
Abstract
Spotted cucumber beetle (Diabrotica undecimpunctata howardi; also known as southern corn rootworm) is a pest feeding on many crops, including cucurbits, corn (maize), peanuts, beans, and other crops and wild plants in North America. On cucurbit crops, where it co-occurs with striped cucumber beetle (Acalymma vittatum), both are key pests that vector Erwinia tracheiphila, a destructive pathogen that causes bacterial wilt. Here we deployed synthetic vittatalactone, the male-produced aggregation pheromone of A. vittatum, as lures in the field, to measure attraction of the target and non-target insects. Both sexes of D. u. howardi were strongly attracted to the vittatalactone lures (5–7.5-fold compared to blank trap) throughout the growing season in Maryland and Virginia, USA. Moreover, D. u. howardi was more attracted to live A. vittatum males feeding on squash plants than to plants with females or without any beetles, confirming the genuine organismal cross-attraction between these two key pests. The kairomonal attraction to vittatalactone suggests that it functions as a “keystone semiochemical,” mediating the colonization of cucurbits by multiple herbivore species. Such cross attraction could allow for use of vittatalactone in management of multiple pest species, with potential reduction of pesticide use in cucurbit crops.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Key message
-
Vittatalactone, pheromone of striped cucumber beetle, is highly attractive to both sexes of spotted cucumber beetle, in eastern USA
-
Use of vittatalactone as a kairomone in host-finding by the spotted-cucumber beetle confirms the latter species’ evolutionary connection to cucurbit hosts
-
Attractiveness of vittatalactone to both sexes of both species of cucumber beetles offers the potential for advances in sustainable pest management of both species
Introduction
Spotted cucumber beetle (Diabrotica undecimpunctata howardi; also known as southern corn rootworm) is a polyphagous pest feeding on cucurbits, corn (maize), peanuts, beans, and many other horticultural and ornamental crops in North America. Although adults are generalist feeders, they prefer certain hosts, especially cucurbits (Cranshaw and Shetlar 2018). This preference may arise from the fact that D. u. howardi sequesters cucurbitacins, bitter defensive compounds present in cucurbits that elicit a compulsive feeding response in D. u. howardi. In addition to cucurbitacins being used by D. u. howardi as a defense against predators (Metcalf and Lampman 1989), male D. u. howardi provide them as a component of nuptial gifts to females (Tallamy et al. 2000), helping to protect their eggs which are laid underground. Diabrotica undecimpunctata howardi overwinters as adults and is highly mobile, colonizing northern latitudes over the course of the growing season. Its widespread abundance, mobility and polyphagy pose major challenges to pest management (Hoffmann and Zitter 1994; Diver and Hinman 2008). A better understanding of D. u. howardi host location behavior is therefore needed to effectively manage this pest.
Volatile cues have shown promise for managing a number of agricultural pests (e.g., Bruce et al. 2005; Weber et al. 2014; Morrison et al. 2016), and evidence suggests that volatile cues could be used to also manipulate D. u. howardi behavior for pest management. Diabrotica undecimpunctata howardi males are attracted to a female-produced sex pheromone, R-10-methyl-2-tridecanone (Guss et al. 1983), and there is evidence that females, at least at close range, base mating preferences on male-specific compounds (Brodt et al. 2006). Both sexes are also attracted to cucurbit floral compounds including 1,2,4-trimethoxybenzene and trans-cinnamaldeyde (Jackson et al. 2005). They share this attraction with striped cucumber beetle (Acalymma vittatum), a cucurbit specialist and key pest with which D. u. howardi often co-occurs.
Acalymma vittatum has a male-produced aggregation pheromone, vittatalactone (Smyth and Hoffmann 2003; Morris et al. 2005), that also attracts the cucurbit specialist squash bugs Anasa tristis and A. armigera (Brzozowski et al. 2022; Weber et al. 2022). In the course of determining the attractiveness of vittatalactone in various habitats and seasons to A. vittatum and to nontarget insects, we noted the attraction of D. u. howardi to vittatalactone synthetic lures. In this report, we document this strong cross-species kairomonal attraction both to synthetic lures and to live A. vittatum males on host cucurbit plants.
