
Abstract
Neotropic cormorants are abundant piscivorous birds in southern Brazil and present challenges for fisheries and aquaculture as potential food competitors. To evaluate this interaction, we address the species trophic overlap with artisanal fisheries in a productive estuary in southern Brazil (48°45’27’’S, 28°30’23’’W) with abundance data, pellet samples, taxonomic composition and fishery production during July 2021 until May 2023. Bird abundance showed a seasonal pattern with peaks during autumn-winter and a population decrease during spring-summer, primarily due to the migration of reproductive individuals and the establishment of a colony from March to September. Pellet analysis identified 31 prey types within 17 taxonomical families, most of which were fish from the Ariidae (35.9%), Engraulidae (15.6%), Scianidae (12.8%), Mugilidae (10.3%), and Gerreidae (10.1%) families. We characterized the cormorant population as opportunistic, as temporal variation and Amundsen’s analysis indicated the consumption of available and diverse resources. Through otolith-total length allometric regressions we estimated a mean fish biomass per pellet of 382.7g and prey size ranging from 26.6mm to 326.9mm. When compared to artisanal fisheries, cormorants consumed 12% the amount of fish exploited by artisanal fisheries during the sampling period, although the Pianka’s index of 0.78 indicated a high overlapping food resource. Cormorant predation on juvenile fishes likely plays an important ecological role at Laguna Estuarine System by regulating abundant prey species through density-dependent mechanisms. This study provides new and updated information on cormorant population, diet and interactions with artisanal fisheries of southern Brazil.
Similar content being viewed by others


Introduction
Neotropic cormorants (Nannopterum brasilianum) are widely distributed, with breeding populations ranging from Argentina (Olrog 1975; Quintana et al. 2002; Casaux et al. 2012) to south of the United States (Kalmbach and Becker 2005). While a robust body of literature exists on their trophic ecology (Alarcón et al. 2012; Muñoz-Gil et al. 2013; Tette-Pomárico et al. 2020; Harding and Mesler 2022) and breeding colonies (Morrison et al. 1979; Azevedo 1995; Kalmbach and Becker 2005; Alves et al. 2011; Casaux et al. 2012; Quintana et al. 2002), research on these aspects in Brazil remains limited and outdated. On the past two decades, a few studies focused on cormorant diet (Barquete et al. 2008a; Branco et al. 2009; Oliveira et al. 2019) and population abundance of cormorants (Branco 2002; Barquete et al. 2008b; Silva et al. 2014). These abundant aquatic birds are distributed throughout Brazilian territory, with conspicuous populations in Amazonian basin (Silva et al.2014), Pantanal humid areas (Donatelli et al. 2014) and southern coasts of Brazil, such as Lagoa dos Patos (Barquete et al. 2008b) and Santa Catarina State (Branco 2002).
Given their feeding preference for fish and intensified foraging activity, cormorants can potentially impose significant predatory pressure on aquatic fauna and generate conflicts with fishing activities. Abundant cormorants from the northern hemisphere like the double-crested (Nannopterum auritum) and great cormorants (Phalacrocorax carbo) have shown considerable dietary overlap with fisheries, often leading to conflicts with the fishing industry (Carter et al. 1995; Adkins et al. 2014; Ovegård et al. 2017; Harding and Mesler 2022). In the Neotropics, studies have started to explore cormorants interactions with fisheries, although the focus has often been limited to local or isolated populations. Gil-Weir et al. (2011) explored N. brasilianum ecological functions (ecosystem services; bird watching, indicator of fish schools and nitrogen and phosphorus input) and economic values (i.e. cormorants as meat, eggs and guano as fertilizer) in Los Olivitos Estuary in Venezuela suggesting a minor impact on artisanal fisheries. Moreover, the growth of a Neotropic cormorant population in highland lakes in Ecuador has had tangible impacts on fish farmers, which migrating and non-breeding individuals have been observed preying on commercial fish ponds, potentially facilitating the species expansion into these highland regions (Guevara et al. 2011). In Patagonia, Argentina, a diet assessment of Neotropic cormorant identified not only a flexible feeding strategies but also an important regulatory role of two introduced exotic fish species, Oncorhyncus mykiss and Salmo trutta (Alarcón et al. 2012). While these studies emphasize some ecological roles and isolated Neotropic cormorants interactions, ecological conflicts arising from resource competition remain a prevalent issue that demands closer attention.
Such conflicts are typically fueled by the perception of resource competition, with cormorants high fish consumption rates often cited as a central concern (Kameda et al. 2003; Carss et al. 2009; Östman et al. 2013; Aguado-Giménez et al. 2018). Additionally, the conspicuousness of large cormorant flocks amplifies these perceptions, as their visible presence in key fishing areas is often misinterpreted as a direct indicator of population growth and increased pressure on fish stocks. This perceived abundance, coupled with the species high foraging activity, makes them a target of negative perceptions among fishermen, even in the absence of robust population data (Carss 2022; Dorr et al. 2022; Ludwig et al. 2023). Recently, in the southern Brazilian coast, a new cormorant colony near artisanal fisheries has raised concerns about resource competition and the depletion of fish stocks, highlighting the need for data to acknowledge the local management (Pimenta et al. 2024).
To address these concerns and provide a clearer understanding of cormorants diet and fishery interactions, it is essential to assess their dietary habits and quantify their predatory effects on fish stocks. While cormorants diet is relatively easy to assess, evaluating their predatory effects is more complex, as various factors influence fish distribution and assemblages (Ovegård et al. 2021). Thus, various methods may be employed to evaluate cormorants dietary habits, including direct observations, stomach content analysis, and the examination of regurgitated pellets (Barrett et al. 2007). Each one of these methods poses methodological challenges or advantages. Direct observations of cormorants submitting preys and stomach lavage are valid methods, but they are both qualitative data based approaches that represent only a snapshot of feeding process. Regurgitated pellets can generate both qualitative and quantitative data as different preys can be found on pellets (Barquete et al. 2008a; Tette-Pomárico et al. 2020), being non-invasive to the bird, relatively cheap and viable to collect large samples (Carss et al. 1997; Barrett et al. 2007). Despite that, pellets were long discussed to be a biased method to reconstruct size and mass of prey and demands extreme caution in considering prey sizes, as it can be overestimated by smaller prey omission (Carss et al. 1997; Votier et al. 2001).
This study aims to describe the feeding ecology of cormorants and evaluate their trophic interactions with fisheries in the lower portion of the Laguna Estuarine System (LES), Santa Catarina, Brazil. In order to do so, first, we estimated cormorant abundance through monthly counts at a major colony, considering abundances differences between seasons, breeding and non-breeding periods. Second, we identified prey taxonomic composition and sizes to estimate the total fish consumption by cormorants via regurgitated pellets samples. Then, we analyzed niche breadth, feeding strategies, and temporal diet variation to understand their predatory role. At last, we assessed resource overlap between cormorants and artisanal fisheries data to evaluate potential associations and support local future policy decisions.
Materials and methods
Study area
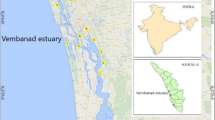
This work was conducted in a major cormorant roosting area located in the lower portion of LES. The roosting area is a semi-open lagoon, known as Noca Lagoon (48°45’27’’S, 28°30’23’’W), partially enclosed by SC-100 highway construction (Fig. 1). The lagoon is connected via water pipes to Santo Antônio Lagoon and the single channel that provides tidal influence from the sea (Barletta et al. 2017; Dantas et al. 2019). An adjacent urbanized area discharges waste directly into the lagoon, while seagrass vegetation dominates much of the lagoon’s area. The cormorant colony, covering an area of 54,000 m2, is situated on a hill predominantly covered by a secondary Atlantic rainforest.

Laguna Estuarine System location map, highlighting South of Brazil and Santa Catarina State. The figure emphasizes the three elliptical lagoons, Tubarão river discharge, the adjacent Camacho Lagoon, highway physical barrier (SC-100), Noca Lagoon and the cormorant roosting site area
Sampling design
Field sampling for both abundance counts and pellet collection was conducted concurrently from July 2021 to May 2023. Abundance data was assessed with two different methods. To determine the number of cormorants in non-breeding seasons (September to March), direct counts were made twice a month at dawn (around 6h00) using 8x42 binoculars and a mechanical counter. Cormorants tend to depart from the roosting site in small amounts (flocks of 10 to 30 birds). The observer was located on road SC-100, about 450 m west from the hill and started counting birds from 20 minutes before dawn until no bird departure occurred in a 30 minutes period. During breeding seasons (April – August), the cormorant colony had an intense traffic of birds (in and outwards) due breeding activities, such as nest construction and chick feeding, making count efforts not viable. So cormorant population was estimated by (1) counting the number of nests (1 pair of cormorants per nest) and (2) adding an estimated juvenile and non-breeding adults population. To estimate non-breeding adults, we conducted counts at perching sites along the estuary, away from the main colony. At these sites, we used plumage differences to calculate the proportion of juveniles and non-breeding adults to breeding adults. Reproductive adults show a blackish color with white filoplumes behind the beak and neck, while juveniles and non-reproductive adults maintain their natural olive-black colors (Sick et al. 1997). This ratio from the perching sites was then applied to the number of breeding adults (from the nest count) to estimate the total number of non-breeding birds, which was added for the final population estimate.
