
Abstract
In northern Norway, an increasing population of Greylag Geese (Anser anser) forages considerably on dairy grassland and can eat up to 60% of the grass (dry matter mass) on a field if allowed to eat undisturbed throughout the growing season. In this study, the seasonal foraging behavior of Greylag Geese on diary grassland was continuously monitored with game cameras from late April to the end of August to be able to pinpoint effective preventive measures to manage, control, and prevent this crop damage. Limited, but regular, lethal scaring was conducted on some fields to reveal the preventive effect of this measure. Foraging from Greylag Geese in a rangeland area was also monitored, and a complete dataset of seasonal foraging behavior of this species is presented here. Greylag Geese foraging on the fields reaches a top between 04:00 and 08:00 h am, all season. Energy and digestibility of the field grass (timothy) did not reveal any correlation with grazing patterns. Greylag Geese do not visit the fields during molting; however, they may visit fields with their chicks to forage. Lethal scaring completely removes visits from Greylag Geese on the fields where this is conducted, while foraging continues if geese are given undisturbed access. In the rangeland area foraging seems to be even and continuous throughout the season, but significantly lower. In the end of June and late July/early August, there is a peak in visits and number of geese per visit on the fields. Preventive and effective measures against crop damage from Greylag Geese must therefore at least be initiated during late June and early August, and between 04:00 and 08:00 am.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In northern Norway, farm fields are mainly used for growing grass as feed for the livestock during the winter months. This feed is vital for the farms economy and is the best feed the husbandry can eat during the winter. Both energy and digestibility as well as function of the ruminant digestive system are best maintained with natural grown grass (e.g., Suárez et al. 2007). Farms situated in areas in northern Norway where the populations of Greylag Geese (Anser anser) are large and increasing, have, from approximately the start of year 2000, experienced increasing loss of grass meant for livestock to grazing Greylag Geese (Aarseth et al. 2018; Aarseth 2019, personal and public communication). The populations of Greylag Geese and other geese species in Europe, which partly nests in or migrate through northern Norway, have increased substantially for many years (Fox et al. 2010; Powolny et al. 2018). The central European breeding population of Greylag Geese also migrate and arrive at the nesting grounds earlier each year (Podhrázský et al. 2017). Also, it seems that the geese have shifted their foraging grounds from mainly natural rangeland to agricultural areas (Fox et al. 2005; Fox et al. 2017; Fox and Abraham 2017; Gauthier et al. 2005). Together, this exerts an increasing loss of the harvest from grass fields and other crops in many areas (e.g., Ankney 1996; Bjerke et al. 2014). In northern Norway there is abundant feed in the natural rangelands around the fields of most farms, especially during the summer months. Nonetheless, Greylag Geese seem to prefer grazing on the field grass. This could be because the fields offer abundance of feed which is concentrated and easily accessible. It could also be that the Greylag Geese choose to graze more intensively during growth periods with higher energy content and digestibility of the field grass. Timothy (Phleum pratense), which is grown on the fields in this study show seasonal variation in, e.g., energy content and digestibility (Gustavsson and Martinsson 2004).
Greylag Geese cause field damage by feeding on grass and seeds, trampling, and faecally contaminating the grass. According to some farmers, the geese also spread seeds from unwanted plants and weeds. Recent studies have shown that the Greylag Geese can leave up to 100 kg of droppings (wet weight) per 100 m2 (Aarseth et al. 2018) and eat between 35 and 60% of the grass (dry matter mass) if left undisturbed to forage (Aarseth et al. 2018; Aarseth 2019). Studies on other geese species such as Pink-footed Geese (Anser brachyrhynchus), Canada Geese (Branta canadensis), Giant Canada Geese (Branta canadensis maxima), and Greater Snow Geese (Chen caerulescens atlantica) have shown the same proportions of crop damage and revealed this to be a challenge for farmers on several continents (e.g., Ankney 1996; Win 2001; Bjerke et al. 2014). To prevent damage to the field grass, farmers have deployed several methods in northern Norway. In most areas, the farms are situated close to natural rangeland areas on the coast and therefore have many other bird species nesting in the proximity of the fields. The use of sound guns, laser, and dogs are therefore not ethically sound and will affect many other species than Greylag Geese. Also, farms in northern Norway have limited areas available for grass fields and use of diversionary fields, i.e., non-production fields where geese are allowed to graze (Conover 2002, Fox et al. 2017), is not possible most places. Therefore, in these areas, lethal scaring has been conducted for the most part. Lethal scaring has proven very effective to prevent a variety of crop damage caused by geese (Månsson 2017; Aarseth et al. 2018; Aarseth 2019). Studies have indicated that the most important part is to conduct lethal scaring on a regular basis, not to shoot a lot of geese (Månsson 2017; Aarseth et al. 2018; Aarseth 2019). When lethal scaring is conducted regularly, after some time, the Greylag Geese seem to perceive the whole area (fields) as hostile and choose to forage elsewhere (Månsson 2017; Aarseth et al. 2018; Aarseth 2019).
