
Abstract
Small-scale private forests cover large areas in Europe and often contain structures and habitats of high nature conservation value that are increasingly put under pressure due to a rising interest in fuelwood and wood products. We investigate the distribution of variables like living tree and deadwood volumes, management intensity, diversity and density of tree-related microhabitats recorded in 81 small-scale private forest parcels in the Lower Saxon Hills (northwest Germany). We provide an assessment of the influence of predictors like the individual forest owners’ goals and activities, as stated by them in a quantitative survey, as well as landscape parameters like parcel size, slope, landscape fragmentation and forest continuity. Our results indicate that there are two types of structures of conservation value in small-scale private forests: slowly evolving structures (type A) like large-diameter living trees and tree-related microhabitats which mostly depend on landscape parameters that act on longer time scales, and fast evolving structures (type B) like deadwood that are influenced by both owner attitudes and landscape parameters. The resulting implications for integrative forest and conservation management are discussed. When considering the conservation of the slowly evolving type A structures, long-term commitments to conservation legislation, financial incentives and generation-spanning education of forest owners are necessary. Efforts to promote the faster evolving type B structures might prove particularly advantageous in small-scale private forests given the structural diversity of the stands, but also the often strong identification of owners with their land.
Graphical abstract

Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
When it comes to an effective conservation policy in forest landscapes, all types of ownership must be considered and integrated. This is especially true for landscapes with mixed ownership and a high proportion of privately owned forest land (Fischer et al. 2019; Loeb and D’Amato 2020). In Germany, half of the country’s forested area is privately owned, distributed equally among owners possessing at least 20 ha, and small-scale private forest (SPF) owners with forest parcels smaller than 20 ha (BMEL 2014). While forestry in owner groups with larger properties (state and federal forests, communal forests, and larger private properties) has become more mechanized and commercially oriented in the past, SPF owners have often been less inclined to harvest timber from their forests. Furthermore, many SPFs in Germany are of very small sizes (average private forest parcel size of 3 ha) (BMEL 2014), often precluding economic use, especially by heavy machinery, and leading to significantly different forest structures in SPFs compared to other ownership types (Mölder et al. 2021). Most notably, SPFs have been shown to be characterized by higher structural diversity (Maltamo et al. 1997; Rendenieks et al. 2015; Schaich and Plieninger 2013) and biodiversity (Torras et al. 2012). Certain structures and habitats that are considered to be of high nature conservation value have been shown to occur there more frequently, like tree-related microhabitats (TreMs) (Johann and Schaich 2016) and deadwood (Schaich and Plieninger 2013).
With changing economic parameters like increasing demand for fuelwood, intensified forest management can also be expected in many regions with a high share of SPF (UNECE 2022). Extension services offered by state and private forestry associations are also increasingly encouraging wood extraction from small parcels (Takala et al. 2022), even though this might be questionable considering the carbon impact (Norton et al. 2019; Sterman et al. 2022). Another aspect to consider are the consequences for habitat structures and the species that depend on them (Kjučukov et al. 2022), i.e., the nature conservation values of forest lands. TreMs have been recognized as valuable for a plethora of species groups (Asbeck et al. 2021a). They are relatively easy to survey and can be used as a proxy for the occurrence of species groups like saproxylic beetles, spiders, bats, birds, fungi and lichens (Martin et al. 2022). Many TreMs like different forms of cavities are known to develop over long time spans (Kõrkjas et al. 2021) and are therefore rare in many conventionally managed forests, where trees are mostly harvested before they can develop features like accumulated crown deadwood or tree cavities (Ranius et al. 2009). Due to lower management intensity, TreMs have a higher chance to develop in SPFs, but are now experiencing increased pressure with rising timber and particularly firewood extraction. This is particularly true for TreMs linked to large-diameter trees, which are important elements of conservation strategies (Kozák et al. 2023). Another feature of high importance to multiple forest functions and nature conservation value is deadwood, which improves forest soils, has a beneficial impact on water retention, and serves as medium-term carbon sink (Chivulescu et al. 2022; Parisi et al. 2018; Piaszczyk et al. 2021). Deadwood also provides the substrate and habitat for a range of organism groups, many of which are endangered and protected. Examples include bryophytes, lichens, fungi, invertebrates, small mammals and birds (Bujoczek et al. 2021). Similar to TreMs, deadwood supplies in SPFs and related biodiversity can be put under pressure with increasing forest management intensity.
SPF owners are a heterogeneous group of people with a diverse set of motivations and goals concerning their land, resulting in a variety of strategies when managing their forests (Tiebel et al. 2023; Westin et al. 2023). While SPF owners are often assumed to have predominantly economic interests, there is actually a large variety in motivations. A substantial group of SPF owners is strongly interested in nature and cultural heritage conservation (Weiss et al. 2019). Likewise, the management activities of SPF owners are diverse and may comprise a wide range from industrialized wood harvesting, logging by horses, manually extracting single small stems as firewood for personal consumption, or even ceasing all felling activities for the sake of nature conservation, or due to lack of interest or long distances to their property (Bieling 2004; Tiebel et al. 2021; Weiss et al. 2019).
Many structures of conservation value in forests take a long time to develop (Kõrkjas et al. 2021) or may depend on random, unpredictable events (breaking of stems or parts of the crown, injuries, colonization by animals; Larrieu et al. (2022)). Therefore, it seems not to be easy to relate structures and habitats to current motivations and management goals of forest owners. The time span during which individual forest owners actively influence their forests is often short compared to natural processes in the forest ecosystem, and long-term influences for example from landscape characteristics of the forest parcels (elevation, slope, water and soil parameters) or their surroundings (fragmentation or location in large, closed forests) may play an equally important role in the development of valuable structures (Bujoczek and Bujoczek 2022; Kapusta et al. 2020; Kennedy et al. 2008).
To our knowledge, the relationships between small-scale private forest owner goals and activities, and structures and habitats in their respective forest parcels have not been investigated yet. To address this research gap, we performed analyses in SPFs smaller than 5 ha in a model region, the Lower Saxon Hills in northwest Germany (see Fig. 1). We used the following data sets: (a) quantitative survey data about SPF owners’ goals and activities; (b) field data collected on their parcels (data on forest structure and nature conservation values); and (c) data on landscape parameters of the parcels to address the following research questions:
-
1.
What are the characteristics of the SPF parcels in our model region concerning stand structure variables and the distribution of valuable habitats?
-
2.
What are the respective influences of owner goals and activities, and landscape parameters on these variables?
-
3.
How can the possibly varying influences be explained, and which conclusions for integrative forest and conservation management in SPFs can be drawn?

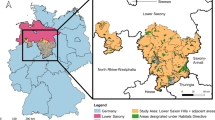
The study area is located in the federal state of Lower Saxony (Germany) and consists of a set of administrative units (dark blue) to closely resemble the Lower Saxon hills geographic region (hatched). Data: Lower Saxony Forest Planning Agency (NFP), GADM (https://gadm.org/)
Methods
Study area
Our study area is the Lower Saxon Hills geographic region (Fig. 1) in the German federal state of Lower Saxony, which consists of two discontiguous areas and includes adjacent administrative units (in total 14,228 km2). Due to the uniform legal situation and consistent governmental forest administration, eventual regional differences in the support and treatment of SPF owners are kept to a minimum in this study. The Lower Saxon Hills are 33% forested, with European beech (Fagus sylvatica, 35%), Norway spruce (Picea abies, 29%) and oaks (Quercus robur and Quercus petraea, 11%) as the dominant tree species. Natural vegetation would consist mostly of European beech, while the admixture of Norway spruce, oaks and other broadleaf (e.g., sycamore Acer pseudoplatanus, wild cherry Prunus avium, birch Betula pendula, hornbeam Carpinus betulus) and conifer (e.g., Scots pine Pinus sylvestris, Douglas fir Pseudotsuga menziesii) tree species is largely due to anthropogenic influence. In privately owned forest parcels (43% of the forested area), 22% of the area consists of parcels with a size between 5 and 20 ha, and 23% of the area is occupied by parcels of 5 ha or less (ML 2014). The distribution of forest ownership classes and small-scale structural diversity is comparable to many European countries (Mölder et al. 2021; Schmithüsen and Hirsch 2010), which allows for a transfer of our results.
