
Abstract
Little is known regarding the migration routes of Siberian songbird populations. Here we provide the first geolocator tracking data for an Arctic Warbler breeding in Central Siberia and compare its movements with eight long-distance ring recoveries of this species. In autumn, the tracked individual migrated eastward to a stopover site in eastern Siberia, before migrating southward through Taiwan to its non-breeding sites in the Philippines and Indonesia. During spring migration, the bird spent at least one month at stopover sites in Mongolia, before migrating to its breeding site in June. Ring recovery data confirmed the movement between the Central Siberian breeding grounds and stopover sites in northern Mongolia.
Zusammenfassung
Über die Zugwege sibirischer Singvogel-Populationen ist bisher nur sehr wenig bekannt.
Mit Hilfe von Hell-Dunkel-Geolokatoren konnten wir zum ersten Mal den Herbst- und Frühjahrszug eines in Zentralsibirien brütenden Wanderlaubsängers dokumentieren und das Zugverhalten mit acht Fernfunden beringter Individuen vergleichen. Im Spätsommer verließ der Vogel das Brutgebiet ostwärts in Richtung eines Rastplatzes in Ostsibirien, bevor er in südlicher Richtung über Taiwan weiterzog, um seine Überwinterungsquartiere in den Philippinen und Indonesien zu erreichen. Während des Frühjahrszuges verbrachte der Wanderlaubsänger mindestens einen Monat in der Mongolei, bevor er das Brutgebiet im Juni erreichte. Ein Fernfund eines beringten Individuums bestätigt die Verbindung zwischen dem zentralsibirischen Brutgebiet und Zwischenstopps in der Mongolei.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Understanding annual cycle events, migration routes and the distribution of important non-breeding sites are crucial for the conservation of migratory species (Kirby et al. 2008). The Arctic Warbler Phylloscopus borealis occupies a huge breeding range in northern Eurasia, but little is known on its distribution during migration and the non-breeding season (BirdLife International 2023). The situation is especially complicated since this taxon was recently split into three species: Arctic Warbler P. borealis, Kamchatka Leaf Warbler P. examinandus and Japanese Leaf Warbler P. xanthodryas. They differ in song but only very slightly in plumage (Alström et al. 2011), and are therefore hardly identifiable away from their breeding sites and based on field observations only. While they differ in their breeding distribution (BirdLife International 2023), the three species seem to co-occur during the non-breeding season, e.g., in Thailand (Round et al. 2015, 2016). To understand the spatiotemporal distribution of the three species, it is therefore necessary to 1) catch birds during the non-breeding season to take measurements and assess plumage details necessary for identification (Round et al. 2015), or 2) to analyze genetic samples (Round et al. 2016), or 3) to track birds from the breeding grounds with geolocators (Adams et al. 2022).
For the Arctic Warbler, such tracking data exist for two individuals belonging to the population on the eastern edge of its distribution in Alaska (Adams et al. 2022). These two birds departed westwards from their breeding grounds, with autumn stopovers in south-eastern Russia and China, and non-breeding sites during the boreal winter on the island of Palau and the Philippines. One individual migrated during spring through the Japanese islands and the Kamchatka peninsula (Adams et al. 2022). The aim of our study was to investigate the non-breeding distribution and migration routes of Arctic Warblers breeding in Siberia, for which no data are available so far. We assumed that birds from the core breeding grounds in Siberia might spend the winter west of their eastern conspecifics from Alaska, for example in Indonesia. We therefore applied light-level geolocators and collated ring recovery data.
Materials and methods
Geolocator tracking
We captured Arctic Warblers in their breeding territories around Mirnoye field station, Central Siberia (88.96931 °E, 62.29825° N). We targeted adult males as we expected the highest chances for recapturing such individuals. Birds were caught in June 2019 with mist-nets and playback of their song. We tagged 19 males and one female with light-level geolocators (Intigeo W30Z11-DIP, Migrate Technology, UK). The weight of the tag and the harness (~ 0.4 g) corresponded to 3.6–4.7% of the birds’ body weights (8.5 to 10.9 g).