Among farmers, there is a lot of knowledge on geese foraging behavior as they observe it daily. However, hunters report that geese frequently visit the fields during dawn and early morning which will not be detected by farmers unless they look for fresh droppings (personal observation and communication). To be able to pinpoint any preventive measures it is important to know the diurnal and seasonal foraging behavior of the Greylag Geese throughout an entire growing season, both on the fields and in the rangeland surrounding the fields. If a consistent grazing pattern is revealed it will help preventive measures to be timely initiated so that they are more effective. In this study, a complete surveillance of Greylag Geese field and rangeland foraging behavior on Musvær (Troms and Finnmark County, Norway) throughout an entire growing season was therefore conducted, by use of game cameras. Also, lethal scaring was conducted on specific fields in combination with this surveillance to get a clear-cut result of the effect of this preventive measure. As the chemical content, energy content and digestibility of the field grass (timothy) are shown to vary through the growing season (Gustavsson and Martinsson 2004), timothy from all fields of Musvær was analyzed throughout the growing season. This was done to look for any correlation between grazing by Greylag Geese and periods of high and/or low energy content and digestibility of the timothy. That is, a positive correlation between high energy content of the grass and high grazing pressure would support selective grazing time preferences by geese. No correlation (or a negative) would support that the abundance of grass (food) on the fields is the preference for the geese to graze there.

A Aerial photo of field 3 and placement of game cameras (green squares). Approximate direction of horizontal view of field from the camera lenses is indicated by light triangles. B Game camera attached to a wooden pole and with plastic cover to protect the lens from moist, rain and snow, field 3

Goosehours on field 1 (A), field 2 (B) and field 3 (C) during the growing season on Musvær. On all these fields lethal scaring was conducted every 3rd–7th day starting at 25th of April, except of July. Start/re-start of lethal scaring is indicated with a black arrow. In July hunting was not conducted

Greylag Geese (n = 73) leaving field 5 at 10:49 am in the beginning of August

Goosehours on field 4 (A) and field 5 (B) during the growing season on Musvær. Lethal scaring was not conducted on these fields

Goosehours in the selected rangeland area from 25th of April to 10th of August. Observations here were cut off 2 weeks before the end of the project (and on the fields) due to free grazing goats that managed to get past the electric wires and destroy the cameras. Lethal scaring was not conducted here

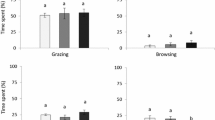
Percent of total visits during the day related to time of day (divided in six 4-h intervals) for all fields (A) and the rangeland area (B) the entire growing season on Musvær. Fields (A): each symbol represents the 5 mean monthly percentages of visits during the respective time interval for all 5 fields (n = 25) the entire season (April-August) ± SEM. Rangeland (B): each symbol represents the 5 mean monthly percentages of visits during the respective time interval (n = 5) the entire season (April–August) ± SEM. Values that do not share the same letter are significantly different (P < 0.05)

Number of chicks observed on field 5 during the growing season on Musvær. There were only observed grazing chicks with parents on this field. All ages of chicks were counted as a chick (see Fig. 8)

Grazing Greylag Geese with chicks on field 5 in late June. In this picture 8 chicks can be observed, the youngest in the background

Feed unit milk (FUM) per kg dry matter of timothy from all fields of Musvær collected every month during the growing season. Each value is the mean of n = 15 ± SEM. Values that do not share the same letter are significantly different (P < 0.05)

Crude protein (▼), digestibility (⬤), non-digestible fiber (▽) and carbohydrates (◯) in % of dry matter in timothy from all fields of Musvær collected every month during the growing season. Each value is the mean of 3 sites per field (n = 15 ± SEM). Values that do not share the same letter are significantly different (P < 0.05)

A family of Greylag Geese escaping from an incoming white-tailed eagle on field 5 at 06:07 am in the middle of June
Material and methods
Study site and instrumentation of sites
The project was conducted from 25th of April to 26th of August 2020. Nineteen game cameras (Browning, model BTC-BE) were mounted around five fields and in one rangeland area of Musvær island, Tromsø municipality, Norway. The size and topography of the fields determined the number of game cameras that were needed to achieve complete surveillance (Table 1). All cameras were mounted on 1.3 m high poles on the edges of the fields to avoid coverage by growing plants during the growing season and placed inside plastic containers to be protected from moist, rain and snow (Fig. 1B). The plastic containers did not disturb the field of view of the lens. In the rangeland area, cameras were protected from free grazing goats by an electric fence powered by a battery.
