
Abstract
Burial assemblages inform us about the biology of past societies, social relations, and ritual and symbolic behavior. However, they also allow us to examine the circumstances of death and social violence. A high level of intergroup violence among prehistoric hunter-gatherers is well-documented in some times and places but is extremely rare in others. Here we present an analysis of the perimortem injury to skeleton PD8 at the site of Sphinx in Central Sudan. This burial, attributed to the Early Khartoum (Khartoum Mesolithic) culture, radiocarbon dated between 8637 and 8463 cal BP, bears evidence of a perimortem sharp force trauma caused by penetration of an unshaped, fractured non-human bone between the right scapula and the rib cage. Among more than 200 anthropologically assessed human burials from the early Holocene Nile Valley reviewed in this paper, PD8 provides the only documented evidence of violence resulting in death. This rare case of death differs from the numerous cases of intergroup conflict documented in terminal Pleistocene burial grounds in Lower Nubia. This suggests different patterns of violence and strategies of conflict resolution in the pre-Neolithic (terminal Pleistocene and early Holocene) Nile Valley. We attribute this difference in the prevalence of interpersonal trauma to climatic and environmental conditions, territorial boundary defense, and post-marital residence practices before and after the Younger Dryas’ arid millennium (~ 12,800–11,600 BP).
Résumé
Les ensembles funéraires nous renseignent généralement sur la biologie des sociétés passées, leurs relations sociales et leurs comportements rituels et symboliques. Ils peuvent également nous permettre d'examiner les circonstances de la mort des individus et de discuter de la violence sociale. Un niveau élevé de violence intergroupes est documenté à certaines périodes et dans certains endroits au sein de populations préhistoriques de chasseurs-cueilleurs, alors qu’il semble inexistant ou rare à d’autres moments. Nous présentons ici une analyse d’une lésion périmortem présente sur le squelette PD8 du site préhistorique de Sphinx (Soudan central). Cette sépulture, associée à la culture Early Khartoum (Mésolithique de Khartoum) et datée au radiocarbone entre 8637 et 8463 cal BP, porte la preuve d'un traumatisme périmortem causé par la pénétration d'un os non humain fracturé et non-façonné entre l'omoplate droite et la cage thoracique. Parmi les plus de 200 sépultures humaines examinées anthropologiquement dans la vallée du Nil au début de l'Holocène, dont nous faisons brièvement la revue dans cet article, PD8 témoigne du seul acte de violence documenté à ce jour ayant entraîné la mort. Ce cas rare de décès diffère des nombreux cas de conflits intergroupes identifiés dans les complexes funéraires de la fin du Pléistocène supérieur en Basse Nubie. Cela suggère des modèles assez différents de rapports humains vis-vis des actes de violence interpersonnels et des stratégies de résolution de conflits. Nous les discutons ici plus en détail en lien avec les conditions climatiques et environnementales, les modes de peuplement, la défense des frontières territoriales et les modèles de résidence post-marital avant et après le millénaire aride du Dryas récent (~ 12 800–11 600 BP).
Similar content being viewed by others


Introduction
Lawrence H. Keeley (1996) has shown that intergroup violence was common among hunter-gatherers. Several ethnographic cases among hunter-gatherers have also documented frequent intra-group acts of violence resulting in death (Lee, 1979; Lomas, 2008-2009). However, the frequency of conflicts aimed at or resulting in killing, both organized and unplanned, does not appear to be constant among hunter-gatherer societies (Chapais, 2008; Fry & Söderberg, 2014; Kang, 1979; Keeley 1996; Kelly, 2000; Manson & Wrangham, 1991; Pettitt, 2011; Rodseth et al., 1991). One way for archaeology to examine this issue is to compare mortuary assemblages that capture the quantity and character of violence in their broader social, economic, and palaeoecological contexts.
From an evolutionary point of view, the ability to prevent or mitigate violence constitutes an important group or individual skill (Hames, 2019). Understanding this skill from a long-term perspective is an important task of social archaeology (Orschiedt, 2020). Likewise, exceptions to the predictive relationship between the ecological theory of human territoriality and intergroup violence may provide insights into the diversity of human social and territorial organization strategies (Ambrose & Lorenz, 1990; Dyson-Hudson & Smith, 1978). These exceptions also counter the pervasive influence of Raymond Dart’s (1953) theory of the predatory transition from ape to human. It posits an atavistic universal human propensity for violence, evinced by assemblages of fractured fossilized bones interpreted as tools and weapons made by Australopithecus africanus. Robert Ardrey’s expansive exegesis of Dart’s thesis in African Genesis (1961), The Territorial Imperative (1966), and The Hunting Hypothesis (1976) was highly influential among evolutionary biologists, paleoanthropologists, and the broader public, promoting acceptance of Dart’s thesis of the atavistic killer ape that we still harbor today despite the doubts first raised by Sherwood Washburn (1957). Lawrence Keeley (in Kim & Kissel, 2018) provides an analogous critique of atavism inferred from chimpanzee (Pan troglodytes) lethal intergroup conflict, using the counterexample of bonobo (Pan paniscus) peaceful intergroup interactions. For these reasons, attestations of violence among prehistoric hunter-gatherers are of special significance for understanding the limits to ecological uniformitarianism in explaining the diversity of modern human conflict mediation strategies. With these usually small-sized groups, violence or its prevention functioned along different lines compared to more complex societies with socially designated privileges and power (Scherer, 2021).
The archaeological assessment of violence in prehistoric hunter-gatherer societies stands on more solid ground where burial assemblages are large enough to study causes of mortality and identification of trauma. There is evidence of elaborate mortuary traditions among anatomically modern humans in the Levant ca. 126 ka and among Neanderthals across western Eurasia by 60 ka (Pettitt, 2011). Burials richly endowed with red ochre are a characteristic feature of the Gravettian of western Eurasia, 30–21 ka (French & Nowell, 2022), and Willandra Lakes, Australia, has evidence for cremation and red ochre burials by 40 ka (Pettitt, 2011). Large cemeteries become widespread at the end of the Pleistocene and the early Holocene (ca. 15–8 ka) (Cullen, 1995; Gibaja et al., 2015; Humphrey et al., 2011; Pettitt, 2011).
Interpersonal violence and likely intergroup feuds are well-documented during the Terminal Pleistocene in the lower Nile Valley at sites such as Jebel Sahaba and Wadi Halfa, near the Second Cataract at the Sudan-Egypt border (Crevecoeur et al., 2021; Saxe, 1971) and the Early Holocene in the Turkana Basin (Lahr et al., 2016a, 2016b). However, skeletal evidence for interpersonal violence is extremely rare during the Early Holocene in the middle Nile Valley of Sudan. This article presents and evaluates a unique case of violence resulting in death in an early Holocene hunter-gatherer context at Sphinx, near the Sixth Nile Cataract in Central Sudan. We set this case within the context of penecontemporaneous and earlier attestations of violent deaths in the Nile Basin and discern two different patterns of violence during the Terminal Pleistocene and Early Holocene. Finally, we argue that these contrasting patterns of violence most likely reflect different strategies of lethal conflict mitigation associated with distinct technology, subsistence, kinship, and social organization strategies used by hunter-gatherers to cope with climatically and environmentally different settings.
Site of Sphinx
The archaeological site of Sphinx (SBK.W-60; 16°18′43.27″N, 32°38′8.34″E) is situated in the western part of the Sabaloka Mountains (Jebel Sabaloka) in the vicinity of the Sixth Nile Cataract in Central Sudan (Fig. 1). This site consists of several parts located on and around one of the small granite hills that dot the northwestern periphery of the mountain (Fig. 2A). Its main area (792 m2), which occupies a rocky platform 15 m above the surrounding terrain, has been explored since 2011 by the Charles University Sabaloka Expedition (Prague, Czech Republic). Intact deposits more than 1-m thick preserve remains of the settlement and a burial ground attributed to Early Holocene hunter-gatherers of the Early Khartoum Culture (Varadzinová et al., 2022a, 2022b). Radiocarbon dates attest to a more than four-millennia-long site occupation between 11,057 and 6659 cal BP (Varadzinová et al., 2023). Human burials form a well-defined phase of occupation of the site between 8720 and 7867 cal BP.

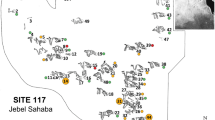
The Nile basin with marked locations of pre-Neolithic burial assemblages discussed in the text. (A) Location of the Nile basin in the African continent. (B) Close up on the archaeological sites in central Sudan. Dark gray circles = late Upper Pleistocene sites; light gray circles = early Holocene sites; stars = sites with occurrences of perimortem trauma. Map adapted from Williams (2009)

Sphinx (SBK.W-60). (A) Location of the site in the western part of Jebel Sabaloka (Google Earth 2018). (B) Plan of Sphinx with trenches; trenches with human burials hatched; the area with PD8 is indicated as a black rectangle inside Trench 2
Remains of at least 45 individuals in varying states of preservation were unearthed in five of the eleven trenches excavated between 2011 and 2019 (Fig. 2B) (Brukner Havelková et al., 2022). They exhibit a compact carbonaceous crust resulting from post-depositional precipitation of calcium carbonates (cf. Varadzinová Suková et al., 2015). Most individuals were placed in a contracted position in grave pits dug into earlier settlement deposits, and backfilled with sediment containing redeposited settlement debris such as flaked and ground stone artifacts, pottery fragments, faunal remains, and other finds. Several burials exhibited signs of post-depositional disturbance due to interment of new individuals or reorganization of previous interments (Brukner Havelková et al., 2022; Varadzinová & Varadzin, 2017; Varadzinová et al., 2022a, 2022b). The demographic structure of the burial sample shows no selection based on sex, but there is a marked underrepresentation of immature individuals, especially those under the age of one year. The population at Sphinx appears to be relatively healthy and peaceful, with most of the recorded pathological alterations likely resulting from everyday activities (Brukner Havelková et al., 2022). The only exception is primary deposition 8 (PD8) from Trench 2, which exhibits an unusual perimortem trauma. This case is described in detail in this study.