Materials and methods
Plants and insects
For all experiments, beetle collecting, and beetle feeding, we used potted summer squash (Cucurbita pepo) cv. ‘Yellow Crookneck’ (Johnny’s Selected Seeds, Fairfield, ME). We sowed seeds directly into 2.8 L plastic pots (3 seeds per pot) with peat/bark based growing medium (Pro-mix BK55, Premier Tech Horticulture, Rivière-du-Loup, Québec, Canada). Plants were kept in a greenhouse (25–30 °C, 16 L: 8 D) prior to experimental set-up.
We used A. vittatum collected in June 2022 from potted squash plants that we placed (exclusively for beetle collecting) at the borders of the previous season’s (2021) squash fields, and from a newly planted squash field at the USDA Beltsville Agricultural Research Center (BARC) North Farm (Beltsville, Maryland, USA; 39.0328° W, 76.9316° W). Following field collection, we identified the sex of each beetle and kept male and female beetles in a greenhouse under a natural photoperiod (25–30 °C) in separate cages provisioned with potted squash plants. We collected beetles no more than two days prior to the start of an experiment.
Field experiments testing attraction to synthetic vittatalactone
Synthesis of the vittatalactone mixture containing eight diastereomers with 2R, 3R configuration was performed by method of Chauhan and Paraselli (2017) by contract of USDA ARS with Chemveda Life Sciences, San Diego, CA. Gray rubber septa (1-F SS 1888 GRY, West Pharmaceutical Services, Lititz, PA) were washed with hexane in a Soxhlet apparatus, and dried in an exhaust hood overnight. The washed septa were then placed in a one-neck Norton flask, and a hexane solution with 1 mg of vittatalactone mixture and 0.6 ml hexane per septum added. The flask was rotated in a rotary evaporator (without vacuum) until the liquid was completely soaked into septa. Septa were then dried in an exhaust hood for 1 h before deployment in the field. This lure was shown to be highly attractive to A. vittatum in earlier field trials (Weber 2018).
Traps in all experiments were double-sided clear sticky traps (STKY Dual Panel Adhesive Traps, Trécé Inc., Adair, OK, USA), 15 × 30 cm, attached vertically with a large metal binder clip to wooden stakes (2 × 2 cm) at ~ 45 cm height (middle of trap above ground). Lures were affixed to the “lure” treatment traps with a small metal binder clip, hung from the large binder clip. Traps with and without lures were paired for each assay, with traps collected and rerandomized, and lures changed each week. Traps were deployed 5 May–1 December 2020 on BARC South Farm, Beltsville, Maryland (39.0175° N, 76.9455° W), and 19 May–1 October on Virginia Tech Kentland Farm, Whitethorn, Virginia (37.2006° N, 80.5657° W). At each site, four randomized complete blocks, consisting of one trap baited with 1 mg mixed vittatalactone, and one unbaited trap, were set up adjacent to former and current year’s cucurbit crops, at a separation of 10 m within blocks (pairs) and ≥ 10 m between blocks. Fields were not treated with any insecticide or fungicide applications. In the Maryland experiment, we identified captured D. u. howardi to sex and recorded the number of males and females captured each week. In the Virginia experiment, we recorded the total number of D. u. howardi captured each week.
Field experiments testing attraction to volatiles from live Acalymma vittatum
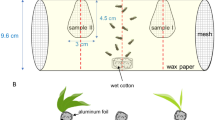
We conducted two experiments in 2022 to test attraction of D. u. howardi to olfactory cues produced by live striped cucumber beetles. All treatments included potted squash plants bagged with paint strainers (600 μm polyester mesh, with c. 18 L maximum volume, Trimaco, Inc., Morrisville, NC, USA) to prevent beetles from leaving plants and so that insect and plant-produced cues were exclusively olfactory. Each of these bait plants was placed inside of a cylindrical wire cage staked to the ground with two flanking wire frames. To catch attracted beetles, we attached two clear sticky traps horizontally using metal binder clips, one to each of the frames (Figure S1).
