
Abstract
To reinforce Eurasian lynx populations in central Europe, 20 lynx from Slovakia and Switzerland were translocated to the Palatinate Forest between 2016 and 2020. Using a systematic camera trapping array consisting of 80 camera trapping sites in a 1,000 km² study area located in the centre of the approximately 1,800 km² Palatinate Forest, we aim to describe the status of the population in the final stages of the reintroduction project in winter and spring 2019/20 and 2020/21. We also use our data to provide a first estimate of population density of the newly established population. With an estimate of 0.52 independent individuals per 100 km², population density in the Palatinate Forest was still significantly lower than the densities of well-established reintroduced populations. The number of independent individuals detected in the study area decreased from 15 individuals in 2019/20 to 11 individuals in 2020/21, thus significantly below the number of lynx translocated. The low abundance in the Palatinate Forest can be explained by the dispersal of several individuals to the Vosges (France), which, together with the Palatinate Forest, form a continuous area of suitable lynx habitat of approximately 8,000 km². Our results may thus reflect the status of a young population that can expand over a potentially large area. Nevertheless, in light of the low population density, we strongly recommend a synchronized and harmonized transboundary monitoring program to keep track of the development of this important Franco-German lynx population. In case population density remains low, supportive measures need to be considered.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
West-central Europe was home to three species of large carnivores: The brown bear (Ursus arctos), the wolf (Canis lupus), and the Eurasian lynx (Lynx lynx) (Chapron et al. 2014). Whereas brown bears and wolves have never completely disappeared from western Europe, Eurasian lynx had been completely extirpated by the beginning of the 20th century (Boitani and Linnell 2015). Autochthonous lynx populations only survived in northern and eastern Europe (Chapron et al. 2014; Lucena-Perez et al. 2020). Moreover, while wolves, and to some extent also brown bears, have naturally re-colonized parts of their historical range (Boitani and Linnell 2015), lynx exhibit a very conservative dispersal behaviour (Zimmermann et al. 2007), which limits their prospects of naturally re-colonizing western Europe. Consequently, efforts have been made since the 1970s to reintroduce lynx to western Europe, mostly by translocating individuals from autochthonous populations in east-central Europe (Breitenmoser and Breitenmoser-Würsten 2008; Linnell et al. 2009; Boitani and Linnell 2015).
Owing to these reintroduction projects, there are now several newly established lynx populations in central Europe (Chapron et al. 2014). Many of these populations have grown to a size where they can be considered demographically stable. However, these populations have so far not expanded significantly beyond the areas of reintroduction, and remain isolated, both from each other, and from autochthonous populations in east-central and eastern Europe (Breitenmoser et al. 2021). As a consequence, there is no genetic exchange between populations, which has led to a significant decline in genetic diversity in many of the reintroduced populations (Mueller et al. 2022). To ensure the long-term viability of the Eurasian lynx in central Europe, further reintroduction projects are required to establish new populations in suitable areas, and eventually, to connect the so far isolated populations to a central European metapopulation (Bonn Lynx Expert Group 2021).
As part of this metapopulation strategy, a reintroduction project was carried out in the Palatinate Forest in southwest Germany between 2016 and 2021. The project pursued the goal to establish a viable population nucleus in the transboundary UNESCO Biosphere Reserve “Pfälzerwald-Vosgeses du Nord” (Idelberger et al. 2021). Ultimately, this population nucleus should become part of the Upper Rhine Metapopulation, spanning from the Palatinate Forest in the north, across the Vosgeses (France) and the Black Forest (Germany) to the Jura Mountains (France and Switzerland) in the south (Krebühl et al. 2021). To reach this goal, a total number of 20 lynxes (12 females, 8 males) had been translocated from the autochthonous population in the Slovak Carpathian Mountains as well as from reintroduced populations in Switzerland to the Palatinate Forest. All translocated individuals belonged to the phylogenetic lineage of the Carpathian lynx (L. l. carpathicus). Upon translocation, they were equipped with GPS/ GSM collars to monitor their fate and ranging behaviour approximately 1–1.5 years after release (Idelberger et al. 2021). Translocations started in spring 2016 and continued until spring 2020, when the last three individuals were released.
The project started very successfully. A first reproduction was already recorded in spring 2017, and in the following years a minimum of 2–3 females per year reproduced successfully in their new environment (Idelberger et al. 2021). Moreover, the project was preceded and accompanied by extensive, transboundary activities to involve stakeholders and to raise public acceptance for the project (Stiftung Natur und Umwelt Rheinland-Pfalz 2021). Despite a few losses due to traffic accidents or emigrations from the project area, it was thus to be expected that the population nucleus in the Palatinate Forest would soon grow in size.
The present study aims to describe the status of the population in the last two years of the project in 2020 and 2021, i.e. in the year of the last releases, and the year following the last releases. We used systematic camera trapping surveys in a 1,000 km² study area located in the centre of the approximately 1,800 km² Palatinate Forest. Our aim was to get a comprehensive overview of the number of lynxes present in the study area. Specifically, we wanted to know how many of the translocated individuals were still present by the end of the project, and how many additional individuals were added to the population (offspring or immigration). Moreover, we used our data within a spatially explicit capture-recapture framework (Borchers and Efford 2008; Royle et al. 2013) to estimate population density of the Palatinate lynx population in the year following the last releases.
