
Abstract
Nests are a critically important factor in determining the breeding success of many species of birds. Nevertheless, we have surprisingly little understanding of how the local environment helps determine the materials used in nest construction, how this differs among related species using similar nest sites, or if materials used directly or indirectly influence the numbers of offspring successfully reared. We also have little understanding of any potential links between nest construction and the assemblage of invertebrates which inhabit nests, in particular, ectoparasites. We addressed these questions by monitoring the success rates of Blue Tits Cyanistes caeruleus and Great Tits Parus major, using nest boxes in rural, urban greenspace and urban garden settings. We collected used nests, identified the arthropods present, and measured the proportions of highly processed anthropogenic materials used in nest construction. Twenty-five percent of Great Tit nest materials were of anthropogenic origin and this was consistent across habitats, while Blue Tits used little (1–2%) anthropogenic material except in gardens (~16%), suggesting that Great Tits preferentially sought out these materials. In fledged nests, an increasing use of anthropogenic material was associated with a lower general arthropod diversity and ectoparasite predator abundance (Blue Tits only), but higher levels of Siphonaptera (fleas). Higher arthropod diversity was associated with lower flea numbers, suggesting that increased diversity played a role in limiting flea numbers. No direct link was found between breeding success and either anthropogenic material usage or arthropod diversity and abundance. However, breeding success declined with increasing urbanisation in both species and increased with nest weight in Blue Tits. The interplay between urbanisation and bird ecology is complex; our work shows that subtle anthropogenic influences may have indirect and unexpected consequences for urban birds.
Zusammenfassung
Die Verwendung anthropogener Baumaterialien beeinflusst die Struktur der Arthropodengemeinschaften in Vogelnestern: Auswirkungen der Verstädterung und die Folgen für Ektoparasiten und Ausfliegeerfolg
Für den Bruterfolg vieler Vogelarten stellen Nester einen Faktor von kritischer Bedeutung dar. Dennoch haben wir nur überraschend wenige Kenntnisse darüber, wie das lokale Umfeld zur Prägung des verwendeten Baumaterials beiträgt, wie sich diesbezüglich verwandte Arten unterscheiden, welche ähnliche Nistplätze nutzen, oder ob die verwendeten Materialien direkt oder indirekt die Anzahl der erfolgreich aufgezogenen Nachkommen beeinflussen. Auch wissen wir nur wenig über potenzielle Zusammenhänge zwischen der Nestbauweise und den Invertebratengesellschaften, speziell Ektoparasiten, welche die Nester besiedeln. Diesen Fragen gingen wir nach, indem wir die Erfolgsraten in Nistkästen brütender Blaumeisen Cyanistes caeruleus und Kohlmeisen Parus major aus ländlichen Bereichen, städtischen Grünflächen und aus Stadtgärten kontrollierten. Wir sammelten benutzte Nester, bestimmten die vorhandenen Arthropoden und maßen den Anteil der beim Bau verwendeten höher verarbeiteten anthropogenen Materialien. Etwa 25% des Nistmaterials bei Kohlmeisen stammte aus einer anthropogenen Quelle und zwar durchweg in allen Habitaten, während Blaumeisen nur wenig davon (1–2%) benutzten, außer in Gärten (~16%), was darauf hindeutet, dass Kohlmeisen diese Materialien bevorzugt sammelten. Bei benutzten Nestern hing die zunehmende Verwendung anthropogenen Materials mit einer geringeren allgemeinen Arthropodendiversität und Häufigkeit ektoparasitischer Prädatoren (nur bei Blaumeisen), jedoch größerer Mengen an Siphonapteren (Flöhen) zusammen. Eine höhere Arthropodendiversität stand mit geringeren Flohzahlen in Verbindung, was nahelegt, dass die höhere Diversität bei der Begrenzung der Flohzahlen eine Rolle spielt. Wir fanden weder einen direkten Zusammenhang zwischen Bruterfolg und der Verwendung anthropogener Materialien, noch zur Arthropodendiversität oder -häufigkeit. Allerdings nahm der Bruterfolg bei beiden Arten mit zunehmender Verstädterung ab und nahm bei den Blaumeisen mit dem Nestgewicht zu. Das Zusammenspiel zwischen Urbanisierung und der Ökologie der Vögel ist komplex; unsere Arbeit zeigt, dass geringfügige anthropogene Einflüsse indirekte und unerwartete Folgen für Vogelarten in Siedlung haben können.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Today, over half of the world’s human population lives in ever-growing towns and cities [United Nations (UN) 2011], which are increasingly recognised as being of considerable value for bird diversity and abundance (Gregory and Baillie 1998; Davies et al. 2009; Evans et al. 2011; Aronson et al. 2014). The changes in habitat and resource availability associated with urbanisation have a marked effect on the life histories of urban birds (Chace and Walsh 2006). Urban-dwelling birds tend to lay eggs earlier, produce smaller clutches and lighter nestlings, and have lower average productivity per nesting attempt than their non-urban conspecifics (Chamberlain et al. 2009). The lower availability of natural foods in urbanised areas results in lower food provisioning to nestlings, and while supplementary feeding can be ubiquitous in urban areas (Davies et al. 2009; Orros and Fellowes 2015a; Hanmer et al., in review), the benefits for bird productivity are not clear (Robb et al. 2008; Harrison et al. 2010; Plummer et al. 2013).