The study area lies at the border between oceanic and humid continental climates (Köppen classifications Cfb and Dfb), with average annual precipitation on our research sites between 645 and 1,042 mm, and annual mean temperatures between 8.2 and 10.3 °C (DWD 2018a, 2018b). Bedrock is mostly Mesozoic limestone and sandstone, in part with a Quaternary loess cover, and occasionally evaporite (gypsum karst) (LBEG 2022). Despite the relatively high average rainfall, the study area has been affected by unusual heat and drought periods since 2018. This combined with severe storms in 2018 and 2020 led to increased damage by bark beetles and widespread tree mortalities in conifer stands (Rohde et al. 2021, 2019).
Owner attitudes and forest parcels
To obtain insights into the attitudes of SPF owners in the study area, we used data from a quantitative mail survey that was conducted among the members of three local private forest owner associations. We sent out questionnaires by post to all 4,204 members of these associations, and 1,671 persons (39.8%) returned usable answers by mail or online (Tiebel et al. 2021). For the present work, we re-contacted those participants who had provided their contact details. We asked for the location of their largest forest parcel and for their approval to carry out field work. This call resulted in 91 answers, of which 81 provided useful geographic information about forest parcels (Fig. 1). These parcels range in size from 0.1 to 3.8 ha (mean: 1.21 ha, median: 1.01 ha, SD: 0.89 ha).
Owner goals concerning their forests were queried in the questionnaire with a list of 16 possible goals, with the possibility to rate their respective influence on a five-point Likert scale (details given in Appendix A1). These goals were then categorized into four types (Tiebel et al. 2021), namely goals referring to the promotion of certain ecosystem services (regulating, cultural and provisioning), and purely financial goals (Table 1). For each goal type consisting of several sub-goals, a value of agreement between 0 and 1 was calculated. The forest management activities of SPF owners (Table 1) were captured using a checklist of 26 possible choices and categorized into five activity types (1. classical silvicultural, 2. close-to-nature silvicultural, 3. traditional silvicultural, 4. active conservation, and 5. passive conservation activities). Agreement with each type of activity was again expressed as a fractional value between 0 and 1.
Landscape parameters
Based on the literature and expert knowledge, we identified landscape parameters (Table 1) that were expected to influence forest structure and the distribution of valuable habitats. Elevation, slope and aspect of the individual parcels were derived from a digital terrain model (LGLN 2022). We used data about water availability (given on a scale from 0 = dry to 3 = wet) and nutrient availability (0 = oligotrophic, 1 = mesotrophic, 2 = eutrophic) to further characterize the parcels (LBEG 2022). Connection to infrastructure was expressed by the distance to the nearest paved road (OpenStreetMap, 2022). Corine land cover data (European Environment Agency 2018) was used to calculate the proportion of open land surrounding a parcel within a radius of 2 km (Verga et al. 2021); this value was pre-selected from a series of logarithmically scaled possible radii of 50–5000 m, based on correlations of the predictor with the response variables (Jackson and Fahrig 2015). The percentage of each forest parcel border that coincides with open land was calculated from Corine data. We used data about the presence of forests in Lower Saxony on historical maps (NFP 2010) to classify the parcels as either recent forest, or ancient forest (more than 200 years of continuous forest cover) of broadleaf, conifer or mixed character, respectively.
Forest structure and valuable habitats
Forest structural data (Table 2) were collected on circular plots of 500 m2 (r = 12.62 m), generally following the methodology in Meyer and Fricke (2018). Parcels below 1.5 ha were equipped with one plot, larger plots received two (below 3 ha) or three plots. The center of a single plot was placed at the centroid of the forest parcel in a Geographic Information System (with some leeway for concavely shaped parcels), while two or three plots were placed to provide the maximum distance from one another and the border of the parcel. Standing trees (living or dead) with a diameter at breast height (dbh) of at least 7 cm were recorded with dbh and tree species. For each tree species and each plot, nine height measurements were taken if possible from trees in three different height classes using a handheld laser rangefinder (LTI TruPulse 360B). Stumps with a diameter of at least 7 cm and lying deadwood with a diameter of at least 20 cm were recorded, too. Deadwood was classified as either broadleaf or conifer deadwood and as sawn and non-sawn. In addition, the stage of decay (four classes: 1 = freshly dead to 4 = identifiable, but completely soft) was determined.
After data collection on the plots, the whole forest parcel was assessed for disturbances (for example, excessive logging trails, signs of recreational use, storm damage; each classified on a scale from 0 = not present, to 3 = strong influence). Trees of at least 50 cm dbh were recorded and assessed for TreMs following the field key by Larrieu et al. (2018) to calculate TreM density (per ha), the abundance of trees with TreMs (per ha), and the abundance of trees with a dbh ≥ 50 cm (per ha). For each forest parcel, the prevailing stand age was classified into 25-year classes (0–25 yrs, 26–50 yrs, and so on) by expert estimation in the field. Height and dbh data from the trees with measured heights were used to calculate individual Näslund height curves for each tree species on each plot (Mehtätalo et al. 2015) in order to interpolate the heights of the remaining trees from dbh measurements alone. Wood volumes were calculated from dbh and height data using species-specific form factors. Accumulated harvested volumes were calculated from stump data: To this aim, stump diameters were first extrapolated to dbh using a uniform tapering value of 1 cm/m, and then standard form factors were applied. Harvested volumes therefore constitute an integral over the stump persistence time of the wood volume that was taken from the forest.
To quantify human influence on forest development, we calculated the forest management intensity index (ForMI) (Kahl and Bauhus 2014). Non-native tree species as used in the context of the ForMI were understood to also encompass species planted outside their natural range, for example Picea abies (Norway spruce). On each parcel, the three most important biotope types in terms of surface area were identified (Drachenfels 2021) and translated to area-weighted biotope values ranging from 0 to 24 points (low to high, see Table A2 in Appendix) according to the German federal regulation on the avoidance and compensation of interventions into nature and landscape (BKompV 2020). An impression of the diverse situations encountered in SPFs is given in Fig. 2.

Selection of structural features and management situations found in SPFs in the study area. a Remnants of historical usage: old coppice stool in a formerly coppiced European beech (Fagus sylvatica) stand. b Extreme habitats: European beech forest on shallow soil over limestone in steep terrain. c Habitat trees: limb breakage with exposed heartwood (pedunculate oak, Quercus robur). d Small-scale harvesting of firewood, mainly European beech. Photographs: Peter Hansen
Data analysis
To assess the respective influence of owner attitudes and landscape parameters on the response variables forest structure and nature conservation values, we used ordinary least squares (OLS) regression analysis (compare Lange et al. (2022), Przepióra and Ciach (2022)). For each response variable (Table 2), we calculated a linear model using all predictors from landscape parameters and owner attitudes (Table 1). Multicollinearity among the predictors was relatively weak, with variance inflation factors below 4, and we decided not to remove any predictors (Naimi et al. 2014; Wooldridge 2013). We calculate semipartial correlations (Grömping 2015) as a measure of the effect size of individual predictors, and quote R2 values to assess the relative explanatory power of the models. All analyses were carried out using R 4.0.3 (R Core Team 2020).
Results
Descriptive statistics of the response variables are presented in Table 2. In this section, we focus on the OLS regression analysis. Relevant results are summarized in Table 3. Each row corresponds to an individual OLS regression computed for a single response variable (first column). To compare the explanatory power of the models, we used R2 values, as stated in column 2. The remaining columns express the individual contributions of the predictors to the model at hand. In the following, we group the results concerning living tree volumes, deadwood, harvested volumes, management intensity, and nature conservation values.