Methods
Study area
The Palatinate Forest is located in south-west Germany in the German federal state of Rhineland-Palatinate (49.29359 N, 7.85149 E). It is one of Germany’s largest connected forest areas with a size of approximately 1,800 km². The Palatinate Forest can roughly be divided into two parts: Whereas the northern part is dominated by elongated mountain ridges divided by narrow valleys, the southern part, known as Wasgau, is characterized by soft conical hills and broad, open valleys. This latter type of landscape continues southwards across the French border into the Vosges du Nord. Further south, the Vosges du Nord are connected with the Vosges Moyenne and Vosges Hautes (central and high Vosges). Together with the Vosges, the Palatinate Forest forms an area of suitable lynx habitat of approximately 8,000 km². In the north and west, the Palatinate Forest merges into a landscape characterized by a mixture of small forest patches and agricultural lands. By contrast, the eastern border is formed by a sharp transition into agricultural lands dominated by vineyards. The highest elevation in the Palatinate Forest is 673 m.
The area of the Palatinate Forest is almost completely covered by forest. Only about 20% of the area consist of settlements or are used for agriculture. The forest consists mainly of broad leaf or mixed forest dominated by European beech (Fagus sylvatica) and Baltic pine (Pinus sylvestris). Even though the forest is used for timber production and a variety of recreational activities (e.g., hunting, hiking, mountain biking, climbing) it supports a diverse community of medium-sized and large mammals, such as roe deer (Capreolus capreolus), red deer (Cervus elaphus), wild boar (Sus scrofa), European badger (Meles meles), and red fox (Vulpes vulpes). Moreover, the Palatinate Forest is one of the most important distribution areas of the European wildcat (Felis silvestris) in Germany (Steyer et al. 2016).
With an annual mean precipitation of 700–800 mm and an annual average temperature from 10 to 11.5 °C (Rheinland-Pfalz Kompetenzzentrum für Klimawandelfolgen 2023), the Palatinate Forest is characterized by moderately cool summers and mild and humid winters.
Camera trapping
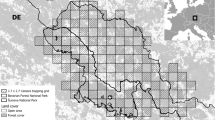
The study area for the camera trapping surveys was located in the centre of the Palatinate Forest and comprised an area of 1,000 km² (Fig. 1). We divided this area into grid cells of 2.5 × 2.5 km length and located a camera trapping site in every second cell of this grid (similar to Pesenti and Zimmermann 2013; Weingarth et al. 2015; Palmero et al. 2021). We planned to set up 80 camera trapping sites in each year of the study, resulting in eight sites per 100 km². In some cases, however, we had to deviate from this schedule, usually because the forest area in the designated grid cell was too small.

Map of the study area (red) and the state space used for SCR analyses (orange) plotted against the forest areas of the Palatinate Forest and the Vosges
The camera trapping sites were located on forest paths or forest roads (dirt or gavel roads used for forest management), as this approach has been proven highly successful in previous camera-trapping surveys of lynx in similar environments (e.g., Weingarth et al. 2015; Port et al. 2021; Palmero et al. 2021). We installed two camera traps per station, one at each side of the road or path, to obtain pictures of both flanks of a passing animal. Camera traps were secured inside metal boxes, locked with a padlock or cable lock, and attached to a tree or a pole approximately 50 cm above ground. Camera traps were checked every 4–6 weeks to replace batteries and SD cards.
We used white-flash cameras of the models Cuddeback Attack and C1 (Cuddeback, Green Bay, USA). Delay time between successive photographs was set to the shortest time frame possible (approximately 1–30 s, depending on camera type and time of the day).
In the first year of the study, cameras were set up in November 2019, such that all cameras were active from 01 December 2019. Cameras were taken down from 19 April 2020 such that cameras operated between 01 December and 18 April over an overall period of 139 days. However, owing to delayed permissions to set up cameras in some forest properties, 18 trapping sites could not be realised in the study period 2019/20, reducing the number of operating trapping sites to 62. As a result, the overall trapping effort during the study period 2019/20 was 8,618 trapping days (camera trapping sites x number of days cameras were installed).
In the second year of the study, cameras were set up between mid-November and mid-December 2020, and taken down from 19 April 2021. All 80 sites could be realized. 30 sites (37.5%) were sites already used during the first survey, the remaining 50 sites were newly selected for the second survey. Cameras operated for a total period of 124 days between 16 December 2020 and 18 April 2021. The overall trapping effort was 9,920 trapping days.