The first experiment took place from 10 to 21 June and included four treatments: (1) four males on plant, (2) four females on plant, (3) plant without beetles, and (4) blank pot with growing medium. Because our early-season beetle collection from the field was male-biased, this experiment was set up as a completely randomized design with seven replicates of the male treatment, five replicates of the female treatment, and four replicates each of the uninfested plant and blank pot treatments. These plants were randomized along the borders of two fields on BARC North Farm (10 plants per field). One field (39.0333° N, 76.9344° W) had been planted in the 2021 season with yellow crookneck squash, and the other (39.0284° N, 76.9333° W) with zucchini (C. pepo). Potted plants were spaced 10 m apart within each field. All potted plants were late in their vegetative stage upon experimental set-up, and most had begun flowering at the conclusion of the experiment. To evaluate attractiveness of each treatment, we counted the number of D. u. howardi, as well as A. vittatum, the squash bug Anasa tristis, and the horned squash bug A. armigera on each sticky trap.
The second experiment took place from 28 June to 15 July along the border of the previous season’s zucchini field (the latter field above). Because we speculated that floral volatiles might play a role in D. u. howardi attraction, this experiment included five treatments: (1) four males on vegetative plant, (2) four females on vegetative plant, (3) vegetative plant without beetles, (4) flowering plant without beetles, (5) blank pot with growing medium. The vegetative plants were in the cotyledon stage at the start of the experiment, and had true leaves but had not yet started producing flower buds at the experiment’s completion. Each flowering plant had 1–3 flowers at a time throughout the experiment. This experiment was set up as eight spatiotemporal blocks: four spatial blocks from 28 June-6 July, and another four spatial blocks from 6 to 15 July. For the second temporal block, we rerandomized plants in each spatial block and replaced the sticky traps. We counted the number of D. u. howardi, A. vittatum, A. tristis, and A. armigera captured on each sticky trap.
Data analysis
Attraction to synthetic vittatalactone
Diabrotica undecimpunctata howardi activity exhibits a nonuniform pattern throughout the season; therefore, we evaluated attraction to the vittatalactone lure versus the unbaited lure based on the season-long sum of D. u. howardi captured. We first used log-linear models in the Catmod procedure in SAS 9.4 (SAS Institute 2016) with sex, lure treatment, and block as variables for the Maryland study and lure treatment and block as variables for the Virginia study. All two- and three-way interactions with block were nonsignificant. Therefore, we proceeded with binomial exact tests to determine the statistical significance and magnitude (with 95% confidence interval) of the treatment effect (Lowry 2022; StatPages 2009). In addition, we used Fisher 2 × 2 Exact tests (Lowry 2022) to test for interaction of treatment*sex for Maryland captures, and for interaction of treatment*location for total Virginia versus Maryland captures.
Attraction to volatiles from live Acalymma vittatum
To determine whether the number of captured D. u. howardi and A. vittatum differed by bait plant treatment, we conducted separate generalized linear mixed models for each species using SAS Proc Glimmix with treatment as the fixed effect (four and five treatments total for the completely randomized and blocked experiments, respectively). For the completely randomized experiment, field was included as a random effect, and for the blocked experiment, spatiotemporal block was a random effect. Because we had count data, both models assumed a negative binomial distribution and used a log link function. When models indicated a significant overall treatment effect, we followed up with Tukey–Kramer tests at a Bonferroni-adjusted alpha level to determine which pairwise treatments were significantly different at an experimentwise α = 0.05. For the low numbers of Anasa spp. also captured, we employed the Kolmogorov–Smirnov one-sample test (Lowry 2022) to test total captures.