We relocated one adult male 14 June 2020 and retrieved its geolocator 17 June 2020. This bird (ring number Moscow VF46994) was initially ringed as an adult in 2016, and its nests were found in all years from 2016 to 2019, with the first eggs laid between 14 and 25 June. Remarkably, this individual had lost its right tarsus between the 2017 and 2018 breeding seasons, but still continued to return. The overall return rate (1/20 = 5%) was lower compared to color-ringed adult males in the study area during previous years (21.4%; O. Bourski, unpublished data). However, only 10 of the 20 tagged individuals were known, previously ringed individuals. A similarly low return rate (4/42, 9.5%) was reported for geolocator-tagged Arctic Warblers in Alaska (Adams et al. 2022).
We analyzed the light data using the FlightR package (Rakhimberdiev et al. 2017) in R version 4.3.1 (R Development Core Team 2011) following the geolocation manual (Lisovski et al. 2020). We log-transformed the downloaded data and used a threshold of 0.8 to estimate twilight events. We used the find.stationary.location function to determine a stationary location (between 1 December 2019 and 1 March 2020, initial.coords = 110,10) that we could use for the calibration of the sun elevation angle. We limited the grid for the possible route to latitudes between − 10 and 65 and longitudes of 85 to 150. We defined that birds can migrate through areas up to 700 km away from the shore and can stay in areas up to 100 km away from land since long over-water flights with possible stays on very small islands seem possible in this species based on a previous study (Adams et al. 2022). We run the particle filter with nParticles = 1e6 and used a cutoff probability of 0.1 to determine stationary periods. We replaced the first and the last estimated positions (i.e., at the breeding grounds) with the coordinates of the known breeding location.
Ring recoveries
We requested data regarding long-distance recoveries of Arctic Warblers > 100 km away from their ringing location from the EURING databank, from the National Bird Banding Center of China, the Yamashina Institute for Ornithology and Biodiversity Center of Japan and the Bird Ringing Centre of Russia. We received information of eight recoveries. One individual was ringed in Mongolia in 2019 and recaptured at our study site in Russia in 2021, and one was ringed in China in 2008 and recaptured in South Korea in 2009. Six more birds were ringed in Japan between 1989 and 2008 and recaptured in Japan or China. Those six recoveries might belong to Arctic Warblers sensu stricto, but they could also belong to Kamchatka Leaf Warblers or Japanese Leaf Warblers as those three species were split only recently (Alström et al. 2011).
Results
Geolocator tracking
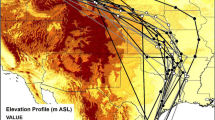
The tracked individual departed from the breeding grounds in Central Siberia during the first days of August 2019 (Table 1). Around 9 August, the bird arrived at a stopover site northeast of Lake Baikal (Fig. 1), where it stayed until 24 August. No positions were found during the following section of the autumn migration and the equinox period, but by 3 October the bird had arrived in Taiwan. It then migrated to the Philippines, where it stayed until November, before migrating further southward to western Sulawesi, where it stayed until December. Already in December, the bird began to move northward again, with stationary sites on Borneo, possibly on Palawan, and on Luzon, the Philippines. No positions were again estimated during the spring equinox, but by late April, the bird had already arrived to Mongolia. Here, at least four stationary sites were identified that were used until the end of May. By 4 June, the bird had arrived in the breeding area in Siberia.

