
Abstract
The unpredictable nature of climate change impacts, e.g. warming producing a latitudinal shift in photoperiod, may modify trees’ rates of survival and capacities for growth. This is also true of the Scots pine (Pinus sylvestris L.), as one of Europe’s main forest-forming tree species. The best results in the study of tree responses to climate change can be obtained during long-term experiments carried out on provenance trials. Such objects make it possible to precisely determine the growth and adaptation reactions of offspring coming from different soil and climatic conditions to the conditions of the research area. Research of this kind has been conducted by analysing growth in the progeny of 16 different Polish provenances of Scots pine on a single provenance experiment located in the centre of Poland, in an area where annual precipitation is at the rather low average level of 580 mm. The experiment was established in 1966, using a randomised block design with five replicates for each of the tested provenances. In autumn 2015, trees present were monitored and analysis of variance was used for their heights and diameters at breast height, with estimates also made regarding straightness of the trunk, crown width and branching angle. At the same time, ecophysiographic features of the places of occurrence of parent stands for each of the provenances were also analysed, with a quantification of the relationship between the growth traits studied and ecophysiographic features also achieved using PCA, while Ward’s Method of clustering was used to establish provenance groups characterised by multi-trait similarity. In the light of what is known about forecast climate change, it would be possible to transfer progeny of different pine populations, if only from places where levels of precipitation are lower, along with values for the De Martonne aridity index. In practice, Central Poland might play host to the progenies of pines from the western part of the country—as represented here by the Bolewice, Rychtal, Karsko and Gubin provenances. On the other hand, the progenies of pines from stands at higher altitudes will not be suitable for cultivation in lowland locations in Central Poland.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The natural range of occurrence of the Scots pine is so extensive that the species manifests far-reaching differences across it, in relation to morphology, productivity and quality (Sabor 1993; Giertych 1997; Reich and Oleksyn 2008). And given its economic significance, this species of pine is the subject of a large number of studies supplying results that allow for the identification and selection of the populations or individuals whose cultivation is likely to prove most valuable under given geographical and environmental conditions. Engagement in appropriate means of selection (above all based on the division of the given country into seed regions) can bring measurable economic effects, while also enhancing both the persistence and the stability of stands of Scots pine (Chodzicki 1975).
The impacts of climate warming are also hard to predict when it comes to the reactions of different species of tree (Davis et al. 2005; Matala et al. 2006; Scholze et al. 2006; Misi et al. 2019). Warming has a direct influence on chances of survival, growth increments and the quality of growth that is achieved (Eilmann et al. 2009; Misi and Náfrádi 2017). The climate warming in the northern hemisphere may also result in a shift of climatic zones to the north (Wang et al. 2006).
Populations of pines occurring in a boreal climate may react positively to warming, even as reactions of trees in central and southern parts of Europe are quite the reverse (Rehfeldt et al. 2002; Reich and Oleksyn 2008). In addition to climatic aspects, the genetic distinctiveness of eastern and western populations, reflecting their different pedigrees, should be taken into account (Naydenov et al. 2007). Savva and Vaganov (2006) reported the genetic determinants of pine response to the factor of drought. Based on observation conducted on two provenance experiments (founded in 1964 and 1974) authors pointed out that in southern populations there is a different incremental dynamics of early and late wood than in northern populations. This would tend to denote the genetic adaptation of particular populations to the climatic conditions they experience.
There is obviously considerable value in assessing trees’ reactions to climate change, by way of long-term study on permanent provenance trials. For—under these circumstances—the progeny of populations originating in different geographical locations can be present over long periods under local climatic conditions that may be characterised and defined precisely. Work of this kind therefore allows for conclusion-drawing in relation to the reactions of different populations to specific climate change.
However, despite the large number of studies run, it remains difficult to determine precisely the way trees may react to anticipate warming of the boreal and temperate climates (Matala et al. 2006; Scholze et al. 2006; Reich and Oleksyn 2008). That said, the results obtained hitherto suggest that even small changes will prove negative for what is Europe’s main forest-forming species—the Scots pine (Strömgren and Linder 2002; Reich and Oleksyn 2008).
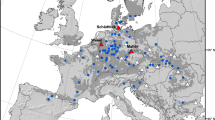
Even before interest in these matters was developing much, a provenance experiment relating to Scots pine was set up in Rogow. Originally, the main goal was to determine the growth and quality of populations from other climatic regions of the country in the conditions of Central Poland. Nowadays, such experiments can be used for preliminary research in the field of assisted migration of tree species in the context of occurring climate change. For this purpose, the De Martonne aridity index was used for the analysis. In the region of the provenance experiment, it takes values below the mean (36.3 mm/°C) compared to the location of the mother stands (from which the studied populations originate) and results from the occurrence of quite high temperatures with low rainfall in the region of Central Poland (Fig. 1, Table 1).

