
Abstract
Unintentional introduction of natural enemies has increased in recent years due to a massive rise in global trade and tourism. One such natural enemy is the Asian egg parasitoid Trissolcus japonicus, a promising agent for classical biological control of Halyomorpha halys. In Europe, adventive T. japonicus populations have been detected in Switzerland, Italy and Germany. Host specificity testing demonstrated that its fundamental host range is fairly broad; however, it is unclear whether spatial or temporal refuges reduce parasitism of non-target species in the field. To address this, the realized host range of T. japonicus was assessed over three years by exposing sentinel egg masses of H. halys and 18 non-target species and collecting naturally laid egg masses in Switzerland and Italy. In total, 15 of 18 non-target species were successfully parasitized by T. japonicus in the field, confirming its broad fundamental host range. However, most non-target species were less parasitized by T. japonicus than H. halys, profiting from either partial temporal or spatial refuges from parasitism. Species with an unusual life cycle and the same ecological niche as H. halys, such as Pentatoma rufipes, which was the most parasitized non-target species in both countries, potentially face an increased risk of parasitism. In contrast, beneficial non-target effects may occur for the invasive pest, Nezara viridula, which suffered high non-reproductive mortality induced by T. japonicus. In both cases, life table studies will be needed to determine the impact of non-target parasitism and the potential consequences at the population level.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Key message
-
The Asian parasitoid Trissolcus japonicus associated with Halyomorpha halys is spreading in Europe
-
Its realized host range was assessed in Switzerland and Italy using sentinel and wild egg masses
-
Species in the same ecological niche as H. halys are more likely to be attacked by T. japonicus
-
DNA analysis of Nezara viridula eggs showed non-reproductive mortality caused by T. japonicus
-
Pentatoma rufipes is regularly attacked, but the impact at population level remains unknown.
Introduction
The brown marmorated stink bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) is native to Asia and arrived in North America and Europe in the late 1990s and early 2000s, respectively (Leskey and Nielsen 2018). The first records from Europe (Switzerland, Liechtenstein) date back to 2004 (Hoebeke and Carter 2003; Arnold 2009; Haye et al. 2014), but at least two additional, independent introductions were later detected in Italy and Greece (Gariepy et al. 2015; Cesari et al. 2018). Since its arrival, it has spread through most of Europe’s mainland (Claerebout et al. 2018), and due to climate change, it has the potential to further expand its range (Kriticos et al. 2017; Stoeckli et al. 2020). Because of its polyphagous behaviour, H. halys quickly became an invasive pest of a wide variety of tree fruit, nut, vegetable, and field crops in Europe, particularly in Italy and Georgia, where it has caused severe economic losses in tree fruit (e.g. apples, peaches, pears) and nuts (hazelnuts) (Maistrello et al. 2017; Bosco et al. 2018).
As the impact of native natural enemies on invasive H. halys populations in Europe and North America is generally low (Abram et al. 2017), H. halys was identified as promising target for classical biological control in the invaded range. Surveys for natural enemies that have co-evolved with H. halys in its native range revealed that it is mostly attacked by egg parasitoids, among which the samurai wasp, Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae), was identified as the most promising candidate for classical biological control, with parasitism levels ranging from 30 to 90% (Yang et al. 2009; Lee et al. 2013; Zhang et al. 2017).
However, prior to the release of exotic biological control agents into a new environment, it is important to consider the potential for unintended non-target effects such as host range expansion to native or beneficial species (van Lenteren et al. 2003). Accordingly, host specificity testing of T. japonicus has been carried out under containment conditions in North America and Europe (Hedstrom et al. 2017; Botch and Delfosse 2018; Haye et al. 2020; Sabbatini Peverieri et al. 2021). Overall, these studies came to similar conclusions that T. japonicus’ fundamental (physiological) host range is restricted to the Pentatomoidea, but the parasitoid is capable of successfully parasitizing the eggs of several pentatomids other than H. halys, as well as a few species of scutellerids. These studies also showed that most non-target species, compared to H. halys, were usually accepted at lower rates and were less suitable for development of T. japonicus offspring. In addition, incorporating more complexity and the parasitoid’s host finding behaviour in laboratory experiments (host chemical footprints, cage trials with plants) seemed to further increase the likelihood that H. halys, more often than non-target species, was detected and parasitized. In Europe, developmental suitability of non-target host species for T. japonicus was demonstrated in no-choice tests by successful offspring emergence from 11 of 13 non-target species tested (Haye et al. 2020). Whereas most non-target species were less accepted than H. halys, four native heteropteran species [Palomena prasina (L.), Rhaphigaster nebulosa (Poda), Pentatoma rufipes (L.), and Arma custos (F.)] were accepted at rates comparable to the target host. In a later study, nine out of 16 non-target species were accepted and suitable for T. japonicus development, confirming the potentially higher risk of non-target parasitism for P. prasina and R. nebulosa (Sabbatini Peverieri et al. 2021).
As laboratory risk assessment studies for T. japonicus in Europe were underway, adventive (self-introduced) populations of T. japonicus were discovered in Switzerland, Italy, and Germany (Sabbatini Peverieri et al. 2018; Stahl et al. 2019b; Moraglio et al. 2020; Dieckhoff 2021). A large-scale survey conducted in northern Italy and parts of Switzerland in 2019 demonstrated that within just three years, T. japonicus has rapidly spread into all types of habitats where H. halys is present (Zapponi et al. 2021). There is no doubt that T. japonicus will continue to quickly expand its range in Europe as predicted by bioclimatic envelope models (Avila and Charles 2018).
Accurately reproducing the complex factors that influence host searching and acceptance behaviour of parasitoids in their natural environment can be difficult. As such, the fundamental host range of a biological control agent is often greater than its realized (ecological) host range, which is the current and evolving set of host species used for successful reproduction in nature (Cameron and Walker 1997; Morehead and Feener 2000; Froud and Stevens 2003; Haye et al. 2005; Barratt et al. 1997, 2010; Cameron et al. 2013). Assuring host specificity from laboratory testing remains challenging, and it has been advocated that laboratory observations should be combined with field observations to provide a basis for correctly interpreting fundamental host range estimations (Onstad and McManus 1996; Hopper 2001; Kuhlmann and Mason 2003). After completing fundamental host range studies for T. japonicus in Europe, it remained unclear if there are behavioural or phenological barriers to parasitism, such as habitat preferences, competition with native egg parasitoids, or oviposition periods of non-target species, which may exist and reduce parasitism of those species that were identified as highly suitable hosts in laboratory tests. However, the unintentional introduction of T. japonicus into Europe now provides a unique opportunity to validate retrospectively estimates of fundamental host range with realized ‘post arrival’ host range as it manifests over time, and determine whether enemy-free space exists in nature, and provides protection to species considered suitable under laboratory conditions.
The objective of the present study was to determine the realized host range of T. japonicus by exposing sentinel egg masses of H. halys and native non-target species in areas of Switzerland and Italy where the parasitoid was first established and compare these results with its predicted fundamental host range (Haye et al. 2020; Sabbatini Peverieri et al. 2021). As sentinel egg masses are sometimes less exploited in the field (Jones et al. 2014), naturally laid egg masses were also collected in order to assess parasitism and parasitoid species composition under more natural conditions. In addition, because egg masses of the invasive Nezara viridula (L.) (Hemiptera: Pentatomidae) suffered from high (non-reproductive) host mortality induced by T. japonicus in laboratory tests (Haye et al. 2020), we aimed to quantify this often neglected aspect of non-target impact under field conditions.
Material and methods
Selection, source and rearing of stink bug species
To assess the realized host range of T. japonicus in Europe, H. halys and 12 non-target species tested by Haye et al. (2020) [Pentatomidae: Acrosternum heegeri Fieber, A. custos, Carpocoris fuscispinus (Boheman), Dolycoris baccarum (L.), Graphosoma lineatum (L.), N. viridula, P. prasina, P. rufipes, Peribalus strictus (F.), Piezodorus lituratus (F.), R. nebulosa; Scutelleridae: Eurygaster maura (L.)] were selected for exposure of sentinel egg masses. In addition, three pentatomids [Carpocoris mediterraneus Tamanini, Carpocoris purpureipennis (De Geer), Eurydema ventralis Kolenati] tested by Sabbatini Peverieri et al. (2021) and three species that had not been part of previous laboratory testing [Pentatomidae: Eysarcoris venustissimus (Schrank), Eurydema ornata (L.); Scutelleridae: Eurygaster austriaca (Schrank)] were added to the test list, for a total of 18 non-target species.
Stink bug colonies (Hemiptera: Pentatomidae, Scutelleridae) were first established in the laboratories of DISAFA (Italy) and CABI (Switzerland) in 2018–2019 from field-collected adults as previously described (Moraglio et al. 2020, 2021a, b; Haye et al. 2020). In subsequent years, new adults were collected in the same way and at the same sites to maintain the laboratory colonies or to establish new ones. Species were identified using the keys by Derjanschi and Péricart (2005), Belousova (2007), Péricart (2010), Wyniger and Kment (2010), Ribes and Pagola-Carte (2013), Lupoli et al. (2013), and Neimorovets (2020). Adults were then reared, each species separately, in gauze cages (BugDorm-4090 Insect Rearing Cage, 47.5 × 47.5 × 47.5 cm; MegaView Science, Taichung, Taiwan) with the bottoms covered with white absorbent paper. At DISAFA, herbivorous pentatomids were fed with broad bean and fennel seedlings, hazelnuts without shells, and apples. At CABI, pentatomids were fed with corncobs, green beans, sunflower seedlings, a mix of wild flowers and fruit-bearing branches of common ivy. Diet for predatory pentatomids was complemented with adults and larvae of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae). Scutellerids collected on wheat were supplied with wheat ears (Feekes growth stages: 10–11; Miller 1992) and wet cotton. All food sources were periodically replaced. Mass rearing was carried out in controlled climatic chambers at 24±1 °C, 65±5% RH, and 16L:8D.
