
Abstract
The transition from conventional arable towards silvoarable systems can increase the delivery of ecosystem services. Nevertheless, the assessment of crop yield under agroforestry condition is crucial to evaluate of the reliability of these systems and to increase the knowledge base needed to support their design. Although the feasibility of poplar short rotation coppice (SRC) silvoarable alley-cropping systems has been widely investigated, few studies have addressed the agronomic response of crops intercropped with poplar SRC in narrow alleys, especially in Mediterranean environments. Thus, this paper treats the effects of SRC poplar rows on soybean and sorghum productivity in a 2-year rotation implemented in an alley-cropping system. A field experiment was carried out in 2018 and 2019 with the objective of measuring and evaluating effects of light availability variation, as affected by the growing rate of 2-year coppice cycle poplar SRC rows, and the soil characteristics on soybean-sorghum and sorghum–soybean rotations. Above-ground biomass, grain yield and crops residue showed a significant reduction in the tree–crop interface up to 74% and sorghum proved to be less tolerant to light reduction compared to soybean. Our results demonstrated that light is the most important factor for sorghum cultivation, despite grain yield was also influenced by the soil characteristics such as pH, while soybean is affected also by soil moisture and water retention capacity. The design of crop rotation in an SRC-based agroforestry system needs to consider the different agronomic performance of different crops and the harvest cycle of tree rows.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The development of agroforestry systems has been shown to be an appropriate strategy to improve the potential ecosystem services delivered by farming systems and to tackle the changing climate (Burgess and Rosati 2018; Kay et al. 2019; Paris et al. 2019). Silvoarable cropping systems can be more effective in generating ecosystem services compared to conventional arable systems (Palma et al. 2007). The sustainability of these systems relies on the following key aspects: combining food, feed, and timber production on the same land surface, stocking carbon in the above- and below-ground biomass of trees, reducing soil erosion soil and flooding risks, and improving nutrient cycling (Tsonkova et al. 2012; Torralba et al. 2016). By contrast, some negative concerns of agroforestry could arise from management and socio-economic issues, due to increased labor, complexity of work, management costs and administrative burden. However, stakeholders usually agree on the positive effect of productions diversification on additional and diversified income opportunities (García et al. 2018).
In Europe, poplar tree plantations managed as short rotation coppice (SRC) showed a high biomass potential for energy use (Nassi et al. 2010) but farmers have shown scarce interest in poplar SRC pure stands, due to low market opportunities for a biomass feedstock characterized by a lower calorific value, higher ash content and logistic problems compared to other woody sources (Elghali et al. 2007). However, the recent European Commission’s decision to include agroforestry as a practice for carbon farming and other eco-schemes related with climate change mitigation and adaptation, prevention of land degradation and protection of biodiversity (European Commission 2021) makes SRC suited for agroforestry-related subsidies. Thus, the conversion from SRC plantation to silvoarable alley cropping-systems could become an interesting option for the transition towards sustainable agroforestry systems. Investigating the feasibility of silvoarable alley-cropping systems based on poplar short rotation coppice (SRC) has highlighted: (i) lower wood biomass production per hectare than pure poplar SRC stands, due to the reduced tree density, despite a higher growth rate per plant; (ii) tree rows significantly affect crop performance, competing for water, nutrients, and light, especially in the tree-crop interface zone (Swieter et al. 2021); (iii) tree arrangement and orientation, as well as the land ratio between trees and herbaceous crops (Lamerre et al. 2015; Seserman et al. 2019), are key factors for the sustainability of silvoarable systems (Tsonkova et al. 2012). In alley-cropping system, the choice of herbaceous crop is crucial for exploiting facilitations and limiting competition of trees. Literature highlighted that at middle latitudes winter crops would be preferable in association with deciduous trees to prevent competition for radiation use from the tree canopy (Lovell et al. 2018) and woody elements such as branches and trunk and can affect crop yield (Talbot and Dupraz 2011). Besides, warm-season crops could benefit from the reduction of potential evapotranspiration, exploiting the windbreaking function of tree rows, especial in rainfed systems (Kanzler et al. 2019).
In previous studies, authors have demonstrated the suitability of the intercropping of both soybean and sorghum with poplar SRC in narrow alleys in rainfed Mediterranean conditions (Mantino et al. 2020; Pecchioni et al. 2020). In the present study, authors investigated the feasibility of soybean and sorghum cultivation in a poplar SRC agroforestry system exploring the relationship between of crop rotation and tree biomass cutting cycle. Other studies have highlighted that, in both temperate and tropical environments, C3 species seem to be more adaptable to the shading effect of trees in agroforestry systems than C4 species (Reynolds et al. 2007; Magalhães et al. 2019; Ogwok et al. 2019). However, few studies have been conducted in Mediterranean environments aimed at assessing the agronomic response of a 2-year rotation including soybean and sorghum intercropped with poplar SRC in narrow alleys.
The hypothesis of the present study is that the effect of tree presence on herbaceous plant in a poplar SRC agroforestry system depends on the combination of tree cutting cycle and crop species. To fill the gap in practical and scientific knowledge, needed for the correct design and implementation of diversified and complex cropping systems such as agroforestry, the aim of present study was to investigate the effects of SRC poplar rows on soybean and sorghum agronomic performance in a 2-year rotation in an alley-cropping system.
Materials and methods
Site description and experimental design
A 2-year field trial was carried out at the Centre for Agro-Environmental Research of the University of Pisa (CiRAA) "E. Avanzi", San Piero a Grado, Pisa, Italy (43° 40′ 49″ N, 10° 20′ 47″ E; 1 m above sea level and 0% slope). The average climate conditions of the site are characterized by an annual rainfall of 920 mm and an annual mean temperature of 15 °C (long term average 1987–2017).