Diet data was assessed via regurgitated pellets, which can be assumed to be produced once per day, depending on physiological conditions and feeding success of cormorants, as an example of captive cormorants pellet production of 0.69 per day (McKay et al. 2003). Pellets were collected once per month during surveys on Noca roosting site, from July 2021 to May 2023. We targeted only fresh (covered in mucous) and undisturbed casks to assure pellets from the same day. During the breeding season, pellets were collected from the outer ring of the colony where non-reproductive adults and juveniles roosted. It was assumed that reproductive adults and non-reproductive individuals have the same food items type.
Pellets were stored frozen until process phase. First we defrosted in ambient water, washed the mucous in running water, while screening prey remnants on a 1 mm, 0.5mm and 63µm stacked sieves. Saggitae and lapillus (for ariidae – Genidens spp.) otoliths were used to count and identify food items. Other key hard structures such as chewing pads, pharyngeal jaws, eye lenses or vertebras were only used if otoliths were absence. We arranged otoliths by taxonomic groups, size and orientation (right and left). The total number of fish for each pellet was assumed to be the maximum number of otolith pairs counted. Fish prey was identified using the Atlas of marine bony fish of Southeastern-Southern Brazil Part I-VIII (Rossi-Wongtschowski et al. 2014; Brenha-Nunes et al. 2016; Siliprandi et al. 2016; Conversani et al. 2017; Giaretta et al. 2017; Santificetur et al. 2017a, b) and Atlas of marine fishes of southern Brazil (Haimovici et al. 2024). Crustaceans exoskeletons (especially penaeids) were not possible to be quantify due its soft and highly digestible nature, but were accounted as present or absent on samples.
Prey size can be underestimated using otolith measurements, as they are susceptible to stomach digestion (McKay et al. 2003; Barrett et al. 2007). To mitigate this methodological bias when applying allometric equations (see Online Resource 1), we used a wear index with five degrees, considering the degree of wear on the otolith edges and the sulcus acusticus (Recchia and Read 1989). The levels were defined as: (01) Intact, otoliths with no wear, extracted directly from fish; (02) Minimal, otoliths with light wear, a visible sulcus acusticus, and well-defined edges; (03) Moderate, with a visible sulcus acusticus and slightly worn edges; (04) High, with a worn sulcus acusticus and completely worn edges; (05) Total, with excessively worn sulcus acusticus and edges. Fish total length (TL) and body mass (WG) were estimated using the mean lengths of right and left otoliths from levels 01, 02, and 03, which accounted for 53.3% of all screened and paired otoliths. Otoliths from Engraulidae species are relatively smaller and more susceptible to digestion, but remained mostly identifiable even at wear level 04. To avoid losing this group taxonomic information, we included level 04 otoliths and grouped these prey items at the family level. Prey total length, body mass and total biomass per pellet were then estimated through allometric regressions from the literature on bony fishes from the southern coast of Brazil (Naves 1999; Carvalho et al. 2014; Haimovici et al. 2024).
Diet analysis
Prey composition was measured via frequency of occurrence (%FO) and numeric frequency (%N):
Where, Ni represents the number of samples which prey i occurred and N the total of samples. Ri, represents the total of prey i within all samples, and R the total number of prey. Prey was also classified according to water column distribution (demersal/benthic or pelagic).
Niche breadth was calculated as Levins (1968) and standardized to a 0 to 1 scale, following (Krebs and Davies 2010):
We delve deeper into feeding strategies by using the graphical method of Amundsen et al. (1996), in which each prey specific-abundance (Pi) is plotted on a diagram with its respective frequencies of occurrence (%FO):
Si is the total number of prey i and Sti the total number of preys occurring on samples that prey i is present. The distribution of prey on the plot indicates their relative importance, rarity, and dominance within the diet. For instance, prey with high Pi and %FO is considered dominant, suggesting specialization, while prey with low values is more rare or incidental. The overall prey distribution provides insights into the predator’s feeding strategy, revealing whether it tends toward a generalist or specialist approach, and helps identify intra- and inter-phenotype differences in diet composition, as discussed by Amundsen et al. (1996).
To investigate the significance of monthly variation in the diet of N. brasilianum, we conducted a permutational multivariate analysis of variance (PERMANOVA) with 999 permutations (Anderson 2001). A Hellinger transformation was applied to balance prey abundance data, minimizing the dominance of abundant species and enhancing the representation of rare species, identified with Amundsen’s graphical analysis. As a post hoc analysis, we identified the most significant variations between groups (months) using pairwise p-values (Martinez Arbizu 2020). To further explore and identify food items contribution for specific months, we performed an Indicator Species Analysis (ISA) (De Cáceres 2013).
Fishing production data and diet overlap
Fishing production data were obtained from the Santa Catarina State Monitoring of Fishing Activities Project (PMAP-SC) for the period from July 2021 to June 2023 (PMAP-SC 2024). This project collects monthly sampling data on fishing production from various cities in Santa Catarina. This includes information on the type of fishing gear used, total effort (monthly), species captured, fishing production (in kilograms), and the number of discharges during the specified period. For this analysis, we focused on artisanal fishing types I and II, excluding industrial fishing. Type I involves fishing with or without vessels up to 10 meters, while Type II utilizes vessels larger than 10 meters. Both types were grouped to provide a comprehensive overview of artisanal fishing in the Laguna Estuarine System. We removed the fishing gears used in ocean waters to better explore those employed within the estuarine waters. Since cormorants forage mostly on estuarine waters, we removed ring nets, beach seines, and manual collection (which focus on mollusks as the target species). To compare artisanal fishing and cormorant biomass consumption, monthly and annual biomass consumption was estimated by multiplying cormorant abundance, days of a given month and calculated mean biomass per pellet (Barquete et al. 2008a). To estimate dietary overlap between Neotropic cormorants and artisanal fisheries we utilized Pianka’s Index (Pianka 1973). Seasonal and overall indexes were calculated with consumed biomass proportions, with index values ranging from 0 (no overlap) to 1 (total overlap):
where Qic represents the biomass proportion of prey species i consumed by Neotropic cormorants, and \(Q_{if}\) represents the biomass proportion of prey species i targeted by artisanal fisheries of LES.
Statistical analysis
All statistical analyses were performed using R version 4.3.2 (R Core Team 2024). The abundance data were tested for normality, heteroscedasticity, and dispersion. Given the temporal nature of the count data, we tested for autocorrelation using the acf() function from the ‘tseries’ package (Trapletti et al. 2015). We identified temporal autocorrelation and accounted the discrete structure of the data by employing a Generalized Linear Mixed Model (GLMM) with a negative binomial distribution, using the glmmTMB() function from the ‘glmmTMB’ package (Brooks et al. 2023). Reproductive periods were included on the model as a fixed effect to test for differences on bird counts between breeding and non-breeding periods. We treated the monthly autocorrelation implementing a first order structure nested by year, ‘ar1(month_num + 0 | year)’. Model residuals were then evaluated using the DHARMa package, which confirmed that the temporal autocorrelation was effectively corrected and that the model appropriately accounted for the discrete structure of the data (see Online Resource 2).
To test for temporal diet composition variation, a PERMANOVA was performed using the function adonis2() from ‘Vegan’ package (Oksanen et al. 2022), while the post hoc test was performed with the function pairwise.adonis() from ‘pairwiseAdonis’ package (Martinez Arbizu 2020). The function multipatt() from package ‘Indicspecies” was used to execute the Indicator Species Analysis (De Cáceres 2013). Dietary overlap between cormorants and artisanal fisheries was calculated using the function pianka() from package ‘EcoSimR’ (Gotelli et al. 2013). The significance of the dietary overlap was assessed by comparing the observed values to those generated through randomization, with 1000 iterations performed on the function niche_null_model() (method “pianka” and algorithm “r3”). The observed overlap index was compared to the distribution of simulated indices, and a standardized effect size (SES) was calculated for future comparisons.
Results
Cormorant abundance
Cormorant abundance at Laguna Estuarine System varied throughout months of all sampled years. We identified seasonal tendencies of higher peaks of abundance during autumn and winter (March - August), while a decreased numbers during September and February. We accounted for a monthly mean abundance of 4390 ± 1626 cormorants. Lowest abundance occurred in September 2021 with 1345 cormorant and highest numbers occurred during the breeding period July 2023, an estimate of 5040 reproductive adults and 2308 juveniles and non-breeding adults (not accounting for born chicks) (Fig 2).