On each field, the cameras were placed in such a way that their horizontal field of view (36°) did not overlap and together covering the entire field (Fig. 1A). This was achieved by mounting 3–4 cameras. Two cameras were placed in a rangeland area selected by the locals, where Greylag Geese traditionally are known to prefer to eat and rest. The cameras were set on timelapse mode, taking one picture per hour each day (24 h) for the entire project period. In addition, the cameras took pictures when moving objects triggered the sensors up to approximately 30 m, also during periods of twilight and darkness (IR-sensors). This allowed monitoring of Greylag Geese activity on the fields in relation to other birds, animals, and humans.
Every 2–3 weeks, the memory cards of the cameras were collected and replaced. Then the pictures were analyzed and counted for the number of geese present on the fields. This allowed for a regular check of the function of the cameras and a time dispersed counting of geese on the pictures, which was done manually. For the whole project period the number of pictures was about 54,000, needing about 20 workdays of counting. All cameras functioned perfectly during the entire project period. It was necessary to change the batteries of the cameras twice during the project period. Rarely, and not for more than 1–3 h at a time, a camera lens was covered by snow or moist, resulting in non-countable pictures, but this was only the case for one camera at the time for each field.
Calculations of visits and numbers of geese
To estimate the total number of Greylag Geese that were present on a field or in the rangeland area at a specific time and date, all pictures from all cameras were first analyzed, i.e., geese were counted, and all data were entered in Excel (Microsoft 365). All Greylag Geese numbers from all cameras at a field were aligned and summarized, and the total number of Greylag Geese present every hour each day on that field was thereby calculated. The same was done for the rangeland area. In such a way, the total number of Greylag Geese present on the fields and in the rangeland area every hour through the entire growing season was estimated.
More geese present on a field at a certain time indicate a higher grazing pressure, although they also rest on the fields. To estimate the relative grazing pressure from geese throughout the growing season at Musvær, the total number of geese per day was multiplied by the proportion of hours the geese were present, named “goosehours.” It is also possible to calculate “goosedays” in a similar way (Vang 2014), but the extensive material in this study allows for a higher resolution. “Goosehours” is calculated by the following simple equation:
where 24 = total number of pictures taken during a day
*In periods with darkness during the day (April/early May and late August) the camera will perceive this as night and not take timelapse-pictures. On such days, 24 in the calculation above was replaced with, e.g., 22. Musvær is situated at 69°52′N and during the project period the sun, partly or whole, is permanently above the horizon from 18th of May to 25th of July. In the period from 25th of April to 18th of May the twilight decreases from 3 h 48 min to 0 h 0 min a day, and from 25th of July to 26th of August it increases from 0 h 0 min to 7 h 45 min a day.
For example, a field has a total visit of 600 Greylag Geese one day and the geese are present on 12 of the 24 pictures. The number of “goosehours” that day is therefore 300. “Goosehours” is not an absolute number, but a good relative expression of the grazing pressure from geese.
Visualizing the timing of Greylag Geese visits to the fields and rangeland area
To present the presence and relative grazing pressure from Greylag Geese for the entire growing season (project period), all goosehours for all days (n = 124) for one field were plotted against date to create area plots in Sigmaplot 13.0 (Systat Software, Inc. SigmaPlot for Windows). This was done for all fields and for the rangeland site.
To calculate and present the time-of-day visits from Greylag Geese, the day was divided into 6 × 4 h intervals (12:00 am–04:00 am, 04:00 am–08:00 am, 08:00 am–12:00 am, 12:00 pm–04:00 pm, 04:00 pm–08:00 pm, and 08:00 pm–12:00 pm), and all Greylag Geese present on the pictures in these periods were registered. The 4 h interval was selected based on observations in previous projects on Musvær that Greylag Geese tend to graze for about a maximum of 4–5 h if left undisturbed. Then all these interval numbers were summarized from all 5 fields over the project period which lasted 5 months (April–August, n = 25). The percentage of visits during each time interval for one day was calculated and averaged altogether from all days and fields (monthly) over the entire growing season.