The study was conducted in an alley-cropping experimental field based on poplar SRC rows NW–SE oriented (357° N-NW) and a 2-year rotation including each year soybean (Glycine max (L.) Merr. cv. Zora) and sorghum (Sorghum bicolor L. Moench cv. Baggio). The experimental field originated from a 1.2 ha former poplar SRC plantation (Populus ×canadensis Moench. clone AF2 and Populus × generosa Henry × nigra L. clone Monviso) established in 2009. From 2009 to 2017, the poplar trees were coppiced every 2 years (2011, 2013, 2015, and 2017) with a cut-and-chip harvester at the end of winter. During the present study, the poplar shoots were harvested in March 2019 between the two cropping seasons. The characteristics of poplar SRC rows, summarized in Table 1, were measured at the end of winter, before sprouting or before cutting, in 2017, 2018 and 2019.
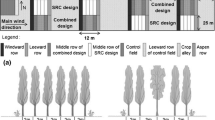
Arable alleys were created after the harvest in March 2017, by removing four out of five rows, thus leaving a 12 m wide net workable space, corresponding to a gross distance between tree rows of 13.5 m. Land clearing was carried out with a forestry shredder followed by disk harrowing at 20 cm depth. During summer 2017, chemical weeding was carried out to kill poplar shoots resprouted from surviving roots. Both soybean and sorghum, were sown in 2018 and 2019 in four plots (12 m wide and 30 m long) for each crop (total number of plots per year was equal to eight). Then, two rotations, sorghum-soybean and soybean-sorghum took place for 2 years, in 2018 and 2019, while 2 years old poplar shoot was harvested in March 2019, between the two cropping seasons.
Soil data collection, soil characteristics and plot selection
In September 2017, along the length of the experimental field, fifty soil cores sampled randomly at two different depths, 0–10 cm and 10–30 cm, were analyzed for: soil texture (international pipette method), pH (H2O, 1:2.5), organic matter content (Walkley–Black method), total nitrogen (N) content (Kjeldhal method), available phosphorus (P) (Olsen method) and exchangeable potassium (K) (BaCl2 method) (Motsara and Roy 2015). Soil samples were collected in all the experimental site, to obtain spatialized soil characteristic maps. Thus, discrete data points from soil analyses were interpolated according to the ordinary kriging method implemented in the R package “gstat” (Pebesma 2004). The “automap” (Hiemstra et al. 2009) package was performed to validate the raster layers from the ordinary kriging models. The “zonal statistic” function of QGIS (v. 3.12) was used to calculate the mean and standard deviation of the soil parameters of each investigated plot from the raster layers of soil characteristics (Table 2).
The soil parameter values of the plots are reported in Table 1. The validation results of the ordinary kriging models for each analyzed soil characteristic were reported in a previous study (Mantino et al. 2020). The assessment of soil characteristics highlighted a high spatial variability along the experimental site probably due to the soil genesis of the Pisa coastal plain originating from the deposition of particles from periodic overflows of the river Arno and by land reclamation carried out during the Medici age (fifteenth century). As a consequence, the presence of sand, silt, and clay in the soil varied among plots with a coefficient of variation of 0.75, 0.31, and 0.22, respectively (Table 2). Likewise, the presence of available phosphorus and exchangeable potassium in the soil varied according to the texture, with an increasing trend positively correlated with the sand content. Conversely, soil organic matter, pH and total N values showed a lower variability among the experimental plots (Table 2).
Eight plots (four per each crop) were selected in both silty-clay-loam and loam to sandy-loam soil areas, to take into account the texture as a source of variability. From the first year of the experiment, two plots per soil type were assigned to sorghum and soybean. N, P, and K fertilization was applied before crop sowing in both years of the study in such a way to reduce the limiting effect due to nutrient availability.
Crop management and data collection
The seedbed preparation in 2018 and 2019, for both soybean and sorghum, consisted in chisel ploughing (0.5 m depth) and disk harrowing and in 2019 crop residues were incorporated into the soil. In 2018, wet conditions in spring delayed sowing date that was carried out in June while in 2019 was carried out in April.
A ternary compound fertilizer was applied in both years at rates of 32 N, 96 P2O5, and 96 K2O kg ha−1 before soybean sowing. On 12 June 2018, soybean inoculated with Bradyrhizobium japonicum (coated seed) was sown with a seeding rate of 50 seeds m−2. The following year was sown on 29 April 2019. In both experimental years, 40 days after emergence root nodules were absent, thus two consecutive nitrogen applications of 92 kg N ha−1 were carried out on 13 July 2018 before the inter-row hoeing and on 21 August 2018 at the first visible pods (69 BBCH phenology scale). In 2019, on soybean a single nitrogen application was carried out at a rate of 92 kg N ha−1 on 9 July 2019 before the inter-row hoeing. N fertilization was reduced compared to the previous year due to a high quantity of residual N. Authors hypothesis about the absence of inoculation is that the use of coated seed with Bradyrhizobium japonicum was not the optimal strategy in a field with no soybean cultivation in the last 10 years. In fact, in 2020 (year out of this study) the soybean inoculated with a handmade liquid inoculum was rich in root nodules in all the agroforestry plots. Moreover, authors did not observe any correlation between inoculation rate and distance from trees. Soybean was harvested on 8 October and 24 September, in 2018 and 2019 respectively.
Regarding sorghum, a ternary compound fertilizer was applied in both years at rates of 32 N, 96 P2O5, and 96 K2O kg ha−1 before sowing. On 6 June 2018, sorghum was sown with a seeding rate of 40 seeds m−2 and N was applied before the inter-row hoeing with a rotary cultivator at a rate of 184 kg N ha−1 on 10 July. The following year sorghum was sown on 29 April and N fertilization was performed at a rate of 92 kg N ha−1 on 9 July. As for the soybean, N fertilization in the second year was reduced due to a high level of residual N from the previous year. Sorghum was harvested on 4 October and 24 September, in 2018 and 2019 respectively.