Identification and sexing of lynx
As many other striped or spotted felids, Eurasian lynx can be identified based on their unique fur pattern (Pesenti and Zimmermann 2013; Port et al. 2021; Palmero et al. 2021). All camera trap images of lynx were thus compared to images of already known individuals. Known individuals could either be translocated individuals or known offspring of translocated individuals. For all translocated individuals images of both body flanks were available. These images were taken when the animals were anesthetized for the equipment of the GPS/ GSM collar. Images of known offspring were usually available as camera trap images obtained during opportunistic camera trapping sessions, usually at lynx kills. Images of known offspring were often available from only one body flank, but usually, the second body flank of these individuals could be complemented when they were camera-trapped by both cameras at our sites. The individual identification was carried out by two observers independently, following a protocol used by Port et al. (2021): Each observer was asked independently to rate the lynx shown on the image as either an already known individual (translocated individual or known offspring) or as a yet unknown individual. If observers were unsure about the identity of the lynx, they were asked to rate the photo as “uncertain”. An image was given an identification code (ID, indicating a yet unknown lynx), or was classified as an already known individual, if both observers agreed on their assessment of the image. Otherwise, the image was discarded from analysis.
The sex of all translocated individuals was known. In some cases, other individuals could by sexed based on camera trap images if genitals were clearly visible, or if females were photographed together with their kittens. However, the sex of some individuals captured during this study could not be determined. Voucher photographs of all individuals are provided in the online supplementary material (as recommended by Choo et al. 2020).
Estimating abundance
The major aim of this study was to get a comprehensive overview of the number of individuals present in the Palatinate Forest at the end of the reintroduction project. A 1,000 km² study area located in the centre of the approximately 1,800 km² Palatinate Forest should ensure that all individuals present had a very good chance to be camera trapped over the course of the study. Moreover, the time of data collection in winter and spring was chosen in a way to maximize detection success (Port et al. 2021) and to be able to possibly also capture peripheral individuals (particularly males) during mating season excursions. The number of individuals reported in the results section is, therefore, first and foremost a description of the minimum number of individuals present in the study area.
To get an impression of how successful our survey was in detecting all individuals present in the study area, and how many individuals we might have missed, we performed an abundance estimation using conventional (i.e., non-spatial) capture-recapture models (Otis et al. 1978; Chao 2001). This estimation was carried out only for the second study period in winter 2020/21. The reason is that, at the end of the first study period in spring 2020, three individuals were still translocated to the Palatinate Forest. In other words, translocations were still ongoing, and any newly translocated individual would have to be excluded from analysis. The resulting abundance estimation, therefore, would have been incomplete and inaccurate. During the second study period in winter 2020/21, we restricted the period of data analysis to the last 84 days of the survey period between 25 January and 18 April 2021. This was done to increase the likelihood of meeting the assumption of population closure underlying closed capture-recapture models (see e.g., Pesenti and Zimmermann 2013; Weingarth et al. 2015).
Next, we divided the continuous period of data collection between 25 January and 18 April into twelve discrete capture occasions of seven days each. Based on these capture occasions, we then created capture histories for all lynx individuals detected during the period of analysis, noting whether or not the lynx was detected in a given occasion.
Non-spatial capture-recapture analyses were carried out in R (R Core Team 2021) using the package Rcapture (Rivest and Baillargeon 2015). Rcapture calculates various models: The simplest model (the null model M0) assumes that the capture probability is the same for all individuals and capture occasions. More complex models assume, for example, that the capture probability differs across occasions (Mt) or between individuals (Mh). We used Akaike’s Information Criterion corrected for small sample sizes (AICc) to assess the fit of the various models to our data and to select the most appropriate model.
Estimating population density
In addition to the analyses described above, we used our data to provide a first estimation of population density for the newly established population nucleus in the Palatinate Forest. We are aware that the time of the survey might not be ideal for density estimations, as possible influxes of individuals during the mating season might bias density estimates. We will come back to this point in the discussion, and stress that density estimation was only a secondary goal of this study.
For reasons stated in the previous section, density estimations were also restricted to the second study period in winter 2020/21. As in the analyses above, the period of data analysis was restricted to 84 days between 25 January and 18 April, and the survey period was divided into twelve capture occasions of seven days each.
Population density estimation was carried out using maximum likelihood-based spatial capture-recapture models implemented in the R package secr (Efford 2023). In addition to the information of the capture histories of the non-spatial analyses, capture histories of spatial analyses require the information at which trapping sites an individual was detected during each occasion. Each line of the capture histories thus contained the lynxID, the sampling occasion and the trapping site of the detection event. Entries were restricted to only one event per individual per sampling occasion per trapping site as required by the detector type ‘proximity’ (Zimmermann and Foresti 2016). In spatial capture-recapture (SCR) models detection is modelled as a decreasing function of distance between an individual’s activity centre (where detection probability is assumed to be highest) and the trapping station. Specifically, we used a half-normal detection function with intercept g0 and scale parameter σ, where g0 gives the detection probability of an individual at its activity centre, and σ the decline in capture probability with increasing distance from the activity centre (Borchers and Efford 2008).
We ran four different models: First, we ran a null model (M0) in which g0 and σ are assumed to be constant across individuals. Next, we ran three finite mixture models (Pledger 2000; Borchers and Efford 2008). These models assume that the sampled population consists of two (or more) latent classes of individuals differing in their detection parameters (g0 and σ). secr then allocates individuals to these latent classes, such that classes with relatively homogeneous detection parameters are created. Finite mixture models are particularly useful in cases where individuals of the sampled population are expected to differ in detection parameters (for example males and females), but not all sampled individuals can be assigned reliably to these classes. This is the case in our study, because we were not able to determine the sex of all sampled individuals.