Results
Attraction to synthetic vittatalactone
In Maryland, we captured a total of 450 male and 338 female D. u. howardi across all traps. The sex*lure interaction was nonsignificant (Fisher Exact test P = 0.77), indicating that attraction to the vittatalactone versus unbaited lure was independent of sex. There were over 5 times as many of both male D. u. howardi (P < 0.0001) and female D. u. howardi (P < 0.0001) at traps baited with vittatalactone relative to unbaited traps (Fig. 1). In Virginia, we captured a total of 120 D. u. howardi (both sexes combined) across all traps. There were over 7.5 times as many D. u. howardi on vittatalactone-baited traps as there were on unbaited traps (P < 0.0001; Fig. 1). The site*lure interaction was nonsignificant (Fisher Exact test P = 0.34), indicating that D. u. howardi response to the lure did not differ between Maryland and Virginia locations.

Captures of Diabrotica undecimpunctata howardi on clear sticky traps baited with 1 mg lure of mixed vittatalactone, versus captures on unbaited clear sticky traps, during entire growing season 2020, in four randomized complete blocks at both Beltsville, Maryland, and Whitethorn, Virginia, USA. Error bars and ratio confidence intervals are 95% exact binomial CIs; comparisons between sexes (Maryland) and between sites are both non-significant by Fisher 2 × 2 Exact tests
Attraction to volatiles from live Acalymma vittatum
The generalized linear model for the completely randomized experiment indicated a significant treatment effect on D. u. howardi attraction (F3,15 = 6.06, P = 0.007). The number of D. u. howardi captured on the male treatment was significantly greater than the number captured at either the female or blank treatment, with over six times as many captured in the male treatment relative to the female treatment and over seven times as many captured in the male treatment relative to the blank treatment (Fig. 2a). Over four times as many D. u. howardi were captured in the male treatment relative to the plant treatment, although this pairwise difference was not statistically significant (Fig. 2a).

Least squares mean captures of Diabrotica undecimpunctata howardi at caged plants with either male or female Acalymma vittatum, or no beetles, or a blank pot. Error bars show 95% confidence limits; different letters above bars indicate pairwise significant differences based on Bonferroni-corrected Tukey–Kramer tests, P < 0.05. a: 10–21 June 2020, in completely randomized design. b: 28 June through 15 July 2020, in four randomized complete blocks
Diabrotica undecimpunctata howardi attraction in the blocked experiment also differed by treatment (F4,28 = 8.66, P < 0.0001). The number of D. u. howardi attracted to the male treatment was over three times greater than the number attracted to the female treatment, over four times greater than the number attracted to the flowering plant treatment, and over twice as many as the number attracted to the vegetative plant and blank treatments each (Fig. 2b). Acalymma vittatum attraction exhibited a strong treatment effect in both experiments (completely randomized: F3,15 = 13.48, P = 0.0002; blocked: F4,28 = 16.09, P < 0.0001), with attraction to the male treatment significantly greater than attraction to all other treatments (Figure S2). In the completely randomized experiment, we also captured only seven total A. tristis, and zero A. armigera. In the blocked experiment we captured ten A. tristis and seven A. armigera. In each case, in which Anasa were captured, numbers for the male treatment were significantly higher than all other treatments (P < 0.01, Kolmogorov–Smirnov one-sample test); see Figure S3.
Discussion
Our results demonstrate that the eastern subspecies of spotted cucumber beetle (D. u. howardi) is attracted to vittatalactone, the male-produced aggregation pheromone of the striped cucumber beetle (A. vittatum). At all experimental sites, traps baited with vittatalactone caught more D. u. howardi than unbaited traps. In addition, squash plants with live A. vittatum males attracted more D. u. howardi than plants with A. vittatum females or with no beetles, confirming that D. u. howardi is in fact attracted to the male-produced pheromone and not any different isomer present in the synthetic lures. Furthermore, the stronger attraction of A. vittatum to squash plants with A. vittatum males relative to other bait plant treatments is consistent with previous work (Smyth and Hoffmann 2003) and confirms that the male A. vittatum on our bait plants produced a detectable pheromone signal. Although we captured only a few Anasa tristis and A. armigera individuals, captures were significantly higher on plants with male A. vittatum, providing further support to prior work demonstrating cross attraction of both species to vittatalactone (Brzozowski et al. 2022; Weber et al. 2022). Here, we show vittatalactone attracts yet another cucurbit pest, indicating the potential for use of vittatalactone as a sustainable management strategy to manipulate the behavior of multiple pests simultaneously.