On 26 June 2019, one overhead irrigation of 30 mm was performed in all experimental plots using a travelling sprinkler to help crop growth due to the lack of rainfall during June (see Fig. 2).
Sorghum and soybean were sampled at harvest to measure the above ground biomass production and its partitioning between grain and residue. The sampling points were positioned along a transect replicated three times per plot spaced 10 m. In each transect, the sampling was carried out on five points at fixed distances from the tree row westward to eastward, in positions referred to as West (2.5 m distance from the tree row), Mid-west (4.5 m), Centre (6.75 m), Mid-east (4.5 m) and East (2.5 m), for a total of 15 samples per plot (overall 60 samples per crop per year). Soybean and sorghum above-ground biomass samples were collected by cutting the plants at ground layer harvesting on a 0.5 m2 surface (Fig. 1). Crop production in the two experiment years was measured as follows: total above-ground biomass dry weight (g m−2), grain dry weight (g m−2), and residue dry weight (total dry weight minus grain dry weight) (g m−2). Samples were fresh weighed and sub-samples were oven dried at 60 °C for dry matter content determination.

Experimental field and representation of main cultivation phases of trees and crops in the experimental field starting from the conversion from short rotation coppice to alley-cropping systems. The pictures provide a schematic view of a plot during the month of July from 2016 to 2019. W means, MW means Mid-west, C means Centre, ME means Mid-east, E East sampling point position
Light availability data collection
The light availability (LAV) at the herbaceous layer was measured by means of hemispherical photos. Photos were taken at dawn or sunset, to prevent the disturbance of direct sunlight. Photos were taken with a digital camera equipped with a 180° fish-eye lens (Nikon, Coolpix 4500 with FC-E8 lens, Minato, Tokyo, Japan) at a fixed distance of 1 m from the ground (Frazer et al. 1999) and when necessary photos were taken above the herbaceous layer. The LAV measurement scheme was arranged in five points replicated in three transects in each plot, following the same layout described above, for soybean and sorghum (Fig. 1). LAV measurements were performed on 3 days in 2018 and 2 days in 2019, at different crop development stages. The first LAV measurement round was performed at sowing (16 June 2018 and 24 June 2019, for both crops). The second took place during soybean and sorghum vegetative growth (26 July 2018 and 25 July 2019). In 2018, the third round was carried out at the end of the growing season, before crop harvesting (2 October). LAV was obtained by processing the images with Gap Light Analyzer 2.0 GLA (Frazer et al. 1999).
Meteorological conditions
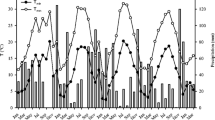
Meteorological data was collected by a public weather station (43°38′16.8"N 10°23′27.6"E) of the Tuscany Region (http://www.sir.toscana.it). Figure 2 shows the meteorological conditions during the study period and the graphs highlight the dry months, when precipitation is equal to or less than twice of the monthly mean air temperature (P ≤ 2 T) (Bagnouls and Gaussen 1957). In the 2 years of the experiment, from May to October, the rainfall patterns differed greatly between years. In May 2018, a high precipitation was recorded (91 mm, + 36% compared to the long term), forcing a delay in sowing to early June (Fig. 2). In April 2019, an unusual amount of rainfall was recorded (101 mm, + 25% higher than the long-term average). In 2018, between late June and late July, 4 days with over 6 mm of rain occurred for a total of 38 mm of rain, favoring the growth conditions during the vegetative stages of the two crops. Conversely, in August and September, only two rainy days occurred, with precipitations lower than the long-term average (− 53% in August and − 79% in September).

The graphs show the monthly values of average air temperature in the study period (solid line), the monthly rainfall (vertical white bars) and monthly evapotranspiration (vertical grey bars)
The crops, in 2019, received 101 mm of rainfall in May in the early development stages, distributed over 12 rainy days. In 2019, after the heavy rains in May, June and August were characterized by just 4 and 5 mm of rain respectively, in a single rainy day for each month. Due to the lack of rain, on 26 June 2019 an overhead irrigation was performed. Unusually, rainfall occurred in July, with 94 mm of precipitation in 4 days, when the monthly long-term average is just 25 mm.
The mean temperatures between June and September 2018 were, on average, 1.2 °C higher compared to the long term. In 2019, between May and September, the daily mean temperature was 0.8 °C higher than the long term, with the maximum temperatures very close to the long term (+ 0.1 °C in 2019 compared to the long term). Conversely, the minimum temperatures were 1.9 °C higher than the long term. Crop water stress, calculated as the difference between evapotranspiration (Hargreaves–Samani equation) and precipitation, was similar in the 2 years of the study, and 369 and 396 mm in 2018 and 2019, respectively (Fig. 2), showing a low water availability during the cropping period.
Statistical analysis
Statistical analysis was carried out in R (R Core Team 2015). LAV at herbaceous layer (%), above-ground biomass (g m−2), grain yield (g m−2), plant residue (g m−2), plant density at harvest (plant m−2), above-ground biomass per plant (g plant−2), and grain yield per plant of soybean and sorghum (g m−2) were analyzed to assess the effects of the position in the alley and the year of cultivation. Detection and removal of outliers were carried out with the Bonferroni outlier test using the “car” package (Fox and Weisberg 2019) with the outlierTest function. Levene’s test was used to check the homogeneity of variance and the Shapiro–Wilk test was used to check the normality of the residuals. Fischer z-transformations were performed on LAV data to respect the normal distribution of residuals. Data transformation was not performed on crop data.