Lethal scaring
Lethal scaring has been conducted on Musvær for several years. In 2020, 25 Greylag Geese were allowed to be shot from 25th of April to 30th of June and 10 Greylag Geese from 1st of August to 15th of August (Grant nr. 20/06950–2 from Tromsø municipality). Regular hunting starts on 15th of August. Lethal scaring was conducted from hunting sheds located nearby the fields, and hunters entered the sheds in the evening and observed the fields during the night. When Greylag Geese landed to graze, the hunter shot one goose using a rifle with a silencer and exited the shed immediately to pick it up. This scared all geese off the field, and the hunter made sure he/she was observed by the escaping geese. During the project period, 25 of the granted 35 Greylag Geese were shot (Table 1).
As part of the project and to avoid preferred selection by farmers, lethal scaring was conducted on three randomly selected fields on Musvær every 3rd to 7th day, from 25th of April to 30th of June and from 1st of August to 26th of August. In July, the Greylag Geese molt their flying feathers and does not visit the fields to forage (local knowledge and personal observation). Two fields were left undisturbed (no lethal scaring) except for the normal traffic related to farm work and activity from people (Table 1).
Harvest and analyses of timothy
Timothy (Phleum pratense) was harvested in the middle of every month during the project period, except for April, when samples were collected at the start of the project, 25th of April. Samples (n = 3) were taken randomly from each field, put in zip lock plastic bags and frozen at − 20 °C for later analyses. After drying the samples in paper bags at + 67 °C for 48 h, the timothy was grinded to a fine powder according to a standard procedure (Fystro and Lunnan 2006). All samples were then analyzed (in vitro) for chemical composition, feed unit milk, energy content, fiber content, non-digestible fiber, digestibility, protein and carbohydrates following the procedure of Fystro and Lunnan (2006).
Statistics
No statistics were employed regarding the counting of geese present on the pictures (ending up in goosehours) because these numbers are “absolute.” On the data for the “% of total visits vs. time of day (Fig. 6)” and “chemical analyses of the timothy vs. month (Figs. 9 and 10)” a one-way ANOVA, including a post hoc Tukey HSD test was used. The significance level was set to P < 0.05.
Results
The values of goosehours on the 5 fields of Musvær differed before and after the molt in July (Figs. 2 and 4). Field 2 had values above 300 at the end of April, while field 3 reached a seasonal maximum of about 50 at the same time. In this period field 1, 4, and 5 reached around 100 goosehours, or slightly above. In the rangeland area values were drastically lower before July, reaching about 18 at the end of May, but then reached a maximum of 113 at the end of July (Fig. 5). The onset of lethal scaring on field 1, 2, and 3 reduced or removed goosehours almost completely (Fig. 2). About 2 weeks after initiation of lethal scaring no Greylag Geese were observed on field 1, and only a few on field 2 and 3 (10–20 goosehours). On field 2, goosehours were reduced significantly (P < 0.05) from an average of 56 ± 27 (n = 12, ± SEM, 26th April to 7th May) to 0.47 ± 0.20 (n = 53, ± SEM, 8th May to 30th June). On field 4 and 5, without lethal scaring, the situation was completely different (Fig. 4). Especially field 5 had regular visits of many Greylag Geese almost every day until the molt in July, reaching a maximum of about 230 goosehours and an average value of 38 ± 6.38 (n = 65, ± SEM). In the rangeland area, until July, goosehours were on average 1.7 ± 0.55 (n = 64, ± SEM, Fig. 5) and maximum level reached was 18. However, in late July/beginning of August, goosehours were on average 22 ± 7.0 (n = 16, ± SEM, Fig. 5) and maximum level reached was 113. Greylag Geese did not visit the rangeland area in the majority of July. In most of July, as expected because of the molt, almost no Greylag Geese were observed on the fields (Figs. 2 and 4). Interestingly, Greylag Geese with chicks in all ages started to visit field 5 to graze already in the beginning of June, and this continued, although reduced, in July (see Figs. 7 and 8). At the end of July/beginning of August, goosehours reach maximum levels. Fields 4 and 5 (Figs. 3 and 4) reached about 1000 and 400 goosehours, respectively, in August. In the fields where lethal scaring was conducted, only field 1 had an abrupt increase in goosehours at the beginning of August. Again, as in April, goosehours were completely removed with the onset of lethal scaring (Fig. 2).