Material
The individual PD8, designated in the field as burial 14 (B14), was found above three other interments. It clearly disturbed two burials (PD6 and PD7) and superimposed and possibly partly disturbed PD9 (Fig. 3). The body of PD8 was found in a contracted position on its left side, the right lower limb folded against the abdomen, with its feet drawn close to the pelvis. The right upper limb was also folded, with the forearm drawn across the chest towards the head. The rib cage exhibits natural flattening related to the decomposition process. The body was oriented to the west, with the face to the north. Elements of another individual were found in association with PD8 (see Fig. 3). These included isolated, nearly complete long bones likely belonging to PD6. Six pieces of unworked granite were found along the left lower limb (3 pieces) or directly on the body of PD8—on the right hand, in the abdominal area, and over the lower limbs. Finally, four ground stones were found in association with PD8, with three sandstone upper grinders (two complete and one fragment) found behind the head close to the right shoulder and at the level of the right elbow, and one complete lower grinder of porphyritic rhyolite placed on a layer of sediment above the knees of PD8. The sediment surrounding the skeletal remains contained two red rhyolite flakes, pottery fragments, ground stone artifacts, ostrich eggshell beads, pieces of mica, animal bones, mollusc shells, and a quartz crystal. None of the finds was in direct contact with the skeleton, and we assume that most, if not all, of these finds represent redeposited settlement debris from earlier Holocene contexts.

Sphinx. Plan and photograph of PD8. Dark gray = PD8; light gray = other primary depositions; black = earlier human bones redeposited along PD8; medium gray = complete and fragmented grindstones; white = boulders and pieces of local granite
Calcareous concretions heavily cemented the remains of PD8, so several parts of its body had to be removed as blocks of sediment. After excavation, the PD8 human remains were transported to the Department of Anthropology of the National Museum in Prague to be cleaned, inventoried, and analyzed. Export was authorized by a permit granted to Lenka Varadzinová by the National Corporation for Antiquities and Museums of Sudan. Cleaning of the right shoulder block that preserved the right clavicle in anatomical connection with the acromion of the right scapula and five ribs revealed a non-human bone fragment of dark color embedded between the scapula and the ribs. It was cemented to the adjacent human bones with carbonate concretions.
Methods
Petra Brukner Havelková cleaned the skeletal remains manually in the laboratory using water and a brush. Before cleaning of the cemented right shoulder of PD8 with the embedded faunal fragment was completed, the block was digitized by X-ray microtomography at the UMS 3626 PLACAMAT from Bordeaux University with a General Electric microscanner (GE v|tome|x s) and a 10.81-µm voxel size. Three-dimensional imaging and manual segmentation of the non-human fragment and the human remains in the cemented block were performed at Bordeaux University with the Avizo 7.1 software (VSG—2012).
X-ray fluorescence (XRF) was used to identify the chemical element composition of the dark pigmentation area of the animal bone embedded in the human skeleton. Analysis was performed in the Natural History Museum in Prague-Horní Počernice with a mobile µXRF spectrometer—the Bruker ARTAX 400 (for details measuring parameters, see Pospíšilová et al., 2016). We used ZooMS (Zooarchaeology by Mass Spectrometry) to identify the embedded non-human bone fragment species. Mike Buckley performed protein extractions and analyses at the University of Manchester.
The biological identification of PD8 followed the same protocol as the one applied to the entire Sphinx anthropological collection (Brukner Havelková et al., 2022). Identification of human skeletal elements of PD8 relied on classical anthropological methods (Buikstra & Ubelaker, 1994; White & Folkens, 2005). However, the state of preservation of the skeleton limited the application of the most reliable methods. For the sexual diagnosis, we used the coxal bones and cranial morphology (Brůžek, 2002; Buikstra & Ubelaker, 1994; Ferembach et al., 1980). Age at death was estimated based on dental and skeletal maturation (Moorrees et al., 1963; Scheuer & Black, 2000). The stature estimate used the length of the best-preserved long bone, following the regressions of Trotter and Gleser (1977).
Tooth enamel of the upper left second molar of PD8 was prepared for radiocarbon dating at the Environmental Isotope Paleobiogeochemistry Laboratory, University of Illinois Urbana-Champaign, using the vacuum milling technique, described in detail in Jones et al. (2021). Radiocarbon dating was performed at the Keck Carbon Cycle Accelerator Mass Spectrometry Laboratory, University of California, Irvine.
Results
PD8 Biological Identification
The state of preservation of PD8 is shown in Fig. 4. Every anatomical part is represented, including the hyoid bone. However, the preservation is not even, with long bone extremities exhibiting a fragmentary state and poor preservation due to post-depositional taphonomical processes. In contrast, other body parts, including some rather fragile ones, such as the right scapula, are better-preserved thanks to the carbonaceous crust that cemented them together.

The anatomical parts preserved on the PD8 individual
The skeleton of PD8 belongs to a robust adult, probably male (Brůžek, 2002). The individual’s age at death is estimated to be > 30 years because dental and osseous maturation is complete. The stature of PD8, based on the length of the left fibula, indicates a body height of 171.3 ± 3.9 cm (Trotter & Gleser, 1977). This individual is characterized by severe dental wear with lingual attrition of the upper left incisors (LSAMAT; Turner & Machado, 1983) and grooves and notches that suggest non-masticatory wear-inducing activities. Linear enamel hypoplasia is present on all the mandibular and maxillary teeth, including the third molars. Two space-occupying lesions are evident on the atlas’ transversal arch and the axis body’s anterior part. A small depression is present above the left orbit, and the distal part of the diaphysis of the right radius is bulging. These features may represent healed fractures or injuries whose etiology is unknown.
PD8 Trauma
A triangular fragment of non-human bone was found wedged between the rib cage and the anterior part of the subscapular fossa of the right scapula (Figs. 5 and 6). The subscapular fossa was perforated and bulged posteriorly following the axis of the apex of the fragment’s distal part at the level of contact with the scapula. Three-dimensional segmentation µ-CT scan imagery of the block containing the scapula, clavicle, ribs, and non-human bone fragment allowed us to observe the position of each element in situ and relative to others, in addition to providing internal structural information (Fig. 6). Notably, the faunal fragment exhibits lines of fractures that were segmented similarly to the human bones.

Photograph of the cemented block of the right upper part of PD8 body containing the shoulder girdle, costal remains, and the bone fragment: (a) anterior view during the cleaning, the color of the artifact is enhanced because of the use of water; (a1) lateral view with a focus on the area where the faunal piece is embedded; (b) anterior view of the faunal piece showing the broken distal tip; (c, d) anterior and posterior views of the right scapula and ribs after cleaning and removal of the non-human bone fragment

Combined photographs of the lateral sides and proximal border views of the faunal piece (photo Martin Frouz). The black bar represents 1 cm
The first area of interest is the tip of the fragment, in contact with the scapula. This part exhibits a V-shaped plan of fracture originating from its broken extremity. The second set of fractures is visible along the proximal broken border of the bone: three planes are present at the top, in the middle, and at the bottom. Regarding the fracture on the scapula, the perforation at the level of the bulging and the triangular uplifting of two bone fragments close to the bulging is in direct relationship with the penetration of the faunal fragment (Fig. 7). In addition, some of the fracture lines visible under the acromial spine of the scapula and diffusing toward the glenoid fossa and the base of the spine may be related to the same event. Their pattern is also consistent with descriptions of past and present-day scapula fracture types (e.g., Bartoníček et al., 2018, 2021; Blondiaux et al., 2012; Südkamp et al., 2011; Tuček et al., 2017). However, with these latter fracture lines, post-depositional taphonomical processes and poor bone preservation call for caution in identifying their origin. The same caution applies to characterizing the fracture pattern seen on the posterior side of the right humeral head fragment.

Composite image of the three-dimensional reconstruction of the non-human bone fragment and its fracture pattern, and PD8 bones with variation in transparency and orientation to illustrate the position of each part relative to one another: (a) lateral view; (b) antero-lateral view; (c) anterior view; (d) anterior view without the ribs; (e) antero-medial view with the ribs in transparency; (f) medial view; (g) posterior view; (h) posterior view with the scapula in transparency
The Embedded Non-human Bone
The object embedded in the subscapular fossa is a fragment of a flat bone, possibly part of an ulna from a large mammal or a scapula or pelvis from a medium-sized mammal. More precise taxonomical identification of the bone is limited by the shape and size preserved. Protein extraction and analysis by mass spectrometry were unsuccessful due to degradation of proteins.