The effects of the position in the alley and year of cultivation were evaluated for each crop by the lmer function for linear mixed-effect models, in the “lme4” package (Bates et al. 2015), with the factors “position” (n = 5) and “year” (n = 2) as fixed effects and “plot” (n = 4) and “transect” (n = 3) as random nested effects (plot/transect). Tukey’s honestly significant difference (HSD) post-hoc test was carried out by pairwise multiple comparisons using the “emmeans” package (Lenth et al. 2021) with the emmeans function. A regression tree analysis was also performed, using the “party” package (Hothorn et al. 2006) and the ctree function, to assess the multiple effects of tree distance, light availability, and soil characteristics (clay presence, sand presence, pH and organic matter content) on soybean and sorghum agronomic performance. A Bonferroni test with p-value equal to 0.05 was used as tree split criterion (Hothorn et al. 2006).
Results and discussion
Variation of light availability in the alley
Statistical analysis showed that during the soybean and sorghum cropping period, the average LAV at the herbaceous layer was significantly affected by the position in the alley and year of cultivation, and the first order interaction (position per year) was also significant (Table 3).
LAV at the herbaceous layer significantly varied according to the distance from trees and tree canopy size, as effect of the shoot age (Tables 1 and 4), showing the same fashion both in the soybean and sorghum plots. This result shows that the poplar rows had a similar effect on light availability among the crop plots due to the homogenous growth of the tree rows in the experimental field. On average, the LAV was lower in 2018 compared to 2019 due to the higher canopy of 2-year old poplar-SRC stems (Table 4). This was because the poplar harvest occurred in March 2019, allowing a higher light availability at the herbaceous layer during the second cropping period. A similar effect was recorded by Dufour et al. (2020) after the pollarding of poplar trees in an alley-cropping system in France.
The orientation of the poplar rows determined a significantly lower light availability especially in the tree-crop interface, in both West and East sides, compared with the center of the alley and the intermediate positions (Mid-west and Mid-east). However, during the 2 years different LAV was observed between East and West side, with an interaction among year and position. In fact, in 2018, the lowest light availability was recorded in the West side of the alley while in 2019 the lowest data was recorded in the East side. Previous studies carried out on single scattered trees showed that the hemispherical photo method should not lead to different estimations of LAV between East and West orientation (Montero et al. 2008). Thus, we suppose that the observed difference between the East and West side might be due to the azimuth of the tree rows, not perfectly corresponding to 0°. This fact, combined with the different crop growing period and the different canopy size and shape of the poplars in the 2 years, might also have caused the observed interaction between year and position. In the first year (2018), the canopy of poplar-SRC rows was wider and higher compared to the second year, significantly reducing the LAV from 94.70 to 74.30% and from 93.96 to 73.94% from the Center to the West side in soybean and sorghum plots, respectively. The following year after the poplar harvest (2019), the tree presence slightly reduced the LAV from 98.56 to 93.54% and from 98.72 to 95.22% passing from the Center to the East in the soybean and sorghum plots, respectively.
Similar patterns of light reduction in relation to tree distance and height were recorded by Swieter et al. (2021) in a short rotation coppice agroforestry experimental site in northern Germany. It should be noted that in our experiments the highest LAV recorded in 2018 for soybean and sorghum in the center part of the alley (an average of 94.33% at 6.75 m to the tree row) are similar to the lowest data recorded in 2019 on the East side of the alley of soybean and sorghum (an average of 94.38% at 2.5 m to the tree row). This comparison highlights how the management of the SRC is important for light availability at the herbaceous layer in an alley-cropping system based on a narrow inter-row (13.5 m). This outcome confirms the results reported by Dupraz et al. (2018) that show how crop light availability decreased with tree growth. These authors also concluded that the size and density of the trees had a larger impact on light availability at the herbaceous layer than the latitude or tree row orientation in an alley-cropping system.
Soybean and sorghum production in the alley-cropping system
Soybean and sorghum were both cultivated in the 2 years of the field experiment, and statistical analysis showed different behaviors between the two crops in the 2 years. The effect of tree growth and presence on the agronomic performance of crops was measured through the evaluation of the variation of crop production in the alley according to tree distance and sun exposure. The variation of soil water content in the agroforestry alley was not measure and this is a limitation of the study.
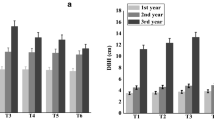
In 2018, average soybean above-ground biomass was equal to 378.53 g DM m−2 which varied significantly according to the tree distance from 543.43 to 136.85 g DM m−2 (− 74%) in the Center and the West respectively. In 2019, a lower but significant reduction was registered in the same positions from 499.83 to 278.33 g DM m−2 (Center and West, respectively). In the tree-crop interface positions, a significant difference was recorded in 2018 between soybean above-ground biomass in the East compared to the West (353.08 and 136.85 g DM m−2), while no significant differences have been observed in 2019, being the above-ground biomass 378.33 and 287.08 g DM m−2, respectively (Fig. 3). This different behavior in the tree-crop interface positions in 2018 may have been caused by a reduction of the plant density occurred in the West side, probably due to a pest damage before the emergence, compared to the East (Table S1).

Above-ground biomass, grain yield, and residues of soybean (A, B, and C, respectively) and sorghum (D, E, and F, respectively) cultivated in a poplar short rotation coppice alley-cropping system in 2018 and 2019 in Pisa, Italy. Different uppercase letters indicate significant differences among crop positions in the alley in the years of the experiment, while different lowercase letters indicate significant difference among crop positions in the alley in 2018 (light green for soybean and dark green for sorghum) or 2019 (blue for soybean and grey for sorghum), p-value < 0.05, for Tukey’s honest significant difference test. Vertical bars represent standard errors. DM means dry matter
Regarding the soybean grain yield, the average production in the alley did not vary in the 2 years of the study, 171.93 g DM m−2. A significant difference between the 2 years was recorded in the West position, with 126.77 g DM m−2 in 2019 compared to 54.91 g DM m−2 in 2018, when a lower plant density influenced the yield and affected the crop production. In the 2 years, no significant difference in yield was observed between the Center and the Mid-east positions, with the two values being close (2-years average values were 238.85 and 216.10 g DM m−2, respectively). The two positions in tree-crop interface (West and East) showed a significant reduction compared to the other positions, with 94.10 and 141.31 g DM m-2, respectively. The same result was reported by Carrier et al. (2019) which highlights how the detrimental effect on soybean yield occurred in a narrow band adjacent to the tree row.
