
Abstract
Al-Khiday, located on the bank of the White Nile in Sudan, offers an exceptionally preserved stratigraphic sequence, providing a unique opportunity to use organic residue analysis to investigate diet and subsistence during the Khartoum Mesolithic and the Early Neolithic, a period of nearly 3500 years (7000–4500 cal BC). While the vast and diverse Mesolithic fish assemblage indicates a strong reliance on products from aquatic habitats, floodplains, vegetated marshes, and open water, results from the lipid residue analysis suggest that the fish were not cooked in ceramic pots, but consumed in other ways. Rather, pots were more specialized in processing plants, including wild grasses, leafy plants, and sedges. These results, confirmed by experimental analysis, provide, for the first time, direct chemical evidence for plant exploitation in the Khartoum Mesolithic. Non-ruminant fauna (e.g., warthog) and low lipid-yielding reptiles (e.g., Adanson’s mud turtle and Nile monitor lizard), found in significant numbers at al-Khiday, were likely also cooked in pots. There is little evidence for the processing of wild ruminants in the Mesolithic pots, suggesting either that ruminant species were not routinely hunted or that large wild fauna may have been cooked in different ways, possibly grilled over fires. These data suggest sophisticated economic strategies by sedentary people exploiting their ecological niche to the fullest. Pottery use changed considerably in the Early Neolithic, with ruminant products being more routinely processed in pots, and while the exploitation of domesticates cannot be confirmed by a small faunal assemblage, some dairying took place. The results provide valuable information on Early and Middle Holocene lifeways in central Sudan.
Resumé
Al-Khiday, située sur la rive du Nil Blanc, présente une séquence stratigraphique exceptionnellement préservée qui fournit une opportunité unique d’utiliser l’analyze des résidus organiques pour étudier le régime alimentaire et la subsistance pendant tout le Mésolithique de Khartoum et le Néolithique ancien, une période d’environ de 3500 ans (7000–4500 cal BC). Alors que le grand assemblage diversifié d’os de poisson du Mésolithique montre une forte dépendance aux produits des habitats aquatiques, des plaines alluviales, des marais végétalisés et de la rivière, les résultats de l’analyze des résidus lipidiques suggèrent que les poissons n’étaient pas cuits dans les marmites en céramique, mais, probablement, consommés dans d’autres façons. Ou plutôt, les pots étaient utilisés pour la transformation des plantes, des herbes sauvages, des feuillues et des carex. Ce résultat, confirmé par l’analyze expérimentale, offre une preuve chimique directe de l’exploitation des plantes dans le Mésolithique de Khartoum. La faune non-ruminante, comme le phacochère, et les reptiles à faible rendement en lipides, comme la tortue d’Adanson ou le varan du Nil, retrouvés en quantités importantes à al-Khiday, était également probablement cuits dans des pots. Il y a peu de preuves de la transformation de ruminants sauvages dans les pots en céramique, ce qui suggère que les espèces des ruminants n’étaient pas systématiquement chassées, ou que la grande faune sauvage pouvait avoir été cuite de différentes manières, probablement grillée. Ces données suggèrent que les populations sédentarisées avaient des stratégies économiques sophistiquées, tirant probablement la meilleure part de leur niche écologique. L’utilization de la poterie a considérablement changé au Néolithique ancien, ou les produits des ruminants sont plus régulièrement transformés avec cuisson dans de pots, et bien que l'exploitation des animaux domestiqués ne puisse être confirmée par un petit assemblage faunique, une certaine production laitière a lieu. En résumé, les résultats fournissent des informations utiles sur les modes de vie de l'Holocène ancien et moyen dans le centre du Soudan.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Mesolithic of Sudan, or Khartoum Mesolithic, first characterized by Arkell (1949) during his excavations at the Khartoum Hospital site (Arkell, 1949) and Shaheinab (Arkell, 1953), lasted at least 3000 years (Salvatori, 2012). It is marked by a wide distribution of sites along the banks and hinterlands of the Blue and White Nile and other early Holocene rivers such as Wadi Muqaddam and Wadi Howar (Hosfield et al., 2015; Jesse, 2000; Smith, 1998). These sites include Saggai I (Caneva, 1983), Umm Marrahi (Elamin & Mohammed-Ali, 2004), Abu Darbein, Aneibis and El Damer (Haaland & Magid, 1995), Umm Singid (Caneva, et al., 1993), Jebel Sabaloka (Varadzinová et al., 2022), el-Barga (Honegger, 2005; Honegger & Bastien, 2009), and El Shaqadud (Marks & Mohammed-Ali, 1991), among others (e.g., Ali Hakem & Khabir, 1989; Caneva, 1988; Khabir, 1987; Salvatori & Usai, 2006, 2009; Salvatori, et al., 2011; Usai & Salvatori, 2002, 2005). Such sites were inhabited by communities of mainly sedentary pottery-using hunter-gatherer-fisherfolk (HGF), who produced high-quality decorated ceramics, grinding stones, and retouched stone and bone tools and instruments.
Notably, these sites are generally marked by a lack of stratified deposits, largely due to natural and anthropogenic post-depositional disturbances, thereby limiting the potential for understanding the chronology, extent, and social and economic organization of the Sudanese Mesolithic (Salvatori, 2012; Usai, 2014). However, the site of al-Khiday provides a detailed, stratigraphically integral record of Mesolithic deposits (Salvatori, 2012; Salvatori et al., 2014), producing the first cultural sequence of the Khartoum Mesolithic that covers a period of over 1000 years (Maritan, et al., 2018; Usai & Salvatori, 2019), and the transition to the Early Neolithic (Table 1). Here, the Mesolithic is divided into three phases, Early (7000–6650 cal BC), Middle (6650–6000 cal BC), and Late Mesolithic (6000–5800 cal BC). The Middle Mesolithic can be further broken down into three sub-phases: Middle Mesolithic A (6650–6500 cal BC), B (6500–6250 cal BC) and C (6200–6000 cal BC). To date, these deposits have provided a unique opportunity to investigate subsistence strategies, burial practices, material production, and social organization at a very detailed level (e.g., Buckley et al., 2014; Dal Sasso et al., 2014, 2016; Linseele, 2020; Maritan et al., 2018; Salvatori et al., 2014; Usai & Salvatori, 2019; Usai et al., 2014, 2017; Zerboni et al., 2018). The neolithization of the region appears rooted in the Mesolithic HGF groups, although some external influences cannot be ruled out (Usai, 2016). Neolithic habitation sites tend to be situated on the alluvial plain close to then-extant Nile channels, which offered reliable, fertile resources. Agro-pastoralists likely utilized large permanent camps such as Esh Shaheinab and Geili, and domesticates, mainly cattle (Bos taurus), were the dominant mammal at sites such as Kadero (4600–3800 cal BC) and Ghaba (5200–4500 cal BC), where wild plants were also intensively exploited (Caneva, 1988; Gautier, 1984; Haaland, 1992, 1995; Krzyzaniak, 1991; Marshall & Hildebrand, 2002; Out et al., 2016). Here, we use an exceptional long-term stratigraphic sequence, in combination with organic residue analysis, faunal and archaeobotanical remains, to investigate the diet and subsistence strategies of Khartoum Mesolithic and Early Neolithic White Nile populations, across a period of nearly 3500 years.
Al-Khiday Site Complex, Sudan
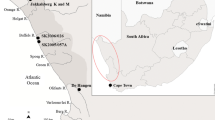
The al-Khiday complex is located along the western bank of the White Nile, 25 km south of the city of Omdurman (central Sudan) and 3 km west of the present-day Nile (Fig. 1). The group of sites at al-Khiday are situated on a sandy ridge rising around 4 m above the surrounding plain (Salvatori, 2012; Salvatori et al., 2011, 2014; Usai & Salvatori, 2005; Usai et al., 2010; Williams et al., 2015; Zerboni, 2011), similar to other sites located above the White Nile floodplain, occupied from the early Holocene (Adamson et al., 1982; Clark, 1989; Zerboni, 2011).

© CSSeS)
Map of Sudan with the location of al-Khiday sites: a kite view of site al-Khiday 2 (16-D-4); b kite view of a portion of the site under excavation (
The group of sites comprises:
-
(i)
16-D-4, a multi-phase site with a cemetery containing 235 inhumation burials of Pre-Mesolithic, Neolithic, and Classic/Late Meroitic Age; a Mesolithic functional area comprising 104 pits; and a near complete Mesolithic circular hut.
-
(ii)
16-D-4B, a second Mesolithic functional area with a large number of garbage pits, 32 of which were excavated.
-
(iii)
16-D-6, a Neolithic site, probably seasonal.
-
(iv)
16-D-3 and 16-D-5 are Mesolithic settlements. 16-D-5 also contains a poorly preserved Neolithic layer, partly disturbed by post-Meroitic tumuli.