Season overall, Greylag Geese visit the fields at all hours during the day. However, they seem to prefer to graze on the fields during dawn and early morning (Fig. 6A), as about 28% of all visits, all season, on all fields, occur between 04:00 and 08:00 am. Visits during this time interval was significantly higher than other time interval visits (P < 0.05, n = 25). In the rangeland area the grazing showed a parabolic spread throughout the day (Fig. 6B) with most visits occurring during midday/early evening. Over 80% of the visits occur between 08:00 am and 20:00 pm.
The field grass of Musvær, timothy (Phleum pratense), collected from all fields in the middle of each month (except for April, when it was collected in the end of the month), was analyzed for chemical content and digestibility (Figs. 9 and 10). Feed unit milk (per kg dry matter) increased significantly from April to May (P < 0.05) but did not significantly change the rest of the growing season. Digestibility of the timothy showed the same pattern (in % of dry matter), while non-digestible fiber decreased significantly (P < 0.05) from April to May and did not significantly vary the rest of the season (% of dry matter). Crude protein showed a significant increase only from April to May (P < 0.05) but decreased again to April levels later in the growing season. The % of carbohydrates of dry matter was significantly higher in June and August compared to April (P < 0.05), although the increase was minor.
Discussion
The foraging behavior of Greylag Geese on the fields of Musvær follows a pattern where grazing pressure is maximized, both through number of visits and numbers of geese per visit, in late June and early August. The duration of individual visits could not be verified since no identification of individual Greylag Geese was done in this study. But the pictures show when geese are present and how long they stay (± 1 h). The increasing pressure (goosehours) on the fields in late June is probably due to an increased energy demand preparing for the molt in June/July (Kahlert 2006). Studies have shown that it is imperative for Greylag Geese to tank up energy reserves, especially fat, prior to the molt (Loonen et al. 1991; Fox and Kahlert 2005). On Musvær, the molt seems to reach the flightless stage mainly in July, as many geese can still fly in late June. This is supported as almost no Greylag Geese were observed on the fields or in the rangeland area in most of July (Figs. 2, 4, and 5). This is a bit later than reported in studies on Greylag Geese breeding further south in Europe (e.g., Fox and Kahlert 2005), and it is not known if this could be a delay caused by higher latitudes. In early August increased grazing pressure (goosehours) is probably a result of two factors, increased number of Greylag Geese after the breeding and increased need for energy in preparation for the migration (McKechnie et al. 2015; Wascher et al. 2018). On Musvær, the breeding population of Greylag Geese leaves the islands around the end of August, and only visits from other southward migrating Greylag Geese occur in September (local knowledge, personal observation). Grazing in the rangeland area is stable through April, May, and June and suggests that geese graze here whenever they feel like it. In most of July there are few or no geese observed both on the fields and in the rangeland area. During the molt in July on Musvær Greylag Geese tend to be very shy and stay closer to the sea, as also indicated in other studies (e.g. Kahlert 2003). In late July and beginning of August goosehours in the rangeland area show an abrupt increase (Fig. 5), reaching 113 and average 22 per day. This is probably due to an increase in the population after the breeding and a need for energy before the migration. However, goosehours in the rangeland area are on average much lower than on the fields where lethal scaring is not conducted (Fig. 4), and it can be speculated that the rangeland area is used mostly for resting and some grazing by limited numbers of geese at a time. Thus, the fields seem to be used for intense grazing only and the preferred habitat for feeding.
Interestingly, Greylag Geese take their chicks up on the fields to graze from very early age (Fig. 8). It therefore seems that the energy and nutritional value of the grass (timothy) is accepted by the parents as energy-rich food for the chicks almost as soon as they hatch. This is supported by a recent study by Strong et al. (2021) which showed that Greylag Geese prefer grass pastures if they can choose between various crops. They also preferred this pasture during pre- and post-molt which indicates that the energy and digestibility of grass can meet the energy requirements of Greylag Geese in energy demanding periods like the molt (Strong et al. 2021). Another indication of the desirability of the timothy on Musvær is pictures that show white-tailed eagles (Haliaeetus albicilla) hunting for chicks and geese on field 5. So, geese still risk eating on the open field where they are more easily detected than in the rangeland areas (Fig. 11). In this study, however, a substantial proportion of grazing in rangeland areas of Musvær cannot be excluded, as no surveillance was done along the shoreline, where most chicks and parents are observed during geese counts (late June and early August) on Musvær. One assumption to be made is that since the pictures reveal that greylag chicks are introduced to grazing on the fields very early, they will therefore, most likely, continue to do so later in life. In July, only Greylag Geese with chicks were observed grazing on fields, and exclusively on field 5. There could be several explanations for this; this field has easy and short access to the sea, is “more protected” by small hills and humps, is situated furthest away from the farm and the activity there, and finally and most importantly, no lethal scaring was conducted here. This variation could also be present in the rangeland, including shoreline, of Musvær, as only one rangeland site was monitored in this study.