Living tree volumes
The model for broadleaf, conifer and total living tree volumes showed relatively high R2 values (Table 3). Owner attitudes had no significant influences on the response variables, while several landscape parameters exhibited significant influences. Living broadleaf tree volume was positively connected to ancient broadleaf and ancient mixed forest sites. The proportion of border length with open land also showed a positive correlation. For living conifer tree volumes, the amount of open land within a 2 km radius displayed a strong negative influence. The total living tree volume was strongly positively influenced by ancient mixed forest sites, while there was only a weak positive influence of broadleaf forest sites and the border length with open land.
Deadwood volumes
The picture for deadwood volumes as an important nature conservation value was different (Table 3), with forest owner attitudes acting as relevant influences. Deadwood was found in all decay stages, but recent deadwood prevailed (decay stage 1: 66.4%, stage 2: 21.3%, stage 3: 10.1%, stage 4: 2.1% of the volume). Total deadwood volume showed a strong dependency on the slope of the forest parcels, with additional influence by the implementation of close-to-nature silviculture. The respective influences became clearer if we discerned between broadleaf (63.2% of the deadwood volume) and conifer (36.8%) deadwood volumes. Broadleaf deadwood showed a slight positive influence by close-to-nature silviculture, the slope and ancient broadleaf forest sites, while parcel size had a negative influence. For conifer deadwood, we found a strong positive effect of slope. We also discerned between deadwood of anthropogenic (sawn) and natural origin (deadwood without saw marks). The occurrence of anthropogenic deadwood was highly dependent on owners’ attitudes. We found a strong positive influence of close-to-nature silviculture activities, a slight positive influence of the identification with regulating goals, and a slight negative influence of the implementation of active conservation measures. Landscape parameters played an important role as well, with a strong positive influence of ancient conifer forest sites. For deadwood of natural origin, slope was the sole but strong positive driver.
Accumulated harvested wood volumes and management intensity
For accumulated harvested wood volumes and the closely related management intensity, owner attitudes showed significant effects (Table 3). Total harvested wood volume, while most strongly influenced by ancient conifer forest sites, was fostered by the implementation of classical silvicultural activities (and slightly influenced by close-to-nature forestry activities). For broadleaf harvested volumes, ancient mixed forest sites had a strong positive influence, while the distance to the next paved road and the adoption of close-to-nature silvicultural activities also had positive influences. We saw a more pronounced influence of owner attitudes on harvested conifer volumes. Classical silvicultural activities were promotors, while the identification with provisioning goals and the pursuit of passive nature conservation hampered them. The strongest effect was a positive influence of ancient conifer forest sites.
For management intensity, we found positive effects of the support for cultural goals and the implementation of classical silvicultural activities. Passive conservation measures showed a negative influence. We additionally identified a strong positive effect of ancient conifer forest sites. The proportion of harvested to total wood volume (Iharv) showed a similar pattern. The volume proportion of non-natural tree species (Inonat) was slightly negatively influenced by ancient broadleaf forest sites and the amount of open land within a 2 km radius, while ancient conifer sites showed a positive effect. The volume proportion of anthropogenic deadwood (Idwcut) showed a different pattern: It was slightly positively influenced by the identification with cultural goals and the adoption of classical silvicultural activities.
Other nature conservation values
Within the group of nature conservation values apart from the various wood volumes, landscape parameters showed strong influences. Ancient broadleaf forest sites and the amount of open land within a 2 km radius exhibited a positive influence on biotope values. The mean stand age showed positive influences by both ancient broadleaf and mixed forest sites. Regarding TreMs, the most abundant TreM categories found in the field were crown deadwood and concavities (see table A3 in Appendix), and European beech (61.2%) and oaks (28.4%) were the main TreM-bearing trees. The mean diameter of the inventoried TreM-bearing trees was 65.0 cm (median 60.0 cm). Concerning the abundance of TreMs, the models showed patterns similar to the mean stand age model. Parcel size had a slightly negative effect, while the location on ancient broadleaf and mixed forest sites was positively correlated. Note the positive influence of the border length with open land for the density of large-diameter trees. Owner attitudes were only sparsely correlated with nature conservation values. Except for the model for the number of tree species (which reached only marginal explanatory power), almost no owner attitude variable exhibited significant influence on these models.
Discussion
In this study, we adopted a novel methodological approach to shed light on the effect of small-scale private forest owners’ attitudes on the structures of their forest stands, especially taking into account nature conservation values. Since we expected owner attitudes to only partly explain the variance seen in the response variables (note that the survey cast only a spotlight on a single time point in the long history of a parcel’s owners and their attitudes), several landscape parameters were included as well. We found that landscape parameters played an important role as predictive variables in almost all of the models that we considered. Such patterns are similar to those found in previous studies (Asbeck et al. 2021b; Kapusta et al. 2020; Kennedy et al. 2008; Levers et al. 2014). Concerning the influence of owner attitudes in the models, we found that some response variables are indeed correlated with forest owner attitudes, while others are mostly unrelated to the owners’ goals and activities. We propose that these two groups of response variables describe two underlying types of structures with distinct characteristics. We sort these structures as belonging into either “type A” or “type B”, as described as in the following (Fig. 3):
-
Type A Structures addressed as belonging to type A show strong correlations with several landscape parameters, but are mostly unrelated to owner attitudes. We postulate that these structures are evolving very slowly, such that the goals and activities of the current forest owner play only a minor role. Landscape parameters continuously persist over long time spans and are therefore plausible influence factors for the development of type A structures.
-
Type B Structures of this type show correlations with landscape parameters, but they are also prominently correlated with owner attitudes. We regard such structures as fast evolving and readily susceptible to the individual forest owner’s influence.

Overview of the two types of structures that we discerned: Slowly evolving type A structures like living tree volume or the density of tree-related microhabitats were not influenced by owner attitudes, but showed a correlation with landscape parameters. Fast evolving type B structures like the amount of deadwood or harvested wood were influenced by both landscape parameters, and owner goals and activities
In the following, we take a closer look at the individual response variables and propose a classification into either type A or type B and finally highlight crucial points concerning the integration of our results in forest conservation planning.
Wood volumes and valuable habitats (type A, slowly evolving structures)
The amount of living tree volume is determined mostly by high-diameter trees which take long time spans to mature. Consequently, current forest owner attitudes show minimal influence on wood volumes, supporting a classification of all response variables concerning living tree volumes into type A (slowly evolving structures). While the total living tree volume already shows correlations with ancient forest sites, it proved beneficial to discern between broadleaf and conifer volumes (which is also reasonable given their different habitat properties).
Ancient conifer sites do not seem to promote today’s conifer volume, in line with evidence for greater climate resilience of mixed stands (Honkaniemi et al. 2020). The dwindling of conifers on ancient conifer sites might be due to water stress and extensive damage by storms and bark beetles in spruce monocultures often present on such sites (Bujoczek et al. 2021; Kharuk et al. 2016). The strongest correlation of conifer wood volume is a negative one with the amount of open land within a 2 km radius. Historically, conifer plantations in the study area tended to cover large, contiguous areas and were planted on less valuable land. Smaller conifer patches in open land are more exposed to adverse climatic and biotic conditions like storm gusts, bark beetle swarming (Stříbrská et al. 2022), and drought (Buras et al. 2018).
Living broadleaf tree volumes are closely interrelated with the occurrence of TreMs and large-diameter trees (dbh ≥ 50 cm), and both benefit from a long continuity of forest cover (ancient broadleaf or mixed forest sites). Broadleaf wood volume correlated with parcels’ direct borders with open land, which also holds for large-diameter trees. The positive influence of forest border structures can be explained by the occurrence of old, large-diameter trees that were intentionally left in place and historically marked the border between forest and agricultural land. Such trees are prone to develop a variety of TreMs, and today belong to the oldest individuals (‘champion trees’) on many parcels, making forest borders and forest patches in agricultural landscapes a highly valuable habitat (Froidevaux et al. 2022; Orłowski and Nowak 2007). This is further supported by TreM density being negatively correlated with parcel size. In contrast to findings by Larrieu et al. (2022), neither elevation nor slope showed any significant correlation with TreM densities.