Locations of parental stands and a schematic representation of the distributions of the analysed provenances for Scots pine growing on the provenance trial at the Rogów Forest Experimental Station. Janow Lub.*—the abbreviation of Janow Lubelski provenance name
Furthermore, in the last decade or so in particular, Central Poland has come to experience very marked water-deficit effects in the face of a rise in average temperatures (Chojnacka-Ożga and Ożga 2018a, b). Therefore, the location of the provenance trial in such a place may increase the possibilities of assessing the adaptability of studied populations to the Central Poland conditions.
The study aim was to verify the research hypotheses, assuming that: (1) the progenies of populations growing in similar or worse conditions of precipitation deficiency and high temperature will better adapt to difficult conditions, and (2) the native populations adapt the best to local conditions on the provenance trial.
Materials and methods
As noted already, the experimental site referred to here was founded in 1966 in the context of the Rogow Forest Experimental Department. That lies within the Forest Sub-District of Lipce, within Compartment 44j (and hence at 51°54′29.1"N, 19°54′46.4"E). Relevant spatial information for the experimental site is detailed in Szeligowski et al. (2015). The experiment was established on what had been farmland, featuring lightly dusty clayey sands down to a depth of 40−55 cm. This is underlain by a dusty layer showing signs of podsolisation. All of this soil is highly acidic—and that applies down to depths of 160–180 cm. While the site type here was referred to as mesic mixed/coniferous forest (so-called BMśw) as of 1966, revision of the forestry management classifications has seen it re-categorised as mesic mixed/broadleaved forest (so-called LMśw) (Zielony 1993). Relevant meteorological information is as detailed in Table 1.
As measurement work on the provenance experiment was conducted in the autumn of 2015, it was in a position to take account of some 50 years of growth. Measurements were made of tree heights and breast-height diameters, with assessments also made of the straightness of trunks. Crown widths (at the base of the living crown) and branching angle (at the mid-point within the living crown) were also considered. Traits based on estimates used a classification system as follows: straightness of trunks: (1) much severe curvature, (2) one example of severe curvature, (3) curvature of average size, (4) one, or two minor, examples of curvature, (5) straight; crown width: (1) very narrow, (2) narrow, (3) average, (4) broad, (5) very broad; branching angle: (1) of less than 30°, (2) of between 31 and 40°, (3) of between 41 and 50°, (4) of between 51 and 60°, (5) above 61°.
The volumes of individual trees and stands were determined in-line with the formula:
where: g1,3 refers to the breast-height cross section, h is tree height, and f1,3 is steam form factor (Bruchwald and Rymer-Dudzińska 1998).
Long-term mean values for air temperature and atmospheric precipitation in the areas around maternal stands whose progeny were tested were obtained as grid data from the Climatic Research Unit database (CRU CL v. 2.0), with a spatial resolution of 10' (New et al. 2002). The synthetic description of climatic conditions in the places of growth of maternal stands was also achieved by taking note of the De Martonne aridity index (De Martonne 1926):
where: P is precipitation and T as mean temperature.
Measurable traits were characterised by calculated mean values as associated with their standard errors. The fallowing linear mixed model (LMM) was used for analysis:
where: Ykj is the observation, µ is the overall mean, Pk is the fixed effect of kth provenance, Bj is the random effect of jth block, and ejk is the error.
In contrast, estimated features were determined by reference to shares of trees assigned to the classes distinguished (qualitative data). This type of data were also analysed using the model presented above, except that a multinomial distribution was assumed for the observation values, thus creating a GLMM (generalized linear mixed model) model. The Tukey’s linear contrast was used to make pairwise comparisons between the provenance for the LMM and GLMM models.
The analysis of comparisons between the provenances was performed for all quality classes of the three examined features (straightness of trunks, width of the crown and angle of the branches). Due to the very large number of classes (15), detailed analyses are presented only for the best economically class for each feature: the share of trees in the straight trunks class (5), in the narrow crown class (2), and in the branch angle setting above 61° (class 5). The diversity of origins in terms of the share of trees in these classes was compared to the tree density at the age of 50 per 1 ha (trees/ha).