Lethal scaring is shown in this study to be very effective to prevent crop damage by Greylag Geese (Fig. 2). There are not many studies available that have looked at the effect of lethal scaring on geese, but the few that have been conducted show a clear-cut preventive effect of this management tool (Månsson 2017; Aarseth et al. 2018). This study is unique as the game cameras give a 24/7 surveillance of the fields through an entire growing season. In other studies, the surveillance of field grazing by geese has either been more limited or patched (e.g., Win 2001; Månsson 2017; Strong et al. 2021). In this study what you see is what you get, and the 54,000 pictures analyzed proves a clear-cut preventive effect by use of lethal scaring on grazing activity of Greylag Geese on the fields of Musvær. As soon as the lethal scaring is initiated, the number of goosehours decline, and for the most part, disappear completely (Fig. 2). A good preventive effect of lethal scaring has been shown in other studies as well (e.g., Månsson 2017; Aarseth et al. 2018; Aarseth 2019) and geese reduce the use of sites affected by hunting disturbance (Madsen and Fox 1995). On Musvær it has been necessary to conduct lethal scaring every year, probably to “remind” the older Greylag Geese and “educate” yearlings and Greylag Geese that do not breed on Musvær. By doing so, extensive harvest losses are avoided. If hunting is conducted every 3rd–7th day on a field, this protective measure can last an entire growing season on Musvær. When lethal scaring is not conducted, Greylag Geese eventually start grazing on the fields again, as after the period of no scaring in July (Fig. 2). Again, initiation of lethal scaring immediately decreases the number of Greylag Geese on the fields (Fig. 2A). By shooting only one goose per visit, the number of geese that needs to be shot to achieve protective success is low, and this study has shown that the 25 Greylag Geese that were shot was enough to keep the geese away from the fields. Also, the post-breeding population of Greylag Geese on Musvær have been counted every August since 2018 and shows stable numbers of about 250 individuals. Lethal scaring conducted in this way does therefore not seem to affect the population size on Musvær, and should not be regarded as derogation shooting. It seems that lethal scaring establishes an area as dangerous for the geese that is hunted, in this case Greylag Geese. The pictures in this study show that other species of geese grazing on the fields, as barnacle geese (Branta leucopsis) and pink-footed geese (Anser brachyrhynchus), which is not hunted, does not interrupt their grazing pattern as the Greylag Geese do. They leave the fields when a shot is fired, but not always, and return shortly thereafter. It is therefore tempting to speculate that geese register if they are hunted or not.
Several scaring methods have been and can be employed to prevent grazing damage from geese (e.g., dogs, sound, fences, diversionary fields, scarecrows (Smith et al. 1999) or laser (Clausen et al. 2019)). On Musvær, where up to 40 different bird species nest, most of these methods are either ethically not sound or too disturbing or time-consuming for the farmers to employ. A recent study on the use of laser beams to scare off geese (Clausen et al. 2019), shows that the laser is effective up to a certain distance, but too time-consuming to be used by the farmers. The cost–benefit analysis is not positive (Clausen et al. 2019), and farmers therefore need to employ personnel to scare geese off. Farms in northern Norway are generally small with small fields and do not necessarily have diversionary fields where the geese can graze freely. By use of lethal scaring, farmers can get help from hunters that want to hunt for geese, thereby saving time and cost. On Musvær, the farmers have successfully received help from voluntary experienced hunters for many years to prevent extensive grazing damage to the fields. Lethal scaring of Greylag Geese is the only method that does not disturb and affect other species. Taking into consideration the increasing European populations of Greylag Geese (Fox et al. 2010; Powolny et al. 2018), the stable numbers of Greylag Geese on Musvær and the low annual number shot here, lethal scaring has sustainably solved the problem of crop damage by Greylag Geese on this farm.