The tip of the faunal fragment in contact with the subscapular fossa is broken, and V-shaped lines of fractures diffuse proximally from this broken area. Fracture lines are also present on the proximal side of the intrusive bone that may be related to the breaking of the piece during or after impact. Macroscopic and microscopic observations of the bone surface indicate excellent preservation of the natural surface without any traces of modification such as shaping, use wear, or hafting, excluding the hypothesis of an intentionally modified bone that could be used as a tool or a weapon.
This bone fragment exhibits a brown–red color on its surface and a heterogeneous dark pigmentation on the superior part of its proximal border, distinguishing it from the white–gray human bones. XRF elemental analysis allowed us to eliminate the presence of manganese (Mn), or another dominant element, as the primary source of coloration of the object (Fig. 8). The results are consistent with fossil bone hydroxyapatite (calcium phosphate) composition (Keenan et al., 2015).

X-ray fluorescence results with the photographic image of the area measured (Conc. %, concentration of the element in percent)
One hypothesis to explain this dark coloration is that the bone was exposed to heat. Experimental studies have shown that heat treatment of bone triggers crystallinity changes (Munro et al., 2007; Reiche et al., 2013; Shipman et al., 1984), with the size of hydroxyapatite crystal being positively correlated with the increasing heating temperature. This change in bone crystallinity happens even at low temperatures (< 525 °C) and is detectable by X-ray analysis of bone density variation (Boschin et al., 2015; Postnov et al., 2003; Shipman et al., 1984). The hypothesis that the bone fragment embedded in the subscapular fossa of PD8 was modified by heating is supported by its black staining and the X-ray microtomographic data showing a lighter gray pixel that reflects a higher bone density of the faunal fragment compared to the human bones of PD8 (Fig. 9). The homogeneous pattern exhibited by the non-human fragment, which preserves both cortical and medullar parts, indicates fire-induced modification.

Composite image of the 3D reconstruction of the cemented block containing the faunal fragment embedded in the right scapular fossa of PD8 (right); and the projection (left) of the ortho-slice (red line) illustrating the difference in bone density between the non-human and human bones
Dating
Radiocarbon AMS dates on tooth enamel bioapatite of fourteen individuals from the Sphinx burial ground set a well-defined cemetery phase at the site between 8720 and 7867 cal BP, with 95.4% probability (Varadzinová et al., 2023). PD8 was directly dated and gave an uncalibrated date of 7795 ± 30 BP (UCIAMS-227174). Tree-ring calibration modeled with OxCal v4.4.4 using the IntCal20 calibration curve (Reimer et al., 2020) produced a calibrated date for this individual at 8637–8463 cal BP with 95.4% probability. The non-human bone fragment associated with PD8 could not be directly dated because no collagen is preserved, and bone tissue is more sensitive to contamination than enamel in bioapatite analysis. In addition, the heat treatment may also have affected its stable isotope and radiocarbon composition (Snoeck et al., 2014).
Discussion
Case of Violence
The remains of the PD8 individual are dated to 8637–8463 cal BP. They belong to a robust adult male over 30 years of age. The skeleton exhibits clear evidence of trauma on the anterior part of the right scapula caused by a piece of a non-human bone that entered the body laterally, underneath the right shoulder. The bulging and perforation of the subscapular fossa observed on PD8 suggest that insertion of the burned bone fragment must have occurred at or near the time of death when the scapula bone was fresh. The insertion came with powerful forces and pierced the soft tissues of the shoulder girdle that is protected by heavy muscles (Blondiaux et al., 2012). Post-depositional insertion of the object in the shoulder girdle of PD8 is ruled out because such a taphonomical introduction would have impacted the position of the ribs. However, the ribs are in place, maintaining the natural flattening of the chest classically recorded in archaeothanatology after body decomposition (Duday, 2009). Scapular fractures are uncommon (between ~ 1 and 2% of fractures) and are often associated with multi-system injuries during high-energy trauma (Bartoníček & Cronier, 2010; Blondiaux et al., 2012; Ideberg et al., 1995; Südkamp et al., 2011; Tatro et al., 2019). The absence of other unhealed trauma on PD8’s right clavicle, ribs, and other body parts is consistent with the observation that the bone fragment was inserted due to a sharp force trauma.
The inserted fragment is a piece of a flat bone from a medium-sized or large mammal concealed in the shoulder girdle without projecting out of it. It seems to constitute the tip of an originally larger object whose shape and size remain unknown. It is likely to have broken off from the larger object during or after impact, as suggested by the fracture lines on the proximal side of the fragment. However, the absence of other pieces of the non-human bone weapon in the sediment surrounding the human remains suggests that breakage likely occurred prior to burial. Two aspects of the faunal fragment are of interest. The absence of any traces of shaping, use wear, or hafting indicates the bone was not modified or used as a tool or weapon. In the Early Khartoum culture, within which the find fits, Arkell (1949) proposed the use of unmodified bones as tools but did not provide direct evidence. Thus, while formal bone tools are well-attested in Early Khartoum culture sites (e.g., Arkell, 1949; Caneva, 1983; Clark, 1989; Fernández et al., 2003; Garcea, 2020; Haaland & Magid, 1995; Varadzinová et al., 2022a), informal bone tools still constitute an undefined category of finds. However, this piece shows traces of prior fire-induced heating at low temperatures. Modification of bones by heat for mechanical advantage or coloration and shine has been documented in numerous contexts and regions, but it is usually carried out on formal tools at the end of the manufacturing sequence (e.g., Campana, 1989; Newcomer, 1974; Sidéra, 2000; Stordeur, 1988). In Early Khartoum contexts, only one fragment of a small pointed and polished bone implement from an undetermined animal has been reported to be heat modified (Arkell, 1949, p. 77, pl. 53, Fig. 6), but it is not clear whether the burning was for functional or aesthetic reasons, or unintentional. At Sphinx, the settlement debris contains numerous bone fragments showing burning traces probably related to cooking and waste removal (Sůvová, 2017). Therefore, the absence of other modification traces on the bone fragment found in PD8 makes it impossible to determine whether it was subject to accidental or intentional heating.
PD8 was deposited with earlier burials in a manner that did not differ significantly from other burials at Sphinx. Other individuals were later buried nearby, attesting that PD8 was part of the same community. Among the 45 individuals examined at this site, six exhibit healed traumatic lesions that were most likely of an accidental origin linked to habitual activities (e.g., healed fractures on the ribs, forearms, hand bones, and pelvis; see Table 1; Brukner Havelková et al., 2022). The trauma on PD8 is the only perimortem penetrating trauma documented at Sphinx. The PD8 case is unusual. No occurrences of traumatic injury to the scapula or use of (fragments of) bones to inflict such trauma have been ascertained so far in any hunter-gatherer burial assemblage from Northeast Africa. Documentation of scapular fractures in the Nile Valley comes from later periods (Blondiaux et al., 2012; Dabbs & Zabecki, 2015) and includes five individuals at Amarna in Egypt during the second millennium BCE that present similar sharp force trauma on the scapula. This was interpreted as evidence of ritualized punishment by stabbing (Dabbs & Zabecki, 2015).
The unorthodox nature of the object found beneath the shoulder blade of PD8 supports an opportunistic use of the pointed heated bone as a weapon at a close range. This evidence supports the hypothesis of an occasional intra- or inter-community lethal conflict, which is ethnographically attested even among small and relatively peaceful hunter-gatherer groups in other parts of Africa (e.g., Lee, 1979; Marshall, 1976).
Early Holocene and Terminal Pleistocene Violence in the Nile Basin
A total of 17 burial sites containing more than 265 burials have been reported from early Holocene hunter-gatherer burial sites in the Nile Valley (Table 1; Fig. 1). The sites of El-Barga and Wadi el-Arab represent Mesolithic occupations near the Third Nile Cataract in northern Sudan (Honegger, 2014; Honegger & Williams, 2015). The site of Al Khiday 2 contains a pre-Mesolithic (> 8750 cal BP) phase of occupation in the lower White Nile (Dal Jakob, 2014; Sasso et al., 2014; Usai et al., 2010). The remaining sites are attributed to the Early Khartoum culture in Central Sudan (Garcea, 2020). Based on published reports, more than 200 Early Holocene individuals have been analyzed by anthropologists. Traces of trauma have been identified on 17 individuals. This number must be treated as the minimum because data on taphonomy and the degree of coating of the human remains by calcareous crusts and their laboratory treatment methods are not always described in publications. Among Holocene hunter-gatherer burials in Central Sudan, PD8 is the only individual with demonstrable traces of interpersonal violence. El-Barga in northern Sudan is the only other Early Holocene site with evidence of perimortem trauma that has been ascertained on two individuals (Table 1).