The relationships between all the traits used to describe tested provenances were then assessed using Principal Component Analysis (PCA). In order to demonstrate the similarity between the progeny, the actual values of weather conditions (long-term averages) characteristic for the maternal stands locations of the studied populations were used. Groups of provenances characterised by multi-trait similarity were then generated using Ward’s cluster analysis method, with the measure of similarity being the Euclidean distance squared. All calculations were made with the aid of the R 3.2.1 statistical package.
Results
After 50 years of growth on the study site, there was found to be considerable differentiation among the 16 provenances in mean values for breast-height diameter (DBH), height (H) and stand volume (V) (Table 2). The ranking in-line with stand volume included, in the most favourable situation, the pine progeny from Bolewice, where the figure attained was 459.2 m3 ha−1 of merchantable wood per hectare. High volume figures were also noted for the populations from Karsko (439 m3 ha−1), Rychtal (438.9 m3 ha−1) and Dluzek (434.6 m3 ha−1), while the lowest were noted for the progeny groups deriving from Rospuda (322.9 m3 ha−1), Janow Lubelski (319.2 m3 ha−1) and Nowy Targ (198.2 m3 ha−1). The studied populations differed statistically significantly in terms of the mean value of the volume. The separate group with a low value of the analysed feature were pines from Nowy Targ (Table 2). A similar degree of differentiation was noted for the DBH and height data, with both features achieving their highest values in the case of the Bolewice provenance (DBH = 226.8 mm, H = 23.2 m). This compared favourably with the poorest results for DBH—the 203.8 mm noted for Rospuda trees, as well as height—Jegiel (H = 21.7 m).
When it came to variability measured by reference to standard error, it was trees in the Gubin, Dluzek and Rychtal populations that varied least, and those from the Rospuda and Nowy Targ provenances that varied most. Likewise, standard errors associated with height were lowest for Gubin and Dluzek, and highest in the cases of Rospuda and Ruciane. Finally, the least variability in stand volume characterised the Karsko population; while the Rospuda and Janow Lubelski trees were at the other end of the spectrum from this point of view. Statistical analysis showed no significant differences in mean DBH. Four homogeneous groups were distinguished in the case of height (Table 2).
Trees with multidimensional curvatures (class 1) were found only in the Gubin and Jegiel populations, while trees with a very large one curvature (class 2) were recorded in eight populations (Fig. 2a). Participation in these classes did not exceed 2%. Class 3 (curvature of average size) comprised about 5–25% of trees, while pines with small curvatures (class 4), depending on the population, ranged from 40 to 65%. The straight-trunked pines, as the most valuable (class 5), ranged from 15 to 50%. Such trees were the most numerous in Rospuda, Starzyna, Rychtal, Dluzek and Suprasl populations (Fig. 3a). The differences between the share of trees with the most straight trunks in the populations were statistically significant (p < 0.05).

Structure [%] for trunk straightness, crown width, and mean angle between stem and branches among the trees studied provenances (1–5 traits classification described in chapter Materials and methods)

The share [%] of trees in the context of the analysed features a, b, c, and d
All populations turned out to be very similar in terms of crown width (Fig. 2b). The highest (over 20%) proportions of trees with very broad crowns (class 5) were obtained for the Bolewice, Lipowa, Spala, Karsko, Ruciane and Gubin provenances. The share of trees with broad crowns (class 4), depending on the population, ranged from 22 to 46%, with average crowns (class 3) from 26 to 41%, and with narrow crowns (class 2) from about 4 to 15%. In these four classes, no statistically significant differences were found between the shares of trees in individual populations. In turn, a figure of around 13% was the highest noted for the share of all trees in a population with very narrow crowns (class 1). This applied in the case of the populations from Janow Lubelski and Jegiel. No trees with very narrow crowns were found in the populations of Spala, Karsko and Rychtal (Fig. 3c).
Trees with branches departing from the trunk at a very acute (< 40°—class 1 and 2) angle were not noted in any of the provenances studied (Fig. 2c). The share of trees in class 3 (41° to 50°), depending on the population, ranged from 2 to 36%. In each provenance the greatest numbers (from 46 to 74%) of trees were allocated to class 4—meaning a branching angle ranged from 51° to 60°. The extreme examples with branches forming an angle greater than 61° were progeny from the Spała, Bolewice and Ruciane provenances (over 30% of share), while pines from Rychtal had the lowest, 11% share (Fig. 3b).