The soybean residues in the five positions showed no differences in the 2 years, in accordance with the yield. This thus suggested that the reduction observed in yield and above-ground biomass in the West was more related to a reduced yield than a reduced plant biomass. The residues in the 2 years followed findings observed for the above-ground biomass except for the West and East, where the West showed the lowest value, while for the above-ground biomass, the West and East were comparable.
Regarding soybean biomass production and grain yield, on average in the 2 years a significant reduction was recorded in the tree-crop interface in both West and East positions (Fig. 3), which were the positions in the alley with the lowest light availability (Table 4) and the potential higher water competition level in the belowground. This detrimental effect of tree presence on soybean agronomic performance in the tree-crop interface has been reported by several authors for temperate climates (Reynolds et al. 2007; Rivest et al. 2009; Carrier et al. 2019). Recently, Gagné et at. (2022) highlighted that the effect of tree presence on crop performance varied according to the inter-annual variation of precipitation showing different patterns in dry or wet conditions. Gagné et al. (2022) showed that tree root pruning led to a decrease of soybean yield, suggesting that competition for water should be further investigated in temperate areas. Moreover, our result suggests that the most important reduction in the tree-crop interface regarded the yield and not the total plant biomass and these outcomes are not in line with Gao et al. (2013) and Carpinelli et al. (2021) who found the same detrimental effect of tree presence on soybean above-ground biomass and yield in Chinese and Brazilian agroforestry systems, respectively.
Sorghum performance was significantly affected by the “position” and the “year” only for above-ground biomass (p = 0.002) and yield (p = 0.005) (Table 3). Compared to soybean, in sorghum no significant influence of tree presence was found on the plant density for the factor “year” (p = 0.111) and “position" (p = 0.665), while the two factors affected the grain yield per plant in both years of the experiment (Table S1). The soybean density was probably affected pest damage. Regarding above-ground biomass and grain yield of sorghum, in 2018 the same differences were found according to the position in the alley. Conversely, in 2019, a decline of above-ground biomass production was registered in the West, while grain yield showed no significant differences among the five positions. In 2018, the sorghum above-ground biomass varied between 314.34 and 1020.99 g DM m−2 in the West and Mid-east, respectively, with a difference from the highest to the lowest of 69%. On the other hand, in 2019 it varied from 981.08 to 1358.83 g DM m−2 in the same positions, corresponding to a variation of 28%.
In 2018, the sorghum grain yield ranged between 63.76 g DM m−2 in the West and 263.73 g DM m−2 in the Mid-west (73% variation), while in 2019 the range of variation was more limited, with 426.56 g DM m−2 in the West and 536.68 g DM m−2 in Mid-east respectively (21%). These results show that close to the tree row sorghum had a significant above-ground biomass and yield reduction, which did not occur where competition for resources was less limited.
The sorghum residues showed almost the same reduction pattern in the West and East for both years, suggesting that the plant biomass accumulation is less sensitive than grain yield to the tree presence (Fig. 3). Moreover, at plant level the above-ground biomass per plant is less influenced by the year compared to the grain yield per plant (p = 0.048 and p = 0.002, respectively) (Table S1).
Other authors have reported the same effect of tree presence on sorghum yield, and our results are in line with grain yield losses reported by Berenji and Dahlberg (2004), showing a reduction of up to 79% with unfavorable physicochemical conditions. Deng et al. (2017) showed a grain yield reduction in the sorghum cultivar Wad Ahmed, an improved variety, when cultivated under the canopy of acacia trees in South Sudan. Righi and Foltran (2018) reported a reduction in the production of a local variety of broomcorn in Tiete (Brazil) in relation to the tree distance, showing, in line with our results, that a small reduction of available light radiation is beneficial to agronomic performance. Our results are in line with Coulibaly et al. (2014) who found a reduction in above-ground biomass and grain yield in sorghum (unknow variety) according with the tree presence in several areas of Burkina Faso. They reported that the crop harvest index varied among investigated tree-crop distances and reached the maximum in shaded plots, as occurred in the West and East of our study, compared with other positions in 2019 (on average 42 vs 39%). An increasing trend in sorghum harvest index according to the reduction of light interception and water stress was reported by Thapa et al. (2017).
Our results indicate that soybean and sorghum performance was similarly affected with regard to the tree row orientation, with the aboveground biomass being lower on the West side than on the East side, as were the grain yield and the amount of residue (Fig. 3). In particular: (i) soybean above-ground biomass production and partitioning were affected by the position in the alley, but not by the year of cultivation, (ii) sorghum above-ground biomass production and partitioning were affected by both the position in the alley and the year, and (iii) the first order interaction (year per position) of the two investigated factors was not significant either for soybean or sorghum (Table 3). These results highlight that the productivity of sorghum may be more affected by light availability reduction than soybean, and confirm previous outcomes reported by Reynolds et al. (2007) and Magalhães et al. (2019): C3 plants such as soybean are more tolerant to light reduction than C4 plants such as sorghum. Conversely, the fact that aboveground biomass and grain production of soybean were affected by tree distance similarly in the 2 years may suggest a higher effect of belowground completion by tree root (Thapa et al. 2017).