Observed temporal abundances of Nannopterum brasilianum at Noca Lagoon dormitory, Laguna, Santa Catarina, Brazil. Horizontal arrows represent non-breeding (blue) and breeding periods (red)
The negative binomial GLMM with an AR(1) correlation structure indicated a significant effect of the reproductive season on abundance, with higher counts during the reproductive period (Estimate = 0.44, z = 4.48, p < 0.001; Fig. 3a). Furthermore, the density distribution of counts across seasons (Fig. 3b) highlighted greater abundance during Autumn and Winter. The AR(1) structure effectively addressed temporal autocorrelation, as confirmed by the residual analysis (Fig. 3c). The random effect of year accounted for interannual variability (Variance = 0.043, SD = 0.208).

Overview of the effect of season on abundance a GLMM showing the relationship between seasons and abundance, highlighting higher values during the breeding season (Autumn and Winter). b Density distribution of counts across seasons, illustrating seasonal variations. c Diagnostic plot of Pearson residuals by month for the negative binomial GLMM with an AR(1) correlation structure
Diet composition, prey sizes and feeding strategies
We collected a total of 201 pellets over 19 months, with a mean of 10.15 ± 3.2 samples per month. These samples contained 31 different food items distributed across 17 families. Teleost fish were the dominant prey, occurring in 90% of samples (n = 181), while crustaceans were present in 29.1% (n = 59). In total, 4494 individual prey items were screened, with an average of 16.4 ± 18.6 prey per sample and a maximum of 188 prey in a single pellet.
From the total, 2397 prey items were identified to the lowest possible taxonomic level using a wear index to account for digestive erosion. Demersal and pelagic prey were equally represented in terms of numerical abundance (%N), accounting for 54.9 and 45.1% of the diet, respectively. A notable example of the identification challenges and solutions was the Engraulidae family: from 376 engraulid prey, we identified 188 as Anchovia clupeoides, 31 to the genus Anchoa sp., and four as Anchoviella lepidentostole. The remaining 153 engraulids could not be identified to a lower taxonomic level due to high wear (Table 1).
For 1,554 prey with suitable otoliths, we retrocalculated total length and mass using allometric equations. The estimated average prey size was 117.1 ± 67.7 mm (ranging from 26.6 to 326.9 mm). The mean consumed biomass per pellet was 382.8 ± 555.8 g, ranging from 1.2 g to 979.8 g.
The overall diet niche breadth for N. brasilianum was 0.287. Amundsen’s graphical analysis revealed the prey with the highest prey-specific abundance: Trachinotus spp., Menticirrhus americanus, Urophycis spp., Genidens spp., and Eucinostomus spp. (Fig. 4). These species were distributed across different quadrants of the plot according to their respective frequencies of occurrence.

Amundsen’s graphical analysis of feeding strategies for Nannopterum brasilianum at southern coast of Santa Catarina, Brazil. Dots represent the relative position of each prey, classified by numbers. 1- Genidens spp., 2- Engraulidae, 3- Micropogonias furnieri, 4- Eucinostomus spp., 5- A. brasiliensis, 6- M. curema, 7- M. liza, 8 – D. rhombeus, 9- Sardinella brasiliensis, 10- C. spilopterus, 11- Ctenogobius spp., 12- Centropomus spp., 13- P. isoceles, 14- S. rastrifer, 15- M. litorallis, 16- Saurida brasiliensis, 17- Urophycis spp., 18 – e.n.i., 19- Trachinotus spp., 20- M. americanus, 21- P. brasiliensis, 22- Cynoscion spp., 23- Ctenosciane spp., 24- Strongylura spp., 25- Achirus lineatus
Micropogonias furnieri exhibited low prey-specific abundance but high frequency of occurrence, confirming its status as a dominant and constant resource in the diet, alongside Genidens spp. In contrast, Atherinella brasiliensis and Mugil curema showed relatively low values for both metrics, indicating lower overall importance. The remaining prey, located in the bottom left quadrant of the plot, are considered rare or incidental food items.
Temporal diet variation
The PERMANOVA results, based on Hellinger dissimilarity, indicated significant temporal variation in the diet of cormorants (F = 3.305, P < 0.001, perm = 999) (Table 2). Pairwise comparisons revealed 23 significant pairwise dissimilarities among months. The winter months of July and August 2021 were the primary drivers of this variation, collectively involved in 12 of the 23 significant comparisons (July 2021: 7 comparisons; August 2021: 5 comparisons). This indicates that the diet composition during the winter of 2021 was distinctly different from most other periods. Furthermore, September and October of 2022 also showed highly distinct diets, with each being significantly dissimilar to a wide range of other months (see Online resource 4). Indicator Species Analysis (ISA) revealed significant contributions of Genidens spp., M. furnieri and M. curema to temporal variations in cormorant diet thought the whole sampling period. A few other preys contributed significantly on specific months, such as M. liza, engrualids, A. brasiliensis and gerreids (Table 3). Mugil liza was more prominent in warmer months, such as January and February, while Gerreids and A. brasiliensis were associated with a broader range of months but exhibited peaks in cooler months, such as May, June, and July. Additionally, engraulids emerged as significant contributors, particularly during cooler months (July 2022 and May–June 2023), reflecting their availability and potential aggregation during these periods. Interestingly, Saurida brasiliensis showed a significant association with a limited number of sampling groups (e.g., August 2022 and June 2023). This last result likely arises from its occurrence in just two unique and distinct diet samples, where it was the only prey item recorded, thereby inflating its indicator value.
Fishing production and dietary overlap
LES artisanal fisheries targeted 39 different species (excluding Elasmobranchii). The most frequently captured species were Callinectes spp. (36.2%), mullets (24.2%), penaeids (12.9%), Whitemouth croakers (6.8%) and clupeiformes (6.5%) (see Online Resource 3). Over the sampled period, artisanal fisheries extracted more than 13,625 tons of aquatic resources, while cormorants were estimated to consume 1,665 tons. A dietary overlap index of 0.78 was estimated between cormorants and artisanal fisheries, with a mean simulated value of 0.52 (nReps = 1000, P < 0.001). Seasonal indexes ranged from 0.63 on summer 2021/2022 to 0.87 on winter 2023, with a mean value of 0.80 ± 0.08.
Discussion
Our multifaceted results reveal that the Neotropic Cormorant population in the Laguna Estuarine System is both abundant and exhibits an opportunistic feeding strategy, which differs from patterns reported in other studies. This detailed assessment of its diet provides crucial insights into its feeding ecology and resource use, particularly by highlighting the potential for competition with artisanal fisheries. Given that this species remains underexplored in Brazilian waters, our findings help to fill knowledge gaps and establish a baseline for understanding the species ecological roles and mitigating potential new conflicts, as have already occurred in the LES (Pimenta et al. 2024).
Cormorant abundances and fluctuations
Neotropic cormorant population of LES exhibited fluctuations patterns of abundance with higher peaks of individuals during autumn and winter, while a decrease in numbers during summer and spring. This seasonal pattern coincides with the establishment of a reproductive colony in LES from March to August as cormorants distributions can be shifted within breeding and feeding grounds (Barquete et al. 2008a,b; Branco et al. 2009). The establishment of cormorant breeding colonies is perceived to increase bird numbers, enhances their feeding activities (Quintana et al. 2002), raise fish predation and generate ecological conflicts among fisheries activities of Laguna region (Pimenta et al. 2024). Last publication focused on population assessment of N. brasilianum in south of Brazil occurred over 20 years ago in Lagoa dos Patos (Barquete et al. 2008b) and Saco da Fazenda Estuary (Branco 2002), accounting a max population of approximately 1390 and 650 respectively. LES cormorant population fluctuated from approximate 1300 individuals at spring of 2021 to 7800 during 2023 breeding period. Our study not only brings new comparable population numbers, but also the information of an established and conspicuous reproductive colony on southern Brazil, which may contribute significantly to cormorants distribution on Brazilian Southern coasts, Pantanal and Argentina. Newly established colonies and increased cormorant numbers could be an indirect response to climate change, as changes in aquatic habitats influence systems from the individual to the community level and also decrease biodiversity (Wrona et al. 2006; Harley 2011; Rodrigues-Filho et al. 2023), thereby favoring generalist predators such as cormorants. Furthermore, extreme climate change events may also be driving these changes, for instance, by causing higher chick mortality during heat waves (Quintana et al. 2022) or depletion of food availability through severe droughts, since this birds forage on waterbodies (Braz-Mota and Val 2024).
Diet composition and feeding strategies
Neotropic cormorant at LES showed opportunistic behaviors with a diverse diet and fish as main food item. We identified 31 different prey types within 17 taxonomic families. The majority of fish consumption by N. brasilianum is observed in other regions of the species distribution range, including Brazil (Barquete et al. 2008a; Branco et al. 2009; Oliveira et al. 2019), Venezuela (Gil-Weir et al. 2011), Argentina (Alarcón et al. 2012; Casaux et al. 2009, 2012), Colombia (Tette-Pomárico et al. 2020) and United States (Harding and Mesler 2022). Within all identified teleost, Catfishes (Genidens spp.), Engraulids, Whitemouth croakers (M. furnieri), Gerreids (Eucinostomus spp. and D. rhombeus), flounders (C. spilopterus), Brazilian silverside (A. brasiliensis) and mullets (M. liza and M. curema) were previously observed to be abundant species in LES (Sunye et al. 2014; Barletta et al. 2017; Dantas et al. 2019; Frischknecht et al. 2023) and major preys of LES cormorants population.