After 50 years of growth the tree density ranged from about 600 trees/ha (Nowy Targ) to almost 1200 trees/ha (Rychtal). The differences between the density of individual populations were statistically significant. Low densities, not achieving 1000 trees/ha, were also found in the following populations: Janow Lubelski, Rogow and Rospuda, while high density, over 1100 trees/ha, was found in the group: Suprasl, Karsko and Dluzek (Fig. 3d).
The Principal Component Analysis carried out revealed two first axes together explaining 63.61% of the multivariate variability present among trees of the different provenances. PC1 was able to account for 43.7% of variability (Fig. 4, Table 3) and proved to be correlated most strongly with the De Martonne aridity index, as well as altitude and annual precipitation totals. In turn, there were inverse correlations noted with mean annual temperature, stand volume and length of the growing season. The second component (PC2) was able to explain a further 19.9% of variability, with the strongest direct correlation being with tree heights and the location along the gradient of altitude above sea level. In turn, the component displayed a negative correlation with latitude, age of the maternal stand, longitude and straightness of the trunk.

Distribution of the 16 provenances in-line with the first two Principal Components
Multifactor cluster analysis distinguished a first group (comprising the local-origin Rogow trees as well as those representing the Spala provenance) whose characteristic traits were a high figure for breast-height diameter (of 215 mm), average values for tree height and stand volume (22.6 m and 372.7 m3/ha, respectively), and low values for straightness (i.e. a large share of trees presenting curvature) (Table 4, Fig. 5). When set against those in other groups, the pines of group 1 had average values for coefficients when it came to crown width (3.50) and branching angle (4.14). A second group comprised eight provenances (of Rospuda, Starzyna, Suprasl, Dluzek, Taborz, Ruciane, Jegiel and Lipowa)—with features here being low values for DBH (211 mm) and height (22.2 m), as well as an average figure for volume (373.9 m3/ha). The mean value for the coefficient of trunk straightness is high in this group—at 4.29, while there are intermediate values for crown width and branching. Group 3 in turn comprises the Bolewice, Karsko and Gubin populations, which are characterised by the largest dimensions and a high figure for volume, even as trunk straightness is less well-marked than in other groups (meaning large numbers of curved trees), which the coefficient for crown width is high and the branching angle is low. The remaining groups (4, 5 and 6) are represented by single populations only, with number 4 (Rychtal) characterised by the highest stand volume (of 438.9 m3/ha), as well as the highest values for the coefficients describing straightness of the trunk and crown width, as well as the lowest value for branching angle (denoting a sharp angle). Group 5 comprises the population from Janow Lubelski only—with intermediate values for DBH and height (213 mm and 22.5 m), as well as low figures for volume (319.2 m3/ha), an average level of trunk straightness (a value for the relevant coefficient equal to 4.22), and broadest crowns (3.13), as well as a very high value for the coefficient relating to branching angle (4.24).

Grouping (1–6) of the studied provenances of Scots pine by reference to the features in Table 3
For its part, the sixth group included the population from Nowy Targ—featuring the lowest values for growth parameters and volume (the latter at just 198.2 m3/ha). The trees of this population are of intermediate straightness and crown width, though they have the highest mean values for the coefficient of branching angle. For this lowest-classified provenance, environmental conditions at the place where the maternal stand is located feature large totals for annual precipitation, a high altitude above sea level, a high value for the De Martonne index, a low value for mean annual temperature and a short growing season.
Discussion
Intensifying climatic warming in Europe is contributing to impairment in the immunity of forest-trees species, including also the Scots pine—notwithstanding the considered status of this tree as drought-resistant. As the pine is not noted for its thermal requirements, forecast rises in temperature may lead to a retreat from certain sites or areas occupied at present. Were such scenarios to arise that would unavoidably denote a need to facilitate migration of more drought-resistant populations (Seidel et al. 2016). This could be accomplished by transferring young trees from drier and warmer areas to regions where the climate anticipated will be similar (Bussotti and Pollastrini 2015). This kind of assisted or facilitated migration would prove especially important where the same species is present over a very extensive range—across which climatic conditions can differ quite markedly. This is of course the case for the Scots pine, which is currently Poland’s key tree species from the economic point of view, accounting for almost 60% of the entire area of forest (State Forest Report 2020). There is therefore justification for a seeking-out of the populations most suited to cultivation under conditions anticipated for the modified environments of the near-future.
Indeed, the ecological and economic significance of the Scots pine is particularly great given the lack of other species of anything like equal (economic and natural) importance in circumstances of poor soils. In the view of Sabor and Stachnik (1990), as well as Giertych (1997), Scots pines across Poland are generally characterised by continuous variation, with one population differing from another in-line with differences in environmental conditions—which are sufficient to shape features in the species.