Mesolithic al-Khiday
Settlements 16-D-3 and 16-D-5, the settlement occupation in the cemetery area (16-D-4), and the functional area (16-D-4B) cover a period of more than one thousand years, between the Early and Late Mesolithic (Salvatori, 2012; Salvatori et al., 2011). The first occupation levels at sites 16-D-3 and 16-D-5 date to the Late Early Mesolithic (Table 1) at ca. 7000 to 6750 cal BC and are characterized by postholes suggesting the presence of huts, likely inhabited by small groups of hunter-gatherer-fisherfolk exploiting the rich riverine environment on a seasonal basis (Linseele, 2020; Linseele & Zerboni, 2018; Maritan et al., 2018). Middle Mesolithic occupation levels are seen at sites 16-D-3, 16-D-4, 16-D-4B, and 16-D-5. This period has three distinct phases (Middle Mesolithic A, B and C; for dates, see Table 1) and is marked by a significant transformation in house architecture, with semi-subterranean mud-walled huts (with fireplaces) being arranged either in a beehive pattern (Salvatori et al., 2014) or isolated (Salvatori et al., 2018). Over 104 pits (Usai, 2014), with diverse fillings, were excavated at 16-D-4 (Usai & Salvatori, 2019). Those filled with ashes, burnt pebbles, (scarce) mollusk shells, and mammal and fish bones (often partially in anatomical connection) are thought to be fireplace pits. Most of the remaining features, including a concentration of 32 pits at 16-D-4B, appear to be large refuse dumps containing significant amounts of faunal remains. All are connected with the main settlement (16-D-5) in a spatial arrangement that suggests an organized use of the space and implies a more sedentary lifestyle (Usai & Salvatori, 2019).
During this period, the climate in the lower White Nile valley was significantly wetter due to high stands of the White Nile floods between ca. 9700 and 9100 cal BP (Adamson et al., 1980; Williams, 2009). The deposits at Mesolithic al-Khiday suggest a local swamp or wetland environment, seasonally flooded by the White Nile (Williams, 2009; Williams & Adamson, 1980; Williams et al., 2015; Zerboni, 2011). Locally wet areas were interspersed with dry thorn savanna and scrub, including species of Acacia, sometimes in association with Balanites aegyptiaca, and grasses such as Hyparrhenia anthistirriodes, Cymbopogon nervatus, and Sorghum spp. (Wickens, 1982). Extensive papyrus swamps probably extended along the White Nile and likely included several varieties of sedges, as suggested by phytoliths from Early and Middle Mesolithic C deposits 16-D-3 (Usai & Salvatori, 2019). The wetlands would have been a source of abundant fish and edible Pila snails, with drier areas providing favorable conditions for species such as kob (Kobus kob), elephant (Loxodonta), and buffalo (Syncerus caffer), as detected in the faunal remains. Thus, during the Mesolithic, the al-Khiday sites were situated in a rich and productive location, providing a sustainable environment over a chronological span of ca. 1200 years.
Neolithic al-Khiday
The Neolithic evidence at al-Khiday originates from 16-D-5 and 16-D-6 sites. The occupation layer at 16-D-5 comprises a ‘carpet’ of fragmented pottery, rhyolite and quartzite lithic tools, Chambardia shells, grinding stones, and numerous animal bones (Salvatori, 2012; Salvatori et al., 2011). At site 16-D-6, a single occupation layer was recognized, comprising post-holes and a small fireplace (Salvatori et al., 2018). A complete burial of an individual in a contracted position was found, suggesting the presence of a small cemetery associated with the settlement (Salvatori et al., 2018, Fig. 14).
Although detailed information on the climate during the Neolithic occupation at al-Khiday is missing, general data support drying conditions, denoted by a fall in the level of the Nile and a retreat of vegetation (Wickens, 1982; Williams et al., 2015). This is confirmed by oxygen isotope analysis of al-Khiday Neolithic human remains (Iacumin et al., 2016), whose values indicate that the population experienced much drier conditions, although not dry enough to limit the hunting of diverse wild animals.
Al-Khiday Pottery
The Mesolithic pottery assemblage comprised vertical and open bowls and (mainly) globular jars. Incised and impressed decorations cover most of the pots’ surfaces, including the rims. They often occur on the inner surface (Fig. 2a–h). A range of decoration types (see Fig. 2) was used, including incised wavy line, dotted wavy line, alternately pivoting stamp, rocker stamp drops, and rocker stamp dotted zigzag (Dal Sasso et al., 2014; Salvatori, 2012). The Mesolithic pottery comprises two main fabrics (Dal Sasso et al., 2014): one characterized by abundant grains of sand-sized quartz with a sub-rounded shape (Fig. 3a), and the other rich in feldspar (Fig. 3b). The latter is derived from the grinding of an alkaline granite or syenite and is only found in pottery with certain types of decoration, in particular, incised wavy line (Fig. 2a), rocker stamp dotted zigzag packed (Fig. 2d), and alternately pivoting stamp (Fig. 2g).

Selection of lipid-yielding Mesolithic and Neolithic pottery fragments. Decoration is described by means of technique plus design. Early Mesolithic: a Incised Wavy line; b Rocker stamp drops deep. Middle Mesolithic A: c Rocker stamp drops fan, deep. Middle Mesolithic C: d Rocker stamp drops, horizontal; e Rocker stamp drops oblique and fan, deep; f Rocker stamp drops band; g Alternately Pivoting Stamp atypical. Late Mesolithic A: h Laqiya. Early Neolithic: i, j Rocker stamp unevenly spaced dots; k Rocker stamp dotted zig-zag spaced; l Rocker stamp dotted zig-zag; m Alternately Pivoting Stamp; n Rocker stamp dotted concentric semi-circles; o Burnished undecorated

Photomicrographs of the three main fabrics observed among the Mesolithic and Neolithic pottery at al-Khiday: a fabric rich in sand-sized rounded quartz grains; b fabric rich in alkali-feldspar; c fabric rich in sub-angular fine sand-sized quartz inclusions. Images taken at the same magnification and in crossed-polarized light
Neolithic pottery (Fig. 2i–o) is characterized by new decoration types (fish scale, incised straight lines). However, some Mesolithic styles remain in use (alternately pivoting stamp, rocker stamp dotted zigzag and scraped). The Neolithic pottery from 16-D-5 includes globular, hemispherical, and conical bowls, and globular and ovoid jars decorated with a large range of motifs/techniques. Evenly spaced rocker stamp and packed dotted zig-zag rocker stamp are the most common (Fig. 2i–l). Concentric dotted semicircles produced by an alternating pivoting stamp technique and, as seen on complete pots found at the site, usually organized in symmetric panels, are also very common (Fig. 2g, m). Pottery without impressions is also common, generally with red-slipped polished surfaces and a series of black semicircles at the rim (Fig. 2o). The Neolithic pottery is produced using a quartz-rich paste, with smaller inclusions (fine sand), which are sub-rounded/sub-angular in shape (Fig. 3c) denoting a technological change from Mesolithic production (Dal Sasso et al., 2014; Salvatori, 2012; Fig. 3c).
Lipid Residue Analysis and Results
A total of 152 potsherds, from several al-Khiday sites, including 16-D-3, 16-D-4, 16-D-4B, 16-D-5, and 16-D-6, were investigated according to well-established analytical procedures described in Online Supplementary Material 1 (Correa-Ascencio & Evershed, 2014), with 37 sherds yielding interpretable lipid profiles (Mesolithic 22, Neolithic 15), i.e., containing sufficient concentrations (> 5 µg g−1) of lipids that can be reliably interpreted (Evershed, 2008). Overall, the lipid recovery rate was 24%, although it varied significantly between the Mesolithic phases and the Early Neolithic (Table 2). The mean lipid concentration of the sherds was 55.0 µg g−1, with a maximum lipid concentration of 459.6 µg g−1 (Table 3). Notably, lipid concentration for 81% of the interpretable residues (n = 23) was less than 50 µg g−1, with 59% containing less than 30 µg g−1. These low lipid concentrations strongly suggest that the commodities processed in most vessels are likely of a low lipid-bearing origin, either plants or low lipid-yielding animal products.
The analysis of the fatty acid methyl esters (FAMEs) extracted from the al-Khiday potsherds shows lipid profiles dominated by free fatty acids, palmitic (C16) and stearic (C18) (Fig. 4a). Also present in the majority of the Mesolithic lipid residues (73%, n = 22) are a series of even-numbered long-chain fatty acids (LCFAs) in the range up to C30, often in high abundance (Table 3 and Fig. 4b–d). These are strongly indicative either of an origin in leaf or stem epicuticular waxes (Bianchi, 1995; Kolattukudy et al., 1976; Kunst & Samuels, 2003; Tulloch, 1976) or suberin (Kolattukudy, 1980, 1981; Pollard et al., 2008; Walton, 1990), particularly given the low abundance of lipids in the majority of the vessels. Notably, of these plant lipid profiles, 21 (57% of the assemblage, 95% of Mesolithic sherds) were found in Mesolithic potsherds, with only six originating from the Neolithic vessels (16% of the assemblage, 40% of Neolithic vessels).

a-d Partial gas chromatograms of acid-extracted FAMEs showing a-c: typical plant lipid profiles (ALK479, ALK532 and ALK552, Mesolithic sherds); d: typical degraded animal fat lipid profile (ALK1927, Early Neolithic sherd). Circles, n-alkanoic acids (fatty acids, FA), triangles, diacids; IS, internal standard, C34 n-tetratriacontane. Number denotes carbon chain length
Several lipid profiles (n = 6, Fig. 4b–d) also include a series of long-chain α, ω-dicarboxylic acids (diacids), often present in high abundance. These major components of plant polymeric compounds, such as cutin and suberin, are highly unusual in plants. Those with chain lengths of C16 to C26 are considered diagnostic for the presence of suberin, found in the outer part of underground plants (Dembitsky et al., 2002; Holloway, 1984; Kolattukudy, 1980, 1981; Pollard et al., 2008; Walton, 1990). The α, ω-dicarboxylic acids are present at < 5% (apart from in the Arabidopsis) in cutin, so this is highly unlikely to be the source of the diacids within the lipid profiles.