Neotropic Cormorants are considered a generalist predator due to its wide distribution and plasticity in food resource use. Our niche breadth value of 0.287 for the LES population is substantially higher than values reported for other populations, such as 0.056 in Lagoa dos Patos (Barquete et al. 2008a) and 0.02 in northern Colombia (Tette-Pomaricó et al. 2020). This key difference is not a contradiction but rather a clear reflection of contrasting prey availability across these ecosystems. The extremely low value in Colombia (0.02) indicates a forced specialization, where the diet was dominated by a single family (Ariidae) due to a documented decline in historically important Engraulidae prey populations caused by overfishing (Tette-Pomaricó et al. 2020). The low value in Lagoa dos Patos (0.056) also indicates a specialized diet on a few abundant demersal fish, which together comprised over 94% of local fish catches (Barquete et al. 2008a; Vieira, 1991). In contrast, the Laguna Estuarine System supports a dynamic fish assemblage (Freischneck et al. 2023), providing a wider spectrum of abundant prey throughout the year. This greater resource diversity allows the local cormorant population to exploit a broader niche, resulting in the higher Levin’s index. This pattern strongly supports the concept that the niche breadth in Neotropic cormorants is a plastic indicator of local resource availability rather than a fixed taxonomic trait. As noted by Barquete et al. (2008a), cormorants can prey heavily on abundant resources (a specialist tactic) within an overall generalist and opportunistic framework.
Using the feeding strategy graphical analysis as a qualitative approach (Amundsen et al. 1996), we identified several abundant fish species, such as Catfishes, Whitemouth croakers, Engraulids, Gerreids and Brazilian silverside, consistent with an opportunistic feeding strategy. We also identified a few prey types with high specific abundances but low frequency of occurrence, meaning they were consumed by only a few individuals (Amundsen, 1996) and appeared in a limited number of samples, often as the sole prey type. These preys in Amundsen’s plot are Menticirrhus americanus, Trachinotus sp. and Urophycis sp., being all marine only species. The presence of exclusively marine species may be attributed to intra-specific variations within the cormorant population, as individual birds may exploit different resources based on factors such as prey spatial distribution, individual experience, sex, and age (Ceia and Ramos 2015). Also, cormorants are observed to forage on flocks and individually (Quintana et al. 2004. Oliveira et al. 2019), on shallow ocean waters (Quintana et al. 2004) and estuarine waters (Barquete et al. 2008a; Branco et al. 2009). Although our results indicate a preference for estuarine waters in the LES, cormorants are observed to forage in shallow oceanic waters less frequently (not published).
The presence of pelagic and demersal preys on cormorant diet also indicates an opportunistic behavior which may be explained by the capacity on changing foraging strategies (Quintana et al. 2004), even under high turbidity scores (Grémillet et al. 2012). For instance, cormorants can switch targets during diving sessions (Cosolo et al. 2010), transitioning from predictable, sediment-associated demersal prey (Cook et al. 2006) to dense and conspicuous schools of pelagic fish. Overall, the diverse and adaptable diet of Neotropic cormorants at LES highlights their opportunistic feeding strategy, shaped by prey availability and environmental conditions, reinforcing their role as a key predator in the estuarine ecosystem.
Prey size and daily food intake
Our data estimates a mean prey size of 117.1mm on the diet of N. brasilianum. Similar sizes of 113.4mm and 120 mm were also estimated by Barquete et al. (2008a) and Harding and Mesler (2022), respectively in Lagoa dos Patos, southern Brazil and Arizona, United States. With a prey size ranging from 26 to 326 mm, Neotropic cormorants feeds not only juveniles, but also adult sizes, although cormorant morphology may limit the ingestion of larger fishes (~300mm), as discussed by Barquete et al. (2008a). Additionally, the growth patterns of fish, particularly the time required to reach a size detectable and capturable by piscivorous birds, as well as the period they remain within a consumable size range (i.e. >300mm), are secondary, but important factors that influence their presence on cormorant diet (Riedel et al. 2007). In addition, we also suggest the possibility of cormorants feeding upon discarded heads by fisherman cleaning fish along the estuary, as artisanal fisheries are an intensive activity at Southern State of Santa Catarina (Barletta et al. 2017). By consuming a broad range of sizes, cormorants may regulate fish population density and minimizes competition among fish of various size classes, which could favor growth and surviving of key species by reducing competition (Casaux et al. 2012).
If assumed that one pellet is produced per day, we evaluated a daily food intake of 382 g of fish, a lower number compared to the 425 g per day of Neotropic cormorants at Lagoa dos Patos (Barquete et al. 2008a), 828 g of P. carbo (Grémillet et al. 1996) and many other species around the world (Ridgway 2010). We aimed to reduce size estimation bias by using a worn index, but most of our smaller preys were removed from the analysis, possibly influencing towards a higher mean biomass per pellet (M°Kay et al. 2003; Barret et al. 2007; Ridgway 2010; Tette-Pomárico et al. 2020).
Temporal variation on prey preferences
Estuarine fish assembly is spatially and temporally diverse (species composition and life stages) as abiotic conditions (i.e. tide regimes, salinity, precipitation and water depth) creates different habitats for fish during their life (Henriques et al. 2017; Rodrigues-Filho et al. 2023). For instance, absence of fresh water prey in this study may be explained due cormorants night roosting site, Noca Lagoon, be closer to ocean waters and suffer influences from coastal waters and salinity, key factor that shapes distributions of fishes from Ariidae family (Dantas et al. 2012). For instance, Genidens genidens is an estuary-dependent species and exhibits different distributions and behaviors throughout its life stages (Silva Junior et al. 2013), with juveniles using shallow waters and adults occupying deeper waters (Dantas et al. 2019). The presence of juvenile catfishes (<45mm) in cormorant diet may be associated to fish inexperience (Van Eerden and Carss 2012) and a limited moving capability (Barbieri et al. 1992). Thus, higher population densities of G. genidens occurs after spawning periods, especially in warmer months (spring-summer) (Silva Junior et al. 2013), increasing the species abundance and raising food availability for cormorants, which could explain higher occurrences of catfishes on cormorant diet.
Another important prey to cormorants, gerreids, is a group strongly associated with salinity gradients and abundant in LES. For example, Eucinostomus melanopterus is more common during higher salinity conditions in spring, while E. argenteus is associated with lower salinity scores in autumn (Frischknecht et al. 2023). Our cormorant prey abundance data indicates higher abundances of gerreids during both periods, suggesting that prey availability may influence cormorant diet in specific months.
Biotic elements such as presence of vegetation (e.g., seagrass and mangroves), reproductive patterns, spawning periods, life stages, and migration also play a crucial role in determining fish assembly. Spawning may enhance abundances of Atherinella brasiliensis at post reproductive periods, presenting higher abundances during Summer and Autumn (Cattani et al. 2019). Although this species abundance and resilient presence during the whole year may be explained due higher niche breadth and capability to occur on different habitats (e.i seagrass and mangroves) (Rodrigues-Filho et al. 2024). Also, White mullets (Mugil curema) are opportunistic estuarine resident species in LES and abundant on the region, which are favored by temperatures above of 20 °C and higher salinity scores (Vieira and Vieira 1991; Mai et al. 2018). Another important mullet species in the LES is Mugil liza. This marine migratory species is highly dependent on the estuarine environment during its juvenile phase, with individuals occurring in the system year-round (Garbin et al. 2014; Herbst and Hanazaki 2014; Lemos et al. 2014). However, the fishery primarily targets large, migratory adults that enter the estuary in massive schools during the winter reproductive migration (Daura-Jorge et al. 2013). Both mullets have different biological demands but similar morphological characteristics, which their different temporal variation appearances in cormorant diet may be explained by fluctuations in environmental conditions and their different life history strategies (Meurer and Netto 2007; Frischknecht et al. 2023). Another key factor that influences fish and other aquatic organisms is precipitation, as rain regimes introduce fresh water to the estuarine system shifting the salinity gradient and turbidity (Barletta et al. 2017). The latter factor is essential for prey accessibility as cormorants use visual cues of prey during flights and dives, but in contrast, also influence prey awareness (Strod et al. 2008). For future diet assessments, we suggest subsequent daily sampling efforts to test daily diet composition and associations between precipitation scores and prey diversity in cormorant diet, associated to salinity and fresh water species.