To be able to prevent grazing damage from geese it is helpful to know what time of day the geese prefer to graze on the field. One hundred Greylag Geese grazing for several hours during the night, can do severe damage before the farmers start their workday. On Musvær, season overall, the Greylag Geese visit the field preferably during dawn/early morning, and mostly between 04:00 and 08:00 am (Fig. 6A). To my knowledge, no other studies have investigated daily time preferences of visits from geese throughout an entire growing season. To prevent grazing damage, this means that the farmers should find solutions to scare the geese off at night/dawn. Automatic scaring devices could be employed, but as mentioned earlier, either the effect is too short (Smith et al. 1999) or ethically questionable. Why the Greylag Geese prefer to graze on the fields during the night/dawn is not evident, but it could be to take advantage of the lower activity of both the farm and diurnal predators (white-tailed eagles) on Musvær. This notion is supported by the grazing pattern in the rangeland area (Fig. 6B). This area is situated far from the farm and has no traffic from farmers, visitors, or hunters. The vegetation here shows great variety and abundance and should be sufficient to meet the energy requirements of Greylag Geese throughout their stay on Musvær. Based on goosehours and the daily grazing patterns it seems that the Greylag Geese stay here during the day, leading to a notion that this is to avoid disturbance and predators (it is very difficult to spot a Greylag Goose here as opposed to the field), and to rest and graze. However, it should be noted that the present study only monitored one rangeland area, and although rangeland areas of Musvær are mainly homogenous, other conditions at other sites might modify this notion.
Chemical analyzes of the timothy on Musvær including potential important variables for grazing geese like energy content, protein content and digestibility, did not vary between the fields and did not correlate with field grazing behavior of Greylag Geese (Figs. 9 and 10). Although both feed unit milk, digestibility, crude protein and carbohydrate content increased significantly from April to May, these values stayed constant from May and throughout the growing season. This does therefore not explain the intense grazing initiated already in April, which showed later peaks in late June and early August. From April to May a significant decrease in non-digestible fiber would also, if important for the geese, effect the grazing activity in April and May. But as Fig. 4 shows, on the fields where no lethal scaring was conducted, grazing is steadily above about 100 goosehours in both April and May. This leads to a sound hypothesis that Greylag Geese do not care particularly about energy content and digestibility of field grass (i.e., timothy) as the feed is so abundant that they can eat their daily requirements almost without walking.
When studying 54,000 pictures from grazing activity of geese, interesting and puzzling biological observations can be made. For instance, it appeared that the Greylag Geese knew very well if it were locals (farmers and family) or foreigners (i.e., researcher, hunters or visits) that were coming. In both cases, they leave the field but return about 3 times faster when locals had “scared” them off. This behaviour have been observed visually by locals before.
References
Aarseth JJ et al (2018) Effekten av skadefelling av grågås (Anser anser) for grovfôr-produksjonen på et nordnorsk gårdsbruk. NIBIO-rapport 154(4):29. https://nibio.brage.unit.no/nibio-xmlui/bitstream/handle/11250/2577626/NIBIO_RAPPORT2018_4_154.pdf?sequence=1&isAllowed=y
Aarseth JJ (2019) Grågåsa er sulten døgnet rundt - og liker aller best å spise av husdyras matfat. NIBIO POP 5(36):1–6. https://nibio.brage.unit.no/nibio-xmlui/handle/11250/2631383?locale-attribute=en
Ankney CD (1996) An embarrassment of riches: too many geese. J Wildlife Manage 60:217–223. https://doi.org/10.2307/3802219
Bjerke JW et al (2014) Reduced dairy grassland yields in Central Norway after a single springtime grazing event by pink-footed geese. Grass Forage Sci 69:129–139. https://doi.org/10.1111/gfs.12045/abstract.