Biotope values as defined by BKompV (2020) can also be classified as type A structures. They lack correlations with owner attitudes but show correlations with ancient broadleaf sites and the amount of open land around the parcels. This is not surprising, as sites embedded in a fragmented landscape are likely to be covered by broadleaf forest, reaching high biotope values (Table A2 in Appendix). Such sites have already been identified to be promising ecosystem service providers (Decocq et al. 2016; Varela et al. 2018). Lastly, the number of tree species did not fit in any of the type A or type B categories.
Deadwood, harvested volume and management intensity (type B, fast evolving structures)
The mean amount of deadwood (30.4 m3 ha−1) reached several threshold values for the survival of different forest species groups as evaluated by Müller and Bütler (2010), who for instance recommend to establish forest stands with deadwood amounts > 20–50 m3 ha−1 in a network of forest landscapes. Variables related to deadwood amounts show significant influences of owner attitude variables and can be classified as type B (fast evolving structures). Across the models, we see positive influences of the forest owners’ implementation of close-to-nature silviculture, especially on sawn deadwood. Doerfler et al. (2017) found management intensity to be a driver of deadwood volume, with deadwood increasing up to a certain maximum and then decreasing with management intensity. We observed this behavior only for conifer deadwood, while broadleaf deadwood was uniformly negatively correlated to management intensity. Concerning the landscape parameters, consistent with Bujoczek et al. (2021), slope plays a major role for non-sawn (conifer) deadwood, but an influence of elevation as proposed in Kennedy et al. (2008) was not observed (possibly due to a lower elevation range among our sites). The influence of slope can be explained by more difficult access to steep, wind-thrown parcels, where accumulating deadwood cannot be easily extracted.
When looking at harvested wood volumes, strong influences by owner attitudes lead to a classification in type B as well. Especially conifer harvested volume is affected by owner attitudes like the implementation of classical silvicultural measures. The related forest management intensity index also shows a distinct type B pattern, with forest owner attitudes playing a major role in the models’ ability to explain variance in the response variables. Provisioning goals are negatively correlated with the sub-index Iharv (the proportion of harvested to total wood volume). It can be assumed that provisioning goals become less important as owners’ stands are more affected by storm damage. The implementation of passive conservation measures exerts a strong negative influence on management intensity. This is in line with research pointing at high management intensities being negatively correlated with most forest ecosystem services except for the production of biomass (Sing et al. 2018). High agreement with cultural goals might be perceived as characteristic of recreationalist forest owners (Blanco et al. 2015); however, our results reveal a correlation with forest management intensity. A possible explanation might be that forest owners who invest more time in management are more inclined to value cultural ecosystem services than more passive owners. Landscape parameters do not show strong influences on management intensity apart from the importance of ancient conifer forest sites.
Consequences for nature conservation
In the face of today’s ecological crises, management of forests is challenged to provide wood products and fuelwood while at the same time fostering a multitude of other forest ecosystem services. Integrative forest management has proven to be a suitable way to meet these requirements (Kraus and Krumm, 2013; Krumm et al. 2020), and within this framework, the conservation of forest biodiversity is a prime concern and depends on certain specific structures and habitats. We found that structures of high conservation value in SPFs can be separated into two types: the slowly evolving type A structures, and structures of the relatively fast evolving type B. It is therefore advisable to take these characteristics into account when designing nature conservation efforts in SPFs.
As development times of valuable type A structures (among them living tree volumes especially of broadleaf trees and TreMs) amount to decades and longer, conservation measures are able to protect already existing structures, but cannot be expected to support their formation on a short-term basis. Conservation efforts might target the protection of habitat trees (for instance, ‘champion trees’ that we often localized near border structures in the landscape), e.g., through contract-based conservation programs (Demant et al. 2020). Over the course of such a tree’s life, economical pressure might rise, leading to it being harvested at some point—especially considering the increasing demand for firewood. A continuous educational process will be needed to persuade forest owners of the inherent value of old broadleaf trees (Salomaa et al. 2016). When it comes to promoting the development of valuable structures like large-diameter trees or accompanying TreMs, very long-term commitments that transcend the individual forest owners would be beneficial, like the designation of protected areas. Apart from the naturally slow formation of TreMs, approaches to actively create TreM-like structures like artificial cavities or high stumps should be promoted (Adelmann et al. 2021), but a widespread application would require strong intrinsic or extrinsic motivation of forest owners (see Tiebel et al. (2023) for examples).
Valuable type B structures in SPFs include different sorts of deadwood which are readily influenced by the individual forest owners. SPFs are often endowed with a large pool of deadwood, and they are less liable to the economic pressures of the timber market (Eggers et al. 2014). Consequently, deadwood structures could be a primary target of individualized conservation efforts to reach out to SPF owners. Management intensity is also influenced by owner attitudes: The application of more extensive forms of forest management, but also of traditional, nowadays mostly abandoned forms of management with high importance for biodiversity protection in cultural landscapes, like coppicing or coppicing with standards, would be desirable from a nature conservation perspective (Buckley 2020). Extremely structure-rich remnants of formerly coppiced forests are still found throughout our study area in SPFs (Mölder 2016; Mölder et al. 2021), but need appropriate management lest they become lost in the conversion to high forest (Buckley 2020).
In contrast to forests in other ownership types, SPF owners are an exceptionally heterogeneous group (Blanco et al. 2015; Deuffic et al. 2018; Weiss et al. 2019; Westin et al. 2023), and it might prove beneficial to develop differentiated policy instruments to address different owner types. Many forest owners in our study area value the conservation of biodiversity higher than resource use or the generation of income, but this attitude is not readily translated into consequential behavior (Tiebel et al. 2023). We developed a typology based on the stated activities owners perform in their forests, and discern between multiple-use-oriented forest owners who would benefit from consultation by forestry stakeholders and contract-based conservation the most; conventional forest owners with a focus on wood production who would mostly adopt measures with a low trade-off, but are open to financial incentives; and conservation-oriented forest owners who are mostly passive in their management and would benefit from practice-oriented consultation to implement active measures (Tiebel et al. 2023). SPF owners, especially of the last type, might be an elusive group (Fairchild et al. 2022) and difficult to reach for private conservation initiatives due to lack of or scattering of organization, but their stands offer many possibilities for the conservation and development of valuable habitat structures, also given SPF owners’ high identification with their property (Tiebel et al. 2022).
Concerning financial incentives for nature conservation, activities like the provisioning and retention of deadwood, the promotive management of habitat trees, or the resumption of coppicing, are very promising candidates for contract-based nature conservation in private forests (Demant et al. 2020), and SPF owners are often inclined to embrace such initiatives (Juutinen et al. 2020). Financial incentives to guide forest management decisions are regarded as a key ingredient in adapting forests to climatic change (Bauhus 2022). Until recently, they have played a minor role in Germany, especially in SPFs (Franz et al. 2018), but this is changing (VNPWaldR 2021). Policy makers, forestry and nature conservation organizations should be aware of the high but threatened nature conservational values present in SPFs.
Conclusion
We proposed a novel method to assess the distribution of forest structural (living and harvested wood volumes, management intensity) and nature conservational (diversity, quantities of deadwood and microhabitats) variables in small-scale private forests in Germany’s Lower Saxon Hills. To this aim, we used predictors generated by a forest owner questionnaire (owners’ goals and management measures), and landscape parameters. We separated the response variables into two types, and interpret these as slowly evolving structures (type A) and fast evolving structures (type B). Easily obtainable landscape parameters explain a large proportion of the variance seen in type A structures in SPFs, like broadleaf wood volume and tree-related microhabitats. Structural data about such forests is often missing due to the SPF owners’ low level of institutional organization Hence, leveraging data on landscape parameters might prove useful in predicting conservation values to reduce potential field work to identify structural hotspots. Type B structures were shown to be influenced by landscape parameters but also, for the first time, prominently by owner attitudes.