It was in-line with this thinking that the progeny of pines of 16 different provenances were tested in the experiments described above—and also found to be characterised by considerable variation in DBH, height and qualitative features. In the light of the results for genetic differentiation presented by Nowakowska (2003), as well as Hebda et al. (2017a), this is a known phenomenon, and one that mainly reflects the in-migration of the Scots pine into what is today Poland from refugia areas elsewhere in Europe.
While Giertych (1997) and Barzdajn et al. (2016) were of the view that provenance experiments demonstrate the superiority of local populations of pine, our work on the research area at Rogow could not confirm that; and nor did work carried out at a site in Polany, where the Piwniczna population is regarded as local (Sabor 1993; Hebda et al. 2017a, b). Our analysis reveals that at-best average values are achieved by the population from Rogow itself (as well as the closest outside population from Spala). Indeed, 44 years of assessment of the silvicultural value of trees with both of these origins has already pointed to their limited value (Szeligowski et al. 2015). This is one of the reasons for it to seem entirely justified and worthwhile to seek out populations of greater value under the conditions of Central Poland than those regarded as local. Furthermore, the search effort involved obviously benefits from information on the climatic conditions prevailing in the area of occurrence of the parent populations whose progeny are under study on the provenance experiments.
Thus, when it comes to the De Martonne aridity index, values similar to those at the trial site also characterised maternal stands hosting the Spala, Jegiel, Janow Lubelski and Starzyna provenances. Beyond that far less precipitation actually falls in the circumstances of the climate at Bolewice, Gubin, Karsko and Rychtal, which are all in western Poland. The high stand volumes characterising these populations in the study area serve to confirm their suitability for silviculture in the central part of the country—and this can be justified, not purely from the economic point of view, but also in-line with the adaptability to worsening climatic conditions. In the light of the latter, it may be that typical qualitative traits of trees (like straightness of trunk, breadth of crown or branching angle) may come to look less significant than those relating to increment and survival.
Conclusions similar to the above were reached in the case of the provenance trial in Suprasl (as an area within the IUFRO 1982 series that tested populations of Scots pine representative of the entire European range of the species). Barzdajn et al. (2016) thus showed how populations transferred in from the boreal zone (i.e. the northern part of the range in Russia, Latvia and Sweden)—though characterised by lower growth rates, earlier cessation of main trunk growth and more limited dimensions in general—did have considerable potential for adaptation to the warmer climate of Central Europe.
In turn, in the circumstances of Poland, a southern European provenance denoted lower rates of survival, together with productivity. As with our analysis, local populations were included among the provenances showing good growth—but not in fact the best of all. For their part, Seidel et al. (2016) reported that, under conditions of moderate drought, 2-year-old saplings of Scots pine of southern (Spanish, French or Bulgarian) provenances were characterised by lower stress levels and more favourable growth rates following stress than were found to characterise the populations from Poland and Germany (Central Europe).
Provenance studies with Scots pine further reveal how—in the conditions of Central Poland—the highest values for features of growth are reported for populations originating at latitudes between 49 and 53°N, even as height, breast-height diameter and figures for the accumulation of above-ground and root biomass were correlated significantly with the latitude of the parental populations (Matras 1989; Oleksyn et al. 1999, 2000; Barzdajn et al. 2016). Our research showed that, within the limits for latitude given above, provenances from the Western Poland area adapt better to the climatic conditions of Central Poland than an origin in the east or north of the country. This links up with the lower precipitation totals and lower values for the De Martonne aridity index characteristic for maternal stands in the west of the country.
The secondary ecological importance of quality features (trunk straightness, crown width, and branching angle) does not release the need to include them in the analyses. Earlier studies (Giertych 1986, 1988; Sygit and Giertych 1995) indicated that the best in terms of trunk straightness, i.e. feature of the most important economic significance, are the populations of Dluzek, Ruciane and Suprasl. Our analysis revealed that in the Central Polish conditions, in terms of this feature, the populations from Rospuda and Starzyna are the best. On the other hand, the populations mentioned by Giertych (1986, 1988), especially Dluzek and Suprasl, were also characterised by a large share of straight trees. Wide crowns of pines increase the weight of the assimilation apparatus responsible for biomass production and at the same time reduce the quality of the wood raw material (poor removal of branches from the trunks). An example of such a population is Bolewice, whose progeny had similar properties also on the Polana study plot (Hebda et al. 2017b). In addition, the results of numerous international and national provenance studies on the racial variability of Scots pine indicate the good quality of the Masurian pine race (including the Ruciane, Suprasl and Taborz populations) and the West Pomeranian pine race (Bolewice and Rychtal populations) (Giertych 1979, 1980, 1997; Kowalczyk 2005). Also in the international provenance experiment established in Bosnia and Herzegovina (testing fifteen populations from the Scots pine range), the Ruciane population as a representation of Poland was the highest (Ballian et al. 2019). Considering the above, it can be clearly stated that in the conditions of Central Poland, selective thinning focused on high quality of wood material can be successfully carried out in north-eastern populations. The management in western pine populations should be based on the biomass of stands.