Exposure sites for sentinel egg masses
In Italy, egg masses of H. halys and non-target species were exposed in Piedmont (NW Italy), from April to October in 2019 and from May to September in 2020 in one of the first sites where T. japonicus was found in 2018 (Moraglio et al. 2020). The site, located in a peri-urban area near Turin (Grugliasco, 45.064358, 7.591715), is a park with several stink bug host plants, such as maple, poplar, mulberry, elder, and others, surrounded by crops (hazelnut, pome and stone fruits, wheat).
In Switzerland, stink bug egg masses were exposed at three sites in the Cantons Basel-Stadt, Zurich, Bern, and Ticino between May and September from 2019 to 2021. In the canton Ticino, eggs were exposed in an organic apple orchard and bordering natural areas in the centre of Manno (46.031119, 8.922250), where T. japonicus was first discovered in Europe in 2017 (Stahl et al. 2019b). In the city of Basel, eggs were exposed at the St. Alban Anlage (47.552206, 7.599038), an urban park with a high tree diversity (including maple and catalpa) that has been highly infested with H. halys over the past nine years. In Zurich, eggs were exposed in an urban park (‘Zürichhorn’, 47.354245, 8.552178) on the north shore of the lake of Zurich, where H. halys was first found in Europe in 2007 (Wermelinger et al. 2008). At both urban sites, the presence of T. japonicus was confirmed in the first year of the study, and accordingly, the exposure of non-target species was only started at end of summer 2019. Additional small exposures were conducted at an urban site in the city of Bern (46.963518, 7.447849), where T. japonicus was first discovered in the last year of the study (2021).
Preparation and field exposure of sentinel egg masses
Rearing cages were checked daily for freshly laid egg masses. Egg masses laid on the absorbent paper or on fennel leaves were collected with the substrate on which they were laid and glued on a green plastic tag (16 × 1.3 cm PVC plant labels, Hortima AG, Hausen, Switzerland) with a thin layer of neoprene adhesive (Bostik® Superchiaro, UHU Bostik S.p.A., Milan, Italy). Egg masses laid on the cage gauze or on broad bean and ivy leaves were gently removed from the substrate on which they were laid, and then directly glued on the tag, because the egg masses would easily detach from the drying leaves once they were cut, and consequently get lost. Number of glued egg masses of each species and the number of intact eggs per mass were counted. Empty, sucked, broken or collapsed eggs were not included and removed when possible.
Italy: Freshly laid egg masses were collected, glued, and exposed daily from Monday to Friday between April and October in 2019 and 2020, according to their availability (ESM1). The prepared egg mass tags were immediately hung on the host plants on which the bugs were, or could be, present at that time. Eggs of E. maura and E. austriaca were exposed on wheat. Eggs of R. nebulosa were exposed on Populus alba L. (April) and on Prunus avium (L.) (May and June), and eggs of N. viridula were exposed on Morus alba L. until the end of June and on Sambucus nigra L. from July onwards. Eggs of all other bug species were exposed on P. avium and M. alba from April until the end of June and on Acer spp. from July to October. Tags were attached with a plastic ring to branches of selected trees at ca 1.5–2 m height and on wheat to the stems under the ears. Exposed egg masses were no older than 72 hours and recollected after 2–4 days. In 2019, additional potted broad bean and fennel plants with egg masses laid directly on the underside of their leaves were exposed on a support placed under the trees on which egg tags were exposed. Overall, in NW Italy, 135 H. halys egg masses and 574 egg masses of non-target species (including 24 and 21 exposed directly on the potted bean and fennel plants, respectively) were exposed in 2019, whereas 37 H. halys and 726 egg masses of non-target species were exposed in 2020 (Table 1).
Switzerland: Newly laid egg masses were collected on a daily basis and refrigerated at 6 °C until exposure (no longer than 14 days), which means they were no longer viable for nymph development, but remained viable for parasitoid development (Wong et al. 2021). This was necessary, as field sites in Basel, Zurich and Ticino, where T. japonicus occurred naturally, were too far away for continuous egg exposure like in Grugliasco. In Basel and Zurich, tags with glued egg masses of bug species living on trees, i.e. N. viridula, P. prasina, P. rufipes, and R. nebulosa, were hung on cigar trees (Catalpa bignonioides Walter), Asian maple (Acer palmatum Thunberg), tulip trees (Liriodendron tulipifera L.), beech (Fagus sylvatica L.) and holly (Ilex aquifolium L.). Tags were attached to branches with a plastic ring at ca. 3m height. Egg masses of these four species were exposed in parallel on the same trees and if possible in the same numbers (no more than 5 egg masses per species/per tree). Egg masses of all other species, which feed primarily on herbs, were exposed in weedy patches or wild flower meadows within or in the vicinity of the parks. Plastic tags were attached to the weeds with paper clips. As the presence of T. japonicus was only discovered in Basel and Zurich by chance in the first year of the study, only egg masses of H. halys and P. rufipes were exposed in late August 2019. In the Ticino, egg masses of H. halys and non-target species were exposed in parallel on plastic tags and potted plants (Vicia faba L., A. palmatum, Fragaria x ananassa Duchesne, Hedera helix L.) in the first year of the study (2019). Potted plants were either placed underneath the trees or hung in the apple trees at 1.5 m height. Since egg parasitism on tags and potted plants did not differ but predation on eggs was much higher on potted plants, from 2020 onwards, eggs were only exposed on plastic tags. Eggs of bug species living on trees were either exposed on apple trees within the orchard or on trees (Robinia pseudoacacia L., Acer pseudoplatanus L. and Corylus avellana L.) in natural habitats nearby. Egg masses of all other species were exposed on weeds growing underneath the apple trees or on weeds in a wild flower meadow 150m southeast of the orchard, where non-target species naturally occurred. Towards the end of the study in 2021, T. japonicus had also dispersed to the city of Bern, and accordingly, eggs of H. halys and P. rufipes were exposed in Bern on maple trees in August/September of the same year. All sentinel egg masses were recollected after 4 days and brought back to the laboratory. The number of exposures at each site depended on the year, bug species and availability of egg masses (ESM1). Since all native species used in the study are univoltine and, with the exception of P. rufipes, overwinter as adults, egg exposures were primarily conducted in June during their peak oviposition periods. Only exposures of P. rufipes egg masses were conducted at the end of the summer due to its unusual phenology with oviposition starting in August. Over the period of three years (2019–2021) 498, 397 and 259 H. halys egg masses and 390, 762 and 468 non-target egg masses were exposed in Switzerland, respectively (Table 2).
Collection of wild stink bug egg masses
In Italy, throughout the exposure period of sentinel egg masses, weekly surveys were conducted at the same site to collect naturally laid egg masses of H. halys and non-target pentatomids and scutellerids. Egg masses were collected by visual inspection on the host plants present in the site (maple, hazelnut, cherry, elder, and ash) as described in Moraglio et al. (2021a).
In Switzerland, wild egg masses of H. halys, N. viridula, and P. prasina were collected primarily from catalpa trees in the cities of Basel and Zurich based on visual inspection of the foliage following the exposure of sentinel egg masses. In addition, H. halys egg masses were also collected from catalpa trees in the city of Bern (2020/21) and Biel (2021) after increasing infestations had been reported from both cities. Egg masses of P. rufipes were collected from maple trees in the urban areas of Bern in early autumn 2021 after the first T. japonicus had been detected in May of the same year.
All field-collected egg masses were transferred to the laboratory and reared for parasitoid adults and bug nymphs. In Italy, egg masses from which parasitoids had already partially emerged were included in the analysis, whereas in Switzerland, only complete egg masses (no emergence prior to collections) were included.
Laboratory rearing of sentinel and wild egg masses
Both sentinel and field-collected egg masses were placed individually in plastic Petri dishes (∅ 60 mm) and reared at 24±1 °C, 65±5% RH and 16L:8D until all bug nymphs or parasitoid adults had emerged. Egg masses were checked every 2 days, and emergence of nymphs and parasitoids was recorded. Parasitoids were sexed, sorted by family (Scelionidae, Eupelmidae, Encyrtidae and Pteromalidae) and stored in 99% ethanol. Parasitoids were identified to species level using the keys by Peng et al. (2020) for genus Anastatus Motschulsky, Samra et al. (2018) and Triapitsyn et al. (2020) for genus Ooencyrtus Ashmead, Johnson (1984) and Kozlov and Kononova (1983) for genus Telenomus Haliday, Talamas et al. (2017) and Tortorici et al. (2019) for genus Trissolcus Ashmead and Sabbatini Peverieri et al. (2019) for genus Acroclisoides Girault and Dodd (hyperparasitoids). All identified specimens were deposited in the collection of the Dipartimento di Scienze Agrarie, Forestali e Alimentari (DISAFA), University of Torino, Italy. At the end of each season, all egg masses were inspected under a stereo microscope at up to 40x magnification to assess their fate. Following Moraglio et al. (2021a), the following egg fate categories were assigned to individual eggs within each egg mass: (1) hatched—a bug nymph had emerged; (2) parasitized—a parasitoid had emerged; (3) preyed—the egg was empty or partly destroyed due to the attack of a sucking or chewing predator; and (4) unemerged—undamaged, but nothing had emerged (ESM 2, 3).