Effect of tree presence, light availability and soil characteristics on crop agronomic performance
Figure 4 shows the regression trees as a result of the recursively splitting analysis of the response of soybean and sorghum agronomic performance to the following explanatory variables: tree distance, light availability, clay and sand content in soil, soil pH, and soil organic matter content.

Regression tree showing the emerging drivers of the soybean (light blue) and sorghum (light green) above-ground biomass (a, b), grain yield (c, d) and residues (e, f) in the alley-cropping system: Distance = distance to the tree rows (m), Clay = presence of Clay in soil (g kg−1), LAV = light availability (%) and pH = soil pH; n = number of observations and y = mean yield (g DM m−2) in each terminal node
The regression tree obtained for the soybean shows that the distance from the tree is the sole significant factor affecting above-ground biomass and residue production. It could be argued that soybean is affected by the presence of trees regardless of the different light availability measured in the 2 years, as an effect of the tree canopy size. Differently, in addition to tree distance, clay content in soil also affected grain yield. Significantly higher grain yields were recorded where the clay content was higher than 268 g kg−1 thus showing that soil characteristics are crucial in rainfed systems. The clay content in soil increases the water retention capacity with possible positive effects on crop growth in rainfed conditions. Because the reduced competition for nutrients in the soil by over-fertilization, this outcome provides evidence about belowground competition for water between poplar and soybean in the tree-crop interface.
Previous studies have reported that soybean cultivated in agroforestry systems can be affected by soil moisture reduction and nutrient competition (Rivest et al. 2009; Gao et al. 2013; Gagné et al. 2022). In our experiment, nutrients were over-distributed to prevent competition between trees and crops, however during the 2 years of the study, the water deficit due to scarce rainfall seemed to affect soybean grain yield. It has been reported that the effect of competition for water between soybean and trees may be more evident during periods of prolonged drought (Rivest et al. 2009). Gagné et al. (2022) reported that competition in the aboveground was more evident than in the belowground between poplar and soybean, even under water-limited conditions. Negative response of legumes due to tree presence was also confirmed by a study conducted by our research group in the same area of the present experiment, which showed the detrimental effect of light reduction on the productivity of a perennial species, alfalfa (Medicago sativa L.), in a rainfed olive-based alley-cropping system (Mantino et al. 2021).
Future irrigation field experiments and soil water content measurements in agroforestry systems could clarify this aspect by assessing the relationship between the competition for light and for water in soybean cultivation in Mediterranean agroforestry systems.
Conversely, in sorghum, light availability was the only significant factor affecting above-ground and residue production, while grain yield also seemed to be affected by soil pH (Fig. 3). This result is in line with previous studies (Deng et al. 2017; Sidibé et al. 2017), and confirms the typical behavior of a shade-intolerant crop. In fact, we observed a large growth reduction when the tree canopy was larger, during the first year of the experiment (2 years old poplar stems). The regression tree shows that higher grain yields were recorded when the light availability was up to 97%, while when the light availability decreased below this level, the soil pH becomes crucial in terms of crop yield. Other authors have reported the significant effects of soil pH on sorghum yield (Steiner et al. 2007; Sun et al. 2019). The lowest sorghum agronomic performance was recorded where the light availability values were lower than 88, 90 and 84% for above-ground biomass, grain yield and residues, respectively (Fig. 4).
Several factors affect soybean and sorghum performance in alley-cropping systems. However, our results confirm previous studies highlighting that C3 plants, such as soybean, are more tolerant, although sensitive, to light reduction than C4 plants, such as sorghum (Reynolds et al. 2007; Magalhães et al. 2019; Ogwok et al. 2019). In contrast with Carrier et al. (2019) who found no effects of soil moisture, our results seem to indicate that soybean may be affected by the competition for water in an agroforestry system as reported by other authors (Reynolds et al. 2007; Gao et al. 2013) and this may be exacerbated in rainfed systems and drought conditions (Rivest et al. 2009; Gagné et al. 2022).
Conclusions
We have focused on the relationship between tree presence and warm-season crops, soybean and sorghum, inter-cropped with poplar SRC rows in a rainfed alley-cropping system. Under the considered experimental conditions, our results highlight that light is a crucial factor for sorghum cultivation, even if grain yield was also affected by soil characteristics such as pH. Other resource competitions, such as for water, can occur in the belowground tree-crop interface but we did not measure soil water content in the agroforestry alley, and this is a limitation of the study.
Soybean yield and aboveground biomass production showed similar pattern in the 2 years, despite the light availability was drastically different, as a consequence of the cutting cycle of the poplar SRC. This fact suggests that in the tree-crop interface the competition of trees against the soybean might has also regarded the available soil water (effect on biomass accumulation) and a higher seed predation by pests during germination stage (effect on plant density). Further studies are necessary to investigate the effect of belowground competition and pest management and to increase the knowledge about the crop response in terms of harvest index to water and light competitions in agroforestry systems.