The three finite mixture models calculated in our study were: model Mg in which g0 was assumed to differ between two latent classes of individuals, model Mσ, in which σ was assumed to differ between two latent classes of individuals, and model Mgσ, in which both g0 and σ were assumed to differ between two latent classes of individual. We used Akaike’s Information Criterion corrected for small sample sizes (AICc) for model selection, and used all models with AICc < 5 for model averaging.
In addition to the capture histories, SCR also requires information of the state space, which is the area, in which the potential activity centres of the individuals detected may be located. To determine the state space, we created a habitat mask with a spacing of 1000 m between potential activity centres, and buffer zones of varying sizes around the network of camera stations. Following a procedure proposed by Zimmermann and Foresti (2016), we then sought the smallest buffer zone for which the density estimate of the null model did no longer change. This was the case with a buffer of 15 km, resulting in a state space of 4450 km². In the next step, we excluded habitat from this area that we deemed unsuitable for lynx. To this end, we excluded all area east of the sharp transition zone between the Palatinate Forest and the agricultural lands of the Rhine valley (see study area). The state space contained large parts of the Vosges du Nord as well as of the fragmented transition zone north and west of the Palatinate Forest, where potential home range centres could be located in forest, greenland and meadow habitat (habitat layers retrieved from OpenStreetMap). The resulting state space had a size of 2357 km² (Fig. 1).
Results
Survey effort and detection success
Owing to technical issues a substantial amount of data was lost during the first study period (2019/20). As a result, the amount of data available for analyses reduced to 6,539 effective trap days, which is only 76% of the potential trap days (see Methods). During the second study period, four cameras were stolen at two sites, but otherwise, at least one camera at each site was active at all other 78 sites. The number of effective trap days in 2020/21 (= number of survey days x number of sites, at which at least one camera was active) was 9,824 (99% of potential trap days).
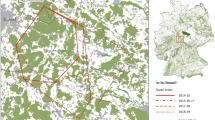
During the first study period, at least one lynx was detected at 33 of the 62 sites (53.2%). The number of lynx detections at the 33 successful sites ranged between one and 14 detections (mean ± SD = 3.71 ± 3.48). Successful sites were mainly located in the central, western and south-western parts of the Palatinate Forest (Fig. 2). During the second study period, the number of successful sites was 34 of 80 sites (42.5% of sites). The number of lynx detections at these 34 sites ranged between one and 12 detections (mean ± SD = 3.44 ± 3.04). The distribution of successful sites was similar as in the first study period (Fig. 2), yet a few detections also occurred in the northern parts of the Palatinate Forest.

Lynx detection frequencies during the first study period (2019/20, left panel) and during the second study period (2020/21, right panel). Dots depict camera trapping sites at which lynx were detected (with the size of the dot indicating detection frequency), crosses depict camera trapping sites at which no lynx was detected
During the first study period, a total number of 264 images of lynx were taken. These 264 photos were distributed across 114 independent detections of lynx. In relation to a realized sampling effort of 6,539 effective trap days, the detection rate was 1.74 detections per 100 effective trap days. During the second study period, 215 images were taken in 116 independent detection events. In relation to a realized sampling effort of 9,824 effective trap days, the detection rate in the second study period declined to 1.18 detections per 100 effective trap days.
Individual identification of lynx
Individual identification of lynx was possible for 106 of the 114 detections recorded during the first study period (93%). Eight detections had to be discarded from further analyses, either because one observer or both observers rated the image as “uncertain”. Uncertain images were usually either blurry, or contained only few body parts insufficient for individual identification. For data collected during the second study period, individual identification was possible for 112 of 116 detections (96.6%). Four detections had to be discarded from analyses because both observers rated the images as “uncertain”. Agreement between observers was very high, and it is worth mentioning that, in no case, observers assigned images to different individuals. In other words, when observers disagreed, one observer attempted an identification whereas the other observer rated the image as “uncertain” (leading to the exclusion of the image).
Number and identity of lynx
In the first study period, a total number of 16 independent lynx were detected (Table 1). However, two individuals (Pip and Twik) were juvenile lynx first detected without their mother towards the end of the study period (Table 1). The number of adult or subadult lynx (> 1 year) was thus 14 individuals. Nine of these 14 individuals were translocated individuals (two of them were translocated in spring 2020, shortly before the end of the survey), and five were known offspring of translocated females born in 2017 and 2018. In other words, no individual was detected that was not already known to the project through opportunistic monitoring.
One translocated male (Juri) died of disease during the course of the survey. Two translocated females were detected with offspring, one female (Mala) with three kittens, the other female (Gaupa) with two kittens. Another adult female (Rosa) was regularly detected during our survey. However, in none of the nine detection was she detected with offspring. Despite successful reproduction in 2018, we assume, therefore, that this female did not reproduce in 2019.
In summary, 15 independent lynx individuals were detected in the study area at the end of the first survey: 14 adult individuals – one fatality + two offspring born in spring 2019. Five of these individuals were females, six were males, and in four cases the sex could not be determined.