Non-reproductive mortality of N. viridula induced by T. japonicus
As laboratory tests have shown that T. japonicus frequently attacks and kills eggs of N. viridula but cannot complete development (Haye et al. 2020), unemerged eggs (undamaged, but no nymphs or parasitoids emerged) from N. viridula egg masses were stored in ethanol for molecular analysis. Molecular forensic analysis was applied to unemerged eggs from the following collections: (1) a subsample of 2019 N. viridula sentinel egg masses (n = 40 out of 67 exposed) from Italy; (2) all 2019 N. viridula wild egg masses collected in Italy (n = 12); and (3) all 2019 N. viridula wild egg masses collected in Switzerland (n = 4). Since all other non-target species tested in Haye et al. (2020) were suitable for parasitoid development, non-reproductive mortality was only analysed in N. viridula.
Genomic DNA was extracted and amplified from individual unemerged eggs using Scelionidae-specific primers, following the protocols described by Gariepy et al. (2019). All conditions for amplification of scelionid DNA were the same, except a single round of PCR was performed using 5 μL of template DNA, instead of the original protocol described and implemented by Gariepy et al. (2014, 2019) in which two rounds of PCR (each using 1 μL of template DNA or PCR product, respectively) were performed. Additionally, as Anastatus bifasciatus (Geoffroy) (Hymenoptera: Eupelmidae) was reared from sentinel and field-collected N. viridula egg masses, a separate PCR was performed on these same samples using Anastatus-specific PCR primers, following the protocol described by Stahl et al. (2019a).
All PCR products were visualized with a QIAxcel Advanced automated capillary electrophoresis system (Qiagen, Hilden, Germany) using the QX DNA Fast Analysis Kit. QIAxcel screengel software (version 1.2.0) was used to score the results, and only samples with the expected fragment size (~562-bp for Scelionidae and ~270-bp for A. bifasciatus) and >0.1 relative fluorescent units signal strength were considered positive. All unemerged sentinel and field-collected eggs that were positive for Scelionidae DNA were purified with ExoSAP-IT (Affymetrix, Santa Clara, CA, USA) following the manufacturer’s protocol. PCR products were bidirectionally Sanger-sequenced using an ABI 3730 DNA analyser (Robarts Research Institute, London Regional Genomics Centre, Ontario, Canada) with the same primers used in the PCR reactions. Forward and reverse sequences were assembled and sequences were edited using CodonCode Aligner, version 9.0.1 (CodonCode Corporation, Centreville, MA, USA). The Anastatus primers were only used diagnostically to detect the presence of parasitoids within this genus. However, as a precaution, all field-collected specimens which yielded Anastatus-positive PCR fragments were purified and sequenced (as described above) to confirm their identity.
All specimen and sequence data were uploaded to the Barcode of Life Data System (BOLD) v4.0 (http://www.boldsystems.org/) under the public project “Egg Parasitoids Associated with Nezara viridula in Europe” (Project code: EPNEZ), and sequences were also deposited in GenBank (accession no. OQ320837-OQ321705). Barcode Index Numbers (BINs) (Ratnasingham and Hebert 2013) were assigned to the scelionid sequences, and the BIN registry was used for taxonomic identification. If species-level identification was not available in the BIN registry, the sequence was screened though the BOLD identification engine to obtain genus or family-level identification. As Anastatus-specific PCR products are only ~270-bp in length, the resulting DNA sequences are considered non-barcode compliant (<500-bp in length) and are not assigned a BIN in BOLD; identification of Anastatus-specific sequences was therefore accomplished using the BOLD Identification Engine.
For each collection [(1) 2019 sentinels collected in Italy; (2) 2019 wild egg masses collected in Italy; and (3) 2019 wild egg masses collected in Switzerland], the total and mean proportion of non-parasitized unemerged and parasitized unemerged eggs was calculated. Overall parasitoid species composition in unemerged eggs for each collection was calculated based on the number of individual eggs assigned to a given species or taxonomic unit (based on molecular analysis), divided by the total number of eggs that yielded positive PCR results for parasitoid DNA.
Statistics
Because the phenology and oviposition periods and thus also the exposure periods of univoltine non-target species only partially overlapped with the oviposition period of the bivoltine H. halys (May to September), the proportions of eggs parasitized by T. japonicus were only compared for those months in which the non-targets and targets were exposed simultaneously. For example, when sentinel eggs of P. rufipes were exposed in August and in September, these were compared to H. halys eggs exposed in the same two months instead of all H. halys eggs exposed (May to September). Proportions of sentinel H. halys and non-target eggs parasitized by T. japonicus were compared with a two-sample test for equality of proportions with continuity correction using the proportion test function of the stats package in R version 3.6.1 (R Core Team 2019) (ESM4). Proportions of sentinel eggs parasitized by T. japonicus when exposed on plants or tags in Manno in 2019 was compared with Chi-square tests in R.
Results
Parasitism of sentinel eggs by T. japonicus
In Tables 1 and 2, the parasitism data for each bug species have been pooled for each year and locality. Accordingly, parasitism data refer to the total number of eggs exposed at each site in each year (total parasitism). Figures 1, 2, 3, 4 show the comparison of proportions of sentinel H. halys and non-target eggs parasitized by T. japonicus when exposed simultaneously; in other words, only data from those months in which sentinel eggs of both target and non-target species were exposed were compared (ESM 4).

Parasitism of sentinel eggs of H. halys and non-target species in Grugliasco (I) that were attacked at same level as H. halys in laboratory tests (Haye et al. 2020; Sabbatini Peverieri et al. 2021) (two-sample test for equality of proportions; ns = not significant, * = P < 0.05; ** = P <0.001; *** = P <0.0001; for details see ESM4)

Parasitism of sentinel eggs of H. halys and non-target species in Grugliasco (I) that were less attacked than H. halys in laboratory tests (grouped on the left), attacked but not suitable for development (N. viridula) (centre) and four additional species that were not part of previous laboratory testing (grouped on the right) (Haye et al. 2020; Sabbatini Peverieri et al. 2021); (two-sample test for equality of proportions; ns = not significant, * = P< 0.05; ** = P <0.001; *** = P <0.0001; for details see ESM4)

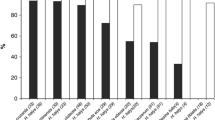
Parasitism of sentinel eggs of H. halys and P. rufipes by T. japonicus at three locations in Switzerland (O = apple orchard; N = natural site) (two-sample test for equality of proportions; ns = not significant, * = P < 0.05; ** = P <0.001; *** = P <0.0001; for details see ESM4)

Parasitism of sentinel eggs of H. halys, P. prasina and C. fuscispinus by T. japonicus at three locations in Switzerland (O = apple orchard; N = natural site) (two-sample test for equality of proportions; ns = not significant, * = P < 0.05; ** = P <0.001; *** = P <0.0001; for details see ESM4)
Italy: In total 963 and 4,289 parasitoids were reared from sentinel eggs of H. halys and non-target species, respectively (EMS 2). Trissolcus japonicus parasitized sentinel egg masses exposed on P. avium, M. alba and Acer spp. between the second week of May and the second week of September in both years. In 2019, in addition to H. halys, six out of 14 non-target species were parasitized by T. japonicus (Table 1; Figs 1, 2). Overall parasitism (all exposures throughout the season combined) of H. halys and P. prasina eggs by T. japonicus was 8.3% (n = 3,374 eggs) and 12.9% (n = 1,911), respectively. When both species were exposed simultaneously, parasitism of P. prasina (13.3%) was significantly higher (Fig. 1, EMS4). Parasitism of P. rufipes and H. halys was low (<10%), but not significantly different (Fig. 1, EMS4). The highest level of T. japonicus parasitism was found in C. mediterraneus (23.3%). Parasitism of other non-target species by T. japonicus was minimal, ranging from 0 to 7.7% (Fig. 2, EMS4).
In 2020, the number of non-target species parasitized by T. japonicus increased to 13 out of 15 (Table 1). Compared to 2019, parasitism of H. halys eggs by T. japonicus increased from 10.1% to 45.7%. At the same time, total parasitism of P. prasina (14.7%) and C. mediterraneus (28.1%) eggs remained stable or increased just slightly. Parasitism of D. baccarum, G. lineatum and R. nebulosa increased from 0% in 2019 to 13.9%, 12.8% and 10.9% in 2020, respectively (Table 1). When sentinel eggs of A. custos, P. prasina, R. nebulosa—three of the species that were attacked at the same level as H. halys in no-choice tests (Haye et al. 2020)—were exposed simultaneously with H. halys, parasitism by T. japonicus was significantly higher for H. halys eggs (Fig. 1). However, as in 2019, parasitism of H. halys and P. rufipes was not significantly different.