It should be noted that while the detection of abundant fatty acids and diacids is consistent with an origin in plant polyesters, such as suberin or cutin, the absence of hydroxy fatty acids is surprising. Since sterols can be lost in organic residues by a range of degradative pathways (Hammann et al., 2018), it is possible that hydroxylated aliphatic components, such as hydroxy fatty acids, are vulnerable to the same fate and, thus, are not seen in lipid extracts. Nonetheless, the presence of diacids indicate that underground storage organs, i.e., tubers, were likely processed in some al-Khiday vessels.
All lipid-yielding samples were analyzed by gas chromatography-mass spectrometry (GC–MS) in selected ion monitoring (SIM) mode to check for the presence of freshwater biomarkers, such as ω-(o-alkylphenyl) alkanoic acids (APAAs) and vicinal dihydroxy acid (DHYAs), which denote the processing of shellfish/crustaceans, fish, waterfowl, and aquatic mammals (see Cramp & Evershed, 2014). No aquatic biomarkers were detectable in the analyzed potsherds, although some aquatic input to the vessels cannot be discounted.
Thirty-seven samples (Mesolithic n = 22 and Early Neolithic n = 15) underwent gas chromatography–combustion–isotope ratio mass spectrometry (GC-C-IRMS) analyses (Table 3 and Fig. 5) to determine the δ13C values of the major fatty acids, C16:0 and C18:0, and ascertain the source of the lipids extracted (Dunne et al., 2012).

Graphs showing: Δ13C (δ13C18:0–δ13C16:0) values from a Mesolithic and b Neolithic vessels from al-Khiday; c, d show the types of foods processed in the al-Khiday vessels. Ranges represent the mean ± 1 s.d. of the Δ 13C values for a global database comprising modern reference animal fats from the UK, Africa, and elsewhere (Dudd & Evershed, 1998; Dunne et al., 2012)
Mesolithic Vessels
Lipid residue results for the Mesolithic period (Fig. 5a, see also annotated Fig. 1a in Online Supplementary Material 2) show that twelve sherds (55%) plot within the non-ruminant/plant region, three sherds plot within the ruminant adipose region (14%), and six sherds plot between the ruminant and non-ruminant/plant region (27%). Significantly, one sherd (ALK542, Late Mesolithic A), with a Δ13C value of − 3.3 ‰ (Fig. 5a), plots just within the ruminant dairy region It should be noted that the vessel plots at the upper end of the range, suggest either some mixing of dairy and ruminant carcass products or possibly, that low quantities of dairy products were masked isotopically through mixing with high amounts of non-ruminant fats, although this seems unlikely given its low lipid concentration (18.4 µg g−1).
Of the twelve Mesolithic sherds plotting within the non-ruminant/plant region, ten display lipid profiles containing a series of even-numbered long-chain fatty acids (LCFAs) up to C30 (see Tables 3 and 4, and Fig. 5a), often in high abundance. Of these, seven have lipid concentrations of less than 30 µg g−1, with two at less than 50 µg g−1 and one at 81.1 µg g−1. Three of these sherds also contain diacids. The combination of LCFAs, diacids, and low lipid concentrations suggests these vessels were either used to process solely plants or plants with small amounts of carcass products. However, care must be taken in their interpretation as fatty acids derived from plant processing can contribute more depleted δ13C values to the overall fatty acid signature of the C16:0 and C18:0 fatty acids. Similarly, of the three Mesolithic potsherds which plot in the ruminant adipose region, two include a series of LCFAs (up to C30) and diacids together with lipid concentrations of less than 50 µg g−1. Thus, they were also likely to have been used to process combinations of plant and animal products or, possibly, solely plants (see below on experimental processing). The other vessel with a ruminant signal (ALK521) has a much higher lipid concentration at 459.6 µg g−1 and was most likely used to process ruminant carcass products (Online Supplementary Material, Fig. 1a and Table 3). Six vessels yielded Δ13C values plotting between the ruminant and non-ruminant/plant regions. Of these, vessel ALK634 did not include LCFAs, but had a low lipid concentration at 7.6 µg g−1 and thus was more likely to have been used solely to process plants, as were the remaining four vessels, with lipid concentrations of less than 50 µg g−1, and series of LCFAs (and diacids in vessel ALK552). The remaining vessel, ALK526, which did include LCFAs, but with a lipid concentration of 200.8 µg g−1, may have been used to process both plants and animal carcass products, or denote multi-use of the vessel.
Early Neolithic Vessels
In the Early Neolithic, seven vessels (47%) plot within the ruminant adipose region (Fig. 5b, see also annotated Fig. 1b in Online Supplementary Material), confirming they were used to process carcass products from either domesticated cattle, sheep (Ovis aries)/goat (Capra hircus), or hunted wild ruminant mammals such as antelope. Of these, the lipid profile from one vessel (ALK630) also includes a series of LCFAs (up to C28), suggesting that, although this one vessel was likely used to process animal products and plants, the majority (n = 6) were used solely to process ruminant carcass products. Three vessels, ALK517, ALK518, and ALK627B, plot in the non-ruminant/plant region and were likely used to process non-ruminant/plant products (with two also including sequences of LCFAs). Three further vessels, plotting between the ruminant and non-ruminant region, were used to process mixtures of ruminant and non-ruminant carcass products. Two sherds (ALK516 and ALK530) plot within the ruminant dairy region with Δ13C values of − 3.6 and − 5.5 ‰ (Fig. 5b), suggesting some minor exploitation of ruminant animals for their dairy products.
δ13C Values
Organic residue analysis can be a powerful proxy for investigating animal management strategies, such as transhumance (Dunne et al., 2012), and in discerning the past isoscapes in which prehistoric groups lived (West et al., 2010). The foods that animals eat exhibit characteristic isotopic signatures (Gannes et al., 1997) and isotopic analyses (δ13C) of fatty acids extracted from archaeological potsherds are therefore a reflection of the consumed diet, providing information about the environment in which the animals foraged (Copley et al., 2003; Mukherjee et al., 2005). In the case of the Mesolithic vessels, the majority appear to have been used to process plants, and thus the δ13C values will reflect their C3/C4 origin.
The δ13C16:0 values of the fatty acids extracted from the al-Khiday potsherds range from − 26.4 to − 16.0 ‰, and the δ13C18:0 values range from − 25.0 to − 17.9 ‰ (Table 3). The δ13C16:0 values (Table 3) suggest either that the plants processed in the vessels were mainly of C4 origin or that the animals giving rise to these fats had subsisted on a range of different forages composed of mainly C4 plants or C4 plants with some contribution from C3 plants. Where possible, the δ13C values of the long-chain fatty acids (denoting plant processing) were measured to provide information on their possible origin (i.e., C3 or C4, Table 4). The δ13C values fall mostly within the known δ13C values for C3 bulk plant lipids, although often at the low end of the range, which suggests that the plants processed in the vessels were generally of C3 origin, with some possible minor C4 input. It should be noted that these plants may have routinely been both prepared and mixed (and sequentially processed) within the vessels.
While the more enriched (C4) δ13C16:0 and δ13C18:0 values suggest the processing of either C4 wild grasses (such as sorghum and millet) or C4 tubers (such as the tuberous sedge plant, Cyperus rotundus), both common in al-Khiday phytolith remains, the LCFA δ13C values suggest a more C3 origin. The presence of LCFAs denotes the processing of leafy plants, candidates for which include jute mallow, also known as bush okra (Corchorus olitorius), widely used as a leafy vegetable and potherb in Africa and found close to the Nile today in abundance. The presence of the diacids in some vessels confirms the likely processing of sedges (although these were not present in enough quantity to be measured isotopically).
To help shed light on possible commodities cooked in the pots, we carried out in situ experimental cooking of the C4 grasses—sorghum (Sorghum sp.) and millet (Panicum miliaceum)—and tuberous sedge plants (Cyperus rotundus), in individual local, hand-made cooking pots, to make comparisons between the δ13C16:0 and δ13C18:0 values obtained from the separate cooking of each plant and those obtained from the archaeological vessels.
Organic Residue Analysis of the Experimental Cooking Vessels
Full details of the production of pots and the cooking experiments can be found in Online Supplementary Material (OSM 3 and Fig. 6). Rims and bases from these experimental vessels, together with ‘burnt-on’ residues from the millet and Cyperus cooking events, were analyzed using well-established analytical procedures described in OSM 1 (Correa-Ascencio & Evershed, 2014). Lipid recoveries from the vessels were generally low (Table 5) but higher from the surface residue, the ‘remains’ of the cooking events, for example, lipid concentration of the surface residue from the millet pot was high at 5437.5 µg g−1.

Ceramic pots being hand-made in Omdurman. Stages of production: a the clay is shaped, using a large stone, to make a bowl shape; b the lip is thinned using the paddle and anvil technique and raised to close up into a jar shape; c the rim and handles are made by adding shaped coils of clay and a red slip is applied using a cloth soaked in ochre and diluted in water, pots were then polished with a pebble; d the pots were then fired in the open-air using wood and dung as fuel. Fragments of broken jars were used to create a cover, thus increasing thermal insulation
Where possible, the δ13C values of the major fatty acids, C16:0 and C18:0, were measured using GC-C-IRMS (Table 5 and Fig. 7). The δ13C values of both the sherd and the surface residue from the vessel used to process millet suggest a C4 origin as does the vessel used to cook sorghum. Both the sherd and surface residue from the millet cooking plot in the plant/non-ruminant region with Δ13C values of 0.4 and 0.3‰ (Fig. 7), as does the surface residue from the cyperus-processing vessel, at 0.2‰. Interestingly, the sherd from the sorghum vessel plotted between the ruminant and plant/non-ruminant ranges, although closer to the ruminant, demonstrating that vessels used to process plants can plot at the furthest extent of the plant range. As seen in Fig. 7, the Δ13C values from the experimental pots are similar to those from the archaeological pots used to process plants and confirm that the plant lipid profiles could originate from C4 wild grass (millet and sorghum) and/or tuber (Cyperus?) processing. These values, together with the δ13C values from the LCFAs, which denote the addition/mixing of C3 leafy plants, suggest that the vessels may have been used to cook meals comprising grains, tubers, and leafy vegetables, sometimes with the addition of meat products.