In the second study period, a total number of 11 independent lynx were detected (Table 1). One of these individuals was an offspring of a translocated female (Gaupa) born in spring 2020. One other lynx was an unknown individual, yet likely this individuals was also a juvenile, presumably an offspring not detected in the opportunistic monitoring. Hence, in the second study period, we detected nine adult individuals in the study area.
Seven individuals detected until the end of the first study period could not be detected again during the second study period, including three adult males (Brano, Wrano, Palu) that were detected regularly during the first survey. Another individual, the female Rosa, was also not re-detected during the second study period, as she shifted her home range to just outside the study area at the western edge of the Palatinate Forest.
One translocated male (Libre) was not detected during the first survey, but was detected regularly in the southern parts of the study area in the second survey. This male had a transboundary home range mostly restricted to the Vosges du Nord in 2019/20 but extended northwards in 2020/21.
In summary, 11 independent lynx individuals were detected in the study area at the end of the second survey period. Four of these individuals (36.4%) were lynx translocated to the Palatinate Forest. Three individuals were females, four individuals were males. The sex of the remaining four individuals could not be determined. None of the three females was detected with offspring. However, two females were detected only twice, leaving a high chance that potential offspring might have been missed.
Abundance estimation
To get an impression of how successful our survey was to detect all lynx in the study area and how many lynx we might have missed, we performed an abundance estimation using conventional (non-spatial) capture-recapture techniques. This estimation was carried out for the second study period only (see Methods). Moreover, we excluded the two (supposedly) juvenile individuals from the analyses, as they likely reached independence during the survey, and including them would have violated the assumption of a demographically closed population. Our analyses thus included nine adult individuals.
Virtually all models estimated an abundance of exactly nine individuals (Table 2). Standard errors of all models were low and the 95% confidence intervals narrow (with an upper 95% CI between 9.5 and 9.7 individuals). According to AICc, the most suitable model was Mh, the model allowing for different capture probabilities between individuals. This model had a coefficient of variation (CV) of 0.02, indicating very high precession. These results make us confident that there was likely no lynx present in the study area that was not detected by our cameras.
Estimation of population density
For reasons described above, density estimations using SCR models were performed using capture histories of only the nine adult individuals detected in the second study period. Density estimates ranged between 0.52 and 0.55 individuals per 100 km², dependent on the model (Table 3). The most appropriate model (according to AICc) was model M0, yet density estimations did not differ greatly between models (Table 3). All finite mixture models had AICc values substantially below model M0. In other words, the higher number of parameters used by these models did not provide a substantially better fit of the models. Population density of the most appropriate model (M0) was 0.52 individuals per 100 km². Owing to the higher complexity of SCR models (compared to conventional CR models), standard errors and 95% confidence intervals were larger in all models (Table 3). The CV of the most appropriate model was 0.34, indicating at best moderate precision.
Discussion
We used systematic camera trapping surveys to describe and estimate the number of lynx present in the last two years of a lynx reintroduction project in the Palatinate Forest in south-west Germany. To our knowledge, our study is the first to provide a systematic estimation of lynx abundance directly after the termination of a reintroduction project. The number of lynx present in the 1,000 km² study area in the centre of the approximately 1,800 km² Palatinate Forest declined from 15 independent individuals at the end of the first survey in spring 2020 to only 11 independent individuals at the end of the second survey in spring 2021. Given that 20 lynx had been translocated to the Palatinate Forest, and that several lynx reproduced successfully in their new environment, lynx abundance at the end of the project is surprisingly low. This finding, therefore, warrants a closer examination and explanation.
Even though the study area of 1,000 km² represents much of the total area of the Palatinate Forest (Fig. 1), there were at least two additional lynx (the females Rosa and Tarda) present outside the study area in the periphery of the forest (Idelberger, pers. communication, Stiftung Natur und Umwelt Rheinland-Pfalz 2023). Their home ranges extended into the fragmented forest areas west and north of the Palatinate Forest. Even though these individuals were not detected in our study area, they were part of the Palatinate lynx population, increasing the number of individuals at the end of the second survey to at least 13. In addition, we cannot exclude the possibility that there were even more individuals living in the fragmented forest areas outside the Palatinate Forest.
Moreover, there are vast areas of suitable lynx habitat south of the Palatinate Forest, where the Palatinate Forest continues into the low mountain range of the Vosges (France). Together with the Palatinate Forest, the Vosges offer an area of connected lynx habitat of approximately 8,000 km² (Fig. 1). The lynx translocated to the Palatinate Forest, therefore, had a very large area over which they could distribute themselves. In fact, opportunistic monitoring data from the Vosges showed, that until spring 2021 at least four additional individuals were present in the Vosges that originated from the Palatinate Forest, three of them being lynx translocated to the Palatinate Forest, one being an offspring born in the Palatinate Forest in spring 2018 (Morelle at al. 2021). These individuals are not lost from the Palatinate population, but form part of a transboundary Franco-German lynx population.