In conclusion, the increased competition for resources in both above- and below-ground layers is a limiting factor in crop agronomic performance in an alley-cropping system with high tree density such as short rotation coppice rows. The reduction in crop growth is more evident at the tree-crop interface and less evident far from the tree rows. Short rotation coppice alley-cropping systems need to be laid out to minimize the trade-off due to tree-crop competition for resources: (i) at middle latitudes, the layout needs to respect a north–south orientation to exploit maximum light availability, (ii) the distance between trees and tree rows needs to be defined to maximize the environmental and economic benefits and (iii) crop rotation management needs to consider the different agronomic performance of different crops and the harvest cycle of tree rows.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Bagnouls F, Gaussen H (1957) les climats biologiques et leur classification. Ann Georgr 66:193–220
Bates D, Mächler M, Bolker BM, Walker SC (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Berenji J, Dahlberg J (2004) Perspectives of sorghum in Europe. J Agron Crop Sci 190:332–338. https://doi.org/10.1111/j.1439-037X.2004.00102.x
Burgess PJ, Rosati A (2018) Advances in European agroforestry: results from the AGFORWARD project. Agrofor Syst 92:801–810. https://doi.org/10.1007/s10457-018-0261-3
Carpinelli S, da Pontes LS, Da Fonseca AF, Weirich Neto PH (2021) Effect of trees and cattle dung input on soybean yield and nutrition in Integrated Crop-Livestock Systems. Agrofor Syst 95:707–716. https://doi.org/10.1007/s10457-021-00622-w
Carrier M, Rhéaume Gonzalez FA, Cogliastro A et al (2019) Light availability, weed cover and crop yields in second generation of temperate tree-based intercropping systems. F Crop Res 239:30–37. https://doi.org/10.1016/j.fcr.2019.05.004
Coulibaly YN, Mulia R, Sanou J et al (2014) Crop production under different rainfall and management conditions in agroforestry parkland systems in Burkina Faso: observations and simulation with WaNuLCAS model. Agrofor Syst 88:13–28. https://doi.org/10.1007/s10457-013-9651-8
Deng B, Tammeorg P, Luukkanen O et al (2017) Effects of Acacia seyal and biochar on soil properties and sorghum yield in agroforestry systems in South Sudan. Agrofor Syst 91:137–148. https://doi.org/10.1007/s10457-016-9914-2
Dufour L, Gosme M, Le Bec J, Dupraz C (2020) Does pollarding trees improve the crop yield in a mature alley-cropping agroforestry system? J Agron Crop Sci 206:640–649. https://doi.org/10.1111/jac.12403
Dupraz C, Blitz-Frayret C, Lecomte I et al (2018) Influence of latitude on the light availability for intercrops in an agroforestry alley-cropping system. Agrofor Syst 92:1019–1033. https://doi.org/10.1007/s10457-018-0214-x
Elghali L, Clift R, Sinclair P et al (2007) Developing a sustainability framework for the assessment of bioenergy systems. Energy Policy 35:6075–6083. https://doi.org/10.1016/j.enpol.2007.08.036
European Commission (2021) https://agriculture.ec.europa.eu/news/commission-publishes-list-potential-eco-schemes-2021-01-14_en - Last access 27 December 2022
Fox J, Weisberg S (2019) An R companion to applied regression, 2nd edn. Sage, Thousand Oaks (CA)
Frazer G, Canham C, Lertzman K (1999) Gap Light Analyzer (GLA), Version 2.0: Imaging software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs, users manual and program documentation. Program 36. citeulike-article-id:4887176
Gagné G, Lorenzetti F, Cogliastro A, Rivest D (2022) Soybean performance under moisture limitation in a temperate tree-based intercropping system. Agr Syst, 201. https://doi.org/10.1016/j.agsy.2022.103460
Gao L, Xu H, Bi H et al (2013) Intercropping competition between apple trees and crops in agroforestry systems on the loess Plateau of China. PLoS ONE 8:1–8. https://doi.org/10.1371/journal.pone.0070739
García de Jalón S, Burgess PJ, Graves A et al (2018) How is agroforestry perceived in Europe? An assessment of positive and negative aspects by stakeholders. Agrofor Syst 92:829–848. https://doi.org/10.1007/s10457-017-0116-3
Hiemstra PH, Pebesma EJ, Twenhöfel CJW, Heuvelink GBM (2009) Real-time automatic interpolation of ambient gamma dose rates from the Dutch radioactivity monitoring network. Comput Geosci 35:1711–1721. https://doi.org/10.1016/j.cageo.2008.10.011
Hothorn T, Hornik K, Zeileis A (2006) Unbiased recursive partitioning: a conditional inference framework. J Comput Graph Stat 15:651–674. https://doi.org/10.1198/106186006X133933
Kay S, Rega C, Moreno G et al (2019) Agroforestry creates carbon sinks whilst enhancing the environment in agricultural landscapes in Europe. Land Use Policy 83:581–593. https://doi.org/10.1016/j.landusepol.2019.02.025
Kanzler M, Böhm C, Mirck J et al (2019) Microclimate effects on evaporation and winter wheat (Triticum aestivum L.) yield within a temperate agroforestry system. Agrofor Syst 93:1821–1841. https://doi.org/10.1007/s10457-018-0289-4
Lamerre J, Schwarz KU, Langhof M et al (2015) Productivity of poplar short rotation coppice in an alley-cropping agroforestry system. Agrofor Syst 89:933–942. https://doi.org/10.1007/s10457-015-9825-7
Lenth RV, Buerkner O, Herve M et al (2021) Estimated marginal means, aka least-squares means. Am Stat, pp 1–89
Lovell ST, Dupraz C, Gold M et al (2018) Temperate agroforestry research: considering multifunctional woody polycultures and the design of long-term field trials. Agrofor Syst 92:1397–1415. https://doi.org/10.1007/s10457-017-0087-4
Magalhães CAS, Pedreira BC, Tonini H, Farias Neto AL (2019) Crop, livestock and forestry performance assessment under different production systems in the north of Mato Grosso, Brazil. Agrofor Syst 93:2085–2096. https://doi.org/10.1007/s10457-018-0311-x