Graph showing Δ13C (δ13C18:0–δ13C16:0) values from the experimental vessels and surface or burnt-on residues from the same vessels. Ranges represent the mean ± 1 s.d. of the Δ13C values for a global database comprising modern reference animal fats from the UK, Africa, and elsewhere (Dudd & Evershed, 1998; Dunne et al., 2012)
Discussion of the Lipid Residue Results
Pottery use across the Mesolithic (see Table 2), covering the 1200 years of the Early Mesolithic (c. 7000 cal BC) to the Late Mesolithic A (c. 5800 cal BC), seems to be entirely consistent. However, by the Early Neolithic (4500 cal BC), changes in diet and subsistence can be seen in the clear differences between foods processed in the Mesolithic and Early Neolithic pottery (Fig. 5). In the Mesolithic period (Fig. 5a, c), most vessels (n = 18; 82%) plot either within the non-ruminant/plant range or between the ruminant and non-ruminant/plant region. Fifteen of these (68% of total Mesolithic sherds) were used to process wild plants or, possibly, combinations of plant and low lipid-yielding animal products, evidenced by the combination of even-numbered LCFAs, diacids and very low lipid concentrations (all at ≤ 50.0 µg g−1, save for ALK545 at 81.1 µg g−1). The animal products processed mainly comprised non-ruminants, likely low-lipid yielding wild fauna. In contrast, a greater number of Neolithic vessels were used to process ruminant products (Fig. 5b, d), and although it should be noted that the experimental vessel used to process sorghum also plotted at the upper end of this region, only one, of the seven used to process ruminant products, displayed evidence for plant processing. The ruminant carcass products (discussed further below) display a slightly broader range of δ13C16:0 values, with some vessels being used to process carcass products from animals consuming more C4 products, in good agreement with the drying environmental conditions. While six vessels plot either in the non-ruminant/plant or between the ruminant and non-ruminant/plant region, only three contain evidence for plant processing, suggesting some vessels may have been used to process mixtures of these animal products and confirming a much greater reliance on meat processing than in the Mesolithic.
Faunal Remains and Their Lipid Signatures at al-Khiday
The Mesolithic deposits at al-Khiday comprise predominantly fish remains (often at 90% or more), considerably higher than at other Central Sudanese Mesolithic sites (Linseele & Zerboni, 2018). Certainly, al-Khiday is located in an area with easy access to the Nile and other productive aquatic habitats, including the Jebel Baroka lake (Cremaschi et al., 2007) and other (possibly seasonal) small lake formations. At least twenty fish species have been identified (OSM 4, Table 1), indicating the exploitation of diverse aquatic habitats, floodplains, vegetated marshes, and open water, although clariid catfish, a shallow water fish, clearly dominate the assemblage. More details on these fish and a diachronic analysis have been published elsewhere (Linseele, 2020; also see OSM 4, Table 1). Freshwater turtles and Nile monitor lizard (Varanus niloticus) are also found, with the Adanson’s mud turtle (Pelusios adansonii) in high abundance. Large numbers of mollusk shells, mainly Pila wernei (OSM 4, Table 1), were also present, particularly at 16-D-3, a shell midden. This freshwater snail is known to be easy to collect from swampy areas and pools during the dry season (Gautier, 1983). Bird remains are not common at the site (possibly due to poor preservation of small and fragile bones), although stork, duck, and geese were identified. The Mesolithic mammal fauna is diverse, indicating a lush savanna environment. Various wild mammal taxa are present (Table 6), particularly antelopes, although these could not often be identified beyond size class. Also found were the remains of baboon, hare, warthog, and (scarce) very large mammal taxa, including elephant, rhinoceros, and hippopotamus.
The faunal assemblage excavated from the Neolithic site 16-D-6 is very small and much more poorly preserved than those from the Mesolithic sites, likely due to the shallow nature of the deposits. The remains of wild ruminants, including three giraffe, two small antelope, and three medium-sized antelope, are more common than those of domesticates. Scant evidence for domesticates comprises three bones attributed to sheep/goat, and one of sheep. Although domesticated cattle bones were not identified, they may be present in those identified as large bovid, cattle, or buffalo. Compared to the Mesolithic, fish remains are much less common, consisting mainly of Nile perch (Lates niloticus) and lacking the diverse range of species seen in the Mesolithic.
Animal Product Processing
During the Mesolithic, eight of the 22 lipid-yielding vessels (36%) were used to process animal carcass products. Of these, only three (14%) appear to have been used solely for animal carcass processing, with the remaining five (23%) comprising lipid profiles suggesting the processing of meat and plants (Fig. 5a, c). Most animal products processed across the 1200 years of the Mesolithic period seem to be either of non-ruminant origin or comprise mixtures of ruminant and non-ruminant carcass products (Table 3). This is surprising considering that wild ruminants contribute to the majority of identified mammalian remains, ranging between 84% and 100% of the identified mammalian taxa in the different Mesolithic assemblages (Table 6). Non-ruminant products may have included reptiles such as Adanson’s mud turtle and Nile monitor lizard, found in significant numbers at al-Khiday, or warthog (Phacochoerus aethiopicus). However, these are not common at the site (Linseele, 2020). Certainly, the processing of low lipid-yielding meats from small fauna such as turtles may explain the low concentrations of lipids found. However, a cautionary note is needed as the Δ13C values from experimental processing of plants show them to plot either at the extent of the non-ruminant/plant range or between the ruminant and non-ruminant/plant regions. This suggests that vessels attributed to a carcass product origin may also have been used to process plants.
In contrast, the processing of ruminant products predominates in Neolithic vessels (47%, Fig. 5b, d). These could originate from domesticates, such as cattle and sheep/goat, although the Neolithic faunal assemblage is very small and poorly preserved, making comparisons difficult. Interestingly, wild fauna was present in much higher abundance than domesticates. Africa presents a unique situation where, unlike other continents, there are many wild ruminants, including about 50–60 different species of bovids. Thus, the presence of ruminant adipose products is not a clear indication of the presence of domesticates and, as wild ruminant fauna are present in greater abundance in the animal bone assemblage, the level of exploitation of domesticates at al-Khiday remains unclear.
Dairy Product Exploitation
Evidence for dairying is, however, a much clearer indication of domesticate exploitation. There is minor evidence for dairying in the Mesolithic, with one sherd plotting at the extent of the dairy range (Table 3 and Fig. 5a). This sherd is of the Laqiya type, from the Late Mesolithic A phase (ALK542, 6000–5800 cal BC), which would suggest a very early presence of domesticates and, indeed, dairying practices, at the site. However, it should be noted that fatty acids deriving from plant product processing can contribute more depleted δ13C values to the overall fatty acid signature of the C16:0 and C18:0 fatty acids, possibly giving a false positive for the presence of dairy products. Nonetheless, the presence of Laqiya pottery at al-Khiday is intriguing as it is thought to be possibly related to a new group that may originate from the Wadi Howar region in northwestern Sudan (Jesse, 2000, 2003; Usai & Salvatori, 2019). This period at al-Khiday corresponds to the phase when domestic animals are first recorded in the Nubian region, north of al-Khiday. Domestic cattle were identified in a burial at El Barga, dated to ca. 5700 cal BC (Honegger, 2004; Honegger & Williams, 2015), and domestic sheep/goat bones have been directly dated to 5700 cal BC (KIA34817, 6820 ± 30 BP) at the site of Boni S05/140 in the Fourth Cataract region (Petrick, 2012; Pöllath, 2012; Wotzka et al., 2012). If we consider the possibility of an allochthonous origin, then it may be that dairy products become available to Late Mesolithic people at al-Khiday through a system of exchanges with groups from more northern sites known to be exploiting domesticates. Trading between some Mesolithic groups is supported by the identification of clay/temper with a northern origin, found in some al-Khiday pottery, suggesting either the circulation of raw materials used in pottery production or the movement of the pottery itself (Dal Sasso et al., 2014). Alternatively, it may be that small numbers of domesticated animals were present in central Sudan during this phase, possibly as a minor component. However, they are not (currently) present in the al-Khiday faunal assemblage (Salvatori & Usai, 2019), and this remains an open question.
Nevertheless, dairying, albeit on a minor scale, can be confirmed in the Neolithic (Fig. 5b), from at least 4500 cal BC, with two sherds (13%) yielding a ruminant dairy signal, very similar to results from the Neolithic phase (4600–4000 BC) at Khor Shambat 1 where one vessel returned evidence for dairy processing (Dunne et al., 2021). However, these data are in contrast to the Early Neolithic site of Kadero (4600–3800 BC; Krzyzaniak, 1991), where 47% of sherds yielded a ruminant dairy signal, suggesting that dairying was an important economic activity at that site (Dunne et al., 2017). This may be due to the location of Kadero, which, although further from the Nile, had access to good grazing land found on the alluvial plain on the east bank.
Were Freshwater Fish Cooked in Pots at al-Khiday?