Switzerland: In total 1,449 and 3,324 parasitoids were reared from sentinel eggs of H. halys and non-target species, respectively. Over the course of three years, T. japonicus was recorded from H. halys and three non-target species, including P. prasina, P. rufipes and C. fuscispinus (Table 2; Figs. 3, 4). In Zurich, overall parasitism of sentinel H. halys eggs by T. japonicus was 11.0% in 2019, but decreased to 3.2% and 4.0% in the two following years (Table 2). Parasitism of H. halys in Basel was 5.8% and 6.5% in 2019 and 2020, respectively, but no T. japonicus were detected in 2021. Similarly, in the apple orchard in Manno, where T. japonicus was originally discovered, parasitism of sentinel eggs on tags was 2.9% in 2019, but no parasitism was detected in the two following years. However, in nearby natural habitats in Manno, T. japonicus was still present in 2020 and 2021, causing 2.5% and 6.1% parasitism, respectively. When eggs were exposed simultaneously on tags and plants in Manno (Table 2), significantly higher parasitism by T. japonicus on tags was observed for H. halys (χ2 = 34.35, df=1, P < 0.0001) and P. prasina (χ2 = 21.58, df=1, P < 0.0001). A significantly higher level of parasitism by T. japonicus on plants was only observed for C. fuscispinus (χ2 = 35.86, df=1, P < 0.0001). Eggs of D. baccarum were not parasitized by T. japonicus, independent of the exposure method.
Pentatoma rufipes was the non-target species most frequently parasitized by T. japonicus. Particularly, in 2019, overall parasitism by T. japonicus in Zurich and Basel was 17.5% and 21.8%, respectively (Table 2). When eggs of both species were exposed in parallel at both sites in August 2019, parasitism of H. halys and P. rufipes by T. japonicus was not significantly different (Fig. 3). In the two following years, parasitism by T. japonicus dropped significantly at both sites and only reached 3.3% and 6.3% in 2021 (Fig. 3). Remarkably, in Basel T. japonicus was only reared at low levels from P. rufipes sentinel eggs exposed in August 2020 and 2021, but not from H. halys. When exposed simultaneously in 2020 at Manno, parasitism of P. rufipes eggs by T. japonicus (16.7%) was significantly higher than parasitism of H. halys eggs (4.9%) (Fig. 3), whereas in 2021, it was nearly the opposite situation (H. halys: 22.1%, P. rufipes: 2.0%). Parasitism of P. prasina was generally low at all sites (<5%) (Table 2), and only in 2020, parasitism levels in Zurich reached more than 10%. In this year, T. japonicus parasitism of H. halys and P. prasina sentinel eggs exposed simultaneously in Zurich was not significantly different (Fig. 4). Carpocoris fuscispinus was only parasitized (13.6%) when exposed on plants at Manno in 2019 (Table 2, Fig. 4), whereas H. halys eggs exposed simultaneously were significantly less attacked (1.1%).
Parasitism of wild H. halys and non-target egg masses by T. japonicus
In total, 845 H. halys egg masses and 142 egg masses from eight non-target species were collected in Italy and Switzerland. Trissolcus japonicus was the most common parasitoid emerging from H. halys egg masses (65.1% to 100% of the emerged parasitoids) (Table 3). In Basel, parasitism in 2019 was 43.2% but declined to 22.3% and 13.3% in 2020 and 2021, respectively. In Zurich, a similar decline in parasitism was observed from 2020 (21.3%) to 2021 (13.9%). In Italy, parasitism of H. halys eggs in 2019 was 14.4% and increased to 17.3% in 2020. Since many parasitoids had already emerged in the field before the collections, the actual parasitism level is likely underestimated. Trissolcus japonicus was also reared from wild egg masses of P. prasina in Italy and Switzerland. In Italy, parasitism of P. prasina by T. japonicus was very low (<2%) and only three individuals emerged from a total of 13 egg masses in the laboratory. However, 85 parasitoids had already emerged in the field, and thus, parasitism was likely higher. In Switzerland, two out of 13 P. prasina egg masses were parasitized by T. japonicus (16.4% egg parasitism). In addition, T. japonicus was reared from two out of 12 wild egg masses of P. rufipes collected in Bern (Switzerland) in 2021 (16.9% egg parasitism), whereas six were attacked by native parasitoids (43.4%). Other non-target species living on weeds, e.g. D. baccarum, Carpocoris sp. or P. lituratus, were exclusively parasitized by native European egg parasitoids (Table 3).
Non-reproductive mortality of N. viridula induced by T. japonicus
The majority of the N. viridula sentinel eggs (n = 2,056) exposed in Italy in 2019 (69%) were unemerged (no nymph or parasitoid emerged), with a mean proportion of 72.3% (±5.2SE) unemerged eggs per egg mass. A total of 22 of the 40 egg masses (55%) contained at least one unemerged egg that was parasitized, with a mean proportion of 24.2% (±5.6SE) unemerged eggs containing parasitoid DNA (including Scelionidae and Anastatus). Parasitoid species composition in unemerged eggs is shown in Table 4. Of the scelionids, T. japonicus dominated the species composition, and accounted for 60.7% of parasitized, unemerged eggs (n = 364), followed by Trissolcus cultratus (Mayr) (16.8%), and Trissolcus mitsukurii (Ashmead) (11.5%), another exotic egg parasitoid of H. halys present in Italy (Zapponi et al. 2021). Approximately, 2.2% of parasitized eggs were specimens which were positive for scelionid DNA, but yielded low-quality DNA sequences which precluded their identification. The remaining 8.8% of parasitized eggs yielded PCR products consistent with the genus Anastatus and are presumably A. bifasciatus based on the fact that this was the only Anastatus species reared from N. viridula (Table 5).
Unemerged eggs from all naturally laid, field-collected egg masses in Italy and Switzerland in 2019 were processed and sequenced for scelionid and Anastatus DNA. From the Italian samples, only five of the 12 egg masses were parasitized. Approximately, 22% of the 1002 individual eggs were unemerged, with a mean proportion of 23.9 (±4.9SE) unemerged eggs per egg mass. The mean proportion of unemerged eggs that were positive for parasitoid DNA was 28.8% (±6.3SE). The dominant parasitoid species was T. japonicus (63.8%), followed by Anastatus (32.0%) (Table 4). All samples that were positive for Anastatus DNA were sequenced (n = 30), and 68% yielded high-quality sequences consistent with A. bifasciatus. The remaining 32% failed to sequence; however, based on the fact that other eggs within the same egg mass produced high-quality A. bifasciatus sequences, it is highly likely that all Anastatus detected here were indeed A. bifasciatus. Trissolcus basalis (Wollaston) occupied 2.1% of the parasitoid species composition, and another 2.1% of the samples positive for scelionid DNA yielded low-quality sequence data, precluding their identification.
Of the 14 N. viridula egg masses collected in 2019 in Switzerland, 13 masses were parasitized. A total of 984 individual eggs were obtained, 508 were unemerged, with a mean proportion of 53.6 (±5.2SE) unemerged eggs per egg mass. Approximately, 74% of unemerged eggs were positive for parasitoid DNA (mean 68.0%±4.8SE), and the species composition is shown in Table 4. The dominant parasitoid species detected was T. japonicus (75.3%). Trissolcus basalis, T. cultratus, and T. turesis occupied 22.1%, 0.5%, and 0.3% of the parasitoid species composition (respectively), and an unidentified species of Telenomus (0.3%) was sporadically detected in the species composition. The remaining 1.5% of the species composition consisted of specimens which were positive for scelionid DNA but failed to produce identifiable DNA sequences. The unidentified Telenomus species (BIN ADW5015) produced a high-quality, barcode compliant DNA sequence, but no species-level match was available in BOLD and GenBank. This is likely because voucher specimens of this species have not yet been sequenced, and as such, no barcode matches are available in public DNA databases.
Parasitism by other parasitoids
In Italy, overall parasitism of sentinel eggs by native egg parasitoids in 2019 was below 10% in most species, and only in P. prasina, it reached 24.3% (Table 1). In parallel to the increase in parasitism of H. halys by T. japonicus in 2020, however, parasitism by native species also increased for nine non-target species. Remarkably, parasitism of H. halys did not follow this trend and actually decreased, whereas parasitism of N. viridula remained stable at low level.
In Switzerland, parasitism of H. halys by native parasitoids (mainly A. bifasciatus) did not exceed 4.0% in all years (Table 2). In contrast, parasitism of native stink bugs such as P. prasina and P. rufipes was much higher, reaching maximum 30.0% and 18.2%, respectively. Parasitism of C. fuscispinus (one of the few other native stink bugs that was attacked by T. japonicus in Switzerland) by native parasitoids, reached up to 31.5% (when exposed on plants).
Associations of parasitoids other than T. japonicus and their host obtained by sentinel egg exposure is shown in Table 5. Apart from T. japonicus, eight native Scelionidae, three Eupelmidae and one Pteromalidae (hyperparasitoid) were reared from sentinel egg masses. Trissolcus mitsukurii was reared from the two native hosts P. rufipes (15.3%) and C. mediterraneus (32.6%) in 2019, whereas in 2020, it was not detected (Tables 1, 3). In addition, T. mitsukurii was also reared from a single egg mass of P. rufipes in the Canton Ticino (Table 5), representing the first record of this species in Switzerland.
In general, each Trissolcus species was associated with 1 to 9 host species, whereas the two Telenomus species were reared from 9 to15 hosts, respectively. The highest number of hosts for a given parasitoid species (20) was recorded for A. bifasciatus. Two additional Anastatus species that have not previously been reported in Europe were sporadically reared from sentinel non-targets; however, further collections would be necessary to clarify trends in their host ranges. Anastatus gansuensis Chen & Zang was reared in low numbers from H. halys in Italy and Switzerland, and also P. prasina in Italy, and C. fuscispinus and G. lineatum in Switzerland, whereas Anastatus japonicus Ashmead emerged from H. halys, P. prasina and P. strictus in Switzerland.