Cormorants interactions with LES artisanal fisheries
The high diet overlap index (0.78) between Neotropic cormorant and artisanal fisheries in the LES suggests strong potential for resources competition. However, a detailed analysis of species composition and temporal dynamics reveals mechanisms that may mitigate this direct competition.
The high overlap is primarily driven by shared key species such as Mugil Liza and Micropogonias furnieri, which account for significant fishery landings of 24.2 and 6.8% of estuarine biomass during the sampling period, respectively (Sunye et al. 2014; Piazza et al. 2021; PMAP-SC 2024). Despite the high numerical overlap, resource partitioning is evidenced by cormorants predominantly preying on M. furnieri juvenils, while fisheries target adults of commercially valuable sizes (size partitioning). For M. liza, niche partitioning occurs on a temporal scale and by fish size. Cormorant predation of juveniles peaks in summer, revealed by Indicator Species Analysis (Table 3), whereas the fishery commercial sizes peaks occurs during the winter which targets a massive migratory adults (>300mm) (Daura-Jorge et al. 2013; Schroeder et al. 2023). In addition, the seasonal shift in cormorant feeding preferences, focusing on other abundant prey like engraulids and gerreids during winter, effectively reduces competition for the key shared resources (M. liza and M. furnieri). The constant presence of Genidens spp. in the cormorant diet across all seasons also contributes to the baseline overlap.
The broader context of the artisanal fishery, which exploits over 39 species but concentrates effort on five key resources (crabs, mullets, penaeids, whitemouth croakers, and clupeiforms, comprising 87% of landed biomass) (Barletta et al. 2017; Sunye et al. 2014; Piazza et al. 2021; PMAP-SC 2024), further explains the mitigated overlap dynamic. Additionally, non-selective fishing methods, such as fyke nets, generate significant bycatch and discards (Garthe et al. 1996; Furness et al. 2007), which likely subsidize the cormorant diet (e.g., crabs) and influences the distribution of many other aquatic birds (Branco 2001; Barbieri 2010; Carniel and Krul 2012). Crucially, the majority of the cormorant’s diet consists of non-target or low-value species (e.g., Genidens spp. at ~0.4% of fishery biomass).
Our data suggests a minimal economical impact of cormorant predation on artisanal fisheries. For that, we considered (1) the size and temporal partitioning of key overlapping species, (2) cormorant predation mostly on non-target species, (3) the natural density-dependent juvenile mortality (Ovegård et al. 2021) and (4) an estimative of cormorant consumption (1,665 tons) representing a 12.2% of fishery extraction (13,625 tons). Cormorants likely play a vital ecological role by preying on abundant juveniles and utilizing discards, thereby contributing to local diversity maintenance and potentially reducing competitive pressures among fish species (Rodrigues-Filho et al. 2024).
Conclusions
This study provides new insights into the trophic ecology of the Neotropic Cormorant and its relationship with artisanal fisheries in southern Brazil. Here we documented seasonal abundance patterns, with the highest population peaks linked to the establishment of a reproductive colony during the austral autumn and winter. Diet assessments revealed a varied use of food resources, dominated by teleost fish and complemented by penaeids, while significant temporal variation underscored the species opportunistic foraging strategy. By delving deep into this multifaceted diet analysis, temporal variation on prey availability and prey sizes are determinants for niche partitioning between cormorants and artisanal fisheries of LES, mitigating direct competition for resources. The data presented here provide a critical base for stakeholders to develop targeted management measures for the Laguna Estuarine System and other similar socio-ecological systems where such predator-fishery dynamics occur.
Data availability
The datasets generated and analyzed during this study may be shared by the corresponding author upon reasonable request for academic and non-commercial research purposes.
References
Adkins JY, Roby DD, Lyons DE, Courtot KN, Collis K, Carter HR, Shuford WD, Capitolo PJ (2014) Recent population size, trends, and limiting factors for the double-crested cormorant in western North America. J Wild Mgmt 78:1131–1142. https://doi.org/10.1002/jwmg.737
Aguado-Giménez F, Eguía-Martínez S, Torres-Campos I, Meroño-García S, Martínez-Ródenas J (2018) Competition for food between the Mediterranean shag, the great cormorant and artisanal fisheries: a case study. Sci Mar 82(1):7. https://doi.org/10.3989/scimar.04680.04B
Alarcón PAE, Macchi PJ, Trejo A, Alonso MF (2012) Diet of the Neotropical Cormorant (Phalacrocorax brasilianus) in a Patagonian freshwater environment invaded by exotic fish. Waterbirds 35(1):149–153. https://doi.org/10.1675/063.035.0115
Alves VS, Soares ABA, Couto GS, Draghi J (2011) Padrão de ocorrência e distribuição de biguás Phalacrocorax brasilianus na Baía de Guanabara, Rio de Janeiro, Brasil. Revista Brasileira de Ornitologia 19: 469–477. http://revbrasilornitol.com.br/BJO/article/view/4502. Accessed 27 August 2024
Amundsen PA, Gabler HM, Staldvik FJ (1996) A new approach to graphical analysis of feeding strategy from stomach contents data—modification of the Costello (1990) method. J Fish Biol 48(4):607–614. https://doi.org/10.1111/j.1095-8649.1996.tb01455.x
Anderson MJ (2001) A new method for non‐parametric multivariate analysis of variance. Austral Ecol 26(1):32–46. https://doi.org/10.1111/j.1442-9993.2001.01070.pp.x
Azevedo TR (1995) Estudo da avifauna do campus da Universidade Federal de Santa Catarina (Florianópolis). Biotemas 8(2):7–35
Barbieri LR, dos Santos RP, Andreata JV (1992) Reproductive biology of the marine catfish, Genidens genidens (Siluriformes, Ariidae), in the Jacarepaguá Lagoon system, Rio de Janeiro, Brazil. Environ Biol Fishes 35:23–35
Barbieri E (2010) Seasonal abundance of Magnificent Frigatebird, Fregata magnificens during the year 2006 at Ilha Comprida (São Paulo, Brazil), and its relation to the number of shrimp fishing boats. Revista Brasileira de Ornitologia 18: 99–110. http://revbrasilornitol.com.br/BJO/article/view/4005 Accessed 27 August 2024.
Barletta M, Lima ARA, Dantas DV, Oliveira IM, Neto JR, Fernandes CAF, Farias EGG, Filho JLR, Costa MF (2017) How can accurate landing stats help in designing better fisheries and environmental management for Western Atlantic Estuaries? Em C. W. Finkl and C. Makowski (Orgs.). Coast Wetl: Alter Remediat 21:631–703. https://doi.org/10.1007/978-3-319-56179-0_20
Barquete V, Bugoni L, Vooren CM (2008) Diet of Neotropic cormorant (Phalacrocorax brasilianus) in an estuarine environment. Mar Biol 153(3):431–443. https://doi.org/10.1007/s00227-007-0824-8
Barquete V, Vooren CM, Bugoni L (2008) Seasonal abundance of the Neotropic Cormorant (Phalacrocorax brasilianus) at Lagoa dos Patos estuary, Southern Brazil. El Hornero 23(1):15–22. https://doi.org/10.56178/eh.v23i1.745
Barrett RT, Camphuysen KCJ, Anker-Nilssen T, Chardine JW, Furness RW, Garthe S, Hüppop O, Leopold MF, Montevecchi WA, Veit RR (2007) Diet studies of seabirds: a review and recommendations. ICES J Mar Sci 64(9):1675–1691. https://doi.org/10.1093/icesjms/fsm152
Branco JO (2001) Descartes da pesca do camarão sete-barbas como fonte de alimento para aves marinhas. Rev Bras Zool 18(1):293–300. https://doi.org/10.1590/S0101-81752001000100033
Branco JO (2002) Flutuações sazonais na abundância de Phalacrocorax brasilianus (Gmelin) no estuário do Saco da Fazenda, Itajaí, Santa Catarina, Brasil. Rev Bras Zool 19(4):1057–1062. https://doi.org/10.1590/S0101-81752002000400010