The Early Khartoum culture in Central Sudan and the Mesolithic occupation at the Third Nile Cataract in northern Sudan are characterized by high densities of sites, reduced residential mobility, and centuries- or even millennia-long occupation of selected locations (e.g., Caneva et al., 1993; Fernández et al., 2003; Garcea, 2020; Honegger, 2014; Honegger & Williams 2015; Salvatori, 2012; Salvatori & Usai, 2019; Suková & Varadzin, 2012; Varadzinová & Varadzin, 2017; Varadzinová et al., 2021, 2023). A broad spectrum of food resources was exploited. Plant food processing was prominent as suggested by abundant grinding and milling tools (Caneva, 1983; Haaland & Magid 1995; Řídký et al., 2022), and by intensive human tooth wear (Brukner Havelková et al., 2022; Jakob, 2014). Hunting of aquatic, riverine, and savanna animals, including large herbivores, collection of aquatic molluscs (Gautier 1983; Peters, 1996), and fishing, using specialized fishing gear such as bone harpoons and hooks (Arkell, 1949; Caneva, 1983; Clark, 1989; Haaland & Magid, 1995) provided the dominant sources of animal protein (Linseele & Zerboni, 2018). Resource exploitation was likely seasonally scheduled (Peters, 1996), with surpluses stored for delayed return (Salvatori & Usai, 2019). These facts clearly attest to the relative wealth and stability of the food resource base of the societies that occupied the Sudanese Nile during the climatic optimum of the African Humid Period (AHP) (deMenocal et al., 2000; Holmes & Hoelzmann, 2017; Ménot et al., 2020).
From the previous period, the Terminal Pleistocene, interpersonal violence has been reported from three of the four burial sites known in the Nile Valley (Table 2; Fig. 1). The most emblematic case comes from Site 117 in Jebel Sahaba, near the Second Nile Cataract, dated broadly between 13,400 and 18,600 cal BP (Crevecoeur et al., 2021; Wendorf, 1968; Zazzo 2014). Here, 64 individuals in contracted position in single, double, or multiple burials were discovered (Wendorf, 1968). Recent reassessment of the anthropological and archaeological evidence from Sahaba confirms that two-thirds of the individuals were injured, and most of the lesions were related to projectile impact marks and interpersonal violence acts (Crevecoeur et al., 2021). In addition, the identification of healed and unhealed trauma on one-fourth of the affected individuals, together with the demographic profile of the Sahaba funerary assemblage, strongly supports the evidence of recurrent episodes of small-scale conflicts such as raids or feuds in the Nile Valley (Crevecoeur et al., 2021). Similar observations of embedded lithics and drag marks were described on two of the 36 individuals from the penecontemporaneous cemetery at site 6-B-36 in Wadi Halfa at the Second Nile Cataract, one of whom also exhibits a healed parry fracture of the forearm (Greene & Armelagos, 1972; Hewes et al., 1964; Saxe, 1971). From the last cemetery of this area at Tushka (Site 8905), with 19 individuals attributed to the Qadan (20,200–12,000 cal BP), no evidence of perimortem trauma has been reported, but their state of preservation is extremely poor with the cortical surface often eroded or embedded in hard matrix (Anderson, 1968). In addition, another case of death caused by a projectile impact was described at site E-82–6 at Wadi Kubbaniya near the First Nile Cataract, dated between 23,500 and 19,300 cal BP (Angel & Kelley, 1986; Leplongeon, 2021; Wendorf & Schild, 1986). The latter case is a single burial that resembles the cases of Jebel Sahaba and Wadi Halfa, with a healed forearm parry fracture and lithic projectiles within the skeleton (Table 2).
The cemeteries at Jebel Sahaba, Wadi Halfa, and Tushka fall within a refugial phase of occupation of the middle Nile near the Second Nile Cataract during the end of the Pleistocene (Crevecoeur et al., 2021; Leplongeon 2021). At that time, the environmental setting along the Nile was characterized by reliable water within the river valley, surrounded by a hyper-arid desert (Schild & Wendorf 2010; Vermeersch & Van Neer, 2015). Seasonal site-occupation intensity points to high mobility and clustering of many foraging groups in a restricted area (Becker & Wendorf, 1993; Van Neer et al., 2000; Yeshurun, 2018). Terrestrial and aquatic faunal assemblages suggest intensive seasonal fishing, occasional hunting, and grinding activities (Linseele & Van Neer, 2010; Roubet, 1989; Van Neer, 2004). The case of violence in Wadi Kubbaniya during the Last Glacial Maximum may also fall within a context of Terminal Pleistocene environmental scarcity in the area adjacent to the Nile Valley (Vermeersch & Van Neer, 2015; Wendorf & Schild, 1986).
The upper Nile basin has yielded another, quite specific case of collective violence—a massacre—at Nataruk at Lake Turkana (Kenya), where ten of twelve individuals—men, women, and children—were found to have been killed roughly around 10 ka (Lahr et al., 2016a). However, this evidence has been contested on taphonomic grounds by Stojanowski et al., (2016; cf. Lahr et al., 2016b). The excavators have offered two alternative interpretations of the massacre: a raid for resources during a period of high population density and resource competition, or an antagonistic response to an encounter between two social groups (Lahr et al., 2016a).
Patterns of Violence in the Terminal Pleistocene and Early Holocene Nile Basin
The qualitative and quantitative data gathered above reveal two contrasting patterns of violence in the Nile Valley at the end of the Upper Pleistocene and the beginning of the Holocene. The first pattern is represented by the evidence from Early Holocene sites that indicates very low frequencies of perimortem (1.5%) and healed (7.5%) traumas among > 200 skeletons. The only case of interpersonal violence documented so far comes from Sphinx where it was related to a close-range trauma, inflicted with an unorthodox weapon. It more likely reflects an occasional intra-community feuding than inter-community conflicts. A hypothesis of ongoing reciprocal interpersonal violence between communities would require more than one individual with interpersonal trauma. This incident occurred during the AHP, which was characterized by favorable environmental conditions, with sustained occupation of large settlements, supported by broad-spectrum terrestrial and aquatic food resource exploitation.
The second pattern is represented by the Terminal Pleistocene sites, with very high frequencies of perimortem (16.2%) and healed (35.9%) traumas among the 117 analyzed individuals. Many of them are connected to systematic intergroup attacks using bows and arrows, or spears, i.e., weapons designed and intended for killing from a distance. Parry fractures and blunt force trauma on the skull or the upper body of several individuals at Jebel Sahaba (Anderson, 1968; Crevecoeur et al., 2021; Greene & Armelagos, 1972) also indicate combat at close quarters. This evidence and the high frequencies of healed bone fractures strongly suggest frequent organized intergroup violence—that is, chronic and debilitating warfare.
This second pattern falls within a period of environmental instability in the refuge zone around the First and Second Nile Cataracts during and after the Last Glacial Maximum (23–15 ka) and the initial Terminal Pleistocene phase of the AHP (15–13 ka). The desert environment outside of the Nile Valley may have been characterized by limited availability of food resources, forcing circumscription, and an apparent increase in population density ca. 23–20 ka cal BP, as at Wadi Kubbaniya, and, again ca. 16–14 ka in the Nile Valley with seasonal fishing in lakes created by damming of the Nile as a result of intense eolian activity (Connor & Marks, 1986; deMenocal et al., 2000; Dufour et al., 2018; Holl, 2013; Kuper & Kröpelin, 2006; Leplongeon, 2021; Mayewski et al., 2004; Vermeersch & Van Neer, 2015; Williams et al., 2006).
After the climatic amelioration at the beginning of the AHP, and also during the Younger Dryas (YD) cold, arid period, ca. 12.8–11.6 ka (deMenocal et al., 2000; Foerster et al., 2015; Ménot et al., 2020), traces of human occupation are sparse in the Nile Valley (Leplongeon, 2021; Vermeersch & Van Neer 2015). However, major floods and subsequent incision of the Nile during the YD may be responsible for the absence of occupations during that period (Butzer, 1997). The main Early Holocene phase of the AHP, characterized by more humid conditions between 11.6 and 7.3 ka, witnessed dramatic increases in population density in the Sudanese part of the middle Nile Valley, by peoples with a remarkably different approach to intra- and intergroup conflict resolution.
The Early Khartoum culture is attested in the Sudan as early as 10.9 ka (Varadzinová et al., 2023), which places its origin close to the onset of the post-YD AHP. If the lower Nile Valley sites evincing systematic interpersonal violence all date to the early AHP humid phase before the YD, then a systematic comparison of the evidence for environment, technology, subsistence, and social and territorial organization may reveal the reasons for this difference in patterns of violence between these periods. More sites spanning the early AHP through the YD and the earliest Holocene phase of the AHP, as well as more accurate and precise chronometric dates, are needed to securely establish whether the pre- and post-YD differences in patterns of violence correlate with major paleoenvironmental regimes.
Both patterns appear to represent two extremes in the continuum of possible configurations of types and frequencies of violence, from rare interpersonal to chronic intergroup conflict, which may have even threatened the existence of the group buried at Jebel Sahaba. Could transformations in social and territorial organization during the YD play a role? Archaeological and ethnographic evidence suggests that environmental crises may have stimulated hunter-gatherer societies to make social or technological innovations such as changes in settlement system, subsistence strategies, and creation of intergroup risk-reducing alliances and exchange networks (Ambrose, 2002; Ambrose & Lorenz, 1990; Dyson-Hudson & Smith, 1978; Kusimba, 1999). But context is important: The mortuary evidence for the middle and lower Nile indicate the social mechanisms and structures mediating dispute resolution likely differed between pre- and post-YD societies. Environmental conditions in the Early Holocene Nile Valley and surrounding green Sahara may have provided a greater space for conflict resolution, including the option of secession and migration from the larger groups, which is frequently attested among ethnographic foraging groups (Lee, 1979; Lomas, 2008-2009; Marshall, 1976). By contrast, in the Lower Nubian Terminal Pleistocene refugia, a certain threshold might have been crossed, perhaps due to territorial circumscription within the Nile Valley, beyond which prevention or mitigation of violence may have been considered disadvantageous or impossible.