Concerning the success of nature conservation in SPFs, the multitude of owner types is increasingly recognized as a decisive factor, but differing characteristics of the target structures in the forests need to be taken into account as well. Slowly evolving structures (type A) depend on long-term commitments to conservation legislation, financial incentives and generation-spanning education of forest owners. Efforts to promote faster evolving valuable structures (type B) like broadleaf deadwood, or the resumption of traditional forms of management might prove particularly advantageous in small-scale private forests given the structural diversity of the stands, but also the often strong identification of owners with their land.
Data availability
All data used in the scope of this article can be found on the platform Zenodo by using the following link: https://doi.org/10.5281/zenodo.7840025.
Code availability
Not applicable.
References
Adelmann W, Hagge J, Langhammer P, Höhna N, Hotes S, Werneyer M, Drexler M, Müller J (2021) Aktiv im Wald: Naturschutz mit der Kettensäge. Bayerische Akademie für Naturschutz und Landschaftspflege, Laufen
Asbeck T, Großmann J, Paillet Y, Winiger N, Bauhus J (2021a) The use of tree-related microhabitats as forest biodiversity indicators and to guide integrated forest management. Curr For Rep 7:59–68. https://doi.org/10.1007/s40725-020-00132-5
Asbeck T, Kozák D, Spînu AP, Mikoláš M, Zemlerová V, Svoboda M (2021b) Tree-related microhabitats follow similar patterns but are more diverse in primary compared to managed temperate mountain forests. Ecosystems 25:712–726. https://doi.org/10.1007/s10021-021-00681-1
Bauhus J (2022) Die Anpassung der Wälder an den Klimawandel – eine waldwirtschaftliche Perspektive. Nat Landsch 97:318–324. https://doi.org/10.19217/NuL2022-07-01
Bieling C (2004) Non-industrial private-forest owners: Possibilities for increasing adoption of close-to-nature forest management. Eur J For Res 123:293–303. https://doi.org/10.1007/s10342-004-0042-6
BKompV (2020) Bundeskompensationsverordnung vom 14. Mai 2020 (BGBl. I S. 1088)
Blanco V, Brown C, Rounsevell M (2015) Characterising forest owners through their objectives, attributes and management strategies. Eur J For Res 134:1027–1041. https://doi.org/10.1007/s10342-015-0907-x
BMEL (2014) Wald in Deutschland – Ergebnisse der dritten Bundeswaldinventur. Bundesministerium für Ernährung und Landwirtschaft (BMEL), Berlin
Buckley P (2020) Coppice restoration and conservation: a European perspective. J for Res 25:125–133. https://doi.org/10.1080/13416979.2020.1763554
Bujoczek L, Bujoczek M (2022) Factors influencing the diversity of deadwood, a crucial microhabitat for many rare and endangered saproxylic organisms. Ecol Indic 142:109197. https://doi.org/10.1016/j.ecolind.2022.109197
Bujoczek L, Bujoczek M, Zięba S (2021) How much, why and where? Deadwood in forest ecosystems: the case of Poland. Ecol Indic 121:107027. https://doi.org/10.1016/j.ecolind.2020.107027
Buras A, Schunk C, Zeiträg C, Herrmann C, Kaiser L, Lemme H, Straub C, Taeger S, Gößwein S, Klemmt H-J, Menzel A (2018) Are Scots pine forest edges particularly prone to drought-induced mortality? Environ Res Lett 13:025001. https://doi.org/10.1088/1748-9326/aaa0b4
Chivulescu S, Pitar D, Apostol B, Leca S, Badea O (2022) Importance of dead wood in virgin forest ecosystem functioning in Southern Carpathians. Forests 13:409. https://doi.org/10.3390/f13030409
Decocq G, Andrieu E, Brunet J, Chabrerie O, De Frenne P, De Smedt P, Deconchat M, Diekmann M, Ehrmann S, Giffard B, Mifsud EG, Hansen K, Hermy M, Kolb A, Lenoir J, Liira J, Moldan F, Prokofieva I, Rosenqvist L, Varela E, Valdés A, Verheyen K, Wulf M (2016) Ecosystem services from small forest patches in agricultural landscapes. Curr For Rep 2:30–44. https://doi.org/10.1007/s40725-016-0028-x
Demant L, Bergmeier E, Walentowski H, Meyer P (2020) Suitability of contract-based nature conservation in privately-owned forests in Germany. Nat Conserv 42:89–112. https://doi.org/10.3897/natureconservation.42.58173
Deuffic P, Sotirov M, Arts B (2018) “Your policy, my rationale”. How individual and structural drivers influence European forest owners’ decisions. Land Use Policy 79:1024–1038. https://doi.org/10.1016/j.landusepol.2016.09.021
Doerfler I, Müller J, Gossner MM, Hofner B, Weisser WW (2017) Success of a deadwood enrichment strategy in production forests depends on stand type and management intensity. For Ecol Manage 400:607–620. https://doi.org/10.1016/j.foreco.2017.06.013
Drachenfels O von (2021) Kartierschlüssel für Biotoptypen in Niedersachsen unter besonderer Berücksichtigung der gesetzlich geschützten Biotope sowie der Lebensraumtypen von Anhang I der FFH-Richtlinie, Stand März 2021. Niedersächsischer Landesbetrieb für Wasserwirtschaft, Küsten- und Naturschutz, Hannover
DWD (2018a) DWD Climate Data Center (CDC): Multi-annual grids of precipitation height over Germany 1981–2010, version v1.0.
DWD (2018b) DWD Climate Data Center (CDC): Multi-annual means of grids of air temperature (2m) over Germany 1981–2010, version v1.0.