Predation of sentinel eggs
In both Italy and Switzerland, predation of sentinel eggs exposed on tags was highly variable among species, sites and years (Tables 1, 2). In 2019, in Italy (all species combined), predation of eggs exposed on tags was ranging from 1.1% to 54.1% (mean 20.5±4.2% SE) (Table 1), whereas on plants, it was 0% to 76.3% (mean 25.7±9.7% SE). For nearly all species, overall predation was higher than overall parasitism. In five out of eight stink bug species that were exposed on tags and plants, predation was higher on plants. In 2020, predation levels ranged from 0% to 95.8% (mean 22.1±6.0% SE). In 2019, in nine out of 14 species, predation levels were higher than overall parasitism, whereas in 2020, it was only six out of 16 (Table 1).
In Switzerland, predation levels of eggs exposed on plant tags were similar to those observed in Italy (Table 2), averaging 23.9±5.0% (SE) and 22.0±2.8% (SE) in 2019 and 2020, respectively. In 2021, however, predation was much lower than in previous years, averaging 12.0±2.7%. Predation was the highest (28.8±4.1% SE), when eggs were exposed on plants in 2019.
Discussion
Unintentional introduction of natural enemies has increased in recent years due to massive increase in global international trade and tourism (Gippet et al. 2019). The recent establishment of such natural enemies provides a unique opportunity to validate estimates of fundamental (laboratory) host range with realized (ecological) post-arrival host range as it manifests over time, particularly for those species that, based on laboratory experiments, would have been considered non-specific and not approved for release [e.g. Trichomalus perfectus (Walker), Leptopilina japonica Novković & Kimura)] (Haye et al. 2015; Beers et al. 2022; Puppato et al. 2022; Weber et al. 2021). Among these species is also the samurai wasp, T. japonicus, which was first recorded in Europe in 2017 (Stahl et al. 2019b).
Studies of the fundamental host range of T. japonicus in Europe had shown that 13 out of 21 non-target species tested were suitable for successful development of T. japonicus offspring in no-choice tests (Haye et al. 2020; Sabbatini Peverieri et al. 2021). In the present study, 15 out of 18 non-target species exposed in Italy and Switzerland were successfully parasitized by T. japonicus in the field, confirming its high potential to develop on a wide variety of hosts as predicted by its fundamental host range. Among the 15 suitable hosts was also an additional species, E. venustissimus, that had not been part of previous host range studies. In addition, T. japonicus was reared from a single egg mass of N. viridula, although this host was not suitable for development in previous laboratory no-choice tests.
Remarkably, in Grugliasco, a much larger number of non-target hosts was parasitized, including many species that were not attacked in Switzerland, which likely resulted from different methodologies, climate conditions or H. halys densities. Whereas in Grugliasco, all egg masses, except those of E. maura and E. austriaca, were exposed on trees, where most likely natural H. halys populations were present, in Switzerland, egg masses of stink bugs usually feeding on weeds or bushes (e.g. D. baccarum, P. lituratus, P. strictus) were not exposed on trees but in their natural habitats, weedy patches or meadows, where H. halys usually does not occur. The absence of T. japonicus in the weedy habitats in Switzerland agrees with earlier studies that T. japonicus was more frequently found in woodland or lightly wooded urban habitats than for example ground crops like soybean (Herlihy et al. 2016; Quinn et al. 2019a, b, 2021). This arboreal habitat preference may explain the differences in the number of non-targets attacked in the two regions, and consequently, some non-target species parasitized in Grugliasco could be ‘false positives’ based solely on exposure locations that are not realistic in nature. Similarly, it has been suggested that ecological refuges from parasitism (e.g. habitats at higher altitude) have mitigated the population-level impacts of a closely related egg parasitoid, T. basalis on native Hawaiian stink bug species (Johnson et al. 2005). In addition, at Grugliasco, a remarkable increase in H. halys parasitism by T. japonicus from 10% in 2019 to 45% in 2020 was observed, and at the same time, non-target parasitism by T. japonicus increased significantly. The reason for this drastic increase remains unknown, but the number of wild H. halys eggs collected at Grugliasco in 2019 and 2020 was 6,580 and 10,677, respectively, indicating that more host eggs were available in 2020, which may have supported an increase in T. japonicus populations and subsequently parasitism of H. halys and non-target species.
Although the realized host range comprised 15 non-target species, the number of hosts attacked alone does not necessarily suggest an increased risk of ecological damage. As suggested by van Driesche and Hoddle (2017), parasitism of native species at levels that do not significantly lower the long-term density of a non-target species (<10%) should be considered as ‘host use’, while rates of more than 50%, if widespread and sustained over time, may have negative population-level impacts for non-target species. Milnes and Beers (2019) showed that in a field paired-host assay in North America, the rate of parasitism on three native species was significantly less than that on H. halys eggs and concluded that the potential impact of T. japonicus on these pentatomids is probably minimal. Similarly, the large majority of non-target species exposed in the present study was clearly less parasitized by T. japonicus than H. halys. In both countries, the forest bug, P. rufipes, was the most parasitized non-target species with higher levels of parasitism particularly observed in Basel (18%) and Zurich (22%) in 2019, and in Manno (17%) and Grugliasco (77%) in 2020. The reasons for this are likely based on its phenology and the overlap of its feeding niche (trees) with H. halys. Whereas most European pentatomids overwinter as adults and lay their eggs in May/June, P. rufipes overwinters as second instar nymph and lays its eggs in August/September (Powell 2020). Field studies in China (Zhang et al. 2017) and the USA (Quinn et al. 2021) have shown that parasitism of sentinel H. halys eggs by T. japonicus is continuously increasing from May to August due to its rapid development and its ability to produce several generations per year. The same seasonal increase of parasitism of H. halys from 19% in May to 83% in September 2020 (ESM 5) was observed at Grugliasco in 2020, and accordingly, by the time P. rufipes started laying eggs (August), T. japonicus populations had likely built up and likely reached their maximum. Consequently, sentinel P. rufipes eggs were likely more frequently encountered by T. japonicus than any of the univoltine non-target species that lay their eggs in spring when T. japonicus populations are still fairly low. It should also be considered that the Grugliasco site did not represent the natural habitat for P. rufipes; in fact, no wild egg masses were ever found in the two years. In Bern, where T. japonicus was first detected in spring 2021, parasitism by this exotic parasitoid of wild P. rufipes egg masses collected in autumn at the same site was 16.9%, whereas parasitism by the native parasitoid T. cultratus, which can act as hyperparasitoid of T. japonicus (Konopka et al. 2017), was 35.5%. To what extent the native T. cultratus, which is the dominant parasitoid of P. rufipes, may prevent intense T. japonicus parasitism remains unknown, but ongoing life table studies in Europe may help to understand if the arrival of T. japonicus could have potential negative impacts at population level.
Compared to P. rufipes, the green stink bug P. prasina, which was among the four species that were parasitized at the same level as H. halys in laboratory tests (Haye et al. 2020; Sabbatini Peverieri et al. 2021), was attacked at relatively low levels (sentinel eggs: <11% in Switzerland and <15% in Italy). Parasitism of the few wild P. prasina egg masses found at Basel and Zurich combined was higher though (3 out of 10), but the sample size was very low. In contrast to P. rufipes, the oviposition period of P. prasina started in late April, when T. japonicus populations had not built up yet. Accordingly, a negative impact of T. japonicus at population level is not expected. Maximum parasitism of R. nebulosa, which starts laying eggs even before P. prasina in early April, was 11% in Grugliasco in 2020, but it should be noted that the amount of sentinel R. nebulosa egg masses available for this study was very limited. The predatory species A. custos was only exposed in July and August 2020 at Grugliasco and the higher level of parasitism by T. japonicus (30%) was likely due to the late exposure, given the timing with higher populations of this parasitoid in the field.
Previous laboratory host range tests had indicated that the exotic common green stink bug, N. viridula, is not suitable for parasitoid development, although regularly attacked by T. japonicus (Haye et al. 2020; Sabbatini Peverieri et al. 2021). Instead, oviposition in the unsuitable host induced a significant level of non-reproductive mortality. In the present study, the analysis of unemerged eggs from field-collected N. viridula egg masses with molecular tools showed that a high proportion of eggs contained T. japonicus DNA, confirming that non-reproductive mortality induced by egg parasitoids can indeed be an important mortality factor of invasive stink bugs (Abram et al. 2019; Kaser et al. 2018). As N. viridula is an invasive pest in Europe, any impact resulting in mortality of this pest would be considered beneficial.
Botch and Delfosse (2018) demonstrated that T. japonicus females reared on non-target hosts showed reduced host specificity and suggested that future efforts should be made to investigate the ability of T. japonicus to reproduce long term on non-target species when H. halys eggs are limited. Such a situation is currently observed in northern Switzerland, where H. halys populations have consistently declined since 2019 (Ackermann et al. 2022). However, concerns that such a decrease in H. halys could result in a host shift and substantial parasitism of alternative non-target hosts such as P. rufipes have not been validated during the course of the present study. In fact, in Switzerland the opposite trend was observed and with declining H. halys populations in 2020 and 2021, T. japonicus parasitism of sentinel P. rufipes eggs also declined.