Because the Palatinate Forest represents the northern part of a large area of lynx habitat, it was to be expected that not all lynx would remain in the Palatinate Forest. The large home ranges of lynx (100–200 km² for females, 200–300 km² for males in central Europe, Herfindal et al. 2005) and their territorial behaviour sets a natural upper limit on the number of individuals that can live in an area. For example, it is possible that the four males detected regularly during the second survey (Alfi, Filou, Libre, Rumo) divided much of the 1,000 km² study area among themselves with little space available for additional males. The disappearance of three other males (Brano, Palu, Wrano) could thus be a result of a replacement through more dominant males. In fact, one of these males (Brano) was later detected in the Vosges (Morelle et al. 2021).
In light of the above discussion, the decline in lynx abundance over the course of this study, even to levels significantly lower than the number of translocated lynx, is not as alarming as it might seem at first glance. It might reflect the status of a still young population that can expand over a potentially large area.
Nevertheless, an estimated population density of 0.52 individuals per 100 km² can certainly be considered low and falls markedly below the levels of established reintroduced populations (Pesenti and Zimmermann 2013; Middelhoff and Anders 2018; Palmero et al. 2021). For example, using a similar computational approach as we did, Palmero et al. (2021) estimated lynx densities between 1.09 and 2.35 individuals per 100 km² for the Bohemian-Bavarian population in the years 2009–2018. In the Dinaric Mountains of Slovenia and Croatia densities are lower than in the Bohemian-Bavarian Forest, but still higher than in the Palatinate Forest (0.83 individuals per 100 km², Fležar et al. 2023). In the autochthonous population of the western Carpathians, lynx densities range between 0.59 and 1.86 individuals per 100 km² in three study sites in central areas of the western Carpathians (Kubala et al. 2017; Dulà et al. 2021). Interestingly, however, in two sites in more peripheral areas of the western Carpathians, densities between only 0.26 and 1.08 individuals per 100 km² were estimated (Kubala et al. 2017; Dulà et al. 2021). Likewise, in the French Jura Mountains, lynx densities are also comparatively low, ranging between 0.24 and 0.91 individuals per 100 km² in three study areas between 2012 and 2015 (Gimenez et al. 2019). These results are in line with a commonly observed trend that in many animal populations, abundance is lower in peripheral regions than in central areas of the distribution range (Martinez-Meyer et al. 2013, Pironon et al. 2017; see also Dulà et al. 2021). This trend may also explain why population density is currently still low in the Palatinate Forest (and may remain low in the future).
We believe there are at least two lessons learnt from this study: The first lesson is of general nature and emphasizes the need for a careful post-release monitoring. The low number of individuals detected at the end of the reintroduction project in the Palatinate Forest makes clear that any reintroduction project is not terminated in the moment the last individuals are released into the wild. Any reintroduction project should rather be followed by a tight monitoring program in which the status of the new population is assessed repeatedly, ideally over a period of several years post release.
The second lesson is more specific to the Vosges-Palatinate population: Given the potentially large distribution area, it is evident that a systematic population assessment should not be restricted to the northern part of this population. We recommend, therefore, that the transboundary cooperation initiated in the LIFE project should be continued and intensified for a coordinated monitoring of this important Franco-German lynx population. Given the currently low population density in the Palatinate Forest, a synchronized and harmonized, transboundary monitoring is now urgently required, ideally by extending the survey period into autumn (Weingarth et al. 2015) to enhance the chance to detect females with dependent offspring. Moreover, in the case that population density remains low, supportive measures need to be considered, such as the release of additional individuals to reinforce and stabilize the population. Such individuals need not necessarily be wild-caught individuals, but may also be carefully selected individuals from captive breeding programs (Lengger et al. 2021).
Notwithstanding the above recommendations, we need to stress two methodological issues. First, our survey was carried out during the mating season, and possible influxes of individuals (particularly males during so-called mating season excursions) could bias density estimates. However, all individuals that entered our analyses were known to have home ranges in the study area. We can thus be confident that no such influxes occurred. Moreover, most density estimations of Eurasian lynx have been carried out during the winter months (e.g. Kubala et al. 2017; Dulà et al. 2021), which facilitates comparison of our study with previous studies. Second, with a sample size of only nine individuals, our sample size falls just below the minimum sample size of ten individuals recommended to derive reliable density estimates (Palmero et al. 2023). In fact, the CV of the most appropriate model was 0.34, indicating at best moderate precision of the estimate, despite a high number of recaptures. Results of the density estimation should thus be interpreted with caution.
Conclusion
At the end of the lynx reintroduction project in the Palatinate Forest, only 11 independent individuals remained in the study area. Despite several successful reproductions, the number of individuals present was thus markedly lower than the number of individuals released. While the low number of lynx present can be explained by dispersal and spatial expansion of the population into the Vosges, population density in the Palatinate Forest was nevertheless low, and might not be sufficient to ensure the long-term viability of the population. We call for a systematic transboundary monitoring of the Vosges-Palatinate population and recommend that population reinforcement should be considered in case population density remains low.
Data availability
Data supporting the findings of this study are available within the article and its supplementary materials.
