
Abstract
Anthropogenic activities, such as outdoor recreation, have the potential to change complex interactions between wildlife and livestock, with further consequences for the management of both animals, the environment, and disease transmission. We present the interaction amongst wildlife, livestock, and outdoor recreationists as a three-way interaction. Little is known about how recreational activities alter the interaction between herbivores in areas extensively used for recreational purposes. We investigate how hiking activity affects spatio-temporal co-occurrence between domestic sheep (Ovis aries) and red deer (Cervus elaphus). We used camera traps to capture the spatio-temporal distribution of red deer and sheep and used the distance from the hiking path as a proxy of hiking activity. We used generalized linear models to investigate the spatial distribution of sheep and deer. We analysed the activity patterns of sheep and deer and then calculated their coefficients of temporal overlap for each camera trap location. We compared these coefficients in relation to the distance from the hiking path. Finally, we used a generalized linear mixed-model to investigate which factors influence the spatio-temporal succession between deer and sheep. We do not find that sheep and red deer spatially avoid each other. The coefficient of temporal overlap varied with distance from the hiking trail, with stronger temporal co-occurrence at greater distances from the hiking trail. Red deer were more likely to be detected further from the path during the day, which increased the temporal overlap with sheep in these areas. This suggests that hiking pressure influences spatio-temporal interactions between sheep and deer, leading to greater temporal overlap in areas further from the hiking path due to red deer spatial avoidance of hikers. This impact of recreationists on the wildlife and livestock interaction can have consequences for the animals’ welfare, the vegetation they graze, their management, and disease transmission.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Interspecific interactions such as predation and competition are key components of a healthy ecosystem (Gurevitch et al. 2000). Amongst these, wildlife-livestock interactions are particularly important in processes of disease transmission (Dohna et al. 2014), biodiversity conservation (Fynn et al. 2016), and livestock management (Ager et al. 2004). Interactions between wildlife and livestock are complex, and changes to these interaction processes can have indirect cascade effects on lower trophic levels (Otuoma et al. 2009; Wilson et al. 2020). For example, interspecific interactions can be altered due to abiotic stressors such as variation in temperature (Tylianakis et al. 2010). Beyond abiotic stressors, human biotic stressors, such as anthropogenic pressure, can alter interspecific interaction patterns and processes. The interaction between human activity, wildlife, and livestock can be considered as a three-way interaction. It can involve one or multiple types of human activities which can alter the spatial or temporal interaction between the wildlife and the livestock. Such three-way interaction systems are important to understand depredation (Gemeda and Meles 2018), impacts on habitat (Otuoma et al. 2009), and disease transmission (Simpson et al. 2018). Anthropogenic pressures in such three-way interaction systems can take multiple forms and are commonly related to processes such as habitat fragmentation (Otuoma et al. 2009) or urbanization (Hassell 2018).
Outdoor recreation activities such as mountain biking, skiing, and hiking can have various impacts on wildlife which are not dissimilar to predation (Frid and Dill 2002). Spatio-temporal interactions between wildlife and outdoor recreation activities have been studied extensively (Larson et al. 2016) and are shown to impact animal diets (Jayakody et al. 2011), feeding behaviours (Szemkus et al. 1998; Griffin et al. 2007), distributions (Sibbald et al. 2011), and predation behaviour (Belotti et al. 2012). However, much less is known about how recreational activities alter the competition between herbivores in spaces where human recreation is present, i.e., do they exhibit competition or spatial avoidance?
Red deer (Cervus elaphus) and sheep (Ovis aries) both graze, at significant densities, within the Scottish Highlands where they are food competitors (Clutton-Brock and Albon 1989). The interaction between red deer and sheep is complex. DeGabriel et al. (2011) found that the spatial absence of sheep was correlated with a higher density of deer, but this was dependant on the time elapsed since sheep removal. Similarly, Hope et al. (1996) found that the removal of sheep on hills increased the number of red deer: the so-called ‘vacuum effect’ (Albon et al. 2007). However, few studies have investigated how external stressors — such outdoor recreation activity — may modify these spatio-temporal relationships.
The Scottish Highlands are a multi-functional landscape, used for outdoor recreation (e.g., hiking, biking), hunting (e.g., red deer and red grouse (Lagopus lagopus scotica)) and the grazing of livestock (e.g., sheep and cattle (Bos taurus)). Previous studies, based in Scotland, on the impacts of hikers on red deer revealed that hiking activity can change diet composition (Jayakody et al. 2011), increase vigilance (Jayakody et al. 2008), and alter their movement patterns (Sibbald et al. 2011). Thus, outdoor recreation activity can cause red deer to be spatially displaced to less nutritious habitats and subsequently forced to compete with other species that they may usually avoid. In Scotland, sheep are often in direct contact with farmers, and it is typically assumed that sheep are habituated to the presence of humans. Thus, it is believed that the impact of hiking activity on their spatio-temporal distribution will be limited to very small scales (i.e., displacement of only meters for a few minutes).