Since it is impossible to collect large (meaningful) numbers of wild (field-laid) egg masses of native European non-target species, sentinel egg masses were used in the present study instead (Milnes et al. 2019; Quinn et al. 2019b). This can underestimate parasitism, parasitoid abundance and species richness as shown for H. halys by Jones et al. (2014). For example, in 2021 in Basel, T. japonicus was only reared in very low numbers from sentinel P. rufipes and P. prasina eggs but not from H. halys. However, parasitism of wild H. halys egg masses collected in the same area and year was 13%. One reason for the observed difference could be that ‘oviposition sites’ chosen for the exposure of sentinel eggs may not have matched with those that would have been chosen naturally by the stink bugs. Since sentinel egg masses were exposed on plastic tags, lower parasitism may have resulted from a lack of important chemical cues used for host location by T. japonicus (Arif et al. 2021; Malek et al. 2021). On the contrary, if these cues were absolutely essential, one would not have expected high levels of parasitism of sentinel eggs as observed in Grugliasco in 2020 or China (Zhang et al. 2017). However, at some sites (Grugliasco, Basel, Zurich), wild H. halys populations were already present on the trees selected for the exposure of sentinel eggs, likely compensating for the lack of chemical cues.
Peterson et al. (2022) demonstrated that feeding and oviposition by H. halys induced direct and systemic changes in volatile compound emissions from peach trees and tree of heaven and speculated that such changes in plant volatile compounds could be used by natural enemies such as T. japonicus. In the first year of the study (2019), we had exposed egg masses laid on host plants in the laboratory, but we did not find increased parasitism on plants compared to sentinel eggs exposed on plastic tags. However, egg predation on potted plants was extremely high, and we cannot exclude that at least some parasitized egg masses had been eaten by predators, thereby masking the potential effect of plant volatiles on parasitism.
The potential impact of T. japonicus on H. halys or specific non-target species cannot be summarized by interactions between species pairs because these species are entangled in diverse communities and are exposed to a multitude of ecological factors (Sentis et al. 2022). The large majority of European stink bug species lay their eggs in spring when T. japonicus populations are low, and can therefore gain partial temporal refuge from parasitism. In addition, species living on herbaceous hosts in the ground cover can escape intensive parasitism due to the preference of T. japonicus for woody habitats, thereby providing a spatial refuge for those species occupying such habitats. On the other hand, species sharing the same ecological niche as H. halys (trees) are more likely to be attacked by T. japonicus, but a potential increased risk should only be considered for those species with an unusual life cycle, such as P. rufipes or Picromerus bidens (L.) (Hemiptera: Pentatomidae), that lay their eggs in late summer when the majority of parasitism by T. japonicus takes place. Incorporating ecological factors such as habitat specificity, host and parasitoid phenology, host density and competition with native parasitoids or predators, can help to yield a more realistic scenario of potential risks for non-target species, but such field studies will only be possible where the biological control agents have become established—either through intentional releases or accidental establishments.
Author contribution
TH and LT conceived and designed research. LT, TH, CM, FT and STM conducted field experiments and analysed data. LT, STM and TH collected and reared non-target species for establishing laboratory colonies. TG did the molecular analysis of stink bug eggs. FT identified all egg parasitoids. TH, STM, TG and LT wrote the manuscript. All authors read and approved the manuscript
Data availability
All data generated or analysed during this study are included in this published article and its supplementary information files.
References
Abram PK, Hoelmer KA, Acebes-Doria A et al (2017) Indigenous arthropod natural enemies of the invasive brown marmorated stink bug in North America and Europe. J Pest Sci 90:1009–1020. https://doi.org/10.1007/s10340-017-0891-7
Abram PK, Brodeur J, Urbaneja A, Tena A (2019) Non-reproductive effects of insect parasitoids on their hosts. Annu Rev Entomol 64:259–276. https://doi.org/10.1146/annurev-ento-011118-111753
Ackermann A, Nölly M, Szalatnay D, Hollenstein R (2022) Merkblatt Baumwanzensituation (2/2022); https://redaktion.strickhof.ch/server/api/dokument/GetDokument?id=5541. Accessed 2 January 2023
Arif MA, Alınç T, Guarino S, Colazza S, Cusumano A, Peri E (2021) Only females oviposit: chemical discrimination of adult stink bug sex by the egg parasitoid Trissolcus japonicus. Front Ecol Evol 9:652561. https://doi.org/10.3389/fevo.2021.652561
Arnold K (2009) Halyomorpha halys (Stål, 1855), eine für die europäische Fauna neu nachgewiesene Wanzenart (Insecta: Heteroptera: Pentatomidae: Cappaeini). Mitt Thüringer Entomol 16:19
Avila GA, Charles JG (2018) Modelling the potential geographic distribution of Trissolcus japonicus: a biological control agent of the brown marmorated stink bug, Halyomorpha halys. BioControl 63:505–518. https://doi.org/10.1007/s10526-018-9866-8
Barratt BIP, Evans AA, Ferguson CM, Barker GM, McNeill MR, Phillips CB (1997) Laboratory nontarget host range of the introduced parasitoids Microctonus aethiopoides and Microctonus hyperodae (Hymenoptera: Braconidae) compared with field parasitism in New Zealand. Environ Entomol 26:694–702. https://doi.org/10.1093/ee/26.3.694
Barratt BIP, Howarth FG, Withers TM, Kean JM, Ridley GS (2010) Progress in risk assessment for classical biological control. Biol Control 52:245–254. https://doi.org/10.1016/j.biocontrol.2009.02.012
Beers EH (2019) Trissolcus japonicus (Hymenoptera: Scelionidae) causes low levels of parasitism in three North American Pentatomids under field conditions. J Insect Sci 19:1–6. https://doi.org/10.1093/jisesa/iez074
Beers EH, Beal D, Smytheman P, Abram PK, Schmidt-Jeffris R, Moretti E, Daane KM, Looney C, Lue C-H, Buffington M (2022) First records of adventive populations of the parasitoids Ganaspis brasiliensis and Leptopilina japonica in the United States. J Hymenopt Res 91:11–25. https://doi.org/10.3897/jhr.91.82812Puppato2022
Belousova EN (2007) Revision of the shield-bug genera Holcostethus Fieber and Peribalus Mulsant et Rey (Heteroptera, Pentatomidae) of the Palaearctic region. Entomol Rev 87:701–739. https://doi.org/10.1134/S0013873807060085
Bosco L, Moraglio ST, Tavella L (2018) Halyomorpha halys, a serious threat for hazelnut in newly invaded areas. J Pest Sci 91:661–670. https://doi.org/10.1007/s10340-017-0937-x
Botch PS, Delfosse ES (2018) Host-acceptance behavior of Trissolcus japonicus (Hymenoptera: Scelionidae) reared on the invasive Halyomorpha halys (Heteroptera: Pentatomidae) and non-target species. Environ Entomol 47:403–411. https://doi.org/10.1093/ee/nvy014
Cameron PJ, Walker GP (1997) Host specificity of Cotesia rubecula and Cotesia plutellae, parasitoids of white butterfy and diamondback moth. P NZ Plan P 50:236–241. https://doi.org/10.30843/nzpp.1997.50.11293
Cameron PJ, Hill RL, Teulon DAJ, Stukens MAW, Connolly PG, Walker GP (2013) A retrospective evaluation of the host range of four Aphidius species introduced to New Zealand for the biological control of pest aphids. Biol Control 67:275–283. https://doi.org/10.1016/j.biocontrol.2013.08.011
Cesari M, Maistrello L, Piemontese L et al (2018) Genetic diversity of the brown marmorated stink bug Halyomorpha halys in the invaded territories of Europe and its patterns of diffusion in Italy. Biol Invasions 20:1073–1092. https://doi.org/10.1007/s10530-017-1611-1
Claerebout S, Haye T, Ólafsson E, Pannier E, Bultot J (2018) Première occurrence de Halyomorpha halys (Stål, 1855) (Hemiptera: Heteroptera: Pentatomidae) pour la Belgique et actualisation de sa distribution en Europe. Bull Soc r Belg Entomol 154:205–227
Derjanschi V, Péricart J (2005) Hémiptères Pentatomoidea Euro-Méditerranéens. Volume 1. Généralités, Systématique: Première Partie. Faune de France 90:1–496
Dieckhoff C, Wenz S, Renninger M, Reißig A, Rauleder H, Zebitz CPW, Reetz J, Zimmermann O (2021) Add Germany to the list - adventive population of Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae) emerges in Germany. Insects 12:414. https://doi.org/10.3390/insects12050414