Cross-cultural ethnographic and primate ethological analyses of the relationships of post-marital residence and exogamy patterns to patterns of conflict and cooperation may provide insights into the remarkable differences between Terminal Pleistocene and Early Holocene patterns of violence along the Nile (Chapais, 2008, 2011; Hill et al., 2011; Kang, 1979; Knauft, 1991; Lévi-Strauss, 1969; Manson & Wrangham, 1991; Rodseth et al., 1991). The most notable pattern emerging from these analyses is that the lowest incidence of intergroup violence is found among societies with predominantly male exogamy (matrilocal post-marital residence) (Kang, 1979; Knauft, 1991; Rodseth et al., 1991). Exogamy promotes enduring intergroup networks of mutual reliance among unrelated and distantly related individuals, families, and clans. Rodseth et al. (1991, p. 230) explain the extremely low incidence of intergroup conflict among matrilocal societies as follows: “With relations between neighboring matrilocal groups friendly and open, men are rarely cut off from their consanguineal kin in the way women may be under patrilocality.” In other words, with male exogamy, males are unlikely to engage in feuds with members of other groups because they have enduring relationships with relatives in other groups. Cross-cultural analysis of post-marital residence patterns in a global sample of 32 forager societies (Hill et al., 2011) explodes the myth of predominantly patrilocal, patrilineal band organization. Their analysis reveals predominantly neolocal residence, and strong and enduring relationships with parents of marriage partners in their respective natal communities. This simple post-marital residence pattern results in a dense, spatially extensive, and enduring network of supportive intergroup relationships that effectively discourages intergroup conflict. Neolocal post-marital residence extends the effect of matrilocality on reducing intergroup conflict.
The results of these studies bring the role of the deep structural foundation of kinship reckoning and post-marital residence patterns in strategies of conflict mediation into sharp focus. They suggest new questions and hypotheses regarding the contrasts in interpersonal and intergroup violence patterns between the Terminal Pleistocene and Early Holocene in the Nile Valley. Were Terminal Pleistocene foraging groups predominantly patrilocal and early Holocene groups predominantly matrilocal and neolocal? Did social strategies of coping with extreme environmental stress during the intervening Younger Dryas promote the adoption of matrilocal and neolocal post-marital residence and promote enhanced cooperation among groups? Can the bioarchaeological and archaeological records of the Terminal Pleistocene and Early Holocene Nile Valley provide evidence to test hypotheses of contrasting patterns of exogamy, post-marital residence, and intergroup interaction networks?
Lower variance in the morphology of female skeletons has been interpreted as evidence for matrilocal post-marital residence (Konigsberg, 1988; Konigsberg & Frankenberg, 2016). When preserved, ancient DNA would provide the best evidence for exogamy and post-marital residence patterns (Konigsberg & Frankenberg, 2016; Lipson et al., 2022; Mittnik et al., 2019). Strontium and lead isotope analysis of tooth enamel can provide evidence for residential life history (Buzon & Simonetti, 2013; Gulson et al., 2003; Hedman et al., 2018; Mittnik et al., 2019). Artifact geochemical provenience and provenance analyses provide another productive avenue for evaluating the geographic scale and intensity of intergroup interaction networks (Zipkin et al., 2020). As noted above, more accurate and precise chronometric dating is essential for evaluating hypotheses that posit correlation and causation with the paleoenvironmental record.
Conclusion
The unusual case of traumatic violence evinced by burial PD8 at Sphinx leads to a broader set of questions regarding how the environmental, economic, kinship, and territorial organizations correlate to the contrasting patterns of interpersonal and intergroup violence among Terminal Pleistocene and Early Holocene hunter-gatherer groups far beyond the middle and lower Nile Valley of Sudan and southern Egypt. The murder weapon lodged beneath the shoulder blade of PD8—a naturally fractured, heat-altered bone—would have been considered consistent with Raymond Dart’s (1953) now-discounted Osteodontokeratic tool and weapon technology wielded by fire-using Australopithecus prometheus during the predatory transition from ape to human. Ironically, PD8 was likely a member of a widespread set of communities in which skeletal evidence for trauma was extremely rare and warfare seemingly unknown. This burial represents the exception to what has been widely accepted as evidence for the atavistic evolutionary propensity for human violence inherited from the last common ancestor of human and apes (Kim & Kissel, 2018). Rather than exceptional, it may exemplify the power of matrilocal and neolocal configurations of exogamy and post-marital residence to promote peaceful, cooperative intergroup interactions.
Data Availability
All data generated and analyzed during the current study are included in the article and the supplementary information.
Change history
08 October 2023
The original version of this paper was updated to correct the project number in the funding statement.
References
Ambrose, S. H., & Lorenz, K. G. (1990). Social and ecological models for the Middle Stone Age in southern Africa. In P. Mellars (Ed.), The emergence of modern humans: An archaeological perspective (pp. 3–33). University of Edinburgh Press.
Ambrose, S. H. (2002). Small things remembered: Origins of early microlithic industries in Sub-Saharan Africa. In R. G. Elston & S. L. Kuhn (Eds.), Thinking small: Global perspectives on microlithization (pp. 9–29). Archaeological Papers of the American Anthropological Association 12.
Anderson, J. E. (1968). Late Paleolithic skeletal remains from Nubia. In F. Wendorf (Ed.), The prehistory of Nubia (Vol. II, pp. 996–1040). Southern Methodist University Press.
Angel, J. L., & Kelley, J. O. (1986). Description and comparison of the skeleton. In F. Wendorf & R. Schild (Eds.), The prehistory of Wadi Kubbaniya (Vol. I, pp. 53–70). Southern Methodist University Press.
Ardrey, R. (1961). African genesis: A personal investigation into the animal origins and nature of man. Atheneum.
Ardrey, R. (1966). The territorial imperative: A personal inquiry into the animal origins of property and nations. Atheneum.
Ardrey, R. (1976). The hunting hypothesis: A personal conclusion concerning the evolutionary nature of man. Atheneum.
Arkell, A. J. (1949). Early Khartoum. An account of the excavation of an early occupation site carried out by the Sudan Government Antiquities Service in 1944–5. Oxford University Press.
Bartoníček, J., & Cronier, P. (2010). History of the treatment of scapula fractures. Archives of Orthopaedic and Trauma Surgery, 130(1), 83–92.
Bartoníček, J., Klika, D., & Tuček, M. (2018). Classification of scapular body fractures. Rozhledy v Chirurgii, 97(2), 67–76.
Bartoníček, J., Tuček, M., & Naňka, O. (2021). Zlomeniny Lopatky. Maxdorf, Praha.
Becker, M., & Wendorf, F. (1993). A microwear study of a late Pleistocene Qadan assemblage from Southern Egypt. Journal of Field Archaeology, 20(4), 389–398.
Blondiaux, J., Fontaine, C., Demondion, X., Flipo, R.-M., Colard, T., Mitchell, P. D., et al. (2012). Bilateral fractures of the scapula: Possible archeological examples of beatings from Europe, Africa and America. International Journal of Paleopathology, 2, 223–230.
Boschin, F., Zanolli, C., Bernardini, F., Princivalle, F., & Tuniz, C. (2015). A look from the inside: MicroCT analysis of burned bones. Ethnobiology Letters, 6, 258–266. https://doi.org/10.14237/ebl.6.2.2015.365
Brukner Havelková, P., Crevecoeur, I., Villotte, S., Velemínský, P., Varadzin, L., & Varadzinová, L. (2022). Palaeodemography and palaeopathology of Khartoum Mesolithic skeletal remains from Jebel Sabaloka in central Sudan: First insights from the site of Sphinx. Journal of the National Museum (Prague). Natural History Series, 191, 65–82.
Brůžek, J. (2002). A method for visual determination of sex, using the human hip bone. American Journal of Physical Anthropology, 117, 157–168. https://doi.org/10.1002/ajpa.10012
Buikstra, J. E., & Ubelaker, D. H. (1994). Standards for data collection from human skeletal remains. Arkansas Archaeological Survey.
Butzer, K. W. (1997). Late Quaternary problems of the Egyptian Nile: Stratigraphy, environments, prehistory. Paléorient, 23, 151–173. https://doi.org/10.3406/paleo.1997.4658
Buzon, M. R., & Simonetti, A. (2013). Strontium isotope (87Sr/86Sr) variability in the Nile Valley: Identifying residential mobility during ancient Egyptian and Nubian sociopolitical changes in the New Kingdom and Napatan periods. American Journal of Physical Anthropology, 151(1), 1–9.
Campana, D. V. (1989). Natufian and Protoneolithic bone tools. The manufacture and use of bone implements in the Zagros and the Levant. BAR International Series 494. Archaeopress.
Caneva, I., Garcea, E. A. A., Gautier, A., & Van Neer, W. (1993). Pre-pastoral cultures along the central Sudanese Nile. Quaternaria Nova, 3, 177–252.
Caneva, I. (1983). Excavating Saggai 1. In I. Caneva (Ed.), Pottery using gatherers and hunters at Saggai (Sudan): Preconditions for food production (pp. 7–29) Origini 12. Roma: Università degli Studi «La Sapienza».
Chapais, B. (2008). Primeval kinship. Harvard University Press.
Chapais, B. (2011). The deep social structure of humankind. Science, 331, 1276–1277.