The population of Janow Lubelski is also noteworthy, where a lot of trees with very narrow crowns were found, despite one of the lowest densities on the research site (847 trees/ha). Zajączkowska et al. (2020) noted the distinctiveness of the pine from Janow Lubelski. The author suggested that apart from acclimatization factors, the reason for the different anatomical structure of the needles of this population may be its origin from the southern migration (Alpine and Balkan refugium).
PCA multivariate analysis provided for the description of a number of relationships between features of the progeny and climatic factors determining growth of trees in the maternal stand. It also supported a particular grouping of these in-line with similarity in respect of growing in similar climatic conditions. One key relationship related to the fact that the greater the annual precipitation total and higher the values of the De Martonne aridity index (as well as the greater the altitude above sea level of the maternal population), the lower the values obtained for features measured among progeny growing at the site in Central Poland (i.e. volume, breast-height diameter and height).
It was in this way possible to confirm a thesis advanced by Giertych (1979), as well as Sygit and Giertych (1995), to the effect that pines from sites at altitudes above 200 m a.s.l. are not suitable for cultivation on the Polish lowlands.
The cluster analysis also confirmed multi-trait similarities between populations whose parental stands are in close proximity with one another. An example would be combined groups of three provenances from Western Poland (i.e. Bolewice, Gubin and Karsko), as well as 7 from the north-east. The analysis further confirmed the similarity of stands made up of individuals from Rogow and Spala, while also succeeding in distinguishing the Nowy Targ and Janow Lubelski populations.
Conclusions
With climate change in prospect, there is a possibility of reacting by transferring progeny from populations of Scots pine around the country, but this will only be indicated if places of origin are those in which the climate has lower levels of precipitation and lower values for the De Martonne aridity index. The analyses have revealed that there is a possibility of pine progenies transfer from populations located in the western part of the country, such as Bolewice, Rychtal, Karsko and Gubin, to Central Poland. It should also be emphasized that the very poor survival rate and population growth from Nowy Targ confirms the assumptions known from the literature that the progeny of pines from higher altitude stands are not suitable for cultivation in lowland areas of Poland.
However, selection of the best populations for transfer will be possible after performing similar analyses on parallel provenance trials from the same series of experiments. Another four experimental sites that test the same populations are located in different climatic regions of Poland and are under the scientific supervision of other research institutions. A comprehensive analysis of data from Fife provenance trials will bring new knowledge about the studied populations adaptability to various climatic conditions and should indicate many more possibilities for the assisted migration of Scots pine populations in Poland.
Data availability
All data generated or analysed during this study are included in this article and are available from the corresponding author upon reasonable request.
Code availability
Not applicable.