© Wieland Heim
Estimated stationary positions and migration track of an Arctic Warbler based on light-level geolocation, as well as long-distance ring recoveries of this species. The diameter of the dots corresponds to the number of days spent at a given site, while the color of the dot indicates the main month of stay (see color legend). Uncertainties around the estimated positions (interquartile range) are plotted as grey vertical and horizontal lines. Note the ring recovery of one individual ringed in Mongolia and recaptured at the very same site in Russia where the geolocator-tracked bird has bred. All ring recoveries are numbered according to Table 2. Photo: Arctic Warbler with geolocator on its back, Mirnoye, Russia, June 2019
Ringing recoveries
We collected information on eight long-distance recoveries (Table 2). Five of the birds were recaptured within the same migration season (after 2–19 days), which allowed us to infer migration direction (all on a north–south or northeast–southwest axis through Japan) and migration speed for autumn (mean 46 km/day, range 30–70 km/day, n = 3) and spring migration (mean 278 km/day, range 202–354 km/day, n = 2).
Discussion
Here we present the first tracking data of an Arctic Warbler from its Eurasian core breeding area in Western Siberia, and a first compilation of available long-distance ring recoveries for this little-studied species.
The estimated departure of the tracked Arctic Warbler by the end of July or early August fits well to observation and ringing data of this color-marked individual, which has been seen no later than 22 July at the breeding grounds between 2016 and 2020 (O. Bourski, unpublished data). Arctic Warblers from Alaska were found to leave their breeding grounds around move one later (Adams et al. 2022). Such late departures are also observed in the Siberian breeding population and might be typical for successful and/or later breeders.
Our tracking data show a rarely documented large-scale eastward longitudinal movement from its breeding grounds toward its first main stopover area around Lake Baikal in autumn (> 1100 km). The recovery of an individual (V180564 BRCH) during breeding season at our Siberian study site, which was ringed during August at a stopover site in Mongolia, confirms this eastward autumnal movement (Table 2). Interestingly, the tracked Arctic Warblers from Alaska also started their autumn migration with a long longitudinal move, but westwards toward East Asia (Adams et al. 2022). The subsequent movement of the individual in our study through Taiwan and the Philippines was similar to one of the tracked birds from Alaska (Adams et al. 2022).
During the boreal winter, the tracked Arctic Warbler migrated to islands of the Philippines and Indonesia. Migration from the Asian mainland to islands in the Indo-Pacific has previously been reported for one other landbird species, the Blue-and-white Flycatcher Cyanoptila cyanomelana breeding in the Russian Far East (Heim et al. 2022). The non-breeding locations of the tracked Arctic Warbler breeding in Siberia were situated further west compared to the two individuals from Alaska (Adams et al. 2022). This suggests that different Arctic Warbler populations segregate spatially during the non-breeding season, with western breeding populations possibly wintering west of their eastern conspecifics. A similar pattern was observed in another species migrating along the East Asian flyway, the Siberian Rubythroat Calliope calliope (Heim et al. 2020).
Our model suggested that the Arctic Warbler already departed in early March from its non-breeding grounds. However, given that we have been unable to estimate positions during the equinox periods, the estimated arrival and departure dates might be biased. For example, we would expect a later spring departure, similar to other Palearctic species following the East Asian flyway (Heim et al. 2020). This also affects our estimates of the duration of migration and, consequently, migration speed, which was found to be much lower compared to other small landbirds in the East Asian flyway (Ktitorov et al. 2022; Sleptsov et al. 2023). The observed migration speeds based on the ring recoveries within one season suggested a higher speed during spring compared to autumn migration, a pattern which is common among migratory landbirds (Nilsson et al. 2013). However, our data here are limited to individuals that have been recaptured within Japan. Nevertheless, both the track and the recoveries point at high migration speeds for the last leg of spring migration.
Furthermore, our model suggested four different spring stop-over sites in Mongolia. While we consider it likely that Arctic Warblers might spend extended periods during spring south of the taiga belt waiting for suitable conditions at their destinations, we urge for a cautious interpretation of the actual estimated positions, given the failure of the model to calculate standard deviation of the position estimates for this period (Supplement 1). However, one of the estimated positions in Mongolia is very close to the Khurkh bird ringing station, where the individual (V180564 BRCH) was ringed which was later recaptured at our study site, confirming the location of stopover sites for Central Siberian populations in this area. Hundreds of Arctic Warblers are ringed annually at this site (Davaasuren et al. 2023).