Eggers J, Lämås T, Lind T, Öhman K (2014) Factors influencing the choice of management strategy among small-scale private forest owners in Sweden. Forests 5:1695–1716. https://doi.org/10.3390/f5071695
European Environment Agency (2018) CORINE Land Cover 2018. https://www.eea.europa.eu/ds_resolveuid/7243bfa34e2840149aaa764fc8ddb597 (Accessed 2 Jan 2023)
Fairchild E, Ulrich-Schad JD, Petrzelka P, Ma Z (2022) The lay of the land: What we know about non-operating agricultural and absentee forest landowners in the U.S. and Europe. J Environ Manage 313:114991. https://doi.org/10.1016/j.jenvman.2022.114991
Fischer AP, Klooster A, Cirhigiri L (2019) Cross-boundary cooperation for landscape management: collective action and social exchange among individual private forest landowners. Landsc Urban Plan 188:151–162. https://doi.org/10.1016/j.landurbplan.2018.02.004
Franz K, von Blomberg M, Demant L, Lutter C, Seintsch B, Selzer AM (2018) Umsetzung von Vertragsnaturschutz im deutschen Wald. AFZ/Wald 73(21):13–15
Froidevaux JSP, Laforge A, Larrieu L, Barbaro L, Park K, Fialas PC, Jones G (2022) Tree size, microhabitat diversity and landscape structure determine the value of isolated trees for bats in farmland. Biol Conserv 267:109476. https://doi.org/10.1016/j.biocon.2022.109476
Grömping U (2015) Variable importance in regression models. Wires Comput Stat 7:137–152. https://doi.org/10.1002/wics.1346
Honkaniemi J, Rammer W, Seidl R (2020) Norway spruce at the trailing edge: the effect of landscape configuration and composition on climate resilience. Landsc Ecol 35:591–606. https://doi.org/10.1007/s10980-019-00964-y
Jackson HB, Fahrig L (2015) Are ecologists conducting research at the optimal scale? Is research conducted at optimal scales? Glob Ecol Biogeogr 24:52–63. https://doi.org/10.1111/geb.12233
Johann F, Schaich H (2016) Land ownership affects diversity and abundance of tree microhabitats in deciduous temperate forests. For Ecol Manage 380:70–81. https://doi.org/10.1016/j.foreco.2016.08.037
Juutinen A, Tolvanen A, Koskela T (2020) Forest owners’ future intentions for forest management. For Policy Econ 118:102220. https://doi.org/10.1016/j.forpol.2020.102220
Kahl T, Bauhus J (2014) An index of forest management intensity based on assessment of harvested tree volume, tree species composition and dead wood origin. Nat Conserv 7:15–27. https://doi.org/10.3897/natureconservation.7.7281
Kapusta P, Kurek P, Piechnik Ł, Szarek-Łukaszewska G, Zielonka T, Żywiec M, Holeksa J (2020) Natural and human-related determinants of dead wood quantity and quality in a managed European lowland temperate forest. For Ecol Manage 459:117845. https://doi.org/10.1016/j.foreco.2019.117845
Kennedy RSH, Spies TA, Gregory MJ (2008) Relationships of dead wood patterns with biophysical characteristics and ownership according to scale in Coastal Oregon, USA. Landsc Ecol 23:55–68. https://doi.org/10.1007/s10980-007-9164-9
Kharuk VI, Im ST, Dvinskaya ML (2016) Decline of spruce (Picea abies) in forests of Belarus. Russ J Ecol 47:241–248. https://doi.org/10.1134/S106741361603005X
Kjučukov P, Hofmeister J, Bače R, Vítková L, Svoboda M (2022) The effects of forest management on biodiversity in the Czech Republic: an overview of biologists’ opinions. iForest 15:187–196. https://doi.org/10.3832/ifor3953-015
Kõrkjas M, Remm L, Lõhmus A (2021) Development rates and persistence of the microhabitats initiated by disease and injuries in live trees: a review. For Ecol Manage 482:118833. https://doi.org/10.1016/j.foreco.2020.118833
Kozák D, Svitok M, Zemlerová V, Mikoláš M, Lachat T, Larrieu L, Paillet Y, Buechling A, Bače R, Keeton WS, Vítková L, Begovič K, Čada V, Dušátko M, Ferenčík M, Frankovič M, Gloor R, Hofmeister J, Janda P, Kameniar O, Kníř T, Majdanová L, Mejstřík M, Pavlin J, Ralhan D, Rodrigo R, Roibu C, Synek M, Vostarek O, Svoboda M (2023) Importance of conserving large and old trees to continuity of tree-related microhabitats. Conserv Biol 25:712–726. https://doi.org/10.1111/cobi.14066
Kraus D, Krumm F (eds) (2013) Integrative approaches as an opportunity for the conservation of forest biodiversity. European Forest Institute, Joensuu
Krumm F, Schuck A, Rigling A (2020) How to balance forestry and biodiversity conservation – A view across Europe. EFI and WSL, Birmensdorf. https://doi.org/10.16904/ENVIDAT.196
Lange F, Ammer C, Leitinger G, Seliger A, Zerbe S (2022) Is Douglas fir [Pseudotsuga menziesii (Mirbel) Franco] invasive in Central Europe? A case study from south-west Germany. Front For Glob Change 5:844580. https://doi.org/10.3389/ffgc.2022.844580
Larrieu L, Paillet Y, Winter S, Bütler R, Kraus D, Krumm F, Lachat T, Michel AK, Regnery B, Vandekerkhove K (2018) Tree related microhabitats in temperate and Mediterranean European forests: a hierarchical typology for inventory standardization. Ecol Indic 84:194–207. https://doi.org/10.1016/j.ecolind.2017.08.051
Larrieu L, Courbaud B, Drénou C, Goulard M, Bütler R, Kozák D, Kraus D, Krumm F, Lachat T, Müller J, Paillet Y, Schuck A, Stillhard J, Svoboda M, Vandekerkhove K (2022) Key factors determining the presence of Tree-related Microhabitats: a synthesis of potential factors at site, stand and tree scales, with perspectives for further research. For Ecol Manage 515:120235. https://doi.org/10.1016/j.foreco.2022.120235
LBEG (2022) NIBIS Kartenserver, Geological map 1:25000, Landesamt für Bergbau, Energie und Geologie (LBEG), Hannover. https://nibis.lbeg.de/cardomap3 (Accessed 2 Jan 2023)
Levers C, Verkerk PJ, Müller D, Verburg PH, Butsic V, Leitão PJ, Lindner M, Kuemmerle T (2014) Drivers of forest harvesting intensity patterns in Europe. For Ecol Manage 315:160–172. https://doi.org/10.1016/j.foreco.2013.12.030
LGLN (2022) Digitale Geländemodelle (DGM) – Landesamt für Geoinformation und Landesvermessung Niedersachsen. https://www.lgln.niedersachsen.de/startseite/geodaten_karten/3dgeobasisdaten/dgm/digitale-gelaendemodelle--dgm-143150.html (Accessed 2 Jan 2023)
Loeb CD, D’Amato AW (2020) Large landscape conservation in a mixed ownership region: opportunities and barriers for putting the pieces together. Biol Conserv 243:108462. https://doi.org/10.1016/j.biocon.2020.108462
Maltamo M, Uuttera J, Kuusela K (1997) Differences in forest stand structure between forest ownership groups in central Finland. J Environ Manage 51:145–167. https://doi.org/10.1006/jema.1997.0140
Martin M, Paillet Y, Larrieu L, Kern CC, Raymond P, Drapeau P, Fenton NJ (2022) Tree-related microhabitats are promising yet underused tools for biodiversity and nature conservation: a systematic review for international perspectives. Front For Glob Change. https://doi.org/10.3389/ffgc.2022.818474
Mehtätalo L, de Miguel S, Gregoire TG (2015) Modeling height-diameter curves for prediction. Can J For Res 45:826–837. https://doi.org/10.1139/cjfr-2015-0054
Meyer P, Fricke C (2018) Probekreisinventuren in niedersächsischen Naturwäldern – Aufnahmeanweisung. Nordwestdeutsche Forstliche Versuchsanstalt, Göttingen
ML (2014) Die Bundeswaldinventur 3 – Ergebnisse für Niedersachsen. Niedersächsisches Ministerium für Ernährung. Landwirtschaft und Verbraucherschutz (ML), Hannover
Mölder A, Tiebel M, Plieninger T (2021) On the interplay of ownership patterns, biodiversity, and conservation in past and present temperate forest landscapes of Europe and North America. Curr For Rep 7:195–213. https://doi.org/10.1007/s40725-021-00143-w
Mölder A (2016) Small forest parcels, management diversity and valuable coppice habitats: An 18th century political compromise in the Osnabrück region (NW Germany) and its long-lasting legacy. iForest 9:518–528. https://doi.org/10.3832/ifor1834-009
Müller J, Bütler R (2010) A review of habitat thresholds for dead wood: a baseline for management recommendations in European forests. Eur J For Res 129:981–992. https://doi.org/10.1007/s10342-010-0400-5