The attraction of D. u. howardi to vittatalactone seems to reflect the continued special role of plant hosts in the Cucurbitaceae in the life history of this very generalist feeder. Genetic analyses estimate that the genera Acalymma and Diabrotica diverged 45 (± 8, 95%CI) million years ago, and that the polyphagous fucata group of Diabrotica, of which D. u. howardi is a member, diverged from the oligophagous virgifera group 17 (± 4) Mya (Eben and Espinosa de los Monteros 2013). Although a D. u. howardi male-produced pheromone has not been identified, the continued affinity of D. u. howardi and other polyphagous Diabrotica species for cucurbitacin consumption and sequestration (Eben 2022) parallels with our findings of kairomonal attraction to the pheromone of the cucurbit specialist Acalymma. The eavesdropping kairomonal cross-attraction provides, through the related specialist beetles, a mechanism “tethering” the polyphagous beetles to their ancestral plant hosts. Such eavesdropping is likely favored by natural selection given the value of cucurbitacins as defensive compounds against fungi, nematodes, and various other natural enemies to all life stages (e.g., Ferguson and Metcalf 1985; Tallamy et al. 1998; Eben 2022).
The very strong evolutionarily-conserved bond between the generalist Diabrotica and their ancestral cucurbit hosts offers an opportunity for behavioral pest management using a combined bait of the volatile vittatalactone as a distance attractant and cucurbitacins as arrestants and feeding stimulants. Baits containing cucurbitacins and small amounts of pesticides have been shown to be effective at controlling D. virgifera (Metcalf et al. 1987; Lance and Sutter 1990; Chandler 2003), but distance attractants such as vittatalactone have never been included in these baits. We note that we have not yet determined the vittatalactone loading necessary to ensure that baits are more attractive than A. vittatum aggregations in the field, and this is worthy of future study. In addition, diversity and host affiliations among the many Diabrotica species are still not well-defined (Derunkov et al. 2015; Eben 2022), and it remains to be determined whether there are other species with attraction to vittatalactone or other semiochemicals of Acalymma species. However, the discovery that two species of Anasa (squash bugs; Hemiptera: Coreidae) are also attracted to vittatalactone (Brzozowski et al. 2022; Weber et al. 2022), shows that cross-attraction to vittatalactone is not limited to the Chrysomelidae or even Coleoptera. This merits further study of vittatalactone’s attractiveness among generalist and specialist cucurbit pests.
To our knowledge, this is the first study to document attraction of a generalist herbivore (D. u. howardi) to the pheromone of a co-occurring specialist (A. vittatum). These results likely reflect the selective advantage to D. u. howardi of feeding on cucurbitacins. Our results suggest that vittatalactone has an inordinate influence on the pest complex of cucurbits in North America, constituting a “keystone semiochemical” with effects on at least four important herbivores. This kairomonal attraction, and the availability of synthetic vittatalactone, present a new opportunity for efficient management of multiple pests that minimizes both insecticide use and nontarget effects. We suggest that vittatalactone be further evaluated for potential attractiveness to other cucurbit pests, as well as for deployment in commercial cucurbit fields as part of sustainable pest management strategy.
Author contributions
DCW conceived the study and designed synthetic vittatalactone experiments, FG made vittatalactone lures, TPK and SMB conducted experiments in Virginia, KP conducted synthetic vittatalactone experiments in Maryland, AIH designed and conducted live A. vittatum experiments, AIH and DCW analyzed data and wrote the first draft of the manuscript, and all other authors read and approved the manuscript.