This study presents a novel investigation of the three-way interaction between sheep, deer, and outdoor recreation activity. To date, there is very limited evidence on the role of outdoor recreation in modulating the interaction between competing foragers. Specifically, we study how hiking activity (represented here as the distance from a popular hiking path) influences the spatio-temporal patterns of co-occurrence between red deer and sheep in the Scottish uplands. Our hypothesis is that sheep will exhibit low/no response to hiking activity and that red deer will spatially avoid the hiking trail. We further hypothesize that the level of spatio-temporal co-occurrence between red deer and sheep will vary in response to the distance from the hiking trail as the hiking activity is highly concentrated along the trail in our study area. We expect red deer and sheep will have greater spatio-temporal co-occurrence in areas less used by hikers due to spatial avoidance of recreationist activity by red deer in high hiking pressure areas.
Methods
Study area
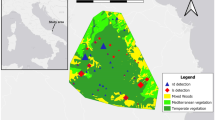
The study area is the Glen Lyon trail at the North Chesthill Estate (56°37′04.5″N 4°10′50.7″W) (Fig. 1). The 17-km Glen Lyon loop hiking trail is popular with hikers because it facilitates access to four summits — called Munros.Footnote 1 This area has a population of red deer estimated to be ~ 380 in 2019 (density of 13.91 deer/km2; Deer Management Plan, Breadalbane DMG). This population varies, however, as the estate perimeter is largely unfenced. The population size is surveyed and managed to avoid overgrazing impacts. Stag culling by the land manager occurs every year between late August and late October, and is followed by culling of hinds until the end of February. In 2018, the hunting season was between the 30th of August to the 20th of October; in 2019, this season was from the 30th of September to the 19th of October. During summer (from the middle of May, i.e., after lambing), around 950 ewes with lambs (35.18/km2) are present across the estate until mid-October, when they are removed from the hills.

Camera trap study design in Glen Lyon, Scotland, showing location, year of deployment, and habitat type of the camera trap sites. In total, we have n = 45 year-location replicates
Data collection
Camera traps
Data collection occurred over two summers: from the end of June 2018 to the end of October 2018 and from the end of May 2019 to the end of October 2019. We used camera traps to study the spatial and temporal distribution of red deer and sheep. We used a stratified sampling design to ensure that camera trap locations provided a cross-section of habitat types, elevations, and distances to the hiking path (Fig. 1). Some habitat types were less present in the study areas (e.g., bracken), and thus, less camera traps were deployed on this habitat. However, each habitat was found at different distances from the hiking path.
In 2018, we deployed 22 camera traps in the west and in the middle of the area. In 2019, we used 23 camera traps in the middle and in east of the study area. Some camera trap locations are used in both 2018 and in 2019 (Fig. 1 and see supplementary material 1 for more details on the camera traps and the study design). We calibrated each camera trap to trigger three photos per animal detection with no delay between triggers. Each camera was positioned at a height of 1.10 m using a wooden post. We determined the dominant type of vegetation in front of each camera trap, using an existing vegetation classification protocol (JNCC 2010).
Hiking activity
For each camera trap location, the distance from the hiking path was used as a proxy of the hiking activity. To assess if this proxy was representative of the hiking activity, we collected GPS tracking data from hikers. We approached groups of hikers at the entrance of the hiking loop between 7 a.m. and 1 p.m. We asked them to carry a GPS tracker (i-Blue 747proS GPS Trip Recorder) during the length of their hikes. One GPS tracker was given per hiking group, and we asked them to leave their tracker in a drop box close to the car park at the end of their day. We repeated GPS data collection across a total of 60 days, spread between the summers 2017, 2018, and 2019. These GPS data were used to estimate the percentages of hikers performing the full hiking loop and leaving the hiking path. We used a kernel density estimation with a 10-m kernel bandwidth of all the GPS tracks.
Data analysis
Analysis of spatial activity
We recorded the number of deer and sheep detected at each location. We classified these detections into detection events, where a new detection event is defined as when two photos of the same species were taken with, at a minimum, a 10-min interval between detections. Previous studies have used a time threshold of 30 min to define detection events (Sollmann 2018); however, we adopted a smaller value because of the relatively high density of deer and sheep in the area. We collected 4756 independent detection events of deer and sheep over 5097 camera trap survey effort days. Our data comprise 2418 detections of deer, 2119 detections of sheep, with 41 detections that included both deer and sheep. The 41 co-occurrence detections are primarily a result of the open landscape (one of the two species triggering the camera in the foreground and the other species being observed in the background). These detections were considered as both deer and sheep detection.