Naimi B, Hamm NAS, Groen TA, Skidmore AK, Toxopeus AG (2014) Where is positional uncertainty a problem for species distribution modelling? Ecography 37:191–203. https://doi.org/10.1111/j.1600-0587.2013.00205.x
NFP (2010) Historisch alte Waldstandorte (Niedersachsen). Niedersächsisches Forstplanungsamt, Wolfenbüttel
Norton M, Baldi A, Buda V, Carli B, Cudlin P, Jones MB, Korhola A, Michalski R, Novo F, Oszlányi J, Santos FD, Schink B, Shepherd J, Vet L, Walloe L, Wijkman A (2019) Serious mismatches continue between science and policy in forest bioenergy. Glob Change Biol Bioenergy 11:1256–1263. https://doi.org/10.1111/gcbb.12643
OpenStreetMap contributors (2022) OpenStreetMap database. https://www.openstreetmap.org/ (Accessed 2 Jan 2023)
Orłowski G, Nowak L (2007) The importance of marginal habitats for the conservation of old trees in agricultural landscapes. Landsc Urban Plan 79:77–83. https://doi.org/10.1016/j.landurbplan.2006.03.005
Parisi F, Pioli S, Lombardi F, Fravolini G, Marchetti M, Tognetti R (2018) Linking deadwood traits with saproxylic invertebrates and fungi in European forests – a review. iForest 11:423–436. https://doi.org/10.3832/ifor2670-011
Piaszczyk W, Lasota J, Gaura G, Błońska E (2021) Effect of deadwood decomposition on the restoration of soil cover in landslide areas of the Karpaty Mountains. Poland Forests 12:237. https://doi.org/10.3390/f12020237
Przepióra F, Ciach M (2022) Tree microhabitats in natural temperate riparian forests: an ultra-rich biological complex in a globally vanishing habitat. Sci Total Environ 803:149881. https://doi.org/10.1016/j.scitotenv.2021.149881
R Core Team (2020) R: A language and environment for statistical computing. https://www.r-project.org/ (Accessed 2 Jan 2023)
Ranius T, Niklasson M, Berg N (2009) Development of tree hollows in pedunculate oak (Quercus robur). For Ecol Manage 257:303–310. https://doi.org/10.1016/j.foreco.2008.09.007
Rendenieks Z, Nikodemus O, Brūmelis G (2015) The implications of stand composition, age and spatial patterns of forest regions with different ownership type for management optimisation in northern Latvia. For Ecol Manage 335:216–224. https://doi.org/10.1016/j.foreco.2014.10.001
Rohde M, Langer G, Hurling R, Plasil P (2019) Waldschutzsituation 2018 in Nordwestdeutschland. AFZ/Wald 74(7):38–41
Rohde M, Langer G, Hurling R, Plašil P (2021) Waldschutzsituation 2020 in Nordwestdeutschland. AFZ/Wald 76(9):42–46
Salomaa A, Paloniemi R, Hujala T, Rantala S, Arponen A, Niemelä J (2016) The use of knowledge in evidence-informed voluntary conservation of Finnish forests. For Policy Econ 73:90–98. https://doi.org/10.1016/j.forpol.2016.09.004
Schaich H, Plieninger T (2013) Land ownership drives stand structure and carbon storage of deciduous temperate forests. For Ecol Manage 305:146–157. https://doi.org/10.1016/j.foreco.2013.05.013
Schmithüsen FJ, Hirsch F (2010) Private forest ownership in Europe (Report), Geneva timber and forest study papers. ETH Zurich. https://doi.org/10.3929/ethz-a-006311424
Sing L, Metzger MJ, Paterson JS, Ray D (2018) A review of the effects of forest management intensity on ecosystem services for northern European temperate forests with a focus on the UK. Forestry 91:151–164. https://doi.org/10.1093/forestry/cpx042
Sterman J, Moomaw W, Rooney-Varga JN, Siegel L (2022) Does wood bioenergy help or harm the climate? Bull At Sci 78:128–138. https://doi.org/10.1080/00963402.2022.2062933
Stříbrská B, Hradecký J, Čepl J, Tomášková I, Jakuš R, Modlinger R, Netherer S, Jirošová A (2022) Forest margins provide favourable microclimatic niches to swarming bark beetles, but Norway spruce trees were not attacked by Ips typographus shortly after edge creation in a field experiment. For Ecol Manage 506:119950. https://doi.org/10.1016/j.foreco.2021.119950
Takala T, Brockhaus M, Hujala T, Tanskanen M, Lehtinen A, Tikkanen J, Toppinen A (2022) Discursive barriers to voluntary biodiversity conservation: the case of Finnish forest owners. For Policy Econ 136:102681. https://doi.org/10.1016/j.forpol.2021.102681
Tiebel M, Mölder A, Plieninger T (2021) Small-scale private forest owners and the European Natura 2000 conservation network: perceived ecosystem services, management practices, and nature conservation attitudes. Eur J For Res 140:1515–1531. https://doi.org/10.1007/s10342-021-01415-7
Tiebel M, Mölder A, Plieninger T (2022) Conservation perspectives of small-scale private forest owners in Europe: a systematic review. Ambio 51:836–848. https://doi.org/10.1007/s13280-021-01615-w
Tiebel M, Mölder A, Bieling C, Hansen P, Plieninger T (2023) Engaging small-scale private forest owners for transformative change towards integrative conservation. (In Review)
Torras O, Gil-Tena A, Saura S (2012) Changes in biodiversity indicators in managed and unmanaged forests in NE Spain. J For Res 17:19–29. https://doi.org/10.1007/s10310-011-0269-2
UNECE (2022) Draft forest products market statement. United Nations Economic Commission for Europe, Committee on Forests and the Forest Industry, Geneva
Varela E, Verheyen K, Valdés A, Soliño M, Jacobsen JB, De Smedt P, Ehrmann S, Gärtner S, Górriz E, Decocq G (2018) Promoting biodiversity values of small forest patches in agricultural landscapes: ecological drivers and social demand. Sci Total Environ 619–620:1319–1329. https://doi.org/10.1016/j.scitotenv.2017.11.190
Verga EG, Huais PY, Herrero ML (2021) Population responses of pest birds across a forest cover gradient in the Chaco ecosystem. For Ecol Manage 491:119174. https://doi.org/10.1016/j.foreco.2021.119174
VNPWaldR (2021) Richtlinie über Zuwendungen nach dem Bayerischen Vertragsnaturschutzprogramm Wald (VNPWaldR 2021). Bayerische Staatsministerien für Umwelt und Verbraucherschutz sowie für Ernährung. Landwirtschaft und Forsten, München
Weiss G, Lawrence A, Hujala T, Lidestav G, Nichiforel L, Nybakk E, Quiroga S, Sarvašová Z, Suarez C, Živojinović I (2019) Forest ownership changes in Europe: State of knowledge and conceptual foundations. For Policy Econ 99:9–20. https://doi.org/10.1016/j.forpol.2018.03.003
Westin K, Bolte A, Haeler E, Haltia E, Jandl R, Juutinen A, Kuhlmey K, Lidestav G, Mäkipää R, Rosenkranz L, Triplat M, Skudnik M, Vilhar U, Schueler S (2023) Forest values and application of different management activities among small-scale forest owners in five EU countries. For Policy Econ 146:102881. https://doi.org/10.1016/j.forpol.2022.102881
Wooldridge JM (2013) Introductory econometrics: a modern approach, 5th edn. South-Western Cengage Learning, Mason, OH, USA
Acknowledgements
We are indebted to two anonymous reviewers for suggestions that greatly improved the article.
Funding
Open Access funding enabled and organized by Projekt DEAL. This work was financially supported through the project “Small private forests: Conservation through resource use (KLEIBER)”, funded by the German Federal Ministry of Food and Agriculture (BMEL) through the Agency of Renewable Resources (FNR) within the funding program “Renewable Resources”, and according to a decision of the German Parliament (FKZ 22001218 and 22023218).
Author information
Authors and Affiliations
Contributions
PH and AM designed the analysis which was performed by PH. PH wrote the manuscript together with AM, MT and TP. Fieldwork was conducted by PH, AM and MT. All authors discussed the results and commented on the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors do not have any conflicts of interest regarding the article.
Additional information
Communicated by Claus Bässler.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Appendices
Appendix
A1: Private forest owner questionnaire, translated version
Remark: The order of the individual sub-questions was changed to group together questions related to the same goal type (Q1) or activity type (Q2). Additional information not present in the original questionnaire is given in brackets.
Question 1: How important do you perceive the following objectives regarding your forest?