References
Boitani L, Linnell JDC (2015) Bringing large mammals back: large carnivores in Europe. In: Pereira HM, Navarro LM (eds) Rewilding European landscapes. Springer, Cham
Bonn Lynx Expert Group (2021) Recommendations for the conservation of the eurasian lynx in Western and Central Europe. CatNews Special Issue 14:78–86
Borchers DL, Efford M (2008) Spatially explicit maximum likelihood methods for capture–recapture studies. Biometrics 64:377–385
Breitenmoser U, Breitenmoser-Würsten C (2008) Der Luchs. Ein Großraubtier in Der Kulturlandschaft. Salm, Bern
Breitenmoser U, Krebühl J, Heider C, Breitenmoser-Wprsten C (2021) Challenges in the conservation oft he eurasian lynx in Continental Europe – an introduction. CatNews Special Issue 14:3–4
Chao A (2001) An overview of closed capture-recapture models. J Agric Biol Environ Stat 6:158–175
Chapron G, Kaczensky P, Linnell JDC (2014) Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 346:1517–1519
Choo YR, Kudavidanage EP, Amarasinghe TR, Nimalrathna T, Chua MA, Webb EL (2020) Best practices for reporting individual identification using camera trap photographs. Glob Ecol Conserv 24:e01294
Dulà M, Bojda., Chabanne DBH, Drengubiak P, Hrdý L, Prokešová J, Kubala J, Labuda J, Marčáková L, Oliveira T, Smolko P, Váňa M, Kutal M (2021) Multi–seasonal systematic camera–trapping reveals fluctuating densities and high turnover rates of Carpathian lynx on the western edge of its native range. Sci Rep 11:9236
Efford MJ (2023) Package secr. Spatially Explicit Capture-Recapture, https://cran.r-project.org/web/packages/secr/secr.pdf, Juli 2023
Fležar U, Aronsson M, Černe R, Pičulin A, Bartol M, Stergar M, Rot A, Hočevar L, Topličanec I, Sindičić M, Gomerčić T, Slijepčević V, Krofel M (2023) Using heterogeneous camera-trapping sites to obtain the first density estimates for the transboundary eurasian lynx (Lynx lynx) population in the Dinaric Mountains. Biodiv Conserv 32:3199–3216
Gimenez O, Gatti S, Duchamp C, Germain E, Laurent A, Zimmermann F, Marboutin E (2019) Spatial density estimates of eurasian lynx (Lynx lynx) in the French Jura and Vosges Mountains. Ecol Evol 9:11707–11715
Herfindal I, Linnell JDC, Odden J, Nilsen EB, Andersen R (2005) Prey density, environmental productivity and home-range size in the eurasian lynx (Lynx lynx). J Zool 265:63–71
Idelberger S, Krebühl J, Back M, Ohm J, Prüsinger A, Sandrini J, Huckschlag D (2021) Reintroduction of eurasian lynx in the Palatinate Forest, Germany. CatNews Special Issue 14:38–42
Krebühl J, Zimmermann F, Herdtfelder M, Idelberger S, Suchant R, Drouet-Holguet N, Breitenmoser-Würstern C, Breitenmoser U (2021) Transboundary cooperation in lynx conservation under the auspice of the Upper Rhine Conference. CatNews Special Issue 14: 55–56
Kubala J, Smolko P, Zimmermann F, Rigg R, Tam B, Foresti D, Breitenmoser-Würsten C, Kropil R, Breitenmoder U (2017) Robust monitoring of the eurasian lynx Lynx lynx in the Slovak carpathians reveals lower numbers than officially reported. Oryx 53:548–556
Lengger J, Breitenmoser U, Sliwa A (2021) EAZA breeding programmes as sources for lynx reintroductions. CatNews Special Issue 14:76–77
Linnell JDC, Breitenmoser U, Breitenmoser-Würsten C, Odden J, Von Arx M (2009) Recovery of Eurasian Lynx in Europe: what part has Reintroduction played? In: Hayward M, Somers MJ (eds) Reintroduction of top-order predators. Wiley-Blackwell, Chichester, UK, pp 72–91
Lucena-Perez M, Marmesat E, Kleinmann-Ruiz D, Martinez-Cruz B, Wecek K, Savaljev AP, Seryodkin IV, Okhlopkov I, Dvornikov MG, Ozolins J, Galsandorj N, Paunovic M, Ratkiewicz M, Schmidt K, Godoy JA (2020) Genomic patterns in the widespread eurasian lynx shaped by late quaternary climatic fluctuations and anthropogenic impacts. Mol Ecol 29:812–828
Martínez-Meyer E, Díaz-Porras D, Peterson AT, Yáňez-Arenas C (2013) Ecological niche structure and rangewide abundance patterns of species. Biol Lett 9:20120637
Middelhoff TL, Anders O (2018) Abundanz und Dichte des Luchses im östlichen Harz. Fotofallenmonitoring 2017, Projektbericht, Nationalpark Harz
Morelle S, Schwoerer ML, Kurtz C, Scheid C, Bouquier C (2021): Suivi et situation actuelle du Lynx dans le Massif des Vosges - Actions mises en oeuvre pour améliorer son acceptation. Proceedings of the final meeting of the project „Life Luchs Pfälzerwald, pp. 13–15
Mueller SA, Prost S, Anders O, Breitenmoser-Würsten C, Kleven O, Klinga P, Konec M, Kopatz A, Krojerová-Prokešová J, Middelhoff TL, Obexer-Ruff G, Reiners TE, Schmidt K, Sindičič M, Skrbinšek T, Tám B, Saveljev AP, Naranbaatar G, Nowak C (2022) Genome-wide diversity loss in reintroduced eurasian lynx populations urges immediate conservation management. Biol Conserv 266:109442
