejudiced the results of the work presented in this paper. In addition, we also declare that this article or any of its findings have never been submitted to any journal and is completely undisputed.
Author information
Authors and Affiliations
Contributions
The main author did the manuscript, with the contribution of the authors for improvement. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This research was carried out in strict compliance of the agronomic norms of experimental design (i.e., orientation of design, no burning of the field, chronogram of crop care activities, and non-use of chemical products). Socially, this research did not involve child labor and was gender-inclusive in its implementation.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mapenzi, N., Katayi, A.L., Bauters, M. et al. Improved crop productivity and soil properties under varying planting densities of Pentaclethra macrophylla Benth. and Acacia auriculiformis A. Cunn. in Congo Basin. Agroforest Syst 98, 295–307 (2024). https://doi.org/10.1007/s10457-023-00908-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-023-00908-1