The absence of aquatic biomarkers in the al-Khiday ceramic assemblage was somewhat surprising, bearing in mind the extensive availability of nearby surface waters and the preponderance of fish fauna in the Mesolithic deposits (Linseele, 2020). Certainly, central Sudan is renowned for its Mesolithic hunter-gatherer-fisherfolk, giving rise to the aptly named Aqualithic Culture (Sutton, 1977), where sites are known for the presence of bone harpoons and substantial fish and other aquatic faunal remains (Linseele, 2020; Linseele & Zerboni, 2018). At al-Khiday, harpoons, gorge hooks, and large catfish spines used as tools, possibly perforators, were found (Usai & Salvatori, 2019). Fish remains averaged 90.5% of the 74,000 Mesolithic bones studied, comprising as much as 99% at one site (16-D4-B). In fact, of all central Sudanese Mesolithic faunal assemblages, those at al-Khiday have the greatest number of fish remains relative to the total number of animal remains collected (Linseele & Zerboni, 2018).
Our data suggest that fish were not boiled in pots at al-Khiday, a trend that increasingly appears to be trans-Saharan (Dunne et al., 2020, 2021). Fish could have been wrapped in leaves and cooked on hot stones or coals in the fire or by other methods. The presence of articulated fish skeletons at al-Khiday suggests sun-drying practices, thought to have been practiced by Iron Age people inhabiting the Sahel (Van Neer, 1995) and traditional fishermen at Lake Chad (Blache et al., 1962). Fish could also have been smoked or salted. Indeed, the mineralogical analysis revealed the presence of salt (halite, NaCl) on fish bones and the inner surfaces of several potsherds at site 16-D-4B (Maritan et al., 2018). Large storage containers, with a rim diameter of between 44 and 50 cm, found at 16-D-4 and 16-D-4B (Salvatori, 2012) and similar to those found at Kobadi (Jousse et al., 2008) and in the Atbara region of the Middle Nile (Haaland, 1995), could also have been used to store cured or salted fish.
Plant Processing: al-Khiday in Regional Context
The survival of archaeobotanical remains at early northern African archaeological sites is patchy due to a lack of systematic sampling and poor preservation of botanical remains, often caused by natural and anthropic post-depositional disturbances (Salvatori, 2012). Lipid analysis of vessels can thus provide direct evidence for the processing of plants within vessels, as demonstrated above. Notably, at al-Khiday, most vessels (82%) were used to process plants (Fig. 5a , c), whether solely or with animal products. Based on the presence of LCFAs and diacids, these plants likely comprised wild grasses, leafy plants, and, possibly, underground storage organs such as sedges. While evidence for plants is rare at al-Khiday, wild grasses of C4 origin were found at nearby sites, such as Kadero, Umm Direiwa, Zakiab, Shaheinab, Saggai, and Shaqadud cave, in Central Sudan (Ryan et al., 2016). These remains mostly comprise wild panicoid grasses such as Panicum sp., Setaria sp., Sorghum sp., Echinochloa sp., and Pennisetum sp., often identified as impressions of plants on pottery rather than found as macro-remains. A recent analysis of phytoliths and dental calculus collected from two Early Neolithic graves at Ghaba, Central Sudan—Grave 233 (6620 ± 40 BP, 5620–5480 cal BC (Beta-59170)) and Grave 295 (5800 ± 40 BP, 4729–4544 cal BC (Beta-371517)) revealed a dominance of C4 grasses, including grasses of the large Paniceae tribe, suggesting the exploitation of mixed stands of probably non-domesticated savanna grasses (Out et al., 2016). Macrobotanical remains are absent at al-Khiday, but microbotanical screening of the deposits at 16-D-3 yielded phytoliths, including wild grasses (both Panicoid and Chloridoid) and sedges, with no changes evident through the sequence. Starch from the sedge plant, Cyperus rotundus, and grasses of the family Triticeae was also recovered from the calculus of Pre-Mesolithic, as well as Neolithic individuals (Buckley et al., 2014). The extensive use of grinding stones at al-Khiday also highlights the importance of plant processing (Usai & Salvatori, 2019).
The Possible Exploitation of Cyperaceae (Sedges)
The presence of LCFAS, together with diacids, in six of the al-Khiday Mesolithic vessels (27%) is suggestive of the presence of suberin, the outer covering of underground plants—corms, roots, and tubers from underground storage organs. The likely sources of tubers and rhizomes at Mesolithic al-Khiday are the Cyperaceae (sedges), including nut-grass (Cyperus rotundus), club-rush (Scirpus maritimus or S. tuberosus), catstail or reedmace (Typha spp.), bulrushes (Schoenoplectus spp.), papyrus (Cyperus papyrus), and the common reed (Phragmites australis), together with other wild tubers grown in Sudan such as Xanthosoma sagittifolium and Colocasta antiquorum (Ferguson, 1954). One of the main candidates is Cyperus rotundus, a C4 plant known as the purple nutsedge or nutgrass, abundant throughout much of the Nile Valley today, growing in dense swards at the water’s edge (Wetterstrom, 1993).
Today, their rhizomes are widely used as food by humans (Ertuğ, 2000; Gragson, 1997; Marlowe & Berbesque, 2009; Mattalia et al., 2013; Ochoa & Ladio, 2015; Vickers & Plowman, 1984; Vincent, 1985) and as animal feed/forage (Abbiw, 1990; Kern, 1974; Kulhari & Joshi, 1992; Parsons & Cuthbertson, 1992). There is also a long history of use of Cyperus rotundus for non-culinary purposes; for example, in perfume manufacturing by the Mycenaeans (Negbi, 1992) and for aromatic purposes in Predynastic Egypt (Fahmy, 2005). Various ancient writers, including Herodotus and the Hippocratic physicians (fifth century BC) and Theophrastus, Pliny, and Dioscorides (first century AD), also discuss its use as both perfume and in medicine (Manniche, 1989; Negbi, 1992).
Evidence for Cyperus Exploitation in Holocene North Africa
In the Pleistocene (ca. 17,000 to 15,000 BC), a diverse assemblage of wild plants, dominated by carbonized rhizomes of several sedges (Cyperus rotundus, Scirpus maritimus, and Scirpus tuberosus), was found at the site of Wadi Kubbaniya, Egypt (Hillman, 1989) and at the early Neolithic Nabta Playa site, E-75-6, in the Western Desert of Egypt (ca. 6900 BC). Over 120 wild plant taxa were identified, including wild grasses, small-seeded legumes, and fruit and tuber plants (Wasylikowa et al., 2001; Wendorf et al., 1992). There were also parenchymatous tissues derived from root and tuber organs, with the majority comprising tubers of Cyperus rotundus (Hather, 1995). Tubers and nutlets of Scirpus cf. tuberosus/maritimus in charred human feces at Wadi Kubbaniya provide conclusive evidence of their dietary use (Hillman et al., 1989). In the Libyan Sahara, Cyperaceae (undiff and Carex sp.) were found at Uan Tabu, and Cyperus rotundus was identified at Uan Afuda (Castelletti et al., 1999), both dating to the Late Acacus period ca. 7000–5500 cal BC.
Notably, a combined analytical and morphological analysis of material extracted from dental calculus of seven burials et al-Khiday demonstrates the ingestion of Cyperus rotundus. The compound calarene (β-gurjunene), known to be present in the essential oil component of C. rotundus (Mekem Sonwa & König, 2001), together with a suite of monoterpenoid and sesquiterpenoids previously identified in the rhizomes and tubers of C. rotundus (Meena et al., 2010; Mekem Sonwa, 2000; Mekem Sonwa & König, 2001), is indicative of the plant species. It is identified in dental calculus samples from all periods, from the pre-Mesolithic, Neolithic, and Meroitic, covering more than 9000 years (Buckley et al., 2014).
Conclusion
Detailed and comprehensive excavations of well-defined stratigraphic sequences at al-Khiday yielded a unique opportunity to use organic residue analysis to investigate dietary and subsistence behavior over the long duration of the Khartoum Mesolithic and into the Neolithic transition in central Sudan. Notably, while zooarchaeological data from the Mesolithic deposits suggests both an intensive and extensive strategy of aquatic exploitation, there is no evidence that these aquatic products were cooked in pots. Instead, the vessels provide direct chemical evidence of plant gathering and processing, which has long been postulated to be a major part of Holocene subsistence in Sudan. The lipid residues, backed up by experimental analysis, confirm the direct processing of abundant wild grasses found in nearby savanna drylands as well as sedges and leafy vegetables, likely located in humid areas along the Nile. Non-ruminant fauna, possibly wild pig and low lipid-yielding reptiles such as Adanson’s mud turtle and Nile monitor lizard, may also have been processed alongside the plant resources. Surprisingly, there is little evidence for the processing of ruminants in the pots, despite wild ruminants contributing the majority of identified mammalian taxa (although it is important to note that the total NISP by phase for identified taxa is still very low). This suggests either that hunting was a minor aspect or that large wild fauna may have been cooked in different ways, similarly to the aquatic resources, possibly roasted over fires. Intriguingly, links to other, more northern groups exploiting domesticates may have resulted in the acquisition of a vessel containing dairy products.
Pottery use changed considerably in the Neolithic, as vessels were used to process mainly carcass products from ruminant animals. The question of whether these were domesticated ruminants, however, remains open. Although the Neolithic faunal assemblage is small and rather poorly preserved, it is dominated by wild ruminants, such as antelopes, with scarce evidence for domesticates, suggesting that hunting was an important activity. Nonetheless, minor evidence for dairying confirms either some management of cattle, sheep, or goat for milk or trade with surrounding pastoralists. Certainly, comparison to lipid residue analysis from other Neolithic pottery assemblages (Kadero and Khor Shambat 1: Dunne et al., 2017, 2021) suggests varied and sophisticated economic strategies by middle Holocene groups in this area, possibly related to local ecosystem variation.