OpenStreetMap contributors (2024) Retrieved from https://download.geofabrik.de/europe/germany.html)
Otis DL, Burnham KP, White GC, Anderson DR (1978) Statistical inference from capture data on closed animal populations. Wildl Monogr 62:1–13
Palmero S, Belotti E, Bufka L, Gahbauer M, Heibl C, Premier J, Weingarth-Dachs K, Heurich M (2021) Demography of a eurasian lynx (Lynx lynx) population within a strictly protected area in Central Europe. Sci Rep 11:19868
Palmero S, Premier J, Kramer-Schadt S, Monterroso P, Heurich M (2023) Sampling variables and their thresholds for the precise estimation of wild felid population density with camera traps and spatial capture–recapture methods. Mammal Rev. https://doi.org/10.1111/mam.12320
Pesenti E, Zimmermann F (2013) Density estimations of the eurasian lynx (Lynx lynx) in the Swiss Alps. J Mammal 94:73–81
Pironon S, Papuga G, Villellas J, Angert AL, García MB, Thompson JD (2017) Geographic variation in genetic and demographic performance: new insights from an old biogeographical paradigm. Biol Rev 92:1877–1909
Pledger S (2000) Unified maximum likelihood estimates for closed capture-recapture models using mixture. Biometrics 56:434–442
Port M, Henkelmann A, Schröder F, Waltert M, Middelhoff TL, Anders O, Jokisch S (2021) Rise and fall of a eurasian lynx (Lynx lynx) stepping-stone population in central Germany. Mammal Res 66:45–55
R Core Team (2021) R: a language and environment for statistical computing. R foundation for statistical Computing, Vienna, Austria. https://www.R-project.org/.
Rheinland-Pfalz Kompetenzzentrum für Klimawandelfolgen (2023) www.klimawandel.rlp.de, accessed 2023/11/14.
Rivest L, Baillargeon S (2015) Rcapture. Loglinear Models for Capture-Recapture Experiments
Royle JA, Chandler RB, Sollmann R, Gardner B (2013) Spatial capture-recapture. Academic
Stiftung Natur und Umwelt Rheinland-Pfalz (2021) LIFE Luchs Pfälzerwald. Projektbericht 2015–2021
Stiftung Natur und Umwelt Rheinland-Pfalz (2023) www.snu.rlp.de/projekte/luchs/wiederansiedlung/raumnutzung. Accessed 14 Nov 2023
Steyer K, Kraus RH, Mölich T et al (2016) Large-scale genetic census of an elusive Carnivore, the European wildcat (Felis s. silvestris). Conserv Genet 17:1183–1199
Wagenmakers EJ, Farrell S (2004) AIC model selection using Akaike weights. Psych Bull Rev 11:192–196
Weingarth K, Zeppenfeld T, Heibl C, Heurich M, Bufka L, Daniszová K, Müller J (2015) Hide and seek: extended camera-trap session lengths and autumn provide best parameters for estimating lynx densities in mountainous areas. Biodiv Conserv 24:2935–2952
Zimmermann F, Foresti D (2016) Capture-recaptre methods for density estimation. In: Rovero F, Zimmermann F (eds) Camera trapping for wildlife research. Pelagic Publisihng, Exeter, UK
Zimmermann F, Breitenmoser-Würsten C, Breitenmoser U (2007) Importance of dispersal for the expansion of a eurasian lynx Lynx lynx population in a fragmented landscape. Oryx 41:358–368
Acknowledgements
We would like to thank the Foundation Nature and Environment (SNU) of Rhineland-Palatinate, especially Sylvia Idelberger, for their support during our study. We thank Michael Back, Sarah Krämer, Raphael Reischmann and Hermann Schmalenberger for fieldwork support. Furthermore, we are indebted to the forestry departments as well as the hunting community of the Palatinate Forest for their helpful cooperation throughout the study.
Funding
Open Access funding enabled and organized by Projekt DEAL. The project was financially supported by the Ministerium für Klimaschutz, Umwelt, Energie und Mobilität Rheinland-Pfalz (MKUEM RLP).
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
The study was designed by UH and MP. Data collection and analysis were performed by MP and CT. Maps were created by CT. The first draft of the manuscript was written by MP and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethical approval
Not applicable. All research carried out in this study involved non-invasive fieldwork for which ethical approval was not required.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Port, M., Tröger, C. & Hohmann, U. Status assessment of a recently reintroduced eurasian lynx (Lynx lynx) population in the Palatinate Forest, South-West Germany. Eur J Wildl Res 70, 55 (2024). https://doi.org/10.1007/s10344-024-01800-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-024-01800-8