van Driesche RV, Hoddle M (2017) A review of nontarget effects of insect biological control agents: concepts and examples. united states department of agriculture, forest service, forest health technology enterprise team, morgantown, WV. http://bugwoodcloud.org/resource/pdf/nontarget.pdf (accessed 3 January 2023)
Froud KJ, Stevens, PS (2003) Importation biological control of Heliothrips haemorrhoidalis by Thripobius semiluteus in New Zealand - a case study of non-target host and environmental risk assessment. In: Van Driesche RG (ed) Proceedings of the 1st International Symposium on Biological Control of Arthropods, Honolulu, Hawaii, 14–18 January 2002, United States Department of Agriculture, Forest Service, Morgantown, WV, FHTET-2033-05, pp 366–369
Gariepy TD, Haye T, Zhang J (2014) A molecular diagnostic tool for the preliminary assessment of host-parasitoid associations in biological control programs for a new invasive pest. Mol Ecol 23:3912–3924. https://doi.org/10.1111/mec.12515
Gariepy TD, Bruin A, Haye T, Milonas P, Vetek G (2015) Occurrence and genetic diversity of new populations of Halyomorpha halys in Europe. J Pest Sci 88:451–460. https://doi.org/10.1007/s10340-015-0672-0
Gariepy TD, Bruin A, Konopka J, Scott-Dupree C, Fraser H, Bon MC, Talamas E (2019) A modified DNA barcode approach to define trophic interaction between native and exotic pentatomids and their parasitoids. Mol Ecol 28(2):456–470. https://doi.org/10.1111/mec.14868
Gippet JM, Liebhold AM, Fenn-Moltu G, Bertelsmeier C (2019) Human-mediated dispersal in insects. Curr Opin Insect Sci 35:96–102. https://doi.org/10.1016/j.cois.2019.07.005
Haye T, Goulet H, Mason PG, Kuhlmann U (2005) Does fundamental host range match ecological host range? - A retrospective case study of a Lygus plant bug parasitoid. Biol Control 35:55–67. https://doi.org/10.1016/j.biocontrol.2005.06.008
Haye T, Abdallah S, Gariepy T, Wyniger D (2014) Phenology, life table analysis, and temperature requirements of the invasive brown marmorated stink bug, Halyomorpha halys, in Europe. J Pest Sci 87:407–418. https://doi.org/10.1007/s10340-014-0560-z
Haye T, Mason PG, Gillespie DR, Gibson GAP, Diaconu A, Brauner AM, Miall JH, Kuhlmann U (2015) Determining the host specificity of the biological control agent Trichomalus perfectus (Hymenoptera: Pteromalidae): the importance of ecological host range. Biocontrol Sci Techn 25:21–47. https://doi.org/10.1080/09583157.2014.945900
Haye T, Moraglio T, Stahl J, Visentin S, Gregorio T, Tavella L (2020) Fundamental host range of Trissolcus japonicus in Europe. J Pest Sci 93:171–182. https://doi.org/10.1007/s10340-019-01127-3
Hedstrom C, Lowenstein D, Andrews H, Bai B, Wiman N (2017) Pentatomid host suitability and the discovery of introduced populations of Trissolcus japonicus in Oregon. J Pest Sci 90:1169–1179. https://doi.org/10.1007/s10340-017-0892-6
Herlihy MV, Talamas EJ, Weber DC (2016) Attack and success of native and exotic parasitoids on eggs of Halyomorpha halys in three Maryland habitats. PLoS ONE 11(3):e0150275. https://doi.org/10.1371/journal.pone.0150275
Hoebeke ER, Carter ME (2003) Halyomorpha halys (Stǻl) (Heteroptera: Pentatomidae): a polyphagous plant pest from Asia newly detected in North America. Proc Entomol Soc Wash 105:225–237
Hopper KR (2001) Research needs concerning non-target impacts of biological control introductions. In: Wajnberg E, Scott JK, Quimby PC (eds) Evaluating Indirect Ecological Effects of Biological Control. CABI Publishing, Wallingford, pp 39–56
Johnson NF (1984) Systematics of Nearctic Telenomus: classification and revisions of the podisi and phymatae species groups (Hymenoptera: Scelionidae). B Ohio Biol Surv 6:1–113. https://doi.org/10.5281/ZENODO.23887
Johnson MT, Follett PA, Taylor AD et al (2005) Impacts of biological control and invasive species on a non-target native Hawaiian insect. Oecologia 142:529–540. https://doi.org/10.1007/s00442-004-1754-5
Jones AL, Jennings DE, Hooks CRR, Shrewsbury PM (2014) Sentinel eggs underestimate rates of parasitism of the exotic brown marmorated stink bug, Halyomorpha halys. Biol Control 78:61–66. https://doi.org/10.1016/j.biocontrol.2014.07.011
Kaser JM, Nielsen AL, Abram P (2018) Biological control effects of non-reproductive host mortality caused by insect parasitoids. Ecol Appl 28:1081–1092. https://doi.org/10.1002/eap.1712
Konopka JK, Haye T, Gariepy T, Mason P, Gillespie D, McNeil J (2017) An exotic parasitoid provides an invasional lifeline for native parasitoids. Ecol Evol 7:277–284. https://doi.org/10.1002/ece3.2577
Kozlov MA, Kononova SV (1983) [Telenominae of the Fauna of the USSR.]. Nauka, Leningrad
Kriticos D, Kean J, Phillips C, Senay S, Acosta H, Haye T (2017) The potential global distribution of the brown marmorated stink bug, Halyomorpha halys Stål (Hemiptera: Pentatomidae): A critical threat to plant biosecurity. J Pest Sci 90:1033–1043. https://doi.org/10.1007/s10340-017-0869-5
Kuhlmann U, Mason PG (2003) Selection of non-target species for host specificity tests. In: Bigler F, Babendreier D, Kuhlmann U (eds) Biological Control of Arthropods Using Invertebrates: Methods for Environmental Risk Assessment. CABI, Wallingford, pp 15–37
Lee DH, Short BD, Joseph SV, Bergh JC, Leskey TC (2013) Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ Entomol 42:627–641. https://doi.org/10.1603/EN13006
Leskey TC, Nielsen AL (2018) Impact of the invasive brown marmorated stink bug in North America and Europe: history, biology, ecology, and management. Annu Rev Entomol 63:599–618. https://doi.org/10.1146/annurev-ento-020117-043226
Lupoli R, Dusoulier F, Cruaud A, Cros-Arteil S, Streito JC (2013) Morphological, biogeographical and molecular evidence of Carpocoris mediterraneus as a valid species (Hemiptera: Pentatomidae). Zootaxa 3609:392–410. https://doi.org/10.11646/zootaxa.3609.4.2
Maistrello L, Vaccari G, Caruso S et al (2017) Monitoring of the invasive Halyomorpha halys, a new key pest of fruit orchards in northern Italy. J Pest Sci 90:1231–1244. https://doi.org/10.1007/s10340-017-0896-2
Malek R, Kaser JM, Anfora G, Ciolli M, Khrimian A, Weber DC, Hoelmer KA (2021) Trissolcus japonicus foraging behavior: implications for host preference and classical biological control. Biol Control 16:104700. https://doi.org/10.1016/j.biocontrol.2021.104700
Miller TD (1992) Growth stages of wheat. Better crops with plant food. Potash & Phosphate Institute 76: 12. http://agrilife.org/yoakumterryipm/files/2013/03/Wheat-Growth.pdfMilnes
Moraglio ST, Tortorici F, Pansa MG, Castelli G, Pontini M, Scovero S, Visentin S, Tavella L (2020) A 3-year survey on parasitism of Halyomorpha halys by egg parasitoids in northern Italy. J Pest Sci 93:183–194. https://doi.org/10.1007/s10340-019-01136-2
Moraglio ST, Tortorici F, Giromini D, Pansa MG, Visentin S, Tavella L (2021) Field collection of egg parasitoids of Pentatomidae and Scutelleridae in Northwest Italy and their efficacy in parasitizing Halyomorpha halys under laboratory conditions. Entomol Exp Appl 69:52–63. https://doi.org/10.1111/eea.12966
Moraglio ST, Tortorici F, Visentin S, Pansa MG, Tavella L (2021) Trissolcus kozlovi in North Italy: host specificity and augmentative releases against Halyomorpha halys in hazelnut orchards. Insects 12(1–14):464. https://doi.org/10.3390/insects12050464
Morehead SA, Feener DH Jr (2000) An experimental test of potential host range in the ant parasitoid Apocephalus paraponerae. Ecol Entomol 25:332–340. https://doi.org/10.1046/j.1365-2311.2000.00261.x
Neimorovets VV (2020) Review of the genus Eurygaster (Hemiptera: Heteroptera: Scutelleridae) of Russia. Zootaxa 4722:501–539. https://doi.org/10.11646/zootaxa.4722.6.1
Onstad DW, McManus ML (1996) Risks of host range expansion by parasites of insects: population ecology can help us estimate the risks that biological control agents pose to nontarget species. Bioscience 46:430–435. https://doi.org/10.2307/1312877
Peng L, Gibson G, Tang LU, Xiang J (2020) Review of the species of Anastatus (Hymenoptera: Eupelmidae) known from China, with description of two new species with brachypterous females. Zootaxa 4767:3. https://doi.org/10.11646/zootaxa.4767.3.1
Péricart J (2010) Hémiptères Pentatomoidea Euro-Méditerranéens. Volume 3: Podopinae et Asopinae. Fédération Française des Sociétés de Sciences Naturelles, Paris
Peterson HM, Ray S, Ali JG et al (2022) Feeding and oviposition by the brown marmorated stink bug, Halyomorpha halys (Stål) induce direct and systemic changes in volatile compound emissions from potted peach and tree of heaven. Arthropod-Plant Inte 16:227–247. https://doi.org/10.1007/s11829-022-09893-1
Powell G (2020) The biology and control of an emerging shield bug pest, Pentatoma rufipes (L.) (Hemiptera: Pentatomidae). Agr Forest Entomol 22:298–308. https://doi.org/10.1111/afe.12408