Data Availability
All data are available in the paper.
References
Abbiw, D. K. (1990). Useful plants of Ghana. Royal Botanic Gardens, Kew.
Adamson, D. A., Gasse, F., Street, F., & Williams, M. (1980). Late Quaternary history of the Nile. Nature, 288(5786), 50–55.
Adamson, D. A., Williams, M. A. J., & Gillespie, R. (1982). Palaeogeography of the Gezira and the lower Blue and White Nile valleys. In M. A. J. Williams & D. A. Adamson (Eds.), A land between two Niles (pp. 165–219). Balkema.
Ali Hakem, A. M., & Khabir, A. R. M. (1989). Sarourab 2: A new contribution to the Early Khartoum tradition from Bauda site. In L. Krzyzaniak & M. Kobusiewicz (Eds.), Late prehistory of the Nile Basin and the Sahara (pp. 381–385). Poznań Archaeological Museum.
Arkell, A. J. (1949). Early Khartoum. Oxford University Press.
Arkell, A. J. (1953). Shaheinab. An account of the excavation of a Neolithic occupation site carried out for the Sudan Antiquities Service in 1949–50. Oxford University Press.
Bianchi, G. (1995). Plant waxes. In R. J. Hamilton (Ed.), Waxes: Chemistry, molecular biology and functions (pp. 176–222). The Oily Press Ltd.
Blache, J., Miton, F., & Stauch, A. (1962). Première contribution à la connaissance de la pêche dans le bassin hydrographique Logone-Chari-Lac Tchad: Aspect général des activités de la pêche et de la commercialisation des produits. Description des engins de pêche et leur emploi. ORSTOM
Buckley, S., Usai, D., Jakob, T., Radini, A., & Hardy, K. (2014). Dental calculus reveals unique insights into food items, cooking and plant processing in Prehistoric Central Sudan. PLOS One, 9(7).
Caneva, I. (1983). Pottery-using gatherers and hunters at Saggai (Sudan): Preconditions for food production. Origini, 12, 7–27.
Caneva, I. (1988). El Geili: The history of a Middle Nile environment 7,000 B.C.-A.D. 1,500. B.A.R. International Series 424.
Caneva, I., Garcea, E. A. A., Gautier, A., & Van Neer, W. (1993). Pre-pastoral cultures along the central Sudanese Nile. Quaternaria Nova, 3, 177–252.
Castelletti, L., Castiglioni, E., Cottini, M., & Rottoli, M. (1999). Archaeobotanical evidence at Uan Afuda: Charcoal, wood and seeds from the early Holocene sequence. In S. di Lernia (Ed.), The Uan Afuda Cave: Hunter-gatherer societies of Central Sahara (pp. 131–148). Arid Zone Archaeology, Monographs 1. All’Insegna del Giglio, Firenze.
Clark, J. D. (1989). Shabona: An early Khartoum settlement on the White Nile. In. L. Krzyzaniak and M. Kobusiewicz (Eds.), Late prehistory of the Nile Basin and the Sahara (pp. 387–410). Poznań Archaeological Museum.
Copley, M. S., Berstan, R., Dudd, S. N., Docherty, G., Mukherjee, A. J., Straker, V., et al. (2003). Direct chemical evidence for widespread dairying in Prehistoric Britain. Proceedings of the National Academy of Sciences of the United States of America, 100(4), 1524–1529.
Correa-Ascencio, M., & Evershed, R. P. (2014). High throughput screening of organic residues in archaeological potsherds using direct acidified methanol extraction. Analytical Methods, 6(5), 1330–1340.
Cramp, L., & Evershed, R. P. (2014). Reconstructing aquatic resource exploitation in Human Prehistory using lipid biomarkers and stable isotopes. In H. D. Holland & K. K. Turekian (Eds.), Treatise on Geochemistry (2nd ed., pp. 319–339). Elsevier.
Cremaschi, M., Salvatori, S., Usai, D., & Zerboni, A. (2007). A further tessera to the huge mosaic: Studying the ancient settlement pattern of the El Salha region (south-west of Omdurman, Central Sudan). In K. Kroeper, M. Chlodnicki, & M. Kobusiewicz (Eds.), Archaeology of the earliest North-eastern Africa (pp. 39–48). Poznań Archaeological Museum.
Dal Sasso, G., Lebon, M., Angelini, I., Maritan, L., Usai, D., & Artioli, G. (2016). Bone diagenesis variability among multiple burial phases at Al Khiday (Sudan) investigated by ATR-FTIR spectroscopy. Palaeogeography, Palaeoclimatology, Palaeoecology, 463, 168–179.
Dal Sasso, G., Maritan, L., Salvatori, S., Mazzoli, C., & Artioli, G. (2014). Discriminating pottery production by image analysis: A case study of Mesolithic and Neolithic pottery from Al Khiday (Khartoum, Sudan). Journal of Archaeological Science, 46, 125–143.
Dembitsky, V. M., Goldshlag, P., & Srebnik, M. (2002). Occurrence of dicarboxylic (dioic) acids in some Mediterranean nuts. Food Chemistry, 76(4), 469–473.
Dudd, S. N., & Evershed, R. P. (1998). Direct demonstration of milk as an element of archaeological economies. Science, 282(5393), 1478–1481.
Dunne, J., di Lernia, S., Chłodnicki, M., Kherbouche, F., & Evershed, R. P. (2017). Timing and pace of dairying inception and animal husbandry practices across Holocene North Africa. Quaternary International, 471, 147–159.
Dunne, J., Evershed, R. P., Salque, M., Cramp, L., Bruni, S., Ryan, K., et al. (2012). First dairying in green Saharan Africa in the fifth millennium BC. Nature, 486(7403), 390–394.
Dunne, J., Jórdeczka, M., Chłodnicki, M., Hardy, K., Kubiak-Martens, L., & M-d Hoyo, M. (2021). Holocene resource exploitation along the Nile: Diet and subsistence strategies of Mesolithic and Neolithic societies at Khor Shambat 1. Sudan. Antiquity, 95(384), 1–20.
Dunne, J., Manning, K., Linstädter, J., Mikdad, A., Breeze, P., & Hutterer, R. (2020). Pots, plants and animals: Broad-spectrum subsistence strategies in the Early Neolithic of the Moroccan Rif region. Quaternary International, 555, 96–109.
Elamin, Y. M., & Mohammed-Ali, A. S. (2004). Umm Marrahi. An early Holocene ceramic site, north of Khartoum (Sudan). Sahara, 15, 97–110.
Ertuğ, F. (2000). An ethnobotanical study in Central Anatolia (Turkey). Economic Botany, 54(2), 155.
Evershed, R. P. (2008). Experimental approaches to the interpretation of absorbed organic residues in archaeological ceramics. World Archaeology, 40(1), 26–47.
Fahmy, A. G. (2005). Missing plant macro remains as indicators of plant exploitation in Predynastic Egypt. Vegetation History and Archaeobotany, 14(4), 287–294.
Ferguson, H. (1954). The food crops of the Sudan and their relationship to environment. McCorquodale and Co. (Sudan) Ltd.
Gannes, L. Z., O’Brien, D. M., & Martínez del Rio, C. (1997). Stable isotopes in animal ecology: Assumptions, caveats, and a call for more laboratory experiments. Ecology, 78(4), 1271–1276.
Gautier, A. (1983). Animal life along the Prehistoric Nile: The evidence from Saggai 1 and Geili (Sudan) in pottery using gatherers and hunters at Saggai (Sudan) - preconditions for food productions. Origini. Rivista di Prehistoria e Protostoria delle Civiltà Antiche, 12(1), 50–115.
Gautier, A. (1984). The fauna of the Neolithic site of Kadero (Central Sudan). In L. Krzyzaniak & M. Kobusiewicz (Eds.), Origin and early development of food-producing cultures in North-Eastern Africa (pp. 317–319). Polish Academy of Sciences and Poznań Archaeological Museum.
Gragson, T. L. (1997). The use of underground plant organs and its relation to habitat selection among the Pumé Indians of Venezuela. Economic Botany, 51(4), 377–384.
Haaland, R. (1992). Fish, pots and grain: Early and Mid-Holocene adaptations in the Central Sudan. African Archaeological Review, 10(1), 43–64.
Haaland, R. (1995). Sedentism, cultivation, and plant domestication in the Holocene Middle Nile Region. Journal of Field Archaeology, 22(2), 157–174.
Haaland, R., & Magid, A. A. (1995). Aqualithic sites along the Rivers Nile and Atbara, Sudan. Alma Mater Forlag.
Hammann, S., Cramp, L. J., Whittle, M., & Evershed, R. P. (2018). Cholesterol degradation in archaeological pottery mediated by fired clay and fatty acid pro-oxidants. Tetrahedron Letters, 59(50), 4401–4404.
Hather, J. (1995). Parenchymatous tissues from the Early Neolithic site E-75-6 at Nabta Playa, Western Desert, south Egypt. Preliminary Report. Acta Palaeobotanica, 35, 157–162.
Hillman, G. C. (1989). Late Palaeolithic plant foods from Wadi Kubbaniya in Upper Egypt: Dietary diversity infant weaning, and seasonality in a riverine environment. In D. R. Harris and G. C. Hillman (Eds.), Foraging and farming: The evolution of plant exploitation (pp. 207–239). Unwin Hyman.
Hillman, G. C., Madeyska, E., & Hather, J. (1989). Wild plant foods and diet at Late Paleolithic Wadi Kubbaniya: the evidence from charred remains. In F. Wendorf, R. Schild & A. E. Close (Eds.), The prehistory of Wadi Kubbaniya. Volume 2. Stratigraphy, paleoeconomy, and environment (pp. 162–242). Southern Methodist University Press.