We defined survey effort as the number of days that the camera trap was actively working and calculated the detection rate (DR) for each camera as the total number of animals detected (i.e., sum of the maximum number of animals counted for each detection event) divided by the total number of working days (O’Brien et al. 2003). Detection rates were calculated separately for both sheep and red deer. We calculated separate detection rates for day and night. We used these detection rates as the dependent variable in a generalized linear model (GLM) using a Gaussian link function. We included as independent variables distance from the hiking path (log transformed), vegetation type (Bog, Bracken, Dry Heath, Wet Heath, Montane), elevation, and the detection rate of the other species (e.g., DR of sheep when DR of deer was the independent variable and vice-versa). After checking for collinearity between the variables, we removed the elevation from further analysis (VIF > 5) (Zuur et al. 2017). We used model selection with the “dredge” function in the R package “MuMIn” (which fitted and compared all the possible combination of models) (Bartoń 2020) and was based on the Akaike Information Criterion corrected for small sample sizes (AICc) (Akaike et al. 1973). To ensure that we retained the most parsimonious model, we retained models for inference if they had ΔAICc < 6 and if they were not a more complex version of any model with a lower AIC value (Richards et al. 2011).
Analysis of temporal activity
We identified red deer and sheep daily activity patterns using the non-parametric kernel density estimation method (Ridout and Linkie 2009). We used kernel density estimation of the time of observation to obtain the probability distribution of each species’ activity pattern. From the daily activity curves, we calculated the coefficient of overlap, Δ, which estimates the temporal overlap in activity between two species (Niedballa et al. 2019). This coefficient is the joint area (i.e., intersection) under the probability density functions of the estimated daily activity density curves of both species. It ranges from 0 (no temporal overlap) to 1 (full temporal overlap). Ridout and Linkie (2009) present three different methods to estimate Δ and suggest using the estimator Δ4 (which uses vectors of densities estimated at the time of observation of the two species) for sample sizes larger than 50. We therefore used Δ4 to calculate temporal overlap for all our camera traps combined. We calculated the 95% confidence interval of this estimate from 1000 bootstrap samples. We used a Watson U2 test to statistically compare the activity patterns of sheep and deer (Landler et al. 2021).
To estimate if the coefficient of temporal overlap (Δ4) varied with hiking impact (i.e., distance from the hiking path), we then calculated the coefficient of overlap (and corresponding confidence interval) for each camera trap location. We again used the estimator Δ4 because most of the camera traps (82%) had more than 50 observations of sheep and/or deer. However, some camera traps detected fewer than two deer and/or two sheep and the coefficient of overlap could not be estimated accurately. Five locations (locations 3,5, 7, 16 and 24) were removed from this analysis due to their overall low deer and/or sheep detection. Moreover, to increase the accuracy of our analysis, we removed detection (n = 30) where the time recorded of the photo did not match the real time (e.g., the camera traps reset itself in a middle of a period of observation). We tested the correlation between Δ4 for each individual camera and the estimated distances from the hiking track using a Pearson’s correlation test. The distance from the hiking path was defined as the Euclidian distance between a camera trap location and the hiking path obtained using the package sf in R (Pebesma 2018). Finally, we performed individual Watson U2 tests to statistically compare the activity patterns of sheep and deer at each camera trap location.
Spatio-temporal co-occurrence
We investigated spatio-temporal co-occurrence between red deer and sheep to test our hypothesis that red deer avoid sheep. To do this, we looked at the pattern of co-occurrence, and the timing of detections relative to the other species. Niedballa et al. (2019) recommend using the time interval between the observation of one species and the second species as the dependent variable in a regression modelling framework. We calculated these time intervals (in hours) for each camera, separating detections between sheep and deer and vice versa (hereafter “time interval”). We kept for analysis only time intervals less than 24 h to focus only on finer-scale daily temporal co-occurrence. For this analysis, we also removed detection where not matching real time (n = 30 as explained above). The time intervals (log transformed) were used as the dependent variable in a generalized linear mixed model (GLMM) using a Gaussian link function.
We included seven different independent variables in this model: the type of detection (i.e., which species preceded the other, deer-sheep, and type sheep-deer), the herd size of the first and the second species detected (sum of the number of deer or sheep within a sequence used for the start of the time interval), the type of habitat (five categories of habitat type described above), the hunting season, the elevation, the distance from the hiking path in meters, and the year. We tested the collinearity between the independent variable and did not remove any variables from further analysis (VIF < 5) (Zuur et al. 2017). Camera location was included as a random effect and model selection used the "dredge" function, following the protocol outlined above.
All analyses were conducted in the statistical software R version 4.0.3 (R Core Team 2020). We used the package Overlap (Meredith and Ridout 2014) to compute the temporal overlap statistics, and the package nlme to fit the statistical models (Pinheiro et al. 2020).
Results
Hiking activity
We found that out of the 252 hikers’ GPS tracks we collected, 83% completed the full delimited hiking loop and 90% did not go off track (Supplementary material 2). Thus, the distance from the hiking path is accurate proxy of the hiking activity as most of the hikers we surveyed stay on the hiking path.
Spatial activity analyses
We found that, during the day, red deer were detected at higher rates away from the path (Fig. 2a; Table 1). We also found the null model to be with ΔAICc < 6 from the best performing model. However, we found that the distance model was 18 times better than the null model (from the weights of the models: 0.669/0.037). The detection of red deer was not related to the detection rate of sheep or vegetation. However, during the night, red deer detection was not dependent on the distance from the hiking path (Fig. 2b) but depended on vegetation type (the only variable retained in the most parsimonious model) (Table 1). More specifically, red deer were more frequently detected in dry heath vegetation during the night. However, we also found the null model to be within ΔAICc < 6 from the best performing model. The vegetation model performed 3.3 times (from the weights of the models: 0.446/0.134) better than the null model.

Scatterplots of the detection rates of a and b deer and c and d sheep vs distances (log transformed) from the hiking path. Each location in the study area (i.e., the location of one camera trap) is represented by a dot in each panel. Blue dots are detection during day and black dots during the night
Overall, sheep were less frequently detected at night than during the day and we did not detect a distance effect (Fig. 2c, d). The detection of sheep was driven by vegetation type during both day and night (Table 1). Sheep were more frequently detected in montane vegetation during the day (Table 1) and in montane and dry heath vegetation during the night (Table 1).
Temporal activity analyses
Sheep and deer had two different activity patterns (Watson U2 test = 36.63, p-value < 0.001). Temporal detections of red deer peaked in the morning and in the evening, with much lower activity in the middle of the day (Fig. 3). Sheep temporal activity increased during the morning and reached an approximately constant level between 9 a.m. and 6 p.m., after which it decreased. The overall coefficient of temporal overlap (including all camera trap locations) was 0.502 (95% CI: 0.479–0.526). Sheep and deer activity overlapped the most during the early morning (5–9 a.m.) and in the evening (from 6 to 11 p.m.).

Daily activity patterns (kernel density curves) of red deer (red line) and sheep (blue dotted line). The overlap of red deer and sheep activities is shown as the grey shaded area. The coefficient of overlap (Δ4) is 0.502 (95% CI: 0.479–0.526)
We calculated the temporal overlap coefficient for each camera trap location (Fig. 4a; Supplementary material 3) and their corresponding Watson’s U2 test of homogeneity (Supplementary material 4). This showed a low level of temporal overlap (Δ4 between 0 to 0.15) for the camera traps which were mostly situated close to the hiking path (e.g., camera traps 9, 10, 20, and 22) (Fig. 4a, b). We found more moderate levels of temporal overlap (Δ4 = 0.30 to 0.60) for the camera traps which were located at moderate distances from the hiking path (e.g., camera traps 6, 26, 27). The highest levels of temporal overlap (Δ4 = 0.45 to 0.75) were observed in camera traps further from the hiking path (e.g., camera traps 2, 17, and 18). There was a positive relationship (Pearson correlation p = 0.0051) between Δ4 and distance from the hiking trail (Fig. 4b). On average, an increase of ~ 500 m from the path led to an increase of ~ 10% in the extent of overlap.

Temporal overlap of sheep and deer in our study area. a Map showing the temporal overlap (Δ4) at each location b scatterplot of the red deer and sheep temporal overlap from each camera trap depending on their distances from the hiking path. The blue line is the linear regression line, and the grey shaded area shows the 95% confidence interval. Each dot represents a camera trap location for which we calculated the Δ4. The kernel density curves for the red (location 17) and purple (location 20) dots are shown in panels c and d as examples of high Δ4 (location 17, in red, Δ4 = 0.63) and low Δ4 (location 20, in purple, Δ4 = 0.057)
Spatio-temporal co-occurrence
We investigated which variables influenced the time interval of sheep-deer and deer-sheep successions using a GLMM and a model selection approach (Table 2). The most parsimonious model included only the type of succession as a variable. While the null model is within ΔAICc < 6 from the best performing model, we also found that the vegetation model was 6.89 times (weight of the models: 0.800/0.116) better than the null model. Based on the variable selection process, the other variables which included the distances from the path and habitat type were not retained in the most parsimonious model.
With this most parsimonious model, we found a significant negative relationship between the time interval and the succession of sheep by deer (in comparison to the deer-sheep succession) (Table 2); the time interval was significantly less important when a deer detection followed a sheep detection than the opposite succession.
Discussion
Our study used a novel approach to study the three-way interaction between wildlife, livestock, and humans at the spatial, temporal, and spatio-temporal scales. We focused on how interspecific interaction between sheep and red deer is influenced by the presence of outdoor recreation activity represented here as the distance from a popular hiking path. We study this three-way interaction system using camera trap data in Scotland. Red deer and sheep are both central to the history of this rural country. Red deer have been hunted for centuries for recreational purposes by private landowners (Clutton-Brock and Albon 1989), who also use the land for sheep grazing, mainly during the summer months. In many cases, human activities in the landscape add a further element that can alter the interaction between livestock and wildlife species (Otuoma et al. 2009; Hassell et al. 2017; Miller and Schmitz 2019). In systems where they co-exist, land management needs to consider how the spatio-temporal interactions occur across the landscape (e.g., Fleischner 1994; Edwards et al. 1996), especially when these species are competing for resources (Madhusudan 2004).
In our study, the spatial distribution of red deer and sheep did not depend on the presence of the other species. We first showed that, as we assumed, sheep were not spatially or temporally displaced by the presence of hikers. During the day, sheep distribution was explained by vegetation; however, this model was not significantly different from the null model. Similarly, the time interval was as well explained by the null model than by the type of succession; thus, we cannot conclude that one species avoids this other more than the other. We found that during the day, detection rates of red deer were greater at distances further from the hiking path. Previous work has demonstrated that red deer tend to avoid recreational activity (Coppes et al. 2017), including hiking areas (Sibbald et al. 2011), and favour areas where human activities are less prevalent (Marion et al. 2021). Thus, at the landscape scale, hiking activity influences interactions between red deer and sheep by spatially displacing red deer populations.
Red deer spatial displacement can have consequences for animal management (Fleischner 1994; Edwards et al. 1996). We did not find that the hunting season was associated with the spatio-temporal co-occurrence of red deer and sheep. However, we did not study the opposite, if the change in spatio-temporal co-occurrence of red deer and sheep due to the presence of hikers impacted their successful management. Conflicts between hunters and hiking groups exist and take multiple sources such as the success of one activity due to the presence of the other (Reis and Higham 2009). Here, the presence of hikers leads to a change in red deer spatial distribution which will have consequences for their management depending on the hunter’s ability to locate the animal.
Red deer and sheep exhibit different patterns of temporal activity. We found red deer to exhibit the typical crepuscular pattern found in previous studies (Georgii 1981), whereas sheep exhibit a more diurnal pattern (Squires 1975). Temporal overlap was higher at distances away from the hiking trail, which may be due to the spatial displacement of red deer in response to hiking pressure. During the day, when most hiking activities occur, we found the distance effect of the hiking trail to be stronger for red deer relative to the night. Stronger temporal overlap might suggest more direct interaction or indirect competition (de Boer and Prins 1990) between red deer and sheep in foraging areas further from the trail. Such indirect competition can affect the welfare of both species (Osborne 1984; Edwards et al. 1996). Red deer welfare may be compromised if they are displaced to less nutritious foraging habitat (Osborne 1984), which can result in a change in diet (Jayakody et al. 2011). Sheep weight (as a proxy for animal welfare) has previously been found to be negatively affected by the presence of wildlife competing for food (Edwards et al. 1996). However, habitat had no impact on this relationship in our study, so we have no direct evidence for dietary change or stress in this case.
The higher level of temporal overlap in areas further away from the hiking path can have consequences for the vegetation grazed by both species in these areas (Wilson et al. 2020). Sheep and red deer are both intensive grazers and can limit vegetation regeneration and growth (Gordon 1988; Hester et al. 1999; Pollock et al. 2005). In the UK, intensive grazing and browsing can prevent tree regeneration (Clutton-Brock et al. 2004) and can degrade the conservation value of heather moorland (Clarke et al. 1995; Palmer and Hester 2000), which are recognized for their conservation status with distinctive bird communities and vegetation diversity (Thompson et al. 1995).
Our study reveals the need to take into consideration all the different components which can influence the spatial and temporal overlap of two species (i.e., here the human component). The increase in spatio-temporal overlap between livestock and wildlife can lead to ecological and economic issues such as an increase in disease transmission (Riley et al. 1998; Barasona et al. 2014). Understanding wildlife movement and the cause of the movement can limit disease spread (Berentsen et al. 2014). Here, we demonstrated how the changes in interspecific interactions can be studied at the temporal and spatial scale using camera traps.
In our study, we did not assess how changes in deer and sheep distribution — as a result of hiking activity — lead to indirect cascade effects (e.g., grazing impact). Future research focusing on herbivore-recreationist interactions should investigate indirect cascade effects (Wilson et al. 2020) on animal welfare, vegetation, animal management, and disease transmission and explore how different densities of herbivores and recreationists can alter these effects.
Conclusion
In this study, we demonstrated that level of spatial, temporal, and spatio-temporal overlap between sheep and deer was modified by hiking pressure, here expressed as a distance from the hiking path. Red deer were spatially displaced during the day which results in a higher temporal overlap with sheep further from the hiking path. Our study opens a new perspective on the study of complex interspecies interaction and their consequences. Further study is required to understand spatial–temporal variations in cattle and wildlife densities, and their significance for animal nutrition, vegetation condition, their management, and disease transmission.
Notes
Munros are mountains in Scotland over 3000 feet; Munros hiking is a widespread activity as people aim to hike each of the 282 Scottish Munros. Thus, its four Munros and its relatively close location to Glasgow and Edinburgh make the area a prime destination for recreational hikers escaping who live in the city.
References
Ager AA, Johnson BK, Coe PK, Wisdom MJ (2004) Landscape simulation of foraging by elk, mule deer, and cattle on summer range. Transactions of the 69th North American Wildlife and Natural Resources Conference 687–707
Akaike H, Petrov BN, Csaki F (1973) Information theory and an extension of the maximum likelihood principle. Springer, pp. 267–281
Albon SD, Brewer MJ, O’brien S et al (2007) Quantifying the grazing impacts associated with different herbivores on rangelands. J Appl Ecol 44:1176–1187. https://doi.org/10.1111/j.1365-2664.2007.01318.x
Barasona JA, Latham MC, Acevedo P et al (2014) Spatiotemporal interactions between wild boar and cattle: implications for cross-species disease transmission. Vet Res 45:122. https://doi.org/10.1186/s13567-014-0122-7
Bartoń K (2020) MuMIn: multi model inference: model selection and model averaging based on information criteria. Version 1.43.17. https://CRAN.R-project.org/package=MuMIn. Accessed 11 Oct 2021
Belotti E, Heurich M, Kreisinger J et al (2012) Influence of tourism and traffic on the Eurasian lynx hunting activity and daily movements. Anim Biodivers Conserv 12
Berentsen AR, Miller RS, Misiewicz R et al (2014) Characteristics of white-tailed deer visits to cattle farms: implications for disease transmission at the wildlife–livestock interface. Eur J Wildl Res 60:161–170. https://doi.org/10.1007/s10344-013-0760-5
Clarke JL, Welch D, Gordon IJ (1995) The influence of vegetation pattern on the grazing of heather moorland by red deer and sheep. I. The Location of Animals on Grass/Heather Mosaics. The Journal of Applied Ecology 32:166. https://doi.org/10.2307/2404426
Clutton-Brock TH, Albon SD (1989) Red deer in the highlands. Blackwells
Clutton-Brock TH, Coulson T, Milner JM (2004) Red deer stocks in the Highlands of Scotland. Nature 429:261–262. https://doi.org/10.1038/429261a
Coppes J, Burghardt F, Hagen R et al (2017) Human recreation affects spatio-temporal habitat use patterns in red deer (Cervus elaphus). PLoS ONE. https://doi.org/10.1371/journal.pone.0175134
de Boer WF, Prins HHT (1990) Large herbivores that strive mightily but eat and drink as friends. Oecologia 82:264–274. https://doi.org/10.1007/BF00323544
DeGabriel JL, Albon SD, Fielding DA, Riach DJ, Westaway S, Irvine RJ (2011) The presence of sheep leads to increases in plant diversity and reductions in the impact of deer on heather. J Appl Ecol 48(5):1269–1277. https://doi.org/10.1111/j.1365-2664.2011.02032.x
Edwards GP, Croft DB, Dawson TJ (1996) Competition between red kangaroos (Macropus rufus) and sheep (Ovis aries) in the arid rangelands of Australia. Austral Ecol 21:165–172. https://doi.org/10.1111/j.1442-9993.1996.tb00597.x
Fleischner TL (1994) Ecological costs of livestock grazing in Western North America. Conserv Biol 8:629–644
Frid A, Dill L (2002) Human-caused disturbance stimuli as a form of predation risk. Conserv Ecol. https://doi.org/10.5751/ES-00404-060111
Fynn RWS, Augustine DJ, Peel MJS, de Garine-Wichatitsky M (2016) Strategic management of livestock to improve biodiversity conservation in African savannahs: a conceptual basis for wildlife–livestock coexistence. J Appl Ecol 53:388–397. https://doi.org/10.1111/1365-2664.12591
Gemeda DO, Meles SK (2018) Impacts of human-wildlife conflict in developing countries. J Appl Sci Environ Manag 22:1233–1238. https://doi.org/10.4314/jasem.v22i8.14
Georgii B (1981) Activity patterns of female red deer (Cervus elaphus L.) in the Alps. Oecologia 49:127–136. https://doi.org/10.1007/BF00376910
Griffin SC, Valois T, Taper ML, Scott Mills L (2007) Effects of tourists on behavior and demography of olympic marmots. Conserv Biol 21:1070–1081. https://doi.org/10.1111/j.1523-1739.2007.00688.x
Gordon IJ (1988) Facilitation of red deer grazing by cattle and its impact on red deer performance. J Appl Ecol 25:1–9. https://doi.org/10.2307/2403605
Gurevitch J, Morrison JA, Hedges LV (2000) The Interaction between competition and predation: A meta-analysis of field experiments. Am Nat 155(4):435–453. https://doi.org/10.1086/303337
Hassell JM (2018) Ecological and epidemiological consequences of rapid urbanisation at wildlife-livestock-human interfaces. The University of Liverpool (United Kingdom), Ph.D.
Hester AJ, Gordon IJ, Baillie GJ, Tappin E (1999) Foraging behaviour of sheep and red deer within natural heather/grass mosaics. J Appl Ecol 36:133–146
Hassell JM, Begon M, Ward MJ, Fèvre EM (2017) Urbanization and disease emergence: dynamics at the wildlife–livestock–human interface. Trends Ecol Evol 32:55–67. https://doi.org/10.1016/j.tree.2016.09.012
Hope D, Picozzi N, Catt DC, Moss R (1996) Effects of reducing sheep grazing in the Scottish Highlands. J Range Manag 49(4):301. https://doi.org/10.2307/4002587
Jayakody S, Sibbald AM, Gordon IJ, Lambin X (2008) Red deer Cervus elephus vigilance behaviour differs with habitat and type of human disturbance. Wildl Biol 14:81–91. https://doi.org/10.2981/0909-6396(2008)14[81:RDCEVB]2.0.CO;2
Jayakody S, Sibbald AM, Mayes RW et al (2011) Effects of human disturbance on the diet composition of wild red deer (Cervus elaphus). Eur J Wildl Res 57:939–948. https://doi.org/10.1007/s10344-011-0508-z
JNCC (2010) Handbook for Phase 1 habitat survey - a technique for environmental audit. JNCC 83
Landler L, Ruxton GD, Malkemper EP (2021) Comparing two circular distributions: advice for effective implementation of statistical procedures in biology. https://doi.org/10.1101/2021.03.25.436932
Larson CL, Reed SE, Merenlender AM, Crooks KR (2016) Effects of recreation on animals revealed as widespread through a global systematic review. PLoS ONE 11:e0167259. https://doi.org/10.1371/journal.pone.0167259
Madhusudan MD (2004) Recovery of wild large herbivores following livestock decline in a tropical Indian wildlife reserve. J Appl Ecol 41:858–869. https://doi.org/10.1111/j.0021-8901.2004.00950.x
Marion S, Demšar U, Davies AL et al (2021) Red deer exhibit spatial and temporal responses to hiking activity. Wildl Biol. https://doi.org/10.2981/wlb.00853
Meredith M, Ridout M (2014) Overview of the overlap package. https://cran.r-project.org/web/packages/overlap/vignettes/overlap.pdf
Miller JRB, Schmitz OJ (2019) Landscape of fear and human-predator coexistence: applying spatial predator-prey interaction theory to understand and reduce carnivore-livestock conflict. Biol Cons 236:464–473. https://doi.org/10.1016/j.biocon.2019.06.009
Niedballa J, Wilting A, Sollmann R et al (2019) Assessing analytical methods for detecting spatiotemporal interactions between species from camera trapping data. Remote Sensing in Ecology and Conservation. https://doi.org/10.1002/rse2.107