Many wild bird species utilise a diversity of urban habitats, and private suburban gardens are particularly important in this context (Cannon et al. 2005; Chamberlain et al. 2005). Aside from the provision of supplementary food, other resources provided by people help support some urban bird populations. In the UK it is estimated that more than one in five gardens contains a bird nest box, equating to a minimum of 4.7 million nest boxes, nationally equivalent to the provision of one nest box for every six breeding pairs of cavity-nesting birds (Davies et al. 2009). The availability of suitable nesting sites limits breeding density (Newton 1998) and with the removal of mature and dead trees typical of suburbia, there will be fewer natural nest sites in such areas (Wiebe 2011). Buildings may provide some cavities, but modern or refurbished houses tend to have fewer potential nesting holes to compensate for this loss of nesting sites (Mason 2006; Shaw et al. 2008). As a result, the provision of nest boxes in urban areas may be a particularly valuable resource, allowing cavity nesters to prosper (Chace and Walsh 2006; Wiebe 2011) and providing an opportunity to investigate factors affecting urban bird breeding biology.
Despite the potential importance of nest site provisioning and the influence of urbanisation on wild bird populations, our understanding of the effect of urbanisation on bird nesting biology remains relatively limited (reviewed in Deeming and Reynolds 2015). Factors that potentially affect breeding success are of considerable interest, and species that utilise nest boxes offer a practical way to explore these effects (Croci et al. 2008). There are several possible ways that urbanisation can influence bird nests. There is evidence for geographic variation within species in nest construction, with nests constructed in cooler regions typically being larger and better insulated (Deeming et al. 2012; Mainwaring et al. 2014; Biddle et al. 2016). Due to the urban heat island (UHI) effect increasing local environmental temperatures, it is plausible that nests from more highly urbanised areas may need less insulation, leading to smaller, lighter nests requiring less energy investment to build. However, there is little if any evidence that a change of the magnitude associated with the UHI effect influences bird reproduction (Deviche and Davies 2014).
Increased urbanisation may also be associated with a change in the proportion of anthropogenic material incorporated into nests (Reynolds et al. 2016). Given the general decline in biodiversity seen with increasing urbanisation (McKinney 2008), the availability of some key natural nesting materials may diminish, while the availability of potentially suitable anthropogenic alternatives is likely to increase. Depending on behavioural preferences and nest location this may cause some birds to expend more energy finding suitable natural nesting material, or instead they may utilise whatever is readily available (Britt and Deeming 2011), and so with higher levels of urbanisation incorporate more anthropogenic material into their nests (Wang et al. 2009; Reynolds et al. 2016). Given the potentially high insulation value of some anthropogenic materials they may be preferred and such preferences (if any) may differ between species (Surgey et al. 2012; Suárez-Rodríguez et al. 2013). Anthropogenic material incorporated into nests may also be hazardous to bird survival and breeding success, particularly by causing entanglement (Votier et al. 2011; Townsend and Barker 2014), but possibly also through more subtle effects on bird health. For example, House Sparrows Passer domesticus incorporating discarded cigarette butts into their nests reduce their ectoparasite load at the cost of exposure to toxins (Suárez-Rodríguez et al. 2017). For the purposes of this study we define anthropogenic material as highly processed anthropogenic material (e.g. polyester, nylon, highly processed cotton), although we acknowledge that hair from domestic animals and material from exotic garden plants could be considered to be anthropogenic in origin.