Data availability
The data supporting the results of this study will be made available in the Dryad repository.
References
Brodt JF, Tallamy DW, Ali J (2006) Female choice by scent recognition in the spotted cucumber beetle. Ethology 112:300–306. https://doi.org/10.1111/j.1439-0310.2006.01158.x
Bruce TJ, Birkett MA, Blande J, Hooper AM, Martin JL, Khambay B, Prosser I, Smart LE, Wadhams LJ (2005) Response of economically important aphids to components of Hemizygia petiolata essential oil. Pest Manag Sci 61:1115–1121. https://doi.org/10.1002/ps.1102
Brzozowski LJ, Weber DC, Wallingford AK, Mazourek M, Agrawal AA (2022) Trade-offs and synergies in management of two co-occurring specialist squash pests. J Pest Sci 95:327–338. https://doi.org/10.1007/s10340-021-01379-y
Chandler LD (2003) Corn rootworm areawide management program: United States department of agriculture-agricultural research service. Pest Manag Sci 59:605–608. https://doi.org/10.1002/ps.683
Chauhan KR, Paraselli BR (2017) Eight diastereomers of vittatalactone and methods of making, and methods of attracting Acalymma vittatum. 2017 Aug 1. U.S. patent 9,718,797.
Cranshaw W, Shetlar D (2018) Garden insects of North America: the ultimate guide to backyard bugs. Second ed. Princeton University Press.
Derunkov A, Prado LR, Tishechkin AK, Konstantinov AS (2015) New species of Diabrotica Chevrolat Coleoptera: Chrysomelidae Galerucinae and a key to Diabrotica and related genera results of a synopsis of North and Central American Diabrotica species. J Insect Biodivers 3(2):1–55. https://doi.org/10.12976/jib/2015.3.2
Diver S, Hinman T (2008) Cucumber beetles: organic and biorational integrated pest management. National center for appropriate technology. www.attra.ncat.org
Eben A (2022) Ecology and evolutionary history of Diabrotica beetles—overview and update. Insects 13:156. https://doi.org/10.3390/insects13020156
Eben A, de Los E, Monteros A (2013) Tempo and mode of evolutionary radiation in Diabroticina beetles (genera Acalymma, Cerotoma, and Diabrotica). ZooKeys 332:207–321. https://doi.org/10.3897/zookeys.332.5220
Ferguson JE, Metcalf RL (1985) Cucurbitacins: Plant derived defense compounds for Diabroticites (Coleoptera: Chrysomelidae). J Chem Ecol 11:311–318. https://doi.org/10.1007/BF01411417
Guss PL, Tumlinson JH, Sonnet PE, McLaughlin JR (1983) Identification of a female-produced sex pheromone from the southern corn rootworm, Diabrotica undecimpunctata howardi Barber. J Chem Ecol 9:1363–1375. https://doi.org/10.1007/BF00994805
Hoffmann MP, Zitter TA (1994). Cucumber beetles, corn rootworms, and bacterial wilt in cucurbits. Vegetable MD Online, Cornell Cooperative Extension. http://vegetablemdonline.ppath.cornell.edu/factsheets/Cucurbit_Beetles.htm
SAS Institute (2016) SAS® 9.4. SAS Institute Inc, Cary, NC.