Branco JO, Evangelista CL, Lunardon-Branco MJ, de Azevedo-Júnior SM, de Larrazábal ME (2009) Atividade diária de Phalacrocorax brasilianus (Aves, Phalacrocoracidae), na região do Saco da Fazenda, Itajaí, SC, Brasil. Ornithologia 3(2):73–82
Braz-Mota S, Val AL (2024) Fish mortality in the Amazonian drought of 2023: the role of experimental biology in our response to climate change. J Exp Biol 227(17):jeb247255. https://doi.org/10.1242/jeb.247255
Brenha-Nunes MR, Santificetur C, Conversani VRM, Giaretta MB, Rossi-Wongtschowski CLDB, Siliprandi CC (2016) Atlas of marine bony fish otoliths (sagittae) of Southeastern-Southern Brazil Part IV: Perciformes (Centropomidae, Acropomatidae, Serranidae, Priacanthidae, Malacanthidae, Pomatomidae, Carangidae, Lutjanidae, Gerreidae and Haemulidae). Braz J Oceanogr 64(1):23–75. https://doi.org/10.1590/S1679-875920161100064(sp1)
Brooks M, Bolker B, Kristensen K, Maechler M, Magnusson A, McGillycuddy M, Skaug H, Nielsen A, Berg C, van Bentham K, Sadat N, Ludecke D, Lenth R, O’Brien J, Geyer CJ, Jagan M, Wiernik B, Stouffer DB, Agronah M (2023) Package ‘glmmtmb.’ R Packag Vers 1(1):7
Carniel VL & Krul R (2012) Use of artisanal fishery discards by seabirds on the Paraná Coast of Brazil. Mar Ornithol 40:57–62. https://digitalcommons.usf.edu/marine_ornithology/vol40/iss1/8. Accessed 28 August 2024
Carss DN (2022) There must be some kind of way out of here: towards ‘reframing’ European cormorant-fisheries conflicts. Ardea 109(3):667–681. https://doi.org/10.5253/arde.v109i2.a31
Carss DN, Bevan R, Bonetti A, Cherubini G, Davies J, Doherty D, El Hili A, Feltham M, Grade N, Granadeiro J (1997) Techniques for assessing cormorant diet and food intake: towards a consensus view. Suppl Ric Biol Selvaggina 26:197–230
Carss DN, Bell S, Marzano M (2009) Competing and coexisting with cormorants: ambiguity and change in European wetlands. Landsc, Process Power: Re-eval Tradit Environ Knowl 10:99–121. https://doi.org/10.1515/9781845459048-007
Carter HR, Sowls AL, Rodway MS, Wilson UW, Lowe RW, McChesney GJ, Gress F, Anderson DW (1995) Population size, trends, and conservation problems of the double-crested cormorant on the Pacific coast of North America. Colon Waterbirds 18:189. https://doi.org/10.2307/1521540
Carvalho BMD, Corrêa MFM, Volpedo A (2014) Lapillus otoliths of the Cathorops spixii (Spix and Agassiz, 1829) and Genidens genidens (Cuvier, 1829) (Actinopterygii—Ariidae). Acta Sci Biol Sci 36(3):343. https://doi.org/10.4025/actascibiolsci.v36i3.21117
Casaux RJ, Prinzio CYD, Bertolin ML, Tartara MA (2009) Diet of the Neotropic Cormorant Phalacrocorax olivaceus at West Chubut, Patagonia. Argent Waterbirds 32(3):444–449. https://doi.org/10.1675/063.032.0310
Casaux R, Ramón A, Tartara MA, Borrell V, Gonc R (2012) Diet of breeding Neotropic Cormorants at the Carrileufu River, Patagonia: is there any impact on recreational fish resources? Ardeola 59(2):279–289. https://doi.org/10.13157/arla.59.2.2012.279
Cattani AP, Ribeiro GC, Cardoso OR, Afonso MG, Hostim-Silva M, Pichler HA, Spach H (2019) Diversity and space-time dynamics of fish assemblages in a coastal lagoon, western Atlantic. Neotrop Biol Conserv 14(2):221–239. https://doi.org/10.3897/neotropical.14.e37667
Ceia FR, Ramos JA (2015) Individual specialization in the foraging and feeding strategies of seabirds: a review. Mar Biol 162:1923–1938. https://doi.org/10.1007/s00227-015-2735-4
Conversani VRM, Brenha-Nunes MR, Santificetur C, Giaretta MB, Siliprandi CC, Rossi-Wongtschowski CLDB (2017) Atlas of marine bony fish otoliths (sagittae) of Southeastern-Southern Brazil part VII: Atheriniformes, Beloniformes, Beryciformes, Zeiformes, Syngnathiformes, Scorpaeniformes and Tetraodontiformes. Braz J Oceanogr 65(3):400–447. https://doi.org/10.1590/s1679-87592017134306503
Cook TR, Cherel Y, Tremblay Y (2006) Foraging tactics of chick-rearing Crozet shags: individuals display repetitive activity and diving patterns over time. Polar Biol 29(7):562–569. https://doi.org/10.1007/s00300-005-0089-y
Cosolo M, Ferrero EA, Sponza S (2010) Prey ecology and behaviour affect foraging strategies in the Great Cormorant. Mar Biol 157:2533–2544. https://doi.org/10.1007/s00227-010-1517-2
Dantas DV, Barletta M, Araújo Lima AR, De Assis Almeida Ramos J, Da Costa MF, Saint-Paul U (2012) Nursery habitat shifts in an estuarine ecosystem: patterns of use by sympatric catfish species. Estuaries Coasts 35(2):587–602. https://doi.org/10.1007/s12237-011-9452-0
Dantas DV, Ribeiro CIR, Frischknecht CDCA, Machado R, Farias EGG (2019) Ingestion of plastic fragments by the Guri sea catfish Genidens genidens (Cuvier, 1829) in a subtropical coastal estuarine system. Environ Sci Pollut Res 26(8):8344–8351. https://doi.org/10.1007/s11356-019-04244-9
Daura-Jorge FG, Ingram SN, Simões-Lopes PC (2013) Seasonal abundance and adult survival of bottlenose dolphins (Tursiops truncatus) in a community that cooperatively forages with fishermen in southern Brazil. Marine Mammal Sci 29(2):293–311. https://doi.org/10.1111/j.1748-7692.2012.00571.x
De Cáceres M (2013) How to use the indicspecies package (ver. 1.7. 1). R Proj 29:1–29
Donatelli RJ, Posso SR, Toledo MCB (2014) Distribution, composition and seasonality of aquatic birds in the Nhecolândia sub-region of South Pantanal, Brazil. Braz J Biol 74(4):844–853. https://doi.org/10.1590/1519-6984.05013
Dorr BS, Fielder DG, Jackson JR, Farquhar JF, Schultz DW, Claramunt RM (2022) Ontario’s double-crested cormorant hunting season may be ineffective but that doesn’t mean there are no conflict issues. Avian Conserv Ecol. https://doi.org/10.5751/ACE-02249-170211
Van Eerden MR, Carss DN (2012) Cormorants and the European environment: Exploring cormorant ecology on a continental scale. NERC Centre for Ecology and Hydrology on behalf of COST
Ferrero EA, Sponza S (2010) Prey ecology and behaviour affect foraging strategies in the Great Cormorant. Mar Biol 157:2533–2544. https://doi.org/10.1007/s00227-010-1517-2
Frischknecht CDCA, Dantas DV, Rodrigues-Filho JL, Gentil E (2023) Composition and diversity of fish assemblages along a shallow beach of a neotropical coastal lagoon. Reg Stud Mar Sci 63:102993. https://doi.org/10.1016/j.rsma.2023.102993
Furness R, Edwards A, Oro D (2007) Influence of management practices and of scavenging seabirds on availability of fisheries discards to benthic scavengers. Mar Ecol Prog Ser 350:235–244. https://doi.org/10.3354/meps07191
Garbin T, Castello JP, Kinas PG (2014) Age, growth, and mortality of the mullet Mugil liza in Brazil’s southern and southeastern coastal regions. Fish Res 149:61–68. https://doi.org/10.1016/j.fishres.2013.09.008
Garthe S, Camphuysen K, Furness R (1996) Amounts of discards by commercial fisheries and their significance as food for seabirds in the North Sea. Mar Ecol Prog Ser 136:1–11
Giaretta MB, Siliprandi CC, Santificetur C, Brenha-Nunes MR, Conversani VRM, Rossi-Wongtschowski CLDB (2017) Atlas of marine bony fish otoliths (sagittae) of Southeastern-Southern Brazil Part VI: Albuliformes, Anguiliformes, Osmeriformes, Stomiiformes, Aulopiformes, Myctophiformes, Ophidiiformes, Polimixiiformes, Batrachoidiformes and Lophiformes. Braz J Oceanogr 65:258–308. https://doi.org/10.1590/s1679-87592017133806502
Gil-Weir K, Weir E, Casler CL, Aniyar S (2011) Ecological functions and economic value of the Neotropic Cormorant (Phalacrocorax brasilianus) in Los Olivitos Estuary, Venezuela. Environ Dev Econ 16:553–572. https://doi.org/10.1017/S1355770X11000179
Gotelli, N.J. and A.M. Ellison. 2013. EcoSimR 1.00. http://www.uvm.edu/~ngotelli/EcoSim/EcoSim.html
Grémillet D, Dey R, Wanless S, Harris MP, Regel J (1996) Determining food intake by Great Cormorants and European Shags with electronic balances. J Field Ornithol 67:637–648