Noteworthy is also the recovery of an individual (42,468 YIO) ringed mid-May in Japan and recaptured one year later mid-May in China. This could hint at flexible route choice between years or fast movements between the Japanese islands and the eastern Chinese mainland as documented for Amur Stonechats Saxicola stejnegeri breeding on Hokkaido (Yamaura et al. 2017).
Overall, our data suggest that the tracked Arctic Warbler was almost constantly on the move, not a single site during the annual cycle has been used for more than two months. This differs strongly from other East Asian songbird migrants, for which long autumn stopovers of up to three months and very long stationary periods during the boreal winter (five to six months) were documented (Heim et al. 2018, 2020, 2022). Most likely, Arctic Warblers are moving further to track seasonal peaks of food abundance—a similar pattern with several consecutive wintering sites was found in the closely related Willow Warbler Phylloscopus trochilus wintering in Africa (Lerche-Jørgensen et al. 2017; Sokolovskis et al. 2018). The rather high uncertainties around the estimated positions during the boreal winter (Fig. 1) might also relate to the constant movement of the tracked bird, but they could also stem from high levels of shading in the understory of tropical forests which would affect the accuracy of the position estimation (Lisovski et al. 2012).
Open questions remain regarding the molt schedule of the Arctic Warbler, which is believed to conduct both a post-breeding and pre-breeding molt in various extent (Snyder 2008; Norevik et al. 2020). Our tracked individual likely suspended its pre-breeding molt (it was recaptured while molting tertials 12–15 July 2019 at the breeding site) and continued molting at one or several of the non-breeding sites, using a similar strategy as described for Pallas´s Grasshopper-warblers Helopsaltes certhiola breeding in Siberia (Eilts et al. 2021). Another possibility is a pre-breeding molt during the long stay in Mongolia, which would explain why Arctic Warblers arrive with very fresh plumage at their Siberian breeding sites (O. Bourski, pers. observation).
Data availability
All tracking data are publicly available at Movebank (https://www.movebank.org/) in the study “Arctic Warbler (Phylloscopus borealis) Central Siberia” (Movebank study ID 3255314165).
References
Adams EM, Stenhouse IJ, Gilbert AT et al (2022) The first documentation of the nearctic-paleotropical migratory route of the Arctic Warbler. Ecol Evol 12:e9223
Alström P, Saitoh T, Williams D et al (2011) The Arctic Warbler Phylloscopus borealis–three anciently separated cryptic species revealed. Ibis 153:395–410
BirdLife International (2023) IUCN Red List for birds. Downloaded from http://www.birdlife.org on 03/10/2023
Davaasuren B, Erdenechimeg T, Dulmaa K (2023) Mongolia Bird Ringing Stations 2021–2022. Ulanbaataar
Eilts H, Feuerbach N, Round PD et al (2021) Complex postbreeding molt strategies in a songbird migrating along the East Asian Flyway, the Pallas’s Grasshopper Warbler Locustella certhiola. Ecol Evol 11:11–21
Heim W, Pedersen L, Heim R et al (2018) Full annual cycle tracking of a small songbird, the Siberian Rubythroat Calliope calliope, along the East Asian flyway. J Ornithol 159:893–899
Heim W, Heim RJ, Beermann I et al (2020) Using geolocator tracking data and ringing archives to validate citizen-science based seasonal predictions of bird distribution in a data-poor region. Glob Ecol Conserv 24:e01215
Heim W, Antonov A, Beermann I et al (2022) Light-level geolocation reveals unexpected migration route from Russia to the Philippines of a Blue-and-white-Flycatcher Cyanoptila cyanomelana. Ornithol Sci 21:121–126