Clausen KK et al (2019) Effectiveness of lasers to reduce goose grazing on agricultural grassland. Wildl Biol 1:1–8. https://doi.org/10.2981/wlb.00560
Conover M (2002) Resolving human-wildlife conflicts: the science of wildlife damage management. CRC-press, Boca Raton, Florida
Fox AD, Kahlert J (2005) Changes in body mass and organ size during wing moult in non-breeding Greylag Geese Anser anser. J Avian Biol 35(6):538–548. https://jstor.org/stable/3677786
Fox AD et al (2005) Effects of agricultural change on abundance, fitness components and distribution of two arctic-nesting goose populations. Global Change Biol 11:881–893. https://doi.org/10.1111/j.1365-2486.2005.00941.x
Fox AD et al (2010) Current estimates of goose population sizes in western Europe, a gap analysis and an assessment of trends. Ornis Svecica 20:115–127
Fox AD, Abraham KF (2017) Why geese benefit from the transition from natural vegetation to agriculture. Ambio 46:188–197. https://doi.org/10.1007/s13280-016-0879-1
Fox AD et al (2017) Agriculture and herbivorous waterfowl: a review of the scientific basis for improved management. Biol Rev Camb Philos. https://doi.org/10.1111/brv.12258
Fystro G, Lunnan T (2006) Analyser av grovfôrkvalitet på NIRS. Bioforsk FOKUS Vol. 1. Nr 3:180–181
Gauthier G et al (2005) Interactions between land use, habitat use, and population increase in greater snow geese: what are the consequences for natural wetlands? Global Change Biol 11:856–868
Gustavsson A-M, Martinsson K (2004) Seasonal variation of biochemical composition of cell walls, digestibility, morphology, growth and phenology in timothy. Eur J Agron 20:293–312
Kahlert J (2003) The constraints on habitat use in wing-moulting Greylag Geese Anser anser caused by anti-predator displacements. Ibis 145:E45–E52
Kahlert J (2006) Effects of feeding patterns on body mass loss in moulting Greylag Geese Anser anser. Bird Study 53:20–31. https://doi.org/10.1080/00063650609461412
Loonen MJJE et al (1991) Timing of wing moult in Greylag Geese Anser anser in relation to the availability of their food plants. Ardea 79:253–259
Madsen J, Fox AD (1995) Impacts of hunting disturbance on waterbirds – a review. Wildl Biol 1:193–206
McKechnie AE et al (2015) Global patterns of seasonal acclimatization in avian resting metabolic rates. J Ornithol 156:367–376
Månsson J (2017) Lethal scaring – behavioral and short-term numerical response of greylag goose Anser anser. Crop Prot 96:258–264
Podhrázský M et al (2017) Central European Greylag Geese Anser anser show a shortening of migration distance and earlier spring arrival over 60 years. Ibis 159:352–365
Powolny T et al (2018) AEWA International Single Species Management Plan for the Greylag Goose (Anser anser) - Northwest/Southwest European population, draft. AEWA Technical Series No. 71, Bonn, Germany
Smith AE et al (1999) Managing Canada geese in urban environments. Jack Berryman Institute Publication 16, and Cornell University Cooperative Extension, Ithaca, N.Y
Strong EA et al (2021) Seeking greener pastures: crop selection by Greylag Geese (Anser anser) during the moulting season. Ornis Fennica 98:1–17
Suárez BJ et al (2007) Effect of roughage Source and roughage to concentrate ratioon animal performance and rumen development in Veal calves. J Dairy Sci 90(5):2390–2403
Vang K (2014) Forvaltningsplan for kortnebbgås, hvitkinngås og grågås i Alstahaug Dønna Herøy Vega Kommuner 2014–2017. http://www.alstahaug.kommune.no/cpclass/run/cpesa62/file.php/h_def/14015451d14015453o2007bf/forvaltningsplan-gas-2014.pdf
Wascher CAF et al (2018) Free-living Greylag Geese adjust their heart rates and body core temperatures to season and reproductive context. Nature Sci. Rep. 8: 2142. https://doi.org/10.1038/s41598-018-20655-z
Win A (2001) Seasonal grazing of Canada goose (Branta canadensis) on high country farmland, Canterbury, New Zealand. Master Thesis, Lincoln University, New Zealand. https://researcharchive.lincoln.ac.nz/bitstream/handle/10182/2678/win_thesis.pdf;jsessionid=2F76579ADB4517ABFC069CDE3406C5EF?sequence=4
Acknowledgements
The author wishes to thank Jarle Morten Enoksen and Ole-Henrik Enoksen for letting me conduct the project on Musvær and for being very helpful when it was needed. This study was funded by the Ministry of Agriculture and Food, Norway, through NIBIO- Norwegian Institute of Bioeconomy and the County Governor of Troms and Finnmark County, Norway (grant no. 2020/528). The author wishes to thank these funding sources.
Funding
Open access funding provided by Norwegian Institute of Bioeconomy Research
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The author declares no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Aarseth, J.J. Complete seasonal surveillance of Greylag Goose (Anser anser) foraging behavior on dairy grassland and the effect of limited lethal scaring. Eur J Wildl Res 69, 53 (2023). https://doi.org/10.1007/s10344-023-01680-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-023-01680-4