Puppato S, Grassi A, Pedrazzoli F, De Cristofaro A, Ioriatti C (2020) First report of Leptopilina japonica in Europe. Insects 11:611. https://doi.org/10.3390/insects11090611
Quinn NF, Talamas EJ, Acebes-Doria AL, Leskey TC, Christopher Bergh J (2019) a) Vertical sampling in tree canopies for Halyomorpha halys (Hemiptera: Pentatomidae) life stages and its egg parasitoid, Trissolcus japonicus (Hymenoptera: Scelionidae). Environ Entomol 48:173–180. https://doi.org/10.1093/ee/nvy180
Quinn NF, Talamas EJ, Leskey TC, Bergh JC (2019b) Sampling methods for adventive Trissolcus japonicus (Hymenoptera: Scelionidae) in a wild tree host of Halyomorpha halys (Hemiptera: Pentatomidae). J Econ Entomol 112:1997–2000. https://doi.org/10.1093/jee/toz107
Quinn NF, Talamas EJ, Leskey TC, Bergh JC (2021) Seasonal captures of Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae) and the effects of habitat type and tree species on detection frequency. Insects 12:118. https://doi.org/10.3390/insects12020118
R Core Team (2019) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Ratnasingham S, Hebert PD (2013) A DNA-based registry for all animal species: the barcode index number (BIN) system. PloS One 8:e 66213. https://doi.org/10.1371/journal.pone.0066213
Ribes J, Pagola-Carte S (2013) Hémiptères Pentatomoidea Euro-Méditerranéens. Volume 2: Systématique: Deuxième Partie. Faune de France 96:1–394
Sabbatini Peverieri G, Talamas E, Bon MC, Marianelli L, Bernardinelli I, Malossini G, Benvenuto L, Roversi PF, Hoelmer K (2018) Two Asian egg parasitoids of Halyomorpha halys (Stål) (Hemiptera, Pentatomidae) emerge in northern Italy: Trissolcus mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J Hymenopt Res 67:37–53. https://doi.org/10.3897/jhr.67.30883
Sabbatini Peverieri G, Mitroiu M-D, Bon M-C, Balusu R, Benvenuto L, Bernardinelli I, Fadamiro H, Falagiarda M, Fusu L, Grove E, Haye T, Hoelmer K, Lemke E, Malossini G, Marianelli L, Moore MR, Pozzebon A, Roversi PF, Scaccini D, Shrewsbury P, Tillman G, Tirello P, Waterworth R, Talamas EJ (2019) Surveys of stink bug egg parasitism in Asia, Europe and North America, morphological taxonomy, and molecular analysis reveal the Holarctic distribution of Acroclisoides sinicus (Huang & Liao) (Hymenoptera, Pteromalidae). J Hymenopt Res 74:123–151. https://doi.org/10.3897/jhr.74.46701
Sabbatini Peverieri G, Boncompagni L, Mazza G, Paoli F, Dapporto L, Giovannini L, Marianelli L, Hoelmer K, Roversi PF (2021) Combining physiological host range, behavior and host characteristics for predictive risk analysis of Trissolcus japonicus. J Pest Sci 94:1003–1016. https://doi.org/10.1007/s10340-020-01311-w
Samra S, Cascone P, Noyes J, Ghanim M, Protasov A, Guerrieri E et al (2018) Diversity of Ooencyrtus spp (Hymenoptera: Encyrtidae) parasitizing the eggs of Stenozygum coloratum (Klug) (Hemiptera: Pentatomidae) with description of two new species. PLoS ONE 13(11):e0205245. https://doi.org/10.1371/journal.pone.0205245
Sentis A, Hemptinne JL, Margo A, Outreman Y (2022) Biological control needs evolutionary perspectives of ecological interactions. Evol Appl 15:1537–1554. https://doi.org/10.1111/eva.13457
Stahl JM, Gariepy TD, Beukeboom LW, Haye T (2019) A molecular tool to identify Anastatus parasitoids of the brown marmorated stink bug. Entom Exp Appl 167:692–700
Stahl J, Tortorici F, Pontini M, Bon MC, Hoelmer K, Marazzi C, Tavella L, Haye T (2019) First discovery of adventive populations of Trissolcus japonicus in Europe. J Pest Sci 92:371–379. https://doi.org/10.1007/s10340-018-1061-2
Stoeckli S, Felber R, Haye T (2020) Current and future potential distribution and voltinism of the brown marmorated stink bug, Halyomorpha halys, in Switzerland. Int J Biometeorol 64:2019–2032. https://doi.org/10.1007/s00484-020-01992-z
Talamas EJ, Buffington ML, Hoelmer K (2017) Revision of Palearctic Trissolcus Ashmead (Hymenoptera, Scelionidae). J Hymenopt Res 56:3–185
Tortorici F, Talamas EJ, Moraglio ST, Pansa MG, Asadi-Farfar M, Tavella L, Caleca V (2019) A morphological, biological and molecular approach reveals four cryptic species of Trissolcus Ashmead (Hymenoptera, Scelionidae), egg parasitoids of Pentatomidae (Hemiptera). In: Talamas E (Eds) Advances in the Systematics of Platygastroidea II. J Hymenopt Res 73:153–200. https://doi.org/10.3897/jhr.73.39052.
Triapitsyn SV, Andreason SA, Power N, Ganjisaffar F, Fusu L, Dominguez C, Perring TM (2020) Two new species of Ooencyrtus (Hymenoptera, Encyrtidae), egg parasitoids of the bagrada bug Bagrada hilaris (Hemiptera, Pentatomidae), with taxonomic notes on Ooencyrtus telenomicida. J Hymenopt Res 76:57–98. https://doi.org/10.3897/jhr.76.48004
van Lenteren J, Babendreier D, Bigler F, Burgio G, Hokkanen HMT, Kuske S, Loomans AJM, Menzler-Hokkanen I, van Rijn PCJ, Thomas MB, Tommasini MG, Zeng QQ (2003) Environmental risk assessment of exotic natural enemies used in inundative biological control. BioControl 48:3–38. https://doi.org/10.1023/A:1021262931608
Weber D, Hajek A, Hoelmer K, Schaffner U, Mason P, Stouthamer R, Talamas E, Buffington M, Hoddle M, Haye T (2021) Unintentional biological control. In: Mason PG (ed) Biological Control - Global Impacts. CSIRO Publishing, Challenges and Future Directions of Pest Management, pp 110–140
Wermelinger B, Wyniger D, Forster B (2008) First records of an invasive bug in Europe: Halyomorpha halys Stal (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Mitt Schweiz Entomol Ges 81:1–8. https://doi.org/10.5169/seals-402954
Wong WHL, Walz MA, Oscienny AB, Sherwood JL, Abram PK (2021) An effective cold storage method for stockpiling Halyomorpha halys (Hemiptera: Pentatomidae) eggs for field surveys and laboratory rearing of Trissolcus japonicus (Hymenoptera: Scelionidae). J Econ Entomol 114:571–581. https://doi.org/10.1093/jee/toaa307
Wyniger D, Kment P (2010) Key for the separation of Halyomorpha halys (Stål) from similar-appearing pentatomids (Insecta: Heteroptera: Pentatomidae) occurring in Central Europe, with new Swiss records. Mitt Schweiz Entomol Ges 83:261–270. https://doi.org/10.5169/seals-403015
Yang ZQ, Yao YX, Qiu LF, Li ZX (2009) A new species of Trissolcus (Hymenoptera: Scelionidae) parasitizing eggs of Halyomorpha halys (Heteroptera: Pentatomidae) in China with comments on its biology. Ann Entomol Soc Am 102:39–47. https://doi.org/10.1603/008.102.0104
Zapponi L, Tortorici F, Anfora G, Bardella S, Bariselli M, Benvenuto L, Bernardinelli I, Butturini A, Caruso S, Colla R, Costi E, Culatti P, Di Bella E, Falagiarda M, Giovannini L, Haye T, Maistrello L, Malossini G, Marazzi C, Marianelli L, Michelon L, Moraglio ST, Pozzebon A, Preti M, Salvetti M, Scaccini D, Schmidt S, Szalatnay D, Roversi PF, Tavella L, Tommasini M, Vaccari G, Zandigiacomo P, Sabbatini Peverieri G (2021) Assessing the distribution of adventive egg parasitoids of Halyomorpha halys in Europe with a large-scale monitoring program. Insects 12(4):316. https://doi.org/10.3390/insects12040316
Zhang J, Zhang F, Gariepy T, Mason P, Gillespie D, Talamas E, Haye T (2017) Seasonal parasitism and host specificity of Trissolcus japonicus in northern China. J Pest Sci 90:1127–1141. https://doi.org/10.1007/s10340-017-0863-y
Acknowledgments
We would like to thank Christine Cock, Emily Grove, Emily Lemke, Lauréline Humair, Marion Risse, Sandrine Fattore, Sara Morello, Sara Scovero and Sara Visentin for technical assistance in the laboratory and for their help in bug collection. We would like to thank Lukas Seehausen for his help with the statistical analysis.
Funding
This work was supported by the Swiss Federal Office for the Environment (contract no. 00. 5005. PZ/S084-0191) and with core financial support from CABI’s member countries (see http://www.cabi.org/about-cabi/who-we-work-with/key-donors/), and by Hazelnut Company Division of the Ferrero Group.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
All authors declare that there is no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals (vertebrates) performed by any of the authors.
Consent to participate
Informed consent was obtained from all individual participants included in the study.
Consent to publish
Consent was obtained from all individual participants included in the study.
Additional information
Communicated by Kent Daane.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Haye, T., Moraglio, S.T., Tortorici, F. et al. Does the fundamental host range of Trissolcus japonicus match its realized host range in Europe?. J Pest Sci 97, 299–321 (2024). https://doi.org/10.1007/s10340-023-01638-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10340-023-01638-0