Kirby JS, Stattersfield AJ, Butchart SHM et al (2008) Key conservation issues for migratory land- and waterbird species on the world’s major flyways. Bird Conserv Int 18:49–73. https://doi.org/10.1017/s0959270908000439
Ktitorov P, Heim W, Kulikova O, Gibson L (2022) Cross the sea where it is narrowest: migrations of Pacific Swifts (Apus pacificus) between Sakhalin (Russia) and Australia. J Ornithol. https://doi.org/10.1007/s10336-021-01913-w
Lerche-Jørgensen M, Willemoes M, Tøttrup AP et al (2017) No apparent gain from continuing migration for more than 3000 kilometres: willow warblers breeding in Denmark winter across the entire northern Savannah as revealed by geolocators. Mov Ecol 5:17. https://doi.org/10.1186/s40462-017-0109-x
Lisovski S, Hewson CM, Klaassen RHG et al (2012) Geolocation by light: accuracy and precision affected by environmental factors. Methods Ecol Evol 3:603–612. https://doi.org/10.1111/j.2041-210X.2012.00185.x
Lisovski S, Bauer S, Briedis M et al (2020) Light-level geolocator analyses: a user’s guide. J Anim Ecol 89:221–236
Nilsson C, Klaassen RHG, Alerstam T (2013) Differences in Speed and Duration of Bird Migration between Spring and Autumn. Am Nat 181:837–845. https://doi.org/10.1086/670335
Norevik G, Hellström M, Liu D, Petersson B (2020) Ageing & sexing of migratory East Asian passerines. Avium Förlag
R Development Core Team (2011) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing
Rakhimberdiev E, Saveliev A, Piersma T, Karagicheva J (2017) FLightR: an R package for reconstructing animal paths from solar geolocation loggers. Methods Ecol Evol 8:1482–1487
Round PD, Pierce AJ, Nualsri C, Sankamethawee W (2015) A record of Japanese Leaf Warbler Phylloscopus xanthodryas in Thailand. BirdingASIA 23:133–134
Round PD, Pierce AJ, Saitoh T, Shigeta Y (2016) Addition of Kamchatka Leaf Warbler Phylloscopus examinandus and Sakhalin Leaf Warbler P. borealoides to Thailand’s Avifauna. Bull Jpn Bird Banding Assoc 28:9–21
Sleptsov Y, Ktitorov P, Round PD, Heim W (2023) Autumn migration tracks of Helopsaltes grasshopper-warblers from Northeast Asia support recent taxonomic assignments. Ecol Evol 13:e9932
Snyder TA (2008) Molt strategies in Alaskan arctic warblers. North Am Bird Bander 33:22
Sokolovskis K, Bianco G, Willemoes M et al (2018) Ten grams and 13,000 km on the wing–route choice in willow warblers Phylloscopus trochilus yakutensis migrating from Far East Russia to East Africa. Mov Ecol 6:20. https://doi.org/10.1186/s40462-018-0138-0
Yamaura Y, Schmaljohann H, Lisovski S et al (2017) Tracking the Stejneger’s stonechat Saxicola stejnegeri along the East Asian-Australian Flyway from Japan via China to southeast Asia. J Avian Biol 48:197–202
Acknowledgements
We thank Alexandra Panaiotidi for support during field work. Further thanks go to Lixia Chen (National Bird Banding Center of China), Dorian Moss (EURING databank), Mariko Senda (Yamashina Institute for Ornithology) and Ilya Panov (Bird ringing center of Russia) as well as Olaf Geiter (Bird ringing center “Vogelwarte Helgoland”), Chenyang Liu, Yang Liu, Tianhao Zhao and Batmunkh Davaasuren for information on Arctic Warbler ring recoveries. Ringing data of birds with connection to Japan were used with the permission from the Yamashina Institute for Ornithology and Biodiversity Center of Japan (permission number #4-90). All experiments comply with the current laws of the countries where they were performed.
Funding
Open access funding provided by University of Zurich.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Communicated by N. Chernetsov.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Heim, W., Bourski, O., Shchemeleva, K. et al. From Siberia to Indonesia: tracking the migration of the Arctic Warbler Phylloscopus borealis. J Ornithol 165, 557–562 (2024). https://doi.org/10.1007/s10336-024-02146-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-024-02146-3