Holloway, P. J. (1984). Cutins and suberins, the polymeric plant lipids. In H. K. Mangold, G. Zweig, & J. Sherma (Eds.), CRC handbook of chromatography lipids, 1, 321–345.
Honegger, M. (2004). Settlement and cemeteries of the Mesolithic and Early Neolithic at el-Barga (Kerma region). Sudan & Nubia, 8, 27–32.
Honegger, M. (2005). El-Barga: Un site clé pour la compréhension du Mésolithique et du début du Néolithique en Nubie. Revue De Paléobiologie, 10, 95–104.
Honegger, M., & Bastien, J. (2009). The early Holocene sequence of Wadi el-Arab. Kerma, 2009, 3–6.
Honegger, M., & Williams, M. (2015). Human occupations and environmental changes in the Nile valley during the Holocene: The case of Kerma in Upper Nubia (northern Sudan). Quaternary Science Reviews, 130, 141–154.
Hosfield, R., White, K., & Drake, N. A. (2015). Middle Stone Age and Early Holocene archaeology in Central Sudan: The Wadi Muqadam geoarchaeological survey. Sudan & Nubia, 19, 16–29.
Iacumin, P., Di Matteo, A., Usai, D., Salvatori, S., & Venturelli, G. (2016). Stable isotope study on ancient populations of central Sudan: Insights on their diet and environment. American Journal of Physical Anthropology, 160(3), 498–518.
Jesse, F. (2000). Early Khartoum Ceramics in the Wadi Howar, Northwest Sudan. In L. Krzyzaniak, K. Kroeper, & M. Kobusiewicz (Eds.), Recent research into the Stone Age of Northeastern Africa (pp. 77–87). Poznań Archaeological Museum.
Jesse, F. (2003). Rahib 80/87. Ein wavy-line-fundplatz im Wadi Howar und die früheste Keramik in Nordafrika. Africa Praehistorica 16. Heinrich-Barth-Institut, Cologne.
Jousse, H., Obermaier, H., Raimbault, M., & Peters, J. (2008). Late Holocene economic specialisation through aquatic resource exploitation at Kobadi in the Méma, Mali. International Journal of Osteoarchaeology, 18(6), 549–572.
Kern, J. H. (1974). Cyperaceae. In C. G. G. J. van Steenis (Ed.), Flora Malesiana Series 1, 7(3), 435–753.
Khabir, A. R. (1987). New radiocarbon dates for Sarurab 2 and the age of the early Khartoum tradition. Current Anthropology, 28, 377–380.
Kolattukudy, P. E. (1980). Biopolyester membranes of plants: Cutin and suberin. Science, 208(4447), 990–1000.
Kolattukudy, P. E. (1981). Structure, biosynthesis, and biodegradation of cutin and suberin. Annual Review of Plant Physiology and Plant Molecular Biology, 32, 539–567.
Kolattukudy, P. E., Croteau, R., & Buckner, R. S. (1976). Biochemistry of plant waxes. In. P. E. Kolattukudy (Ed.), Chemistry and biochemistry of natural waxes (pp. 289–347). Elsevier.
Krzyzaniak, L. (1991). Early farming in the Middle Nile Basin: Recent discoveries at Kadero (Central Sudan). Antiquity, 65(248), 515–532.
Kulhari, O. P., & Joshi, P. (1992). Fodder plants of Shekhawati region (Rajasthan). Journal of Economic and Taxonomic Botany: Addditional Series, 10, 355–370.
Kunst, L., & Samuels, A. L. (2003). Biosynthesis and secretion of plant cuticular wax. Progress in Lipid Research, 42(1), 51–80.
Linseele, V. (2020). A diachronic overview of aquatic adaptations at Al-Khiday (Central Sudan, ca. 7000–4000 cal BC). In: D. Usai, S. Tuzzato, & M. Vidale (Eds.), Tales of three worlds - archaeology and beyond: Asia, Italy, Africa. A tribute to Sandro Salvatori (pp. 289–301). Archaeopress.
Linseele, V., & Zerboni, A. (2018). Done with fish? A diachronic study of fishing in the Holocene Nile basin of Sudan. Quaternary International, 471, 229–240.
Macri, P. (2019). Studio Isotopico di resti Mesolitici di Pila wernei: ricostruzione delle condizioni climatiche e ambientali tra 8700 e 7500 BP nel Sudan Centrale. Ph.D. thesis, University of Parma.
Manniche, L. (1989). An ancient Egyptian herbal. University of Texas Press.
Maritan, L., Iacumin, P., Zerboni, A., Venturelli, G., Dal Sasso, G., & Linseele, V. (2018). Fish and salt: The successful recipe of White Nile Mesolithic hunter-gatherer-fishers. Journal of Archaeological Science, 92, 48–62.
Marks, A. E., & Mohammed-Ali, A. (1991). The late prehistory of the Eastern Sahel: The Mesolithic and Neolithic of Shaqadud. Southern Methodist University Press.
Marlowe, F. W., & Berbesque, J. C. (2009). Tubers as fallback foods and their impact on Hadza hunter-gatherers. American Journal of Physical Anthropology, 140(4), 751–758.
Marshall, F., & Hildebrand, E. (2002). Cattle before crops: The beginnings of food production in Africa. Journal of World Prehistory, 16(2), 99–143.
Mattalia, G., Quave, C. L., & Pieroni, A. (2013). Traditional uses of wild food and medicinal plants among Brigasc, Kyé, and Provençal communities on the Western Italian Alps. Genetic Resources and Crop Evolution, 60(2), 587–603.
Meena, A. K., Yadav, A. K., Niranjan, U. S., Singh, B., Nagariya, A. K., & Verma, M. (2010). Review on Cyperus rotundus—A potential herb. International Journal of Pharmaceutical and Clinical Research, 2(1), 20–22.
Mekem Sonwa, M. (2000). Isolation and structure elucidation of essential oil constituents: comparative study of the oils of Cyperus alopecuroides, Cyperus papyrus, and Cyperus rotundus. Ph.D. thesis, University of Hamburg.
Mekem Sonwa, M., & König, W. A. (2001). Chemical study of the essential oil of Cyperus rotundus. Phytochemistry, 58(5), 799–810.
Mukherjee, A. J., Copley, M. S., Berstan, R., Clark, K. A., & Evershed, R. P. (2005). Interpretation of δ13C values of fatty acids in relation to animal husbandry, food processing and consumption in prehistory. In J. Mulville & A. Outram (Eds.), The zooarchaeology of milk and fats (pp. 77–93). Oxbow Books.
Negbi, M. (1992). A sweetmeat plant, a perfume plant and their weedy relatives: A chapter in the history of Cyperus esculentus L. and C. rotundus L. Economic Botany, 46(1), 64–71.
Ochoa, J. J., & Ladio, A. H. (2015). Current use of wild plants with edible underground storage organs in a rural population of Patagonia: Between tradition and change. Journal of Ethnobiology and Ethnomedicine, 11, 70.
Out, W. A., Ryan, P., García-Granero, J. J., Barastegui, J., Maritan, L., Madella, M., & Usai, D. (2016). Plant exploitation in Neolithic Sudan: A review in the light of new data from the cemeteries R12 and Ghaba. Quaternary International, 412(Part B), 36–53.
Parsons, W. T., & Cuthbertson, E. G. (1992). Noxious weeds of Australia. Inkata Press.
Petrick, B. (2012). The Cologne Fourth Cataract Project 2005 field season on Boni Island. In: H.-P. Wotzka (Ed.), Proceedings of the Third International Conference on the Archaeology of the Fourth Nile Cataract, University of Cologne, 13–14 July 2006. Africa Praehistorica 22 (pp. 117–128). Heinrich-Barth-Institut, Cologne.
Pollard, M., Beisson, F., Li, Y., & Ohlrogge, J. B. (2008). Building lipid barriers: Biosynthesis of cutin and suberin. Trends in Plant Science, 13(5), 236–246.
Pöllath, N. (2012). Bones from Boni: Preliminary results of the faunal analyses. In: H.-P. Wotzka (Ed.), Proceedings of the Third International Conference on the Archaeology of the Fourth Nile Cataract, University of Cologne, 13–14 July 2006. Africa Praehistorica 22 (pp. 187–196). Heinrich-Barth-Institut, Cologne.
Reimer, P. J., Austin, W. E. N., Bard, E., Bayliss, A., Blackwell, P. G., Bronk Ramsey, C., et al. (2020). The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon, 62, 725–757.
Ryan, P., Out, W. A., Garcia-Granero, J. J., Madella, M., & Usai, D. (2016). Plant microremains from the white deposits and dental calculus from Ghaba and R12 cemeteries. Identification and implication. In: S. Salvatori, D. Usai, & Y. Lecointe (Eds.), Ghaba. An Early Neolithic cemetery in Central Sudan (pp. 109–120). Africa Magna Verlag.
Salvatori, S. (2012). Disclosing archaeological complexity of the Khartoum Mesolithic: New data at the site and regional level. African Archaeological Review, 29(4), 399–472.
Salvatori, S., & Usai, D. (2006). Archaeological research south of Omdurman. A preliminary assessment on ceramic and lithic materials from 10-X-6 multistratified mound site along the western bank of the White Nile in Central Sudan. Archéologie Du Nil Moyen, 10, 203–220.
Salvatori, S., & Usai, D. (2009). El Salha Project 2005: New Khartoum Mesolithic sites from Central Sudan. Kush, 19, 87–96.