Grémillet D, Nazirides T, Nikolaou H, Crivelli AJ (2012) Fish are not safe from great cormorants in turbid water. Aquat Biol 15:187–194. https://doi.org/10.3354/ab00430
Guevara EA, G Ts, Mueces T, Terán K, Henry P-Y (2011) Population growth and seasonal abundance of the Neotropic Cormorant (Phalacrocorax brasilianus) at Highland Lakes in Ecuador. Waterbirds 34:499–503. https://doi.org/10.1675/063.034.0413
Haimovici M, Rodrigues LS, Lucato SHB, Freire MA, Fischer LG, Cardoso LG (2024) Otolith atlas for marine fishes of the southwestern Atlantic occurring along southern Brazil (28° S-34° S). Mar Fish Sci (MAFIS). https://doi.org/10.47193/mafis.3712024010101
Harding LE, Mesler JI (2022) Cormorant abundance, diet, and foraging habits in Arizona. J Field Ornithol. https://doi.org/10.5751/JFO-00141-930306
Harley DG (2011) Climate change, keystone predation, and biodiversity loss. Science 334:1124–1127. https://doi.org/10.1126/science.1210199
Henriques S, Cardoso P, Cardoso I, Laborde M, Cabral HN, Vasconcelos RP (2017) Processes underpinning fish species composition patterns in estuarine ecosystems worldwide. J Biogeogr 44(3):627–639. https://doi.org/10.1111/jbi.12824
Herbst DF, Hanazaki N (2014) Local ecological knowledge of fishers about the life cycle and temporal patterns in the migration of mullet (Mugil liza) in Southern Brazil. Neotrop Ichthyol 12:879–890. https://doi.org/10.1590/1982-0224-20130156
Kalmbach E, Becker PH (2005) Growth and survival of neotropic cormorant (Phalacrocorax brasilianus) chicks in relation to hatching order and brood size. J Ornithol 146:91–98. https://doi.org/10.1007/s10336-004-0061-6
Kameda K, Ishida A, Naruse M (2003) Population increase of the Great cormorant Phalacrocorax carbo hanedae in Japan: conflicts with fisheries and trees and future. Vogelwelt 124(Suppl):27–33
Krebs JR, Davies NB (2010) An introduction to behavioural ecology, 3rd edn. Blackwell, London
Lemos VM, Varela AS, Schwingel PR, Muelbert JH, Vieira JP (2014) Migration and reproductive biology of Mugil liza (Teleostei: Mugilidae) in south Brazil. J Fish Biol 85:671–687. https://doi.org/10.1111/jfb.12452
Levins R (1968) Evolution in changing environments. Princeton University Press, Princeton
Ludwig J, Cooke S, Hobson K (2023) On the lack of scientific evidence for the Ontario cormorant cull and other cormorant management actions: a response to Dorr et al. (2022). Avian Conserv Ecol. https://doi.org/10.5751/ACE-02375-180101
Mai ACG, Santos MLD, Lemos VM, Vieira JP (2018) Discrimination of habitat use between two sympatric species of mullets, Mugil curema and Mugil liza (Mugiliformes: Mugilidae) in the rio Tramandaí Estuary, determined by otolith chemistry. Neotrop Ichthyol. https://doi.org/10.1590/1982-0224-20170045
Martinez Arbizu P (2020). pairwiseAdonis: Pairwise multilevel comparison using Adonis. R package version 0.0, 4.
Marzano M, Carss DN, Cheyne I (2013) Managing European cormorant-fisheries conflicts: problems, practicalities and policy. Fish Manag Ecol 20:401–413. https://doi.org/10.1111/fme.12025
McKay HV, Rushton KA, Carss D, Parrott D (2003) The limitations of pellet analysis in the study of cormorant Phalacrocorax spp. diet. Vogelwelt 124
Meurer AZ, Netto SA (2007) Seasonal dynamics of benthic communities in a shallow sublittoral site of Laguna estuarine system (South, Brazil). Braz J Aquat Sci Technol 11:53. https://doi.org/10.14210/bjast.v11n2.p53-62
Morrison ML, Shanley E, Slack RD (1979) Breeding biology and age-specific mortality of Olivaceous Cormorants. Southwest Nat 24:259–266. https://doi.org/10.2307/3670923
Muñoz-Gil J, Marín-Espinoza G, Andrade-Vigo J, Zavala R, Mata A (2013) Trophic position of the Neotropic Cormorant (Phalacrocorax brasilianus): integrating diet and stable isotope analysis. J Ornithol 154:13–18. https://doi.org/10.1007/s10336-012-0863-x
Naves LC (1999) Ecologia alimentar do Talha-mar Rhynchops nigra (Aves: Rhynchopidae) na desembocadura da Lagoa dos Patos. Dissertation, Univ Fed Rio Grande, Rio Grande
Oksanen J, Simpson G, Blanchet FG, Kindt R, Legendre P, Minchin P, O’Hara R, Solymos P, Stevens H, Szöcs E, Wagner H, Barbour M, Bedward M, Bolker B, Borcard D, Carvalho G, Chirico M, De Cáceres M, Durand S, Weedon J (2022) Vegan community ecology package. Version 2.6-2. https://CRAN.R-project.org/package=vegan
Oliveira KR, Corrêa LLC, Petry MV (2019) Dieta de Nannopterum brasilianus (Aves: Phalacrocoracidae), no sul do Brasil. Oecol 23:432–439. https://doi.org/10.4257/oeco.2019.2303.03
Olrog CC (1975) Vagrancy of neotropical cormorant, egrets, and white-faced ibis. Bird Banding 46:207. https://doi.org/10.2307/4512136
Östman Ö, Boström MK, Bergström U, Andersson J, Lunneryd S-G (2013) Estimating competition between wildlife and humans–a case of cormorants and coastal fisheries in the Baltic Sea. PLoS One. https://doi.org/10.1371/journal.pone.0083763
Ovegård MK, Öhman K, Mikkelsen JS, Jepsen N (2017) Cormorant predation overlaps with fish communities and commercial-fishery interest in a Swedish lake. Mar Freshw Res 68:1677. https://doi.org/10.1071/MF16227
Ovegård MK, Jepsen N, Bergenius Nord M, Petersson E (2021) Cormorant predation effects on fish populations: a global meta-analysis. Fish Fish 22:605–622. https://doi.org/10.1111/faf.12540
Pianka E (1973) The structure of lizard communities. Annu Rev Ecol Evol Syst 4:53–74
Piazza TB, Pessoa MB, Santiago DF, Perroca JF, Wagner B, Dantas DV, Barbieri E, Rodrigues-Filho JL (2021) Aspectos econômicos, tecnológicos e socioculturais da pesca de siri-azul no sistema estuarino de Laguna (SC), e sua dinâmica espaço-temporal através do conhecimento ecológico local de pescadores artesanais. In: Engenharia de pesca: Aspectos teóricos e práticos. Editora Científica Digital, pp 36–61. https://doi.org/10.37885/210404220
Pimenta VRA, Rodrigues Filho JL, Ataides JS, Branco JO (2024) Human-wildlife conflict between neotropic cormorant and artisanal fisheries induces dubious management policies in southern Brazil. Ocean Coast Res 72:e24066. https://doi.org/10.1590/2675-2824072.23193
PMAP-SC (2024) Projeto de Monitoramento da Atividade Pesqueira no Estado de Santa Catarina. Universidade do Vale do Itajaí (UNIVALI), Laboratório de Estudos Marinhos Aplicados (LEMA). http://pmap-sc.acad.univali.br/sistema.html?id=57c71daf1db18d0003045194. Accessed 3 Nov 2024
Quintana F, Yorio P, Borboroglu PG (2002) Aspects of the breeding biology of the neotropic cormorant Phalacrocorax olivaceus at Golfo San Jorge, Argentina. Mar Ornithol 30:25–29
Quintana F, Yorio P, Lisnizer N, Gatto A, Soria G (2004) Diving behavior and foraging areas of the Neotropic Cormorant at a marine colony in Patagonia, Argentina. Wilson Bull 116:83–88. https://doi.org/10.1676/0043-5643(2004)116[0083:DBAFAO]2.0.CO;2
Quintana F, Uhart MM, Gallo L, Materra MB, Rimondi A, Gómez-Laich A (2022) Heat-related massive chick mortality in an Imperial Cormorant Leucocarbo atriceps colony from Patagonia, Argentina. Polar Biol 45:275–284. https://doi.org/10.1007/s00300-021-02982-6
R Core Team (2024) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Recchia CA, Read AJ (1989) Stomach contents of harbour porpoises, Phocoena phocoena (L.), from the Bay of Fundy. Can J Zool 67:2140–2146. https://doi.org/10.1139/z89-304
Ridgway MS (2010) A review of estimates of daily energy expenditure and food intake in cormorants (Phalacrocorax spp.). J Great Lakes Res 36:93–99. https://doi.org/10.1016/j.jglr.2009.10.002
Riedel R, Caskey LM, Hurlbert SH (2007) Length-weight relations and growth rates of dominant fishes of the Salton Sea: implications for predation by fish-eating birds. Lake Reserv Manage 23(5):528–535. https://doi.org/10.1080/07438140709354036