Clark, J. D. (1989). Shabona: An early Khartoum settlement on the White Nile. In L. Krzyżaniak & M. Kobusiewicz (Eds.), Late Prehistory of the Nile Basin and the Sahara (pp. 387–410). Studies in African Archaeology 2. Poznań Archaeological Museum.
Connor, D. R., & Marks, A. E. (1986). The terminal Pleistocene on the Nile: The final Nilotic adjustment. In L. G. Straus (Ed.), The end of the paleolithic in the Old World (pp. 171–199). BAR International Series 284. Archaeopress.
Coppa, A. & Macchiarelli, R. (1983). Human skeletal remains from the Mesolithic site of Saggai 1 (Sudan). A descriptive and comparative anthropological study. In I. Caneva (Ed.), Pottery using gatherers and hunters at Saggai (Sudan): Preconditions for food production (pp. 116–139) Origini 12. Università degli Studi «La Sapienza».
Crevecoeur, I. (2012). First anthropological insights on the early Holocene funerary assemblages from El-Barga. Kerma: Documents de la Mission Archéologique Suisse au Soudan, 4, 19–28.
Crevecoeur, I., Dias-Meirinho, M.-H., Zazzo, A., Antoine, D., & Bon, F. (2021). New insights on interpersonal violence in the Late Pleistocene based on the Nile Valley cemetery of Jebel Sahaba. Scientific Reports, 11, 9991. https://doi.org/10.1038/s41598-021-89386-y
Cullen, T. (1995). Mesolithic mortuary ritual at Franchthi Cave, Greece. Antiquity, 69, 270–289.
Dabbs, G. R., & Zabecki, M. (2015). Slot-type fractures of the scapula at New Kingdom Tell El-Amarna. Egypt. International Journal of Paleopathology, 11, 12–22. https://doi.org/10.1016/j.ijpp.2015.07.003
Dal Sasso, G., Maritan, L., Usai, D., Angelini, I., & Artioli, G. (2014). Bone diagenesis at the micro-scale: Bone alteration patterns during multiple burial phases at Al Khiday (Khartoum, Sudan) between the Early Holocene and the II century AD. Palaeogeography, Palaeoclimatology, Palaeoecology, 416, 30–42. https://doi.org/10.1016/j.palaeo.2014.06.034
Dart, R. A. (1953). The predatory transition from ape to man. International Anthropological and Linguistic Review, 1(4), 201–207.
deMenocal, P. B., Ortiz, J., Guilderson, T., Adkins, J., Sarnthein, M., Baker, L., & Yarusinsky, M. (2000). Abrupt onset and termination of the African Humid Period: Rapid climate responses to gradual insolation forcing. Quaternary Science Reviews, 19, 347–361. https://doi.org/10.1016/S0277-3791(99)00081-5
Duday, H. (2009). The Archaeology of the dead: Lectures in archaeothanatology. Oxbow Books.
Dufour, E., Van Neer, W., Vermeersch, P. M., & Patterson, W. P. (2018). Hydroclimatic conditions and fishing practices at Late Paleolithic Makhadma 4 (Egypt) inferred from stable isotope analysis of otoliths. Quaternary International, 471, 190–202.
Dyson-Hudson, R., & Smith, E. A. (1978). Human territoriality: An ecological reassessment. American Anthropologist, 80(1), 21–41.
Ferembach, D., Schwindezky, I., & Stoukal, M. (1980). Recommendation for age and sex diagnoses of skeletons. Journal of Human Evolution, 9, 517–549.
Fernández, V. M., Jiméno, A., & Menéndez, M. (2003). Archaeological excavations in prehistoric sites of the Blue Nile area, central Sudan. In V. M. Fernández (Ed.), The Blue Nile Project. Holocene archaeology in Central Sudan (pp. 273–344). Universidad Complutense, Madrid.
Foerster, V., Vogelsang, R., Junginger, A., Asrat, A., Lamb, H. F., Schaebitz, F., et al. (2015). Environmental change and human occupation of southern Ethiopia and northern Kenya during the last 20,000 years. Quaternary Science Reviews, 129, 333–340.
French, J. C., & Nowell, A. (2022). Growing up Gravettian: Bioarchaeological perspectives on adolescence in the European mid-Upper Paleolithic. Journal of Anthropological Archaeology, 67, 101430. https://doi.org/10.1016/j.jaa.2022.101430
Fry, D. P., & Söderberg, P. (2014). Myths about hunter-gatherers redux: Nomadic forager war and peace. Journal of Aggression, Conflict and Peace Research, 6, 256–266.
Garcea, E. A. A. (2020). The prehistory of the Sudan. Springer International Publishing.
Gautier, A. (1983). Animal life along the prehistoric Nile: The evidence from Saggai 1 and Geili (Sudan). In I. Caneva (Ed.), Pottery using gatherers and hunters at Saggai (Sudan): Preconditions for food production (pp. 50–115). Origini 12. Università degli Studi «La Sapienza», Roma.
Gibaja, J. F., Subirà, M. E., Terradas, X., Santos, F. J., Agulló, L., Gómez-Martínez, I., et al. (2015). The emergence of Mesolithic cemeteries in SW Europe: Insights from the El Collado (Oliva, Valencia, Spain) radiocarbon record. PLoS One, 10(1), e0115505. https://doi.org/10.1371/journal.pone.0115505
Greene, D. L., & Armelagos, G. L. (1972). The Wadi Halfa Mesolithic population. University of Massachusetts.
Gulson, B. L., Mizon, K. J., Korsch, M. J., Palmer, J. M., & Donnelly, J. B. (2003). Mobilization of lead from human bone tissue during pregnancy and lactation: A summary of long-term research. The Science of the Total Environment, 303, 79–104.
Haaland, R. (1993). Aqualithic sites of the middle Nile. Azania, 28, 47–86.
Haaland, R., & Magid, A. A. (1995). Aqualithic sites along the rivers Nile and Atbara, Sudan. Alma Mater Forlag, Bergen.
Hames, R. (2019). Pacifying hunter-gatherers. Human Nature, 30, 155–175.
Hedman, K. M., Slater, P. A., Fort, M. A., Emerson, T. E., & Lambert, J. M. (2018). Expanding the strontium isoscape for the American midcontinent: Identifying potential places of origin for Cahokia and Pre-Columbian migrants. Journal of Archaeological Science: Reports, 22, 202–213.
Hewes, G. W., Irwin, H., Papworth, M., & Saxe, A. (1964). A new fossil human population from the Wadi Halfa area, Sudan. Nature, 203, 341–343. https://doi.org/10.1038/203341a0
Hill, K. R., Walker, R. S., Božičević, M., Eder, J., Headland, T., Hewlett, B., Hurtado, A. M., Marlowe, F., Piessner, P., & Wood, B. (2011). Co-residence patterns in hunter-gatherer societies show unique human social structure. Science, 331, 1286–1289.
Holl, A. F. C. (2013). Caring for the dead, building social ties: Behavioral implications of Late Pleistocene burials in the Sahara. Sahara, 24, 257–266.
Holmes, J., & Hoelzmann, P. (2017). The Late Pleistocene-Holocene African Humid Period as evident in lakes. Oxford Research Encyclopedia of Climate Science. https://doi.org/10.1093/acrefore/9780190228620.013.531
Honegger, M., & Williams, M. A. J. (2015). Human occupations and environmental changes in the Nile Valley during the Holocene: The case of Kerma in Upper Nubia (northern Sudan). Quaternary Science Reviews, 130, 141–154. https://doi.org/10.1016/j.quascirev.2015.06.031
Honegger, M. (2014). Recent advances in our understanding of prehistory in northern Sudan. In J. R. Anderson & D. A. Welsby (Eds.), The Fourth Cataract and beyond: Proceedings of the 12th International Conference for Nubian Studies (pp. 19–30). Peeters.
Humphrey, L., Bello, S. M., Turner, E., Bouzouggar, A., & Barton, N. (2011). Iberomaurusian funerary behaviour: Evidence from Grotte des Pigeons, Taforalt. Morocco. Journal of Human Evolution, 62(2), 261–273. https://doi.org/10.1016/j.jhevol.2011.11.003
Ideberg, R., Grevsten, S., & Larsson, S. (1995). Epidemiology of scapular fractures: Incidence and classification of 338 fractures. Acta Orthopaedica Scandinavica, 66, 395–397. https://doi.org/10.3109/17453679508995571
Jakob, T. (2014). A bioarchaeological appraisal of the human skeletal remains from El-Khiday 2, central Sudan. In J. R. Anderson & D. A. Welsby (Eds.), The Fourth Cataract and Beyond: Proceedings of the 12th International Conference for Nubian Studies (pp. 271–277). Peters.
Jones, M. B., Brandt, S. A., Henry, E. R., & Ambrose, S. H. (2021). Improved ostrich eggshell and ungulate tooth enamel radiocarbon dating methods reveal Later Stone Age occupation in arid MIS 2 southern Somalia. Journal of Archaeological Science: Reports, 36, 102844. https://doi.org/10.1016/j.jasrep.2021.102844
Jórdeczka, M., Stanaszek, L. M., Bobrowski, P., Chłodnicki, M., & Sobkowiak-Tabaka, I. (2020). Neolithic inhabitants of Khor Shambat 1, Sudan. Archaeologica Polona, 58, 135–163. https://doi.org/10.23858/APa58.2020.008
Kang, G. E. (1979). Exogamy and peace relations of social units: A cross-cultural test. Ethnology, 18, 85–99.