For each row, please check the answer that applies to you.
[type of goal] | not important | rather unimportant | neither | rather important | very important | no opinion | |
|---|---|---|---|---|---|---|---|
[fostering regulating ecosystem services] | Long-term preservation of a stable and healthy forest stand | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ |
Protection of soil, water, air quality | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
Biodiversity conservation | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
Carbon sequestration, conservation of carbon sinks | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
[fostering cultural ecosystem services] | Protection as a cultural asset | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ |
Preservation of family heritage | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
Safeguarding or enhancement of landscape beauty | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
Possibility for nature observation | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
Possibility for own recreation | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
Preservation as a place of education | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
Possibility for hunting | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
[fostering provisioning ecosystem services] | Wood production for personal consumption | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ |
Wood production for selling | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
Collection of non-wood products | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | |
[financial goals] | Preservation for financial security | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ |
Profit maximization | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ | ⬜ |
Question 2: Which of the following activities are carried out in your forest (both by you and by third parties)?
Please check all that apply.
[Type of activity] | ||
|---|---|---|
[Classical silvicultural] | ⬜ | Thinning (e.g., felling of competitive trees, extraction of firewood) |
⬜ | Protection of young plants against browsing (e.g., by fences) | |
⬜ | Timber sale | |
⬜ | Planting / promotion of native tree species (e.g., common oak, sessile oak, European beach, Norway spruce, Scots pine) | |
⬜ | Planting / promotion of introduced tree species (e.g., Douglas fir, northern red oak, grand fir) | |
⬜ | Harvest of single mature trees | |
⬜ | Pruning (removal of branches for higher economic value) | |
[Close-to-nature silvicultural] | ⬜ | Reduction of damage due to logging |
⬜ | Avoidance of chemical pesticides | |
⬜ | Promotion of broadleaf trees in coniferous forests | |
⬜ | Use of logging horses | |
⬜ | Promotion of natural tree regeneration | |
⬜ | Avoiding clear-cuts | |
[Traditional silvicultural] | ⬜ | Coppicing/coppicing with standards |
⬜ | Wood pasture | |
[Active conservation] | ⬜ | Promotion of a shrub layer |
⬜ | Protection/restoration of light stand structures (open/light areas) | |
⬜ | Protection/maintenance of special structures (e.g., bizarre growth forms) | |
⬜ | Promotion of rare native tree and shrub species (e.g., European crab apple, wild service tree, common yew) | |
⬜ | Species protection measures (e.g., for bats or birds) | |
⬜ | Habitat restoration (e.g., peat bogs, flowing waters, stagnant waters) | |
⬜ | Removal of introduced species (e.g., black cherry, giant hogweed, Japanese knotweed) | |
[Passive conservation] | ⬜ | Protection of habitat trees |
⬜ | Protection of dead wood | |
⬜ | Non-use of parts of the stand | |
⬜ | None of the above |
A2: Encountered biotopes and their values
Biotope codes (Drachenfels 2021) used in the surveys, translation given according to BKompV (2020).
Biotope code | Biotope | Value (0–30 yrs) | Value (> 30–80 yrs) | Value (> 80 yrs) |
|---|---|---|---|---|
BMS | mesophilic Crataegus shrub | 13 | 13 | 13 |
WAR | nutrient-rich Alnus swamp forest | 14 | 20 | 23 |
WCE | medium-wet Quercus/Carpinus forest, base-poor | 15 | 20 | 23 |
WCR | wet Quercus/Carpinus forest, base-rich | 15 | 20 | 23 |
WEG | Alnus/Fraxinus riparian forest | 12 | 15 | 18 |
WGF | wet mixed non-Quercus/Fagus forest, base-rich | 15 | 19 | 22 |
WGM | medium-wet mixed non-Quercus/Fagus forest, base-rich | 14 | 16 | 18 |
WJL | young-age broadleaf forest | 11 | 11 | 11 |
WJN | young-age conifer forest | 9 | 9 | 9 |
WKC | lichen-rich Pinus sylvestris forest on nutrient-poor dry sand | 14 | 19 | 22 |
WLB | acidophilic hill/mountain Fagus sylvatica forest | 14 | 17 | 20 |
WLM | acidophilic lowland Fagus sylvatica forest on clay | 14 | 17 | 20 |
WMB | mesophilic hill/mountain Fagus sylvatica forest, base-poor | 14 | 17 | 20 |
WMK | mesophilic hill/mountain Fagus sylvatica forest, base-rich | 14 | 16 | 18 |
WMT | mesophilic lowland Fagus sylvatica forest, base-poor | 14 | 17 | 20 |
WNS | miscellaneous swamp forest | 15 | 18 | 21 |
WPB | Betula/Populus tremula pioneer forest | 13 | 13 | 13 |
WPE | Acer/Fraxinus pioneer forest | 13 | 13 | 13 |
WPS | miscellaneous pioneer forest | 13 | 13 | 13 |
WQE | miscellaneous acidophilic Quercus mixed forest | 15 | 19 | 22 |
WQF | medium-wet Quercus mixed forest on sand | 15 | 20 | 23 |
WRM | highly structured forest border, average locations | 16 | 16 | 16 |
WRT | highly structured forest border, dry, warm, base-rich | 16 | 16 | 16 |
WRW | highly structured forest border with hedge bank | 16 | 16 | 16 |
WSS | medium-wet ravine forest on silicate | 15 | 17 | 20 |
WTB | Fagus sylvatica forest on dry, warm limestone | 15 | 19 | 22 |
WXH | plantation of native broadleaf trees | 11 | 13 | 16 |
WZD | plantation of Pseudotsuga menziesii | 6 | 10 | 12 |
WZF | plantation of Picea abies | 9 | 11 | 14 |
WZK | plantation of Pinus sylvestris | 9 | 11 | 14 |
WZL | plantation of Larix decidua | 9 | 11 | 14 |
WZS | miscellaneous plantation of non-native species | 6 | 10 | 12 |
Wn | coppice forest | 19 | 19 | 19 |
A3: Tree-related microhabitats
TreM categories were used as defined in Larrieu et al. (2018).
Form | Group | Number | % |
|---|---|---|---|
Cavities | Woodpecker breeding cavities | 8 | 1.0 |
Rot-holes | 74 | 8.8 | |
Insect galleries | 0 | 0.0 | |
Concavities | 167 | 19.9 | |
Tree injuries and exposed wood | Exposed sapwood only | 17 | 2.0 |
Exposed sapwood and heartwood | 52 | 6.2 | |
Crown deadwood | Crown deadwood | 434 | 51.7 |
Excrescences | Twig tangles | 34 | 4.1 |
Burrs and cankers | 4 | 0.5 | |
Fruiting bodies of saprophylic fungi | Perennial fungal fruiting bodies | 9 | 1.1 |
Ephemeral fungal fruiting bodies | 0 | 0.0 | |
Epiphytic and epixylic structures | Epiphytic and parasitic crypto- and phanerogams | 28 | 3.3 |
Nests | 5 | 0.6 | |
Microsoils | 4 | 0.5 | |
Exudates | Exudates | 3 | 0.4 |
A4: Full version of Table 3
Influences of predictors on response variables. Each line corresponds to a linear model for the response variable given in the first column. The second column lists the R2 value of each model. The predictors are given in the remaining columns. Significant influences are given in bold face. If a predictor represents a significant contribution to the model, it is shown in this table with its semipartial correlation value and an indication of the Pr( >|t|) value p: “(*)” for p ≤ 0.1, “*” for p ≤ 0.05, “**” for p ≤ 0.01, “***” for p ≤ 0.001.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hansen, P., Tiebel, M., Plieninger, T. et al. Owner attitudes and landscape parameters drive stand structure and valuable habitats in small-scale private forests of Lower Saxony (Germany). Eur J Forest Res 142, 1011–1028 (2023). https://doi.org/10.1007/s10342-023-01571-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-023-01571-y