References
Ballian D, Lizdo E, Bogunić F (2019) Analysis of differences of growth and phenology of provenances of Scots pine (Pinus sylvestris L.) in provenance experiment at Kupres (Bosnia and Herzegovina). Šumarski List 143(1–2):25–34. https://doi.org/10.31298/sl.143.1-2.3
Barzdajn W, Kowalkowski W, Chmura DJ (2016) Variation in growth and survival among European provenances of Pinus sylvestris in a 30-year-old experiment. Dendrobiology 75:67–77. https://doi.org/10.12657/denbio.075.007
Bruchwald A, Rymer-Dudzińska T (1998) Empirical formulae for defining the pine over bark d.b.h. form factor with account on natural forest regions. Sylwan 142(8):5–13
Bussotti F, Pollastrini M (2015) Evaluation of leaf features in forest trees: methods, techniques, obtainable information and limits. Ecol Indic 52:219–230. https://doi.org/10.1016/j.ecolind.2014.12.010
Chodzicki E (1975) Preliminary results of comparative plantation of Scots pine (Pinus sylvestris L.) of different Polish provenances in the lower montane belt of the Beskid Sądecki. Sylwan 119:1–13
Chojnacka-Ożga L, Ożga W (2018a) Trends in the length of the thermal growing season in the north−eastern Poland. Sylwan 162:479–489. https://doi.org/10.26202/sylwan.2018a022
Chojnacka-Ożga L, Ożga W (2018b) Air temperature anomalies in experimental forests in Rogow in 1924–2015. For Res Pap 79:37–44. https://doi.org/10.2478/frp-2018-0005
Davis MB, Shaw RG, Etterson JR (2005) Evolutionary responses to changing climate. Ecology 86:1704–1714. https://doi.org/10.1890/03-0788
De Martonne E (1926) Une nouvelle fonction climatologique: l ́indice d ́aridité. La Météorologie 21:449–458
Eilmann B, Zweifel R, Buchmann N, Fonti P, Rigling A (2009) Drought-induced adaptation of the xylem in Scots pine and pubescent oak. Tree Physiol 29:1011–1020. https://doi.org/10.1093/treephys/tpp035
Giertych M (1979) Summary of results on Scotch pine (Pinus sylvestris L.) height growth in IUFRO provenance experiments. Silvae Genet 28:136–152
Giertych M (1980) Polish races of Scots pine, Norway spruce and European larch in international provenance experiments. Arboretum Kórnickie 25:135–160
Giertych M (1986) Provenance variation of Scots pine (Pinus sylvestris L.) on a 46-years old international experiment in Poland. Arboretum Kórnickie 31:183–193
Giertych M (1988) Interaction of genotype with environment and age for Polish provenances of Scots pine (Pinus sylvestris L.). Arboretum Kórnickie 33:159–169
Giertych M (1997) Variability on Polish Provenances of Scots Pine in Poland. Sylwan 141:5–20
Hebda A, Skrzyszewski J, Wachowiak W (2017a) Phenotypic differentiation and genetic background variation of Polish provenances of Pinus sylvestris L. Sylwan 161:277–286. https://doi.org/10.26202/sylwan.2016135
Hebda A, Wachowiak W, Skrzyszewski J (2017b) Long-term growth performance and productivity of Scots pine (Pinus sylvestris L.) populations. Acta Soc Bot Pol 86:3521. https://doi.org/10.5586/asbp.3521
Kowalczyk J (2005) Assessment of the results of provenance research on Scots pine (Pinus sylvestris L.). In: Materials from international scientific and technical conference, Milanówka, June 2005, Conservation of forest tree genetic resources in Poland- state and perspectives, Świat Pub, Warsaw, Poland, pp 132–141
Matala J, Ojansuu R, Peltola H, Raitio H, Kellomäki S (2006) Modelling the response of tree growth to temperature and CO2 elevation as related to the fertility and current temperature sum of a site. Ecol Model 199:39–52. https://doi.org/10.1016/j.ecolmodel.2006.06.009
Matras J (1989) Provenances studies on Scots pine conducted by the section of seed science and selection of the forest research institute. Sylwan 133:53–66
Misi D, Náfrádi K (2017) Growth response of Scots pine to changing climatic conditions over the last 100 years: a case study from Western Hungary. Trees 31:919–928. https://doi.org/10.1007/s00468-016-1517-z
Misi D, Puchałka R, Pearson C, Robertson I, Koprowski M (2019) Differences in the climate-growth relationship of Scots Pine: a case study from Poland and Hungary. Forests 10:243. https://doi.org/10.3390/f10030243
Naydenov K, Senneville S, Beaulieu J, Tremblay F, Bousquet J (2007) Glacial vicariance in Eurasia: mitochondrial DNA evidence from Scots pine for a complex heritage involving genetically distinct refugia at mid-northern latitudes and in Asia Minor. BMC Evol Biol 7:233. https://doi.org/10.1186/1471-2148-7-233
New M, Lister D, Hulme M, Makin I (2002) A high-resolution data set of surface climate over global land areas. Clim Res 21:1–25. https://doi.org/10.3354/cr021001