Mantino A, Tozzini C, Bonari E et al (2021) Competition for light affects alfalfa biomass production more than its nutritive value in an olive-based alley-cropping system. Forests 12:1–13. https://doi.org/10.3390/f12020233
Mantino A, Volpi I, Micci M et al (2020) Effect of tree presence and soil characteristics on soybean yield and quality in an innovative alley-cropping system. Agronomy 10:52. https://doi.org/10.3390/agronomy10010052
Montero MJ, Moreno G, Bertomeu M (2008) Light distribution in scattered-trees open woodlands in Western Spain. Agrofor Syst 73:233–244. https://doi.org/10.1007/s10457-008-9143-4
Motsara MR, Roy RN (2015) Guide to laboratory establishment for plant nutrient analysis. Scientific Publishers, Chennai
Nassi O, Di Nasso N, Guidi W, Ragaglini G et al (2010) Biomass production and energy balance of a 12-year-old short-rotation coppice poplar stand under different cutting cycles. GCB Bioenergy 2:89–97. https://doi.org/10.1111/j.1757-1707.2010.01043.x
Ogwok G, Aele P O, Kizza S (2019) Influence of Shea tree (Vitellaria paradoxa) on maize and soybean production. 1–8. https://doi.org/10.1371/journal.pone.0201329
Palma JHN, Graves R, Bunce RGH et al (2007) Modeling environmental benefits of silvoarable agroforestry in Europe. Agric Ecosyst Environ 119:320–334. https://doi.org/10.1016/j.agee.2006.07.021
Paris P, Camilli F, Rosati A, et al (2019) What is the future for agroforestry in Italy? Agrofor Syst 93:. https://doi.org/10.1007/s10457-019-00346-y
Pebesma EJ (2004) Multivariable geostatistics in S: the gstat package. Comput Geosci 30:683–691. https://doi.org/10.1016/j.cageo.2004.03.012
Pecchioni G, Bosco S, Volpi I et al (2020) Carbon budget of an agroforestry system after being converted from a poplar short rotation coppice. Agronomy 10:1–23
Reynolds PE, Simpson JA, Thevathasan NV, Gordon AM (2007) Effects of tree competition on corn and soybean photosynthesis, growth, and yield in a temperate tree-based agroforestry intercropping system in southern Ontario, Canada. Ecol Eng 29:362–371. https://doi.org/10.1016/j.ecoleng.2006.09.024
Righi CA, Foltran DE (2018) Broomcorn [Sorghum bicolor (L.) Moench] responses to shade: an agroforestry system interface simulation. Agrofor Syst 92:693–704. https://doi.org/10.1007/s10457-016-0036-7
Rivest D, Cogliastro A, Vanasse A, Olivier A (2009) Production of soybean associated with different hybrid poplar clones in a tree-based intercropping system in southwestern Québec, Canada. Agric Ecosyst Environ 131:51–60. https://doi.org/10.1016/j.agee.2008.08.011
Seserman DM, Freese D, Swieter A, et al (2019) Trade-Off between energy wood and grain production in temperate alley-cropping systems: An empirical and simulation-based derivation of land equivalent ratio. Agric 9:. https://doi.org/10.3390/agriculture9070147
Sidibé D, Sanou H, Bayala J, Teklehaimanot Z (2017) Yield and biomass production by African eggplant (Solanum aethiopicum) and sorghum (Sorghum bicolor) intercropped with planted Ber (Ziziphus mauritiana) in Mali (West Africa). Agrofor Syst 91:1031–1042. https://doi.org/10.1007/s10457-016-9978-z
Steiner C, Teixeira WG, Lehmann J et al (2007) Long term effects of manure, charcoal and mineral fertilization on crop production and fertility on a highly weathered Central Amazonian upland soil. Plant Soil 291:275–290. https://doi.org/10.1007/s11104-007-9193-9
Sun J, He L, Li T (2019) Response of seedling growth and physiology of Sorghum bicolor (L.) Moench to saline-alkali stress. PLoS ONE 14:1–10. https://doi.org/10.1371/journal.pone.0220340
Swieter A, Langhof M, Lamerre J (2021) Competition, stress and benefits: Trees and crops in the transition zone of a temperate short rotation alley cropping agroforestry system. J Agron Crop Sci 1–16. https://doi.org/10.1111/jac.12553
Talbot G, Dupraz C (2011) Simple models for light competition within agroforestry discontinuous tree stands: are leaf clumpiness and light interception by woody parts relevant factors? Agrofor Syst 84(1):101–116. https://doi.org/10.1007/s10457-011-9418-z
Team RC (2015) R core team. R A Lang Environ Stat Comput R Found Stat Comput Vienna, Austria URL http://www.R-project.org/
Thapa S, Stewart BA, Xue Q, Chen Y (2017) Manipulating plant geometry to improve microclimate, grain yield, and harvest index in grain sorghum. PLoS ONE 12(3): e0173511. doi:https://doi.org/10.1371/journal.pone.0173511
Torralba M, Fagerholm N, Burgess PJ et al (2016) Do European agroforestry systems enhance biodiversity and ecosystem services? A meta-analysis. Agric Ecosyst Environ 230:150–161. https://doi.org/10.1016/j.agee.2016.06.002
Tsonkova P, Böhm C, Quinkenstein A, Freese D (2012) Ecological benefits provided by alley cropping systems for production of woody biomass in the temperate region: a review. Agrofor Syst 85:133–152. https://doi.org/10.1007/s10457-012-9494-8
Funding
Open access funding provided by Università di Pisa within the CRUI-CARE Agreement. No funds, grants, or other support was received. The authors have no relevant financial or non-financial interests to disclose.
Author information
Authors and Affiliations
Contributions
Conceptualization: AM, MM, GR; Methodology: AM, GR, CT, MM; Formal analysis and investigation: AM, GR, CT; MM, GP;Writing—original draft preparation: IM, GP; Writing—review and editing: GR, MM; Supervision: MM, GR.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest regarding the publication of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mantino, A., Pecchioni, G., Tozzini, C. et al. Agronomic performance of soybean and sorghum in a short rotation poplar coppice alley-cropping system under Mediterranean conditions. Agroforest Syst 97, 1025–1039 (2023). https://doi.org/10.1007/s10457-023-00845-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-023-00845-z