Keeley, L. H. (1996). War before civilization: The myth of the peaceful savage. Oxford University Press.
Keenan, S. W., Engel, A. S., Roy, A., & Bovenkamp-Langlois, G. L. (2015). Evaluating the consequences of diagenesis and fossilization on bioapatite lattice structure and composition. Chemical Geology, 413, 18–27. https://doi.org/10.1016/j.chemgeo.2015.08.005
Kelly, R. C. (2000). Warless societies and the origin of war. The University of Michigan Press.
Kim, N. C., & Kissel, M. (2018). Emergent warfare in our evolutionary past. Routledge.
Knauft, B. M. (1991). Violence and sociality in human evolution. Current Anthropology, 32, 391–428.
Konigsberg, L. W. (1988). Migration models of prehistoric postmarital residence. American Journal of Physical Anthropology, 77, 471–482.
Konigsberg, L. W., & Frankenberg, S. R. (2016). Postmarital residence analysis. In M. A. Pilloud & J. T. Hefner (Eds.) Biological distance analysis. Forensic and bioarchaeological perspectives (pp. 335–347). Elsevier.
Kuper, R., & Kröpelin, S. (2006). Climate-controlled Holocene occupation in the Sahara: Motor of Africa’s evolution. Science, 313, 803–807. https://doi.org/10.1126/science.1130989
Kusimba, S. B. (1999). Hunter-gatherer land use patterns in Later Stone Age East Africa. Journal of Anthropological Archaeology, 18, 165–200.
Lahr, M., Rivera, F., Power, R. K., Mounier, A., Copsey, B., Crivellaro, F., et al. (2016a). Inter-group violence among Early Holocene hunter-gatherers of West Turkana, Kenya. Nature, 529, 394–398.
Lahr, M., Rivera, F., Power, R. K., Mounier, A., Copsey, B., Crivellaro, F., et al. (2016b). Lahr et al. reply. Nature, 539, E10–E11.
Lee, R. B. (1979). The !Kung San: Men, women, and work in a foraging society. Cambridge University Press.
Leplongeon, A. (2021). The main Nile Valley at the end of the Pleistocene (28–15ka): Dispersal corridor or environmental refugium? Frontiers in Earth Science, 8, 607183. https://doi.org/10.3389/feart.2020.607183
Lévi-Strauss, C. (1969). The elementary structures of kinship. Beacon Press.
Linseele, V., & Van Neer, W. (2010). Exploitation of desert and other wild game in ancient Egypt: The archaeozoological evidence from the Nile Valley. In H. Riemer, F. Förster, M. Herb, & N. Pöllath (Eds.), Exploitation of desert and other wild game in Ancient Egypt (pp. 47–78). Heinrich Barth Institute.
Linseele, V., & Zerboni, A. (2018). Done with fish? A diachronic study of fishing in the Holocene Nile basin of Sudan. Quaternary International, 471, 299–240.
Lipson, M., Sawchuk, E. A., Thompson, J. C., Oppenheimer, J., Tryon, C. A., Ranhorn, K. L., et al. (2022). Ancient DNA and deep population structure in sub-Saharan African foragers. Nature, 603, 290–296.
Lomas, W. (2008). Conflict, violence, and conflict resolution in hunting and gathering societies. Totem, 17, 43–50.
Manson, J. H., & Wrangham, R. W. (1991). Intergroup aggression in chimpanzees and humans. Current Anthropology, 32, 369–390.
Marshall, L. (1976). The !Kung of Nyae Nyae. Harvard University Press.
Mayewski, P. A., Rohling, E. E., Stager, J. C., Karlén, W., Maasch, K. A., Meeker, L. D., et al. (2004). Holocene climate variability. Quaternary Research, 62, 243–255. https://doi.org/10.1016/j.yqres.2004.07.001
Ménot, G., Pivot, S., Bouloubassi, I., Davtian, N., Hennekam, R., Bosch, D., et al. (2020). Timing and stepwise transitions of the African Humid Period from geochemical proxies in the Nile deep-sea fan sediments. Quaternary Science Reviews, 228, 106071. https://doi.org/10.1016/j.quascirev.2019.106071
Mittnik, A., Massy, K., Knipper, C., Wittenborn, F., Friedrich, R., Pfrengle, S., et al. (2019). Kinship-based social inequality in Bronze Age Europe. Science, 366, 731–734.
Moorrees, C. F. A., Fanning, E. A., & Hunt, E. E. (1963). Age variation of formation stages for ten permanent teeth. Journal of Dental Research, 42, 1490–1502. https://doi.org/10.1177/00220345630420062701
Munro, L. E., Longstaffe, F. J., & White, C. D. (2007). Burning and boiling of modern deer bone: Effects on crystallinity and oxygen isotope composition of bioapatite phosphate. Palaeogeography, Palaeoclimatology, Palaeoecology, 249(1–2), 90–102. https://doi.org/10.1016/j.palaeo.2007.01.011
Newcomer, M. H. (1974). Study and replication of bone tools from Ksar Akil (Lebanon). World Archaeology, 6, 138–153.
Orschiedt, J. (2020). Violence in Palaeolithic and Mesolithic hunter-gatherer communities. In G. G. Fagan, et al. (Eds.), The Cambridge world history of violence, Vol. 1: The prehistoric and ancient worlds (pp. 58–78). Cambridge University Press
Peters, J. (1996). New light on Mesolithic resource scheduling and site inhabitation in central Sudan. In L. Krzyżaniak, K. Kroeper & M. Kobusiewicz (Eds.), Interregional contacts in the later prehistory of Northeastern Africa (pp. 381–394). Studies in African Archaeology 5. Poznań Archaeological Museum.
Pettitt, P. (2011). The palaeolithic origins of human burial. Routledge.
Pospíšilová, D., Pech, M., & Kotyk, M. (2016). The use of non-destructive instrumental methods in the determination of metal objects from the Asian collection of the Náprstek Museum. Annals of the Náprstek Museum, 37, 93–102. https://doi.org/10.1515/anpm-2017-0007
Postnov, A. A., Vinogradov, A. V., Van Dyck, D., Saveliev, S. V., & De Clerck, N. M. (2003). Quantitative analysis of bone mineral content by X-ray microtomography. Physiological Measurement, 24(1), 165–178. https://doi.org/10.1088/0967-3334/24/1/312
Reiche, I., Lebon, M., Bellot-Gurlet, L., Gourrier, A., & Zazzo, A. (2013). Potentiel informatif des ossements chauffés en contexte archéologique: Imagerie chimique, moléculaire et cristallographique à micro-échelle et analyses isotopiques (OsIRIs). Actes Du Colloque Sciences Des Matériaux Du Patrimoine Culturel, 2, 12–17.
Reimer, P., Austin, W., Bard, E., Bayliss, A., Blackwell, P., Bronk Ramsey, C., et al. (2020). The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon, 62, 725–757. https://doi.org/10.1017/RDC.2020.41
Řídký, J., Varadzin, L., & Varadzinová, L. (2022). Introduction to study of (late) prehistoric ground stone artefacts in the western part of Jebel Sabaloka in central Sudan. Památky Archeologické, 113, 5–44.
Rodseth, L., Wrangham, R. W., Harrigan, A. M., & Smuts, B. (1991). The human community as a primate society [and comments]. Current Anthropology, 32(3), 221–254.
Roubet, C. (1989). The grinding stones of site E-78-3, Wadi Kubbaniya. In F. Wendorf, R. Schild, & A. E. Close (Eds.), The prehistory of Wadi Kubbaniya (Vol. 3, pp. 473–489). Southern Methodist University Press.
Salvatori, S. (2012). Disclosing archaeological complexity of the Khartoum Mesolithic: New data at the site and regional level. African Archaeological Review, 29, 399–472. https://doi.org/10.1007/s10437-012-9119-7
Salvatori, S., & Usai, D. (2019). The Mesolithic and Neolithic in Sudan. In D. Raue (Ed.), Handbook of Ancient Nubia (Vol. 1, pp. 171–193). De Gruyter.
Saxe, A. A. (1971). Social dimensions of mortuary practices in a Mesolithic population from Wadi Halfa, Sudan. In J. A. Brown (Ed.), Approaches to the social dimensions of mortuary practices (pp. 39–57). Memoirs of the Society for American Archaeology 25. Society for American Archaeology.
Scherer, A. K. (2021). Recent research on the archaeology of war and violence. Annual Review of Anthropology, 50, 403–421.
Scheuer, L., & Black, S. (2000). Developmental juvenile osteology (2nd ed.). Academic Press.
Schild, R., & Wendorf, F. (2010). Late Palaeolithic hunter-gatherers in the Nile Valley of Nubia and Upper Egypt. In E. A. A. Garcea (Ed.), South-eastern Mediterranean peoples between 130,000 and 10,000 years ago (pp. 89–125). Oxbow Books.
Shipman, P., Foster, G., & Schoeninger, M. (1984). Burnt bones and teeth: An experimental study of color, morphology, crystal structure and shrinkage. Journal of Archaeological Science, 11(4), 307–325. https://doi.org/10.1016/0305-4403(84)90013-X
Sidéra, I. (2000). Feu et industrie osseuse. Un Marqueur Dʼidentité Culturelle. Paléorient, 26, 51–59.