Jackson DM, Sorensen KA, Sorenson CE, Story RN (2005) Monitoring cucumber beetles in sweetpotato and cucurbits with kairomone-baited traps. J Econ Entomol 98:159–170. https://doi.org/10.1093/jee/98.1.159
Lance DR, Sutter GR (1990) Field-cage and laboratory evaluations of semiochemical-based baits for managing western corn rootworm (Coleoptera: Chrysomelidae). J Econ Entomol 83:1085–1090. https://academic.oup.com/jee/article-pdf/83/3/1085/2581214/jee83-1085.pdf
Lowry R (2022) VassarStats: website for statistical computation. http://vassarstats.net
Metcalf RL, Ferguson JE, Lampman R, Andersen JF (1987) Dry cucurbitacin-containing baits for controlling Diabroticite beetles (Coleoptera: Chrysomelidae). J Econ Entomol 80:870–875. https://academic.oup.com/jee/article-pdf/80/4/870/19233206/jee80-0870.pdf
Metcalf RL, Lampman RL (1989) The chemical ecology of Diabroticites and Cucurbitaceae. Experientia 45:240–247. https://doi.org/10.1007/BF01951810
Morris BD, Smyth RR, Foster SP, Hoffmann MP, Roelofs WL, Franke S, Francke W (2005) Vittatalactone, a beta-lactone from the striped cucumber beetle, Acalymma vittatum. J Nat Prod 68:26–30. https://doi.org/10.1021/np049751v
Morrison WR, Lee DH, Short BD, Khrimian A, Leskey TC (2016) Establishing the behavioral basis for an attract-and-kill strategy to manage the invasive Halyomorpha halys in apple orchards. J Pest Sci 89:81–96. https://doi.org/10.1007/s10340-015-0679-6
Smyth RR, Hoffmann MP (2003) A male-produced aggregation pheromone facilitating Acalymma vittatum [F.] (Coleoptera: Chrysomelidae) early-season host plant colonization. J Insect Beh 16:347–359. https://doi.org/10.1023/A:1024824025210
StatPages (2009) Exact binomial and poisson confidence intervals. https://statpages.info/confint.html
Tallamy DW, Whittington DP, Defurio F, Fontaine DA, Gorski PM, Gothro TW (1998) Sequestered cucurbitacins and pathogenicity of Metarhizium anisopliae (Monilialeas Moniliaceae) on spotted cucumber beetle eggs and larvae (Coleoptera: Chrysomelidae). Environ Entomol 27:366–372. https://doi.org/10.1093/ee/27.2.366
Tallamy DW, Gorski PM, Burzon JK (2000) Fate of male-derived cucurbitacins in spotted cucumber beetle females. J Chem Ecol 26:413–427. https://doi.org/10.1023/A:1005461522609
Weber DC (2018) Field attraction of striped cucumber beetles to a synthetic vittatalactone mixture. J Econ Entomol 111:2988–2991. https://doi.org/10.1093/jee/toy283
Weber DC, Cabrera Walsh G, DiMeglio AS, Athanas MM, Leskey TC, Khrimian A (2014) Attractiveness of harlequin bug, Murgantia histrionica, aggregation pheromone: field response to isomers, ratios, and dose. J Chem Ecol 40:1251–1259. https://doi.org/10.1007/s10886-014-0519-9
Weber DC, Haber AI, Pasteur K, Boyle SM, Kuhar TP, Cornelius M (2022) Both the squash bug Anasa tristis and horned squash bug Anasa armigera are attracted to vittatalactone, the aggregation pheromone of striped cucumber beetle. Environ Entomol. https://doi.org/10.1093/ee/nvac079
Acknowledgements
We thank Ashot Khrimian (Research Chemist, USDA ARS IIBBL) for overseeing the vittatalactone synthesis. We are grateful to Alex Bier, Megan Herlihy, and Daniel Wilczek for collection of a portion of the Maryland field traps, and to George Meyers and the BARC-West Farm Crew for planting and cultivation of the experimental fields. We also thank Brian Currin and Lucas Raymond for collection and deployment of a portion of traps at the Virginia site.
Funding
This work was funded in part by Southern SARE research grant # LS20-337.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare they have no competing interests.
Ethical approval
This article does not contain any studies with vertebrates performed by the authors.
Additional information
Communicated by Cesar Rodriguez-Saona.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Haber, A.I., Pasteur, K., Guzman, F. et al. Spotted cucumber beetle (Diabrotica undecimpunctata howardi) is attracted to vittatalactone, the pheromone of striped cucumber beetle (Acalymma vittatum). J Pest Sci 96, 21–27 (2023). https://doi.org/10.1007/s10340-022-01586-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10340-022-01586-1