Rodrigues Filho JL, Santiago DF, Piazza TB, Dantas DV, de Souza Penha VA, Branco JO (2024) Habitat distinctness doesn’t influence on the abundance and relative growth of the juveniles Atherinella brasiliensis (Quoy & Gaimard, 1824) in a subtropical estuarine ecosystem. Reg Stud Mar Sci 77:103663. https://doi.org/10.1016/j.rsma.2024.103663
Rodrigues-Filho JL, Macêdo RL, Sarmento H, Pimenta VRA, Alonso C, Teixeira CR, Pagliosa PR, Netto SA, Santos NLC, Daura-Jorge FG, Rocha O, Horta P, Branco JO, Sartor R, Muller J, Cionek VM (2023) From ecological functions to ecosystem services: linking coastal lagoons biodiversity with human well-being. Hydrobiologia 850(12):2611–2653. https://doi.org/10.1007/s10750-023-05171-0
Rossi-Wongtschowski CLDB, Siliprandi CC, Brenha MR, Gonsales SDA, Santificetur C, Vaz-dos-Santos AM (2014) Atlas of marine bony fish otoliths (Sagittae) of Southeastern - Southern Brazil Part I: Gadiformes (Macrouridae, Moridae, Bregmacerotidae, Phycidae and Merlucciidae); Part II: Perciformes (Carangidae, Sciaenidae, Scombridae and Serranidae). Braz J Oceanogr 62:1–103. https://doi.org/10.1590/S1679-875920140637062sp1
Santificetur C, Conversani VRM, Brenha-Nunes MR, Giaretta MB, Siliprandi CC, Rossi-Wongtschowski CLDB (2017) Atlas of marine bony fish otoliths (sagittae) of Southeastern-Southern Brazil Part V: Perciformes (Sparidae, Sciaenidae, Polynemidae, Mullidae, Kyphosidae, Chaetodontidae, Mugilidae, Scaridae, Percophidae, Pinguipedidae, Blenniidae, Gobiidae, Ephippidae, Sphyraenidae, Gempylidae, Trichiuridae, Scombridae, Ariommatidae, Stromateidae and Caproidae). Braz J Oceanogr 65:201–257. https://doi.org/10.1590/s1679-87592017131006502
Santificetur C, Giaretta MB, Conversani VRM, Brenha-Nunes MR, Siliprandi CC, Rossi-Wongtschowski CLDB (2017) Atlas of marine bony fish otoliths of Southeastern-Southern Brazil Part VIII: Siluriformes (Ariidae) and Pleuronectiformes (Achiridae, Paralichthyidae, Cynoglossidae). Braz J Oceanogr 65:448–494. https://doi.org/10.1590/s1679-87592017143106503
Schroeder R, Avigliano E, Volpedo AV, Fortunato RC, Barrulas P, Daros FA, Schwingel PR, Dias MC, Correia AT (2023) Lebranche mullet Mugil liza population structure and connectivity patterns in the southwest Altantic ocean using a multidisciplinary approach. Estuar Coast Shelf Sci 288:108368. https://doi.org/10.1016/j.ecss.2023.108368
Sick H, Pacheco JF, Haffer J (1997) Ornitologia brasileira. Ed Nova Fronteira
Siliprandi CC, Brenha-Nunes MR, Rossi-Wongtschowski CLDB, Santificetur C, Conversani VRM (2016) Atlas of marine bony fish otoliths (sagittae) of Southeastern-Southern Brazil Part III: Clupeiformes (Clupeidae, Engraulidae, Pristigasteridae). Braz J Oceanogr 64(spe1):1–22. https://doi.org/10.1590/S1679-875920150988064(sp1)
Silva C, Aguiar-Abdo M, Oliveira D, Girard P, Empresa Brasileira de Pesquisa Agropecuária (2000) Caracterização ambiental do ninhal Corutuba, Pantanal de Barão de Melgaço, MT. In: Empresa Brasileira de Pesquisa Agropecuária (ed) Anais do III Simpósio sobre Recursos Naturais e Sócio-Econômicos do Pantanal, Corumbá, Brasil
Silva EF, Naiff RH, Barata FDB, Santos Júnior LS, França PF, Campos CEC (2014) Abundância sazonal de Phalacrocorax brasilianus (Gmelin, 1789) (Aves, Phalacrocoracidae) na porção norte da bacia amazônica, Macapá, Amapá, Brasil. Biota Amazonia 4:64–67. https://doi.org/10.18561/2179-5746/biotaamazonia.v4n2p64-67
Silva Junior DR, Carvalho DMT, Vianna M (2013) The catfish Genidens genidens (Cuvier, 1829) as a potential sentinel species in Brazilian estuarine waters. J Appl Ichthyol 29:1297–1303. https://doi.org/10.1111/jai.12280
Strod T, Izhaki I, Arad Z, Katzir G (2008) Prey detection by great cormorant (Phalacrocorax carbo sinensis) in clear and in turbid water. J Exp Biol 211:866–872. https://doi.org/10.1242/jeb.014324
Sunye PS, Pereira TJ, Russo A, Netto SA (2014) A pesca do camarão-rosa no Sistema Estuarino de Laguna, SC: história e acasos das políticas de manejo. In: Haimovici M, Andriguetto Filho JM, Sunye PS (eds) A pesca marinha e estuarina no Brasil: Estudos de casos multidisciplinares. Editora da FURG
Tette-Pomárico A, López-Peña A, Franke-Ante R, Duarte LO (2020) Dieta y amplitud del nicho trófico del cormorán neotropical (Nannopterum brasilianus [Gmelin, 1789]) en el Santuario de Flora y Fauna Ciénaga Grande de Santa Marta, Caribe de Colombia. Bol Invest Mar Cost 49(SuplEsp):193–208. https://doi.org/10.25268/bimc.invemar.2020.49.SuplEsp.1072
Trapletti A, Hornik K, LeBaron B, Hornik MK (2015) Package ‘tseries.’ R project. https://doi.org/10.32614/CRAN.package.tseries
Vieira JP, Vieira JP (1991) Juvenile mullets (Pisces: Mugilidae) in the estuary of Lagoa dos Patos, RS, Brazil. Copeia 1991:409. https://doi.org/10.2307/1446590
Votier SC, Bearhop S, Ratcliffe N, Furness RW (2001) Pellets as indicators of diet in Great Skuas (Catharacta skua). Bird Study 48:373–376. https://doi.org/10.1080/00063650109461237
Wrona FJ, Prowse TD, Reist JD, Hobbie JE, Lévesque LMJ, Vincent WF (2006) Climate change effects on aquatic biota, ecosystem structure and function. Ambio 35(7):359–369. https://doi.org/10.1579/0044-7447(2006)35[359:CCEOAB]2.0.CO;2
Acknowledgements
We are grateful to Post graduate program in Ecologia and Natural Resources of Universidade Federal de São Carlos (UFSCar) and State University of Santa Catarina (UDESC) for infrastructure support; we also thank Jorge Luiz Rodrigues-Filho on permitting the utilization of the Laboratory of Conservation and Applied Ecology (LEAC) and David Valença Dantas on otolith identification support at Laboratory of Management Ecology and Marine Technology (GTMAR). We thank all participant fisherman (Joacir, Chipa and Filipe) of Ponta da Barra which supported the research with vessels to access the sampling site. We are thankful to Edison Barbieri, Evelise N. F. de Moura, Manoel M. Dias-Filho and Vivian M. Cionek for providing critical suggestions on the first draft of this manuscript, as well as the anonymous reviewers for their insightful comments and constructive criticism, which significantly strengthened the quality of this manuscript.
Funding
The Article Processing Charge (APC) for the publication of this research was funded by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) (ROR identifier: 00x0ma614). The research was financed in part by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brazil (CAPES) – Finance 88887.498111/2020-00 and Fundação de Amparo à Pesquisa e Inovação de Santa Catarina (FAPESC TO 2023TR000694). JLRF would like to thank Fundação de Amparo à Pesquisa e Inovação do Estado de Santa Catarina (FAPESC TO 2023TR001395) for providing the financial and infrastructure support to carry out this work.
Author information
Authors and Affiliations
Contributions
VRAP designed and conceived the study, conducted fieldwork, analyzed the data, interpreted the data and wrote the original draft. JRLF interpreted the data, supported the scientific framework development, supervised and revised the draft writing. NKT aided on fieldwork, aided on laboratory phase and edited the original draft. JOB supervised and revised the draft writing and supported the scientific framework development. All authors are in agreement to the final draft and granted permission to submit for publication.
Corresponding author
Ethics declarations
Conflict of interests
The authors declare no competing interests.
Additional information
Communicated by RAFAEL LEITAO.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pimenta, V.R.A., Rodrigues-Filho, J.L., Toma, N.K. et al. Feeding ecology of Nannopterum brasilianum (Gmelin, 1789) and its trophic interactions with artisanal fisheries in the Laguna Estuarine System, Southern Brazil. Aquat Ecol 60, 17 (2026). https://doi.org/10.1007/s10452-025-10257-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1007/s10452-025-10257-z