Salvatori, S., & Usai, D. (2019). The Mesolithic and Neolithic in Sudan. In D. Raue (Ed.), Handbook of Ancient Nubia (pp. 171–193). DeGruyter.
Salvatori, S., Usai, D., & Zerboni, A. (2011). Mesolithic site formation and palaeoenvironment along the White Nile (Central Sudan). African Archaeological Review, 28(3), 177–211.
Salvatori, S., Usai, D., Faroug, M. A., Di Matteo, A., Iacumin, P., Linseele, V., & Magzoub, M. K. (2014). Archaeology at Al Khiday: New insight on the prehistory and history of Central Sudan. In J. R. Anderson & D. A. Welsby (Eds.), The Fourth Cataract and beyond (pp. 243–257). Proceedings of the 12th International Conference of Nubian Studies. Peeters.
Salvatori, S., Usai, D., & Zerboni, A. (2018). New evidence from the prehistoric sites at Al Khiday and Al Jamrab, Central Sudan. In J. Kabacinski, M. Chlodnicki, M. Kobusiewicz and M. Winiarska-Kabacinska (Eds.), Desert and the Nile. Prehistory of the Nile Basin and the Sahara. Papers in honour of Fred Wendorf (pp. 71–94). Studies in African Archaeology, 15. Poznań Archaeological Museum.
Smith, L. M. V. (1998). SARS Survey from Omdurman to Gabolab 1997: Pottery and small finds. Sudan & Nubia, 2, 45–52.
Sukova, L., & Varadzin, L. (2012). Preliminary report on the exploration of Jebel Sabaloka (West Bank) 2009–2012. Sudan & Nubia, 16, 118–131.
Sutton, J. E. G. (1977). The African aqualithic. Antiquity, 51, 25–34.
Tulloch, A. P. (1976). Chemistry of waxes of higher plants. In P. E. Kolattukudy (Ed.), Chemistry and biochemistry of natural waxes (pp. 235–287). Elsevier.
Usai, D. (2014). Recent advances in understanding the Prehistory of Central Sudan. In J. R. Anderson and D. A. Welsby (Eds.), The Fourth Cataract and beyond (pp. 31–44). Proceedings of the 12th International Conference of Nubian Studies. Peeters.
Usai, D. (2016). A picture of Prehistoric Sudan: The Mesolithic and Neolithic periods. Oxford Handbooks Online. https://doi.org/10.1093/oxfordhb/9780199935413.013.56
Usai, D., & Salvatori, S. (2002). The Is.I.A.O. El-Salha Archaeological Project. Sudan & Nubia, 6, 67–72.
Usai, D., & Salvatori, S. (2005). The Is.I.A.O. archaeological project in the El Salha area (Omdurman South, Sudan): Results and perspectives. Africa, 40(3–4), 474–493.
Usai, D., & Salvatori, S. (2019). The Mesolithic period on the White Nile region and the al-Khiday sites. Azania: Archaeological Research in Africa, 54(4), 445–467.
Usai, D., Salvatori, S., Iacumin, P., Di Matteo, A., Jakob, T., & Zerboni, A. (2010). Excavating a unique pre-Mesolithic cemetery in central Sudan. Antiquity, 84, 16–18.
Usai, D., Salvatori, S., Jakob, T., & David, R. (2014). The Al Khiday Cemetery in Central Sudan and its ‘‘Classic/Late Meroitic” Period Graves. Journal of African Archaeology, 12, 183–204.
Usai, D., Maritan, L., Dal Sasso, G., Artioli, G., Salvatori, S., Jakob, T., & Salviato, T. (2017). Late Pleistocene/Early Holocene evidence of prostatic stones at Al Khiday Cemetery, Central Sudan. PLoS ONE, 12(1). DOI: https://doi.org/10.1371/journal.pone.0169524
Van Neer, W. (1995). Analysis of the fish remains. In S. K. McIntosh (Ed.), Excavations at Jenne´-Jeno, Hambarketolo, and Kaniana (Inland Delta, Mali), the 1981 season (pp. 319–347). University of California Press.
Varadzinová, L., Varadzin, L., Brukner Havelková, P., Crevecoeur, I., & Garcea, E. A. A. (2022). Archaeology of Holocene hunter-gatherers at the sixth Nile Cataract, Central Sudan. Bulletin D’Archeologié Marocaine, 27, 101–118.
Vickers, W. T., & Plowman, T. (1984). Useful plants of the Siona and Secoya Indians of eastern Ecuador. Fieldiana, 15, 1–63.
Vincent, A. S. (1985). Plant foods in savanna environments: A preliminary report of tubers eaten by the Hadza of northern Tanzania. World Archaeology, 17(2), 131–148.
Walton, T. J. (1990). Waxes, cutin and suberin. In J. L. Harwood & J. R. Bowyer (Eds.), Methods in plant biochemistry (Vol. 4, pp. 105–158). Academic Press.
Wasylikowa, K., et al. (2001). Site E-75-6: Vegetation and subsistence of the Early Neolithic at Nabta Playa, Egypt, reconstructed from charred plant remains. In F. Wendorf & R. Schild (Eds.), Holocene Settlement of the Egyptian Sahara (pp. 544–591). Springer.
Wendorf, F., Close, A. E., Schild, R., Wasylikowa, K., Housley, R. A., Harlan, J. R., & Krolik, H. (1992). Saharan exploitation of plants 8,000 years BP. Nature, 359, 721–724.
West, J. B., Bowen, G. J., Dawson, T. E., & Tu, K. P. (2010). Isoscapes: Understanding movement, pattern and process on Earth through isotope mapping. Springer.
Wetterstrom, W. (1993). Foraging and farming in Egypt: The transition from hunting and gathering to horticulture in the Nile valley. In B. Andah, A. Okpoko, T. Shaw, & P. Sinclair (Eds.), The archaeology of Africa: Food, metal and towns (pp. 165–226). Routledge.
Wickens, G. E. (1982). Palaeobotanical speculations and Quaternary environments in the Sudan. In M. A. J. Williams & D. Adamson (Eds.), A land between two Niles (pp. 23.-50). Balkema, Rotterdam.
Williams, M. A. J. (2009). Late Pleistocene and Holocene environments in the Nile basin. Global and Planetary Change, 69(1–2), 1–15.
Williams, M. A. J., & Adamson, D. (1980). Late Quaternary depositional history of the Blue and White Nile rivers in central Sudan. In. M. A. J. Williams & H. Faure (Eds.), The Sahara and the Nile: Quaternary environments and prehistoric occupation in northern Africa (pp. 281–304). Balkema, Rotterdam.
Williams, M. A. J., Usai, D., Salvatori, S., Williams, D. M., Zerboni, A., Maritan, L., & Linseele, V. (2015). Late Quaternary environments and prehistoric occupation in the lower White Nile valley, central Sudan. Quaternary Science Reviews, 130, 72–88.
Wotzka, H.-P., Goedicke, C., & Michels, M. (2012). Boni Island radiocarbon and OSL dates. In H.-P. Wotzka (Ed.), Proceedings of the Third International Conference on the Archaeology of the Fourth Nile Cataract, University of Cologne, 13–14 July 2006. Africa Praehistorica 22. (pp. 137–176). Heinrich-Barth-Institut.
Zerboni, A. (2011). Micromorphology reveals in situ Mesolithic living floors and archaeological features in multiphase sites in central Sudan. Geoarchaeology, 26(3), 365–391.
Zerboni, A., Salvatori, S., Vignola, P., Ali, M. A., & Usai, D. (2018). The long-distance exchange of amazonite and increasing social complexity in the Sudanese Neolithic. Antiquity, 92(365), 1195–1209.
Acknowledgements
The authors thank the Leverhulme Trust (RPG-2016-115) for funding for ‘Peopling the Green Sahara? A multi-proxy approach to reconstructing the ecological and demographic history of the Saharan Holocene’. The authors wish to thank the NERC for partial funding of the National Environmental Isotope Facility (NEIF; contract no. NE/V003917/1) and NERC (contract no. NE/V003917/1) and funding from the European Research Council under the European Union’s Seventh Framework Programme (FP/2007-2013) and European Research Council Grant Agreement number 340923 for funding GC-MS capabilities, together with NERC (contract no. NE/V003917/1) and the University of Bristol for funding the GC-IRMS capabilities. Ian Bull, Alison Kuhl and Helen Whelton are thanked for technical help. Fieldwork was funded by the Italian Ministry of Foreign Affairs and the Centro Studi Sudanesi e Sub-Sahariani. We also thank the Director and Staff of the National Corporation for Antiquities and Museums, Khartoum, Sudan, who granted permissions for excavation.
Author information
Authors and Affiliations
Contributions
Julie Dunne: Conceptualisation, Writing-Original draft preparation, Investigation; Sandro Salvatori: Conceptualisation, Investigation, Resources, Writing-Review and Editing; Lara Maritan: Investigation, Resources; Writing-Review and Editing; Katie Manning: Conceptualisation, Writing-Review and Editing; Veerle Linseele: Investigation, Resources, Writing-Review and Editing; Toby Gillard: Investigation; Paul Breeze: Writing-Review and Editing; Nick Drake: Writing-Review and Editing; Richard Evershed: Conceptualisation, Writing-Review and Editing; Donatella Usai: Conceptualisation, Investigation, Resources, Writing-Original draft preparation.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Dunne, J., Salvatori, S., Maritan, L. et al. Wild Food: Plants, Fish and Small Animals on the Menu for Early Holocene Populations at al-Khiday, Central Sudan. Afr Archaeol Rev 39, 255–281 (2022). https://doi.org/10.1007/s10437-022-09490-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10437-022-09490-w