Nowakowska J (2003) Genetic diversity of Scots pine (Pinus sylvestris L.) Polish provenances based on RAPD analysis. Sylwan 147:25–37. https://doi.org/10.26202/sylwan.2003229
Oleksyn J, Reich PB, Tjoelker MG, Chałupka W (1999) Differential above- and below-ground biomass accumulation of European Pinus sylvestris populations in a 12-year-old provenance experiment. Scand J for Res 14:7–17. https://doi.org/10.1080/02827589908540804
Oleksyn J, Reich PB, Rachwał L, Tjoelker MG, Karolewski P (2000) Variation in aboveground net primary production of diverse European Pinus sylvestris populations. Trees 14:415–421. https://doi.org/10.1007/PL00009775
Rehfeldt GE, Tchebakova NM, Parfenova YI, Wykoff RA, Kuzmina NA, Milyutin LI (2002) Intraspecific responses to climate in Pinus sylvestris. Glob Change Biol 8:912–929. https://doi.org/10.1046/j.1365-2486.2002.00516.x
Reich PB, Oleksyn J (2008) Climate warming will reduce growth and survival of Scots pine except in the far north. Ecol Lett 11:588–597. https://doi.org/10.1111/j.1399-3054.2006.00682.x
Sabor J (1993) Variation of Scotch pine (Pinus sylvestris L.) and the breeding value of its Polish provenances in the habitat conditions of the Sącz Beskid range. Zeszyty Naukowe Akademii Rolniczej im. H Kołłataja w Krakowie Rozprawy 185:1–186
Sabor J, Stachnik E (1990) Survival and growth of various Scots pine provenances in site conditions of Sącz Beskid exemplified on comparative area in Polany near Grybów. Sylwan 134:11–26
Savva JV, Vaganov EA (2006) Genetic and environmental effects assessment in Scots Pine Provenances planted in Central Siberia. Mitigation Adapt Strategies Global Change 11:269–290. https://doi.org/10.1007/s11027-006-1026-2
Seidel H, Schunk Ch, Matiu M, Menzel A (2016) Diverging drought resistance of Scots Pine provenances revealed by infrared thermography. Front Plant Sci 7:1247. https://doi.org/10.3389/fpls.2016.01247
Scholze M, Knorr W, Arnell NW, Prentice IC (2006) A climate-change risk analysis for world ecosystems. Proc Natl Acad Sci USA 103:13116–13120. https://doi.org/10.1073/pnas.0601816103
State Forest Report. State Forest Information Centre: Warsaw, Poland, 2020. https://www.lasy.gov.pl/pl/informacje/publikacje/informacje-statystyczne-i-raporty/raport-o-stanie-lasow/raport-o-stanie-lasow.pdf/view. Accessed on 01.12.2020
Strömgren M, Linder S (2002) Effects of nutrition and soil warming on stem wood production in a boreal Norway spruce stand. Global Change Biol 8:1195–1204. https://doi.org/10.1046/j.1365-2486.2002.00546.x
Sygit W, Giertych M (1995) Differentiation of Polish provenances of Pinus sylvestris L. on the experimental area in Kórnik. Arboretum Kórnickie 40:87–105
Szeligowski H, Buraczyk W, Drozdowski S, Gawron L (2015) Silvicultural value of Scots pine provenances from Poland on the experimental plot in Rogow. Sylwan 159:997–1007. https://doi.org/10.26202/sylwan.2015050
Wang T, Hamann A, Yanchuk A, O’Neill GA, Aitken SN (2006) Use of response functions in selecting lodgepole pine populations for future climates. Global Change Biol 12:2404–2416. https://doi.org/10.1111/j.1365-2486.2006.01271.x
Zajączkowska U, Piątkowski M, Dolkin A, Buraczyk W (2020) Diversity of needles structure of the selected Polish populations of Pinus sylvestris L. on the experimental plots in the Forest Experimental Station in Rogow. Sylwan 164:842–849. https://doi.org/10.26202/sylwan.2020051
Zielony R (1993) Warunki przyrodnicze lasów doświadczalnych SGGW w Rogowie/Natural conditions of experimental forests of WULS in Rogow. WULS Pub, Warsaw, Poland
Funding
This article was prepared as part of the project “Spectroscopic Methods for Rapid Phenotyping of Trees Reflecting their Ecological Resilience” acronym DENDROSPEC, financed by the National Science Centre Poland (NCN) No UMO-2021/43/I/NZ9/02809.
Author information
Authors and Affiliations
Contributions
Conceptualization was contributed by HS; methodology was contributed by HS and WB; data collecting was contributed by HS, WB, AK and SD; formal analysis and figures preparation were contributed by HS, WB, AK, MS and SD; writing—original draft preparation was contributed by HS; writing—review and editing was contributed by WB, AK and SD. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest.
Additional information
Communicated by Oliver Gailing.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Szeligowski, H., Buraczyk, W., Konecka, A. et al. A multi-trait assessment of selected provenances of Scots pine following 50 years of growth on a provenance experiment in Central Poland, in the light of climate change. Eur J Forest Res 142, 509–520 (2023). https://doi.org/10.1007/s10342-023-01538-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-023-01538-z