Snoeck, C., Brock, F., & Schulting, R. J. (2014). Carbon exchanges between bone apatite and fuels during cremation: Impact on radiocarbon dates. Radiocarbon, 56, 591–602.
Stojanowski, C. M., Seidel, A. C., Fulginiti, L. C., Johnson, K. M., & Buikstra, J. E. (2016). Contesting the massacre at Nataruk. Nature, 539, E8–E10.
Stordeur, D. (1988). Outils et Armes en Os du Gisement Natoufien de Mallaha (Eynan), Israël. Mémoires et Travaux du Centre de Recherche Français de Jérusalem 6. Paris: Association Paléorient.
Südkamp, N. P., Jaeger, N., Bornebusch, L., Maier, D., & Izadpanah, K. (2011). Fractures of the scapula. Acta Chirurgiae Orthopaedicae Et Traumatologiae Čechoslovaca, 78, 297–304.
Suková, L., & Varadzin, L. (2012). Preliminary report on the exploration of Jebel Sabaloka (west bank), 2009–2012. Sudan & Nubia, 21, 118–131.
Sůvová, Z. (2017). The archaeozoological osteological finds from Sphinx. Sudan & Nubia, 21(43–44), 48–49.
Tatro, J., Schroder, L. K., Molitor, B. A., Parker, E. D., & Cole, P. A. (2019). Injury mechanism, epidemiology, and hospital trends of scapula fractures: A 10-year retrospective study of the National Trauma Data Bank. Injury International Journal of the Care of the Injured, 50(2), 376–381. https://doi.org/10.1016/j.injury.2019.01.017
Trotter, M., & Gleser, G. C. (1977). Corrigenda to “estimation of stature from long limb bones of American Whites and Negroes”. American Journal Physical Anthropology (1952). American Journal of Physical Anthropology, 47, 355–356. https://doi.org/10.1002/ajpa.1330470216
Tuček, M., Chochola, A., Klika, D., & Bartoníček, J. (2017). Epidemiology of scapular fractures. Acta Orthopaedica Belgica, 83, 8–15.
Turner, C. G., & Machado, L. M. C. (1983). A new dental wear pattern and evidence for high carbohydrate consumption in a Brazilian Archaic skeletal population. American Journal of Physical Anthropology, 61, 125–130. https://doi.org/10.1002/ajpa.1330610113
Usai, D., Salvatori, S., Iacumin, P., Di Matteo, A., Jakob, T., & Zerboni, A. (2010). Excavating a unique pre-Mesolithic cemetery in central Sudan. Antiquity Project Gallery, 84(323), http://www.antiquity.ac.uk/projgall/usai323
Van Neer, W. (2004). Evolution of prehistoric fishing in the Nile Valley. Journal of African Archaeology, 2, 251–269.
Van Neer, W., Paulissen, E., & Vermeersch, P. M. (2000). Chronology, subsistence and environment at the Late Palaeolithic fishing sites of Mahatma 2 and 4. In P. M. Vermeersch (Ed.), Palaeolithic living sites in Upper and Middle Egypt (pp. 271–288). Leuven University Press.
Varadzinová, L., & Varadzin, L. (2017). Exploration of the site of Sphinx (SBK.W-60): Findings of the 2014 and 2015 field seasons. Sudan & Nubia, 16, 23–49.
Varadzinová, L., & Varadzin, L. (2020). The first notes on the second Khartoum Mesolithic cemetery at Jebel Sabaloka (Sudan). Archaeologia Polona, 58, 121–133.
Varadzinová, L., Varadzin, L., Garcea, E. A. A., D’Ercole, G., Řídký, J., et al. (2021). Prehistoric research in the western part of Jebel Sabaloka (spring 2019). Der Antike Sudan, 32, 5–28.
Varadzinová, L., Varadzin, L., Brukner Havelková, P., Crevecoeur, I., & Garcea, E. A. A. (2022a). Archaeology of Holocene hunter-gatherers at the sixth Nile cataract, central Sudan / Données archéologiques sur les derniers chasseurs-cueilleurs près de la sixième cataracte du Nil au Soudan central. Bulletin Dʼarchéologie Marocaine, 27, 101–118.
Varadzinová, L., Varadzin, L., Crevecoeur, I., Kapustka, K., & McCool, J.-P. (2022b). Excavations at the prehistoric site of Fox Hill in the western part of Jebel Sabaloka (2017–2018). Sudan & Nubia, 26, 160–181.
Varadzinová, L., Varadzin, L., & Ambrose, S. H. (2023). New radiocarbon dates for postglacial reoccupation of the Sudanese Nile. Quaternary Science Reviews. https://doi.org/10.1016/j.quascirev.2023.107953
Varadzinová Suková, L., Varadzin, L., Bajer, A., Lisá, L., Pacina, J., & Pokorný, P. (2015). Tracing post-depositional processes at Mesolithic occupation sites in central Sudan: View from the site of Sphinx (SBK.W-60) at Jebel Sabaloka. Interdisciplinaria Archaeologica: Natural Sciences in Archaeology, 6(2), 133–150.
Vermeersch, P. M., & Van Neer, W. (2015). Nile behaviour and Late Palaeolithic humans in upper Egypt during the Late Pleistocene. Quaternary Science Reviews, 130, 155–167.
Washburn, S. L. (1957). Australopithecines: The hunters or the hunted? American Anthropologist, 59, 612–614.
Wendorf, F. (1968). The prehistory of Nubia. Southern Methodist University Press.
Wendorf, F., & Schild, R. (1986). The prehistory of Wadi Kubbaniya, Volume 1: The Wadi Kubbaniya skeleton--A Late Paleolithic burial from southern Egypt. Southern Methodist University Press.
White, T., & Folkens, P. (2005). The human bone manual. Elsevier Academic Press.
Williams, M. A. J. (2009). Late Pleistocene and Holocene environments in the Nile basin. Global and Planetary Change, 69, 1–15. https://doi.org/10.1016/j.gloplacha.2009.07.005
Williams, M., Talbot, M., Aharon, P., Abdl Salaam, Y., Williams, F., & Brendeland, K. I. (2006). Abrupt return of the summer monsoon 15,000 years ago: New supporting evidence from the lower White Nile Valley and Lake Albert. Quaternary Science Reviews, 25, 2651–2665. https://doi.org/10.1016/j.quascirev.2005.07.019
Yeshurun, R. (2018). Taphonomy of old archaeofaunal collections: New site-formation and subsistence data for the Late Paleolithic Nile Valley. Quaternary International, 471, 35–54.
Zazzo, A. (2014). Bone and enamel carbonate diagenesis: A radiocarbon prospective. Palaeogeography, Palaeoclimatology, Palaeoecology, 416, 168–178. https://doi.org/10.1016/j.palaeo.2014.05.006
Zipkin, A. M., Ambrose, S. H., Lundstrom, C. C. Bartov, G., Dwyer, A., & Taylor, A. H. (2020). Red earth, green glass, and compositional data: A new procedure for solid-state elemental characterization, source discrimination, and provenience analysis of ochres. Journal of Archaeological Method and Theory, 27, 930–970. https://link.springer.com/article/https://doi.org/10.1007/s10816-020-09448-9
Acknowledgements
We thank the National Corporation for Antiquities and Museums of Sudan for their long-term support of the Sabaloka (West Bank) Research Project and for allowing our mission to transport archaeological materials, especially the remains of PD8, to Prague for cleaning, documentation, and analyses. We are grateful to Ronan Ledevin, who performed the X-ray microtomographic acquisition of the cemented block in the PLACAMAT platform of Bordeaux University, and to Prof. MUDr. Jan Bartoníček, DrSc., for his precious expertise and comments. Dr. John Southon is acknowledged for high-precision AMS radiocarbon analyses. Finally, we thank Ondřej Kohout for his help in the XRF analyses.
Funding
Open access publishing supported by the National Technical Library in Prague. This work was supported by the Czech Science Foundation through the research project “Communities and Resources in Late Prehistory of Jebel Sabaloka, Central Sudan: From Analysis to Synthesis” (GAČR 17-03207S), by the Ministry of Culture of the Czech Republic (DKRVO 2019–2023/7.II.d-e, National Museum, 00023272), the Cooperation Program provided by Charles University, research area Archaeology, implemented at the Faculty of Arts of Charles University, the project “Big Dry” of the French Agence Nationale de la Recherche (ANR-14-CE31), and the International Research Project (IRP) ABASC (2020–2024) founded by the CNRS-INEE. IC has benefited from the SMI grant from the CNRS-INSHS to visit Prague in 2019. The research also benefited from the scientific framework of the University of Bordeaux’s IdEx “Investments for the Future” Programme/GPR “Human Past.”
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Archaeological Time Period
Late Stone Age, Early Holocene, Early Khartoum culture, Mesolithic.
Country and Region Discussed
Northeastern Africa, Nile Valley, Central Sudan, Sixth Nile Cataract.
Supplementary information
Below is the link to the electronic supplementary material.
Video of the 3D model of PD8 right upper shoulder and the embedded non-human bone fragment. (MPG 62.8 MB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brukner Havelková, P., Crevecoeur, I., Varadzin, L. et al. Patterns of Violence in the Pre-Neolithic Nile Valley. Afr Archaeol Rev 40, 597–619 (2023). https://doi.org/10.1007/s10437-023-09533-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10437-023-09533-w