Changes in nest construction may in turn influence the invertebrate assemblage present in nests, of which changes in the presence and abundance of bird ectoparasites is of primary interest in this context. The influence of nest construction on ectoparasite load has been explored in a number of studies (e.g. Moreno et al. 2009; Suárez-Rodríguez et al. 2013) but only one has previously been carried out in a European urban environment (Reynolds et al. 2016), focusing on fleas and Blue Tits. Generalist nest-dwelling ectoparasites such as fleas (Siphonaptera) and biting mites (Acari, principally the family Dermanyssidae) are common in passerine nests (Moreno et al. 2009; Cantarero et al. 2013), but the latter are rarely considered.
Ectoparasites may influence reproductive success in a number of ways, such as by reducing nestling growth and survival (Merino and Potti 1995) or by affecting adult health (Tomás et al. 2007), possibly leading to nest desertion (Oppliger et al. 1994). Nest composition may influence ectoparasite load by affecting larval mortality and growth through effects on nest humidity (Heeb et al. 2000) or through the attraction/repellence effects of the materials themselves (Remeš and Krist 2005; Mennerat et al. 2009a; Tomás et al. 2012). As nest construction and host defence behaviour may differ significantly between species utilising similar nest sites in a local area, ectoparasite loads may also differ (Moreno et al. 2009).
Bird nests are also potentially home to a diverse array of non-parasitic arthropods (Tryjanowski et al. 2001). To our knowledge, no studies have considered the influence of nest construction on this diversity in the context of urbanisation. In turn, it is unclear what (if any) effect these arthropods have directly or indirectly on the breeding success of birds. Notably, Krištofík et al. (2017) found that adding saprophagous larvae of Fannia sp. to nests containing young European Bee-eaters Merops apiaster increased nestling condition, as the larvae consumed nest and nestling waste and detritus and so improved conditions in the nest cavity. While unstudied, some invertebrates may also predate other nest-dwelling invertebrates, including ectoparasites, depressing their numbers and thus potentially increasing bird productivity. Just how urbanisation affects the diversity of nest-dwelling invertebrates is not understood; it is possible that their diversity will decline with increasing urbanisation, as more generally found with invertebrates (McKinney 2008; Jones and Leather 2012). Irrespective of this relationship, we hypothesise that increased diversity in the nest may be associated with increased nesting success.
Taken together, it is evident that nest material could influence breeding productivity, and may also influence ectoparasite load. What is not clear is if there is a consistent influence of urbanisation on these factors, if species show similar responses, or if the wider nest arthropod community also varies with urbanisation. To examine this we compared (1) nesting materials, (2) ectoparasite load, (3) the assemblage of other arthropods and (4) fledging success, of two common British urban ‘adapter’ passerine species, the Blue Tit and Great Tit, at different levels of urbanisation in and around a large urban area.
Methods
Study areas
This study was conducted in and around the large urban district centred on Reading, South East England, between April and June 2016. Greater Reading covers approximately 72 km2 and has a population of ~290 000 people (Office for National Statistics 2013; following Orros and Fellowes 2015a).
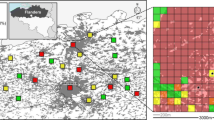
Volunteer garden owners were recruited across Reading through other studies previously run by the People and Wildlife Research Group, leafleting and word of mouth. Additional nest boxes were monitored on the grounds of the University of Reading, the Hill Primary School and Beale Wildlife Park and Gardens in addition to Maