
Abstract
This study provides for the first time in the Carpathians analysis of spatial behaviour of 20 male red deer monitored during 2005–2013 using radio-telemetry. Two distinct spatial patterns were displayed in the same local population, i.e. residential and migratory. Residential annual home ranges were significantly smaller compared to migratory ones using both Minimum Convex Polygon and Kernel Home Range, due to periodic movement of migrants between distinct seasonal ranges. Residents remained in the same area throughout the year and showed a positive age effect on the home range size. While seasonal ranges of migrants were comparable in size, residents surprisingly expanded their space use in winter compared to other seasons. Fidelity to seasonal ranges over the years, especially winter, was striking and comparable in both migrants and residents with increasing tendency throughout the year (37–68 %). Vertical differences in home range altitudes were most obvious in migrants, although in both migrant types, significant descent was recorded during the winter. The longest horizontal movements were recorded in three young stags emigrating to neighbouring mountain ranges (30, 47 and 65 km). Because 45 % of the population seems to be migratory, the data support importance of large-scale hunting management established in Slovakia in 2009.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The European red deer (Cervus elaphus) is the flagship game species in Slovakia; however, data on spatial behaviour of the species are scarce from this area. In general, spatial behaviour of mammals is influenced by a variety of physiological, ecological and social factors, such as metabolic needs (McNab 1963), body mass (Harestad and Bunell 1979; Lindstedt et al. 1986), mating system (Clutton-Brock and Harvey 1978; Clutton-Brock 1989), age (Georgii and Schröder 1983; Szemethy et al. 1998; Kamler et al. 2008), population density (Vincent et al. 1995; Loe et al. 2009), distribution of feeding habitats (Schmidt 1993; Mysterud et al. 2001) and predation and human disturbance (Hebblewhite and Merrill 2007), resulting in a significant seasonal variation. In this context, ungulates of northern hemispheres reduce food intake, activity and movement during winter within restricted home ranges (Georgii 1980; Clutton-Brock et al. 1982; Georgii and Schröder 1983). Mountainous environments emphasize the need to preserve energy during nutritional stagnation and to compensate winter food intake loss during summer (Luccarini et al. 2006).
A strategy to compensate loss is migration to lower altitudes during the winter and, above all, towards upper altitudes during summer. The triggering mechanism of downhill migration is considered to be 20–25 cm of continuous snow cover (Schmitd and Gossow 1991). In temperate regions, the vertical movement of cervids from low elevation winter range to high elevation summer range is the most common pattern of migration (Mysterud et al. 2001). Several processes might contribute to altitudinal expansion of populations during summer (Van Beest et al. 2011). The most prominent among these is clearly the forage maturation hypothesis, which formalizes how access to a more continuous supply of highly nutritious, newly emergent forage is a driving force of migration in large herbivores (Fryxell and Sinclair 1988; Hebblewhite et al. 2008; Mysterud 2013). Temporal shift in phenology allows prolonged access to young, high-quality forage which influences direction and timing of seasonal uphill movements (Mysterud et al. 2001). On the other hand, seasonal movement of large herbivores might be simultaneously a strategy to reduce predation risk by moving beyond ranges of non-migratory predators (Fryxell et al. 1988; Rettie and Messier 2000). Yet, complete “escape” from predation will be uncommon, in part, because migration itself is often risky (Hebblewhite and Merrill 2009). Alternative strategies, such as migratory and non-migratory, can occur within the same species and within the same population, due to environment fluctuation and individual difference (Luccarini et al. 2006).
The effects of migration have been widely discussed (Albon and Langvatn 1992; Mysterud et al. 2001; Boyce et al. 2003; Hebblewhite and Merrill 2007; Hebblewhite et al. 2008; Jarnemo 2008; Hebblewhite and Merrill 2009), and from a biological view, they have been described as a positive effect to ecosystems, resulting in a more reasonable spread impacts from grazing and increasing ecosystem resilience (Fryxell and Sinclair 1988). On the other hand, seasonal migration noticeably hampers harvest and management of a population resulting in overexploitation of migrants (Jarnemo 2008; Bolger et al. 2008) with negative consequences on population dynamics (Ginsberg and Milner-Gulland 1994; Forsyth 1999; Mysterud et al. 2002).
The aim of this paper is to analyse first data on red deer spatial behaviour in the Carpathians, because no data are yet available from this area; however, some preliminary results have been presented in the past (Kropil et al. 2005, 2009) and in general, very few related studies have been carried out in this region (Okarma 1984, 1991; Nowak et al. 2005; Smietana 2005). In particular, the aims of this study are as follows: (i) to present first data on home range sizes at two temporal scales (annual and seasonal) of 20 male red deer monitored for several consecutive years in the Western Carpathians using three home range-estimating methods; (ii) to show distinct patterns of spatial behaviour, i.e. residential and migratory, in the same red deer population evaluating horizontal and vertical movements and (iii) to investigate possible ecological influences on the male home range size and to discuss possible management implications.
Material and methods
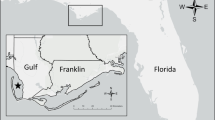
The study was carried out in temperate forests of the Western Carpathians in the area of Kremnica Mountains (Fig. 1). The area of 627 km2 is morphologically variable and has a mountainous climate with an altitudinal range of 350–1315 m a.s.l. which determines two climate subtypes: the rather humid north with cold mountainous climate, average temperatures −7 °C in January and +13 °C in July, annual precipitations 1000–1400 mm, snow depth 60–100 cm and duration of snow cover 160 days; and moderate warm mountainous climate on southern slopes with average temperature −5 °C in January and +18 °C in July, annual precipitations 650–850 mm, snow depth 30–60 cm and 110 days of snow cover. The area has a mosaic structure with 65 % of forests (73 % deciduous, 27 % coniferous), meadows and agricultural land surrounding human settlements. The area is dominated by beech (Fagus sylvatica, 37 %), followed by oak (Quercus spp., 19 %), spruce (Picea abies, 11 %), hornbeam (Caprinus betulus, 10 %), pine (Pinus sylvestris, 7 %) and fir (Abies alba, 7 %). The area is intensively managed from a forestry and hunting perspective; however, there are several natural preserves within the area with restricted management. The red deer is the main game species in Kremnica Mountains, and during this study, its population was estimated to be 2200 individuals in the area. Other wild ungulates living in the area are roe deer (Capreolus capreolus) with a population reaching 2100 individuals and wild boar (Sus scrofa) with 1100 individuals. There is also the constant presence of three large carnivores, namely brown bear (Ursus arctos), wolf (Canis lupus) and lynx (Lynx lynx).

Red deer study area in Slovakia (Kremnica Mountains) and locations collected from 20 male red deer between 2005–2013
This study was conducted from February 2005 to April 2013. A total of 20 male red deer were darted on feeding stations during winter, aged by tooth eruption, ear-tagged, radio-collared and released. Three animals were caught in 2005, five in 2006, five in 2008, two in 2009, one in 2010 and four in 2012 (see Table 1) and were monitored for 6 to 62 months (417 months in total, 20 months per 1 animal). Animals were tracked once per 9 days on average and 1457 locations were collected during the study.
All animals were fitted with ATS M2600 radio-collars with transmitters set on the 150 MHz waveband and surveyed with receiver (ATS FM-100) and three element hand-held Yagi antennas. The location of a radio-collared animal was estimated from bearings obtained from at least three reference points (White and Garrott 1990) with the “loudest signal” technique (Springer 1979). The actual location was obtained with triangulation software Locate III providing 95 % confidence ellipse of the measure (Nams 2006). Accuracy of fixes was determined as the radius of a circle with an area equal to 95 % confidence ellipse area from all fixes (±140 m).
Annual home ranges were estimated using three commonly used methods: Minimum Convex Polygon (MCP, Mohr 1947) using 100 % of locations, Kernel Home Range (KHR, Worton 1989), and Local Convex Hull (LoCoH, Getz and Wilmers 2004), using 90 % isopleths as recommended by Börger et al. (2006). Because LoCoH was considerably demanding on sufficient number of fixes, seasonal home ranges were estimated using MCP 100 % and KHR 90 %. All three methods were used because this procedure gives a more complete and widely comparable picture of red deer space use (Davini et al. 2004; Luccarini et al. 2006). Emigrants were not included in the statistical analysis of annual home ranges and migration, and their movement is only verbally described (Animal ID 504 started dispersion after collar failure and during monitoring was considered resident). Animals monitored less than 10 months (IDs 2611, 294, 324, 650 and 596) were also excluded from estimations of annual home ranges, and the resulting 14 individuals and 26 annual ranges were used for analysis. Seasonal home ranges were estimated using only animals with a completed season (for MCP 100 %) and with more than 16 fixes per animal in a particular season (for KHR 90 %).
Home range size was determined for the entire year (annual range: from May to April following year) and for three seasons determined as follows: summer = May to August, rut (mating season and the recovery time after rut) = September to November, winter = December to April; synchronised with natural seasonal cycles of the species in this environment. In order to avoid risk of including fixes from different seasons, seasonal home ranges of migratory animals were classified according to migration date between two seasonal ranges.
Movement strategies were determined following the definition of Zweifel-Schielly et al. (2009): resident, the distance between centroids of seasonal home ranges is less than 3 km; migrant, the distance between centroids of seasonal home ranges is more than 3 km with repeated seasonal return; emigrant, long-distance consecutive movements of young animals without repeated return. Migration was evaluated in both directions, i.e. horizontal and vertical. Horizontal migration was defined as Euclidean distance between centroids of succeeding seasonal home ranges. Vertical movement was evaluated according to altitudes of locations within the particular seasonal home range for each animal. Timing of migration in the migratory segment of the red deer population was calculated from dates of fixes between two seasonal ranges. Seasonal overlaps were calculated from seasonal MCPs recorded in one animal over the subsequent years (summer over summer, etc.). The largest recorded home range for animal and season was used as reference home range (Davini et al. 2004).
MCP 100 % was calculated by spatial analyst tool in ArcGIS 10 (ESRI 2011), KHR 90 % was calculated in Geospatial Modelling Environment (Beyer 2012) and LoCoH 90 % was calculated using the AdehabitatHR package of the R (R Development Core Team 2009, R version 3.0.1). Statistical analysis was also performed with R. Normality was checked using a Shapiro-Wilk test. Taking into account repeated measures of the same individual in different years and in order to avoid pseudoreplication, a linear mixed-effects model (LME) was fitted specifying subjects (i.e. deer identity) and explanatory variables (i.e. age…). ANOVA was used to test the significance of explanatory variables. The significance level was set at P ≤ 0.05 in all tests.
Results
Annual and seasonal home ranges
Three different movement strategies were identified in the sample: residents (10 individuals), migrants (8 individuals) and emigrants (2 individuals) with a ratio residents:migrants = 1:1.25 (emigrants omitted). Animal ID 504 emigrated from the area after collar failure, however, during monitoring displayed typically residential behaviour and thus was considered to be a resident in the analysis. Methods used for estimation of annual home ranges gave considerably different results. MCP and KHR estimates were larger than LoCoH ones (t = 2.904, P = 0.007 using MCP; t = 4.675, P < 0.001 using KHR). A total of 29 annual home ranges were estimated (2 in 2005, 4 in 2006, 5 in 2008, 6 in 2009, 5 in 2010, 2 in 2011 and 5 in 2012). Residents had significantly smaller annual home range than migrants (Table 2). In particular, LME revealed a significant difference between red deer adopting two different strategies using MCP 100 % (F = 6.809, P = 0.021) and KHR 90 % (F = 8.573, P = 0.011). LME also indicated that residents expanded their home range size with increasing age (F = 5.127, P = 0.045 using MCP 100 %; F = 5.436, P = 0.040 using LoCoH 90 %), while no age effect was seen in migrants (F = 0.223, P = 0.645 using MCP; F = 0.143, P = 0.712 using KHR; F = 0.007, P = 0.933 using LoCoH). Annual home ranges did not differ over the 8 years of the study (F = 0.063, P = 0.806 using MCP 100 %; F = 0.426, P = 0.526 using KHR 90 %; F = 0.229, P = 0.641 using LoCoH 90 %).
Although the average seasonal home range in migrants was larger than in residents over the three seasons, the analysis did not reveal significant differences in size between the two movement strategies (F = 2.019, P = 0.176 for MCP; F = 1.217, P = 0.205 for KHR 90 %). Surprisingly, residents had comparable summer and rut home ranges but significantly expanded winter home ranges (t = 2.383, P = 0.021) using MCP 100 %. KHR 90 % revealed expansion of winter home ranges even in both movement strategies (Table 3).
Migration movements
The analysis of migration distances between seasonal ranges revealed the use of different areas for migratory red deer (F = 15.479, P = 0.001; Table 3). While migrants underwent considerably long movements between seasonal home ranges (8014 ± 1285 m), residents showed only a marginal seasonal shift in space use (1463 ± 1665 m; Table 4).
Interesting to note is that, in spite of the long distances between seasonal home ranges in migratory red deer, the site fidelity to these areas over the years was comparable to that recorded for residents. In fact, analysing the data of animals monitored for more than 1 year in particular season, migratory seasonal overlap (47 ± 11 %) and residential one (56 ± 13 %) did not differ significantly, and migration strategy had no effect on fidelity (F = 0.4985, P = 0.500). Home range overlap tended to increase throughout the year (F = 4.985, P = 0.013) with a significant increase in the winter (t = 2.24, P = 0.035; Table 4).
Vertical movement throughout the year was significantly influenced by season. LME clearly confirmed that seasonal altitudes are a function of time (F = 15.111, P < 0.001). Both migratory and residential red deer reached their highest altitudes in June and July, while the lowest altitudes were recorded during January and February (Fig. 2). In particular, analysing altitudes of fixes within each seasonal home range, both red deer strategies showed a significant descent in winter months (t = −3.990, P < 0.001 in migrants; t = −3.097, P = 0.003 in residents) compared to their summer and rut altitudes. Interestingly, migrants showed a significant movement to lower, distinct and relatively hospitable conditions than residents (t = −3.520, P = 0.003).

Monthly average altitude of resident, migrant and emigrant red deer recorded in Kremnica Mountains during 2005–2013 (pooled across years)
Summer migration occurred on 5th May (±11 days), migration to mating areas occurred on 12th September (±3 days) and migration to wintering areas took place on 13th November (±6 days).
Discussion
The present work describes two distinct movement strategies of red deer, i.e. migratory and residential, in the Western Carpathians. The differences in annual home range sizes between migrant and resident were significant and seasonal changes in the size and altitude of the home ranges were also proven. While MCP and KHR gave comparable results, LoCoH significantly differed from both, probably as consequence of small sample size. The use of several methods allowed wider comparison with other studies and provided a more complete picture of red deer spatial behaviour. The home range size of red deer varied considerably as several other studies indicate (Table 5) and emphasize the significance of the particular environment (Georgii and Schröder 1983). In lowlands with no presence of predators, red deer tend to exhibit non-migratory behaviour (Nahlik et al. 2009; Kamler et al. 2008), while in mountainous regions with strong seasonal spatial variation of critical resources, red deer tend to migrate regularly (Luccarini et al. 2006; Zweifel-Schielly et al. 2009), which is consequently reflected in home range size. Luccariny et al. (2006) reported relatively smaller annual home ranges in the Italian Alps compared to those recorded in Slovakia, possibly due to summing both sexes in calculations, because females commonly use smaller ranges than males (Georgii 1980; Kamler et al. 2008). To the contrary, red deer occupying flat landscapes in Hungary surprisingly displayed use of wide ranges reaching double the size of those in Kremnica Mountains (Szemethy et al. 1998).
Movement strategies seem to be slightly unequally distributed in the population with a M:R ratio of 1:1.25. This ratio obviously correlates with topographic diversity of the given area. For example, while in Hungary, red deer do not exhibit migration movements (Nahlik et al. 2009), Luccarini et al. (2006) reported a M:R ratio of 1:1 in Tarvisio forest and 1.25:1 in Sussa Valley while Zweifel-Schielly et al. (2009) reported 9:1 in the Swiss Alps.
Annual home range size was confirmed to increase with age of the animal related to potential dominance and growing nutritional requirements (Clutton-Brock et al. 1982; Mysterud et al. 2001). This conforms with other studies describing positive age effects on annual home range size in large herbivores (Georgii and Schröder 1983; Szemethy et al. 1998; Cederlund and Sand 1994).
The size of winter home range in residents was significantly larger compared to other seasons, while season had no effect on home range size in migrants. Seasonal areas used by migrants did not differ in size across all seasons. Residents used comparable ranges during summer and autumn, while significantly expanding their space use in winter. This finding does not fit the general concept that climatic conditions influence ungulate spatial behaviour by reducing winter home range size as reported in red deer (Georgii and Schröder 1983; Koubek and Hrabe 1996), roe deer (Mysterud 1999; Rivrud et al. 2010), moose (Phillips et al. 1973; Thompson and Vukelish 1981) and ibex (Parrini et al. 2003). However, Anderson et al. (2005) reported that elk home ranges increased in winter and attributed this to predator pressure of wolves, population densities and reduced quantity and quality of forage. It is also known that quantity of forage negatively influences the home range size, while quality positively influences the size of the range (Mysterud et al. 2001). This might be seen in Kremnica Mountains, given that the topography of the area probably causes an increase of density in the southern parts used as wintering refugees. Consequent intensive browsing reduces availability of woody browse and forces animals to search for high quality forage provided by supplementary winter feeding widespread over the area. Moreover, Kamler et al. (2008) suggested that periodic presence of predators hunting in particular areas may result in an increase of overall movement to avoid excessive predation.
Seasonal site fidelity between years was remarkable and tended to increase during the year in both, migrants and residents. Interestingly, irrespective of long distances between summer and winter areas, migrants showed site fidelity comparable to residents. Overlaps tended to increase during the year with maximum during winter as also shown by Luccarini et al. (2006). Strong seasonal and annual site fidelity in cervids is common (Rudd et al. 1983; Edge et al. 1985) because fidelity likely carries advantages including knowledge of forage resources and security cover (Webb et al. 2011).
The longest dispersal movement was recorded in young stags emigrating to neighbouring mountain ranges. Animal ID 233 moved 30 km east to neighbouring Polana Mountains, animal ID 683 moved 15 km west to Vtacnik Mountains for winter and then travelled another 50 km reaching Tribec Mountains (65 km in total). One year after collar failure, animal ID 504 was identified according to ear tag in Velka Fatra Mountains 23 km north from the last recorded position and in the same year was culled another 24 km north in the same mountain region (47 km in total). Jarnemo (2008) reported similar 47 km movement of red deer in southern Sweden. All emigrants started long-distance movements at a young age (2–3 years), which conforms with the findings of Georgii and Schröder (1983) that stags left the “motherland” after 2.5 years. Dispersal behaviour in males may occur due to local resource competition “pulling” individuals to low-density areas and/or local mate competition “pushing” young males to leave the natal range (Loe et al. 2009).
Significant descent during winter was observed in both movement strategies; however, vertical differences in home range altitudes were most obvious in migrants. While residents simply increased the use of areas at lower altitudes within the same area as they used during summer, migrants displayed periodic vertical movement from high summer altitudes to distinct wintering areas in southern parts of the mountain range. Although Luccarini et al. (2006) reported migrants moving significantly higher in summer compared to residents in Italian Alps, the topography and moderate mountainous climate of the Kremnica Mountains caused that both, migrants and residents remained in comparable altitudes during summer while migrants travelled downhill in winter. This is in contrary with the general concept that migratory animals take advantage of high altitudes while residents remain in the low altitudes during summer as seen in Alps (Luccarini et al. 2006; Zweifel-Schielly et al. 2009). The possible explanation of residents remaining in considerably higher altitudes during winter than migrants is higher availability of forage. Research in the area showed that winter forage availability was lower on wintering areas, mostly including meadows and agricultural land, compared to vertically well-differentiated forested habitats in higher altitudes providing access to woody browse, the main source of natural forage for red deer during winter. Moreover, winter logging in the area can also temporally provide additional highly nutritious forage of sufficient abundance. Also, comparing absolute altitudes of wintering residents in Kremnica Mountains (800 m a.s.l.) with those in Alps (1100 m a.s.l., Luccarini et al. 2006), it is clear that red deer can easily survive in “high” altitudes of Kremnica Mountains. Why do migrants move downhill? There is a general agreement that downhill migration is a strategy to avoid harsh winter conditions at high altitudes because snow cover reduces access to forage, increases the energetic cost of movement (Parker et al. 1984) and exposes red deer to higher predation risk (Cederlund and Lindström 1983). And regular presence of wolves in the area may also cause that migrants adopted large scale strategy to avoid predation by moving closer to humans and occupying poorer forage resources, while residents increased winter movement activity at finer scale (Table 3).
The findings of this study emphasize the need for coordination of management effort and objectives at a large scale, since almost half of the population seems to be migratory moving across distinct landscape units and landholdings. Considering the minimum extent of the hunting ground in Slovakia to be 2000 ha (Garaj and Kropil 2013), there is a logical demand of more complex approach in order to take into account migratory animals occupying multiple size of this area. The Hunting Act 274/2009 for the first time established large-scale game management in Slovakia, establishing hierarchical coordination of hunting units over the geographic ranges considered as compact hunting areas with identical management objectives. Considering migration distances, the smallest area for unified management should be at least 300 km2. The coordination at the landscape scale is aimed to prevent contradictory management within the same population. However, 74 % increase of red deer population in Slovakia from 2000 to 2011 correlating with growing damages of young forest stands can be considered as an indicator of contradictory management in the past decade. Authors of the new concept of hunting management in terms of damages of young forest stands (Konopka and Kastier 2013) concluded that red deer population in 2011 was 57 % higher than its normative numbers and suggested to decrease the population gradually from current 58,100 individuals to normative 32,900 by 2025. Given the strong fidelity to seasonal ranges, it would be also useful to identify wintering areas within the mountain ranges and suitably adjust hunting quotas for these areas. A temporal solution to mitigate this conflict might be supplementary winter feeding historically spread over the whole Carpathians and recently obligatory practice given by law in Slovakia, Ukraine and Romania. However, the effect of supplementary feeding is still unclear and results from related studies are often equivocal (Putman and Staines 2004). Some authors even consider this practice as negative, causing substantial browsing deterioration in forests (Luccarini et al. 2006). But feeding experiments under controlled conditions (Rajsky et al. 2008) indicate that correctly designed supplementary winter feeding could be used as a tool for effective game damage control. However, data in this study confirmed that migration is a very important component of resilient and sustainable ecosystems and its understanding is the key to a future successful management.
References
Albon SD, Langvatn R (1992) Plant phenology and the benefits of migration in a temperate ungulate. Oikos 65:502–513
Anderson DP, Forester JD, Turner MG, Frair JL, Merrill EH, Fortin D, Mao JS, Boyce MS (2005) Factors influencing female home range sizes in elk (Cervus elaphus) in North America landscape. Landsc Ecol 20:25–271
Beyer HL (2012). Geospatial Modelling Environment (Version 0.7.2.0). (software). URL: http://www.spatialecology.com/gme.
Bolger DT, Newmark WD, Morrison TA, Doak DF (2008) The need for integrative approaches to understand and conserve migratory ungulates. Ecol Lett 11:63–77
Börger L, Franconi N, De Michelle G, Gantz A, Meschi F, Manica A, Lovari S, Coulson T (2006) Effects of sampling regime on the mean and variance of the home range size estimates. J Anim Ecol 75:1393–1405
Boyce MS, Mao JS, Merrill EH, Fortin D (2003) Scale and heterogenity in habitat selection by elk in Yellowstone National Park. Ecoscience 10:421–431
Carranza J, Hidalgo de Trucios SJ, Medina R, Valencia J, Delgado J (1991) Space use by red deer in a Mediterranean ecosystem as determined by radio-tracking. Appl Anim Behav Sci 30:363–371
Catt DC, Staines BW (1987) Home range use and habitat selection by red deer (Cervus elaphus) in a Sitka-spruce plantation as determined by radio-tracking. J Zool 211:681–693
Cederlund G, Lindström E (1983) Effects of severe winters and fox predation on roe deer mortality. Acta Theriol 28:129–145
Cederlund G, Sand H (1994) Dispersal of subadult moose (Alces alces) in a nonmigratory population. Can J Zool 70:1309–1314
Clutton-Brock TH (1989) Mammalian mating system. Proc R Soc Lond 236:339–372
Clutton-Brock TH, Harvey PH (1978) Mammals, resources and reproductive strategies. Nature 273:191–195
Clutton-Brock TH, Guinness FE, Albon SD (1982) Red deer: behaviour and ecology of two sexes. Edinburgh University Press, Edinburgh
Davini S, Ciuti S, Luccarini S, Apollonio M (2004) Home range patterns of male fallow deer Dama dama in a sub-Mediterranean habitat. Acta Theriol 49:393–404
Edge WD, Marcum CL, Olson SL (1985) Effects of logging activities on home-range fidelity of elk. J Wildl Manag 49:741–744
ESRI (2011) ArcGIS Desktop: release 10. Environmental Systems Research Institute, Redlands
Forsyth DM (1999) Long-term harvesting and male migration in a New Zeland population of Himalayan tahr Hemitragus jemlahicus. J Appl Ecol 36:351–362
Fryxell JM, Sinclair ARE (1988) Causes and consequences of migration by large herbivores. Trends Ecol Evol 3:237–241
Fryxell JM, Greever J, Sinclair ARE (1988) Why are migratory ungulates so abundant? Am Nat 131:781–798
Garaj P, Kropil R (2013) Game management. Technical University Press, Zvolen
Georgii B (1980) Home range patterns of female red deer (Cervus elaphus) in the Alps. Oecologia 47:278–285
Georgii B, Schröder W (1983) Home range and activity patterns of male red deer (Cervus elaphus) in the Alps. Oecologia 58:238–248
Getz WM, Wilmers CC (2004) A local nearest-neighbor convex hull of home ranges and utilization distributions. Ecography 27:489–505
Ginsberg JR, Milner-Gulland EJ (1994) Sex biased harvesting and population dynamics in ungulates: implications for conservation and sustainable use. Conserv Biol 8:157–166
Harestad AS, Bunell FL (1979) Home range and body weight—a reevaluation. Ecology 60:389–402
Hebblewhite M, Merrill EH (2007) Multiscale predation risk for elk: does migration reduce risk? Oecologia 152:377–387
Hebblewhite M, Merrill EH (2009) Trade-offs between predation risk and forage differ between migrant strategies in a migratory ungulate. Ecology 90:3445–3454
Hebblewhite M, Merrill E, McDermid G (2008) A multi-scale test of forage maturation hypothesis in a partially migratory ungulate population. Ecol Monogr 78:141–166
Jarnemo A (2008) Seasonal migration of male red deer (Cervus elaphus) in southern Sweden and consequences for management. Eur J Wildl Res 54:327–333
Kamler JF, Jendrzejewski W, Jendrzejewska B (2008) Home ranges of red deer in a European old growth forest. Am Midl Nat 159:75–82
Konopka J, Kastier P (2013) New concept of hunting development in Slovakia and damages caused by ungulates on forest stands. Proceedings of 22ndInternational Conference on Actual Problems in Forest Protection. 2009 April 25–26, Novy Smokovec, Slovakia
Koubek P, Hrabe V (1996) Home range dynamics in the red deer (Cervus elaphus) in a mountain forest in Central Europe. Folia Zool 45:219–222
Kropil R, Garaj P, Garaj P, Pataky T (2005) Spatio-temporal activity of red deer in study area of VSLP of TU in Zvolen. Proceedings of 19th International Conference on Game Management and Game Protection. 20th May 2005, Zvolen, Slovakia
Kropil R, Ostrihon M, Kastier P (2009) GPS telemetry of red deer (Cervus elaphus): first results from Slovakia. Proceedings of 23rd International Conference on Game Management and Game Protection. 21st May 2009, Zvolen, Slovakia
Lindstedt SL, Miller BJ, Buskirk SW (1986) Home-range, time, and body size in mammals. Ecology 67:413–418
Loe LE, Mysterud A, Veiberg V, Langvatn R (2009) Negative density-dependent emigration of males in an increasing red deer population. Proc Biol Sci 276:2581–2587
Luccarini S, Mauri L, Lamberti P, Apollonio M (2006) Red deer (Cervus elaphus) spatial use in the Italian Alps: home range patterns, seasonal migrations, and effect of snow and winter feeding. Ethol Ecol Evol 18:127–145
McNab BK (1963) Bioenergetics and the determination of home range size. Am Nat 97:133–140
Mohr CO (1947) Table of equivalent populations of North American small mammals. Am Midl Nat 37:223–249
Mysterud A (1999) Seasonal migration pattern and home range of roe deer (Capreolus capreolus) in an altitudinal gradient in southern Norway. J Zool 247:479–486
Mysterud A (2013) Ungulate migration, plant phenology, and large carnivores: the times they are a-changin’. Ecology 94:1257–1261
Mysterud A, Coulson T, Stenseth NC (2002) The role of males in the dynamics of ungulate populations. J Anim Ecol 71:907–915
Mysterud A, Langvatn R, Yoccoz NG, Stenseth NCH (2001) Plant phenology, migration and geographical variation in body weight of a large herbivore: the effect of variable topohraphy. J Anim Ecol 70:915–923
Nahlik A, Sandor G, Tari T, Kiraly G (2009) Space use and activity patterns of red deer in highly forested and in a patchy forest-agricultural habitat. Acta Silv Lign Hung 5:109–118
Nams VO (2006) Locate III Users’ Guide. Pacer Computer Software, Tatamagouche
Nowak S, Myslajek RW, Jendrzejewska B (2005) Patterns of wolf Canis lupus predation on wild and domestic ungulates in the Western Carpathian Mountains (S. Poland). Acta Theriol 50:263–276
Okarma H (1984) The physical condition of red deer falling a prey to the wolf and lynx and harvested in the Carpathian Mountains. Acta Theriol 29:283–290
Okarma H (1991) Marrow fat content, sex and age of red deer killed by wolves in winter in the Carpathian Mountains. Ecography 14:169–172
Parker KL, Robins CT, Hanley TA (1984) Energy expenditure for locomotion by mule deer and elk. J Wildl Manag 48:474–488
Parrini F, Grignolio S, Luccarini S, Bassano B, Apollonio M (2003) Spatial behaviour of adult male Alpine ibex Capra ibex ibex in the Gran Paradiso National Park, Italy. Acta Theriol 48:411–423
Phillips DH, Berg WE, Sniff DB (1973) Moose movement patterns and range use in southwestern Minnesota. J Wildl Manag 37:266–278
Putman RJ, Staines BW (2004) Supplementary winter feeding of wild red deer Cervus elaphus in Europe and North America: justifications, feeding practice and effectiveness. Mammal Rev 34:285–306
Rajsky M, Vodnansky M, Hell P, Slamecka J, Kropil R, Rajsky M (2008) Influence of supplementary feeding on bark browsing by red deer (Cervus elaphus) under experimental conditions. Eur J Wildl Res 54:701–708
R Development Core Team (2009) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org.
Rettie WJ, Messier F (2000) Hierarchical habitat selection by woodland caribou: its relationship to limiting factors. Ecography 23:466–478
Rivrud IM, Loe LE, Mysterud A (2010) How does local weather predict red deer home range size at different temporal scales? J Anim Ecol 79:1280–1295
Rudd WJ, Ward AL, Irwin LL (1983) Do split hunting seasons influence elk migrations from Yellowstone National Park? Wildl Soc B 11:328–331
Schmidt K (1993) Winter ecology of nonmigratory red deer. Oecologia 95:226–233
Schmidt K, Gossow H (1991) Winter ecology of alpine red deer with and without suplementary feeding: management implications. In Csáni S, Ernhaft J (eds) Transactions of the XXth Congress of the International Union of Game Biologists: Part 1. University of Agricultural Sciences, 1991 21st–26th August, Hungary
Smietana W (2005) Selectivity of wolf predation on red deer in the Bieszczady Mountains, Poland. Acta Theriol 50:277–288
Springer JT (1979) Some sources of bias and sampling error in radio triangulation. J Wildl Manag 43:926–935
Szemethy L, Heltai M, Matrai K, Peto Z (1998) Home ranges and habitat selection of red deer (Cervus elaphus) on a lowland area. Congress of the International Union of Game Biologists, 1st September 1997. Lyon France
Thompson ID, Vukelish MF (1981) Use of logged habitats in winter by moos cows with calves in northeastern Ontario. Can J Zool 59:2103–2114
Van Beest FM, Rivrud IM, Loe LE, Milner JM, Mysterud A (2011) What determines variation in home range size across spatiotemporal scales in a large browsing herbivore? J Anim Ecol 80:771–785
Vincent JP, Bideau E, Hewison JM, Angibault JM (1995) The influence of increasing density on body weight, kid production, home range and winter grouping in roe deer (Capreolus capreolus). J Zool 236:371–382
Webb SL, Dzialak MR, Harju SM, Hayden-Wing LD, Winstead JB (2011) Influence of land development on home range use dynamics of female elk. Wildl Res 38:163–167
White GC, Garrott RA (1990) Analysis of wildlife radio-tracking data. Academic Press Inc, San Diego
Worton BJ (1989) Kernel method for estimating the utilization distribution in home range studies. Ecology 70:164–168
Zweifel-Schielly B, Kreuzer M, Ewald KC, Suter W (2009) Habitat selection by an Alpine ungulate: the significance of forage characteristics varies with scale and season. Ecography 32:103–113
Acknowledgments
This paper was produced within the project VEGA 1/1210/12 supported by the Ministry of Education, Science, Research and Sport of the Slovak Republic and the projects “Centre of Excellence: Adaptive Forest Ecosystems” (ITMS 26220120006) and “Completing the Centre of Excellence: Adaptive Forest Ecosystems” (ITMS 26220120049) supported by the Operational Programme Research and Development within European Regional Development Fund. We thank the University of Veterinary Medicine in Košice, especially Dr. J. Ciberej, the Animal Production Research Centre in Nitra, especially Dr. M. Jurčík and Dr. D. Rajský from the Technical University in Zvolen for supervising the immobilizations. We also thank the University Forest Enterprise for the support and assistance during the study. We thank all dedicated field assistants T. Pataky, M. Ostrihoň, P. Garaj Jr., M. Rajský and Ľ. Ferlica for collecting data, and the land owners and users for allowing telemetry monitoring in their landholdings. We also thank the anonymous referees for their valuable comments and advices.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Gortázar
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Kropil, R., Smolko, P. & Garaj, P. Home range and migration patterns of male red deer Cervus elaphus in Western Carpathians. Eur J Wildl Res 61, 63–72 (2015). https://doi.org/10.1007/s10344-014-0874-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-014-0874-4