O’Brien TG, Kinnaird MF, Wibisono HT (2003) Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim Conserv 6(2):131–139. https://doi.org/10.1017/S1367943003003172
Osborne BC (1984) Habitat use by red deer (Cervus elaphus L.) and hill sheep in the West Highlands. J Appl Ecol 21:497–506. https://doi.org/10.2307/2403424
Otuoma J, Kinyamario J, Ekaya W et al (2009) Effects of human–livestock–wildlife interactions on habitat in an eastern Kenya rangeland. Afr J Ecol 47:567–573. https://doi.org/10.1111/j.1365-2028.2008.01009.x
Palmer SCF, Hester AJ (2000) Predicting spatial variation in heather utilization by sheep and red deer within heather/grass mosaics. J Appl Ecol 37:616–631. https://doi.org/10.1046/j.1365-2664.2000.00515.x
Pebesma E (2018) Simple Features for R: Standardized support for spatial vector data. The R Journal 10:439–446
Pinheiro J, Bates D, DebRoy S et al (2020) nlme: linear and nonlinear mixed effects models. https://cran.r-project.org/web/packages/nlme/nlme.pdf
Pollock ML, Milner JM, Waterhouse A et al (2005) Impacts of livestock in regenerating upland birch woodlands in Scotland. Biol Cons 123:443–452. https://doi.org/10.1016/j.biocon.2005.01.006
R Core Team (2020) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Reis AC, Higham JES (2009) Recreation conflict and sport hunting: moving beyond goal interference towards social sustainability. Journal of Sport & Tourism 14:83–107. https://doi.org/10.1080/14775080902965025
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. JABES 14:322–337. https://doi.org/10.1198/jabes.2009.08038
Richards SA, Whittingham MJ, Stephens PA (2011) Model selection and model averaging in behavioural ecology: the utility of the IT-AIC framework. Behav Ecol Sociobiol 65:77–89. https://doi.org/10.1007/s00265-010-1035-8
Riley SP, Hadidian J, Manski DA (1998) Population density, survival, and rabies in raccoons in an urban national park. Can J Zool 76:1153–1164. https://doi.org/10.1139/z98-042
Sibbald AM, Hooper RJ, McLeod JE, Gordon IJ (2011) Responses of red deer (Cervus elaphus) to regular disturbance by hill walkers. Eur J Wildl Res 57:817–825. https://doi.org/10.1007/s10344-011-0493-2
Simpson GJG, Quan V, Frean J et al (2018) Prevalence of selected zoonotic diseases and risk factors at a human-wildlife-livestock interface in Mpumalanga Province, South Africa. Vector-Borne and Zoonotic Diseases 18:303–310. https://doi.org/10.1089/vbz.2017.2158
Squires VR (1975) Ecology and behaviour of domestic sheep (Ovis aries): a review. Mammal Rev 5:35–57. https://doi.org/10.1111/j.1365-2907.1975.tb00186.x
Sollmann R (2018) A gentle introduction to camera-trap data analysis. Afr J Ecol 56:740–749. https://doi.org/10.1111/aje.12557
Szemkus B, Ingold P, Pfister U (1998) Behaviour of Alpine ibex (Capra ibex ibex) under the influence of paragliders and other air traffic. Zeitschrift Fur Saugetierkunde-International Journal of Mammalian Biology 63:84–89
Thompson DBA, MacDonald AJ, Marsden JH, Galbraith CA (1995) Upland heather moorland in Great Britain: a review of international importance, vegetation change and some objectives for nature conservation. Biol Cons 71:163–178. https://doi.org/10.1016/0006-3207(94)00043-P
Tylianakis JM, Laliberté E, Nielsen A, Bascompte J (2010) Conservation of species interaction networks. Biol Cons 143:2270–2279. https://doi.org/10.1016/j.biocon.2009.12.004
Wilson MW, Ridlon AD, Gaynor KM et al (2020) Ecological impacts of human-induced animal behaviour change. Ecol Lett 23:1522–1536. https://doi.org/10.1111/ele.13571
zu Dohna H, Peck DE, Johnson BK et al (2014) Wildlife–livestock interactions in a western rangeland setting: quantifying disease-relevant contacts. Prev Vet Med 113:447–456. https://doi.org/10.1016/j.prevetmed.2013.12.004
Zuur AF, Ieno EN, Saveliev AA (2017) Beginner’s guide to spatial, temporal, and spatial-temporal ecological data analysis with R-INLA. Highland Statistics, Newburgh, UK
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Marion, S., Demšar, U., Davies, A.L. et al. Spatial and temporal variations in interspecific interaction: impact of a recreational landscape. Eur J Wildl Res 68, 36 (2022). https://doi.org/10.1007/s10344-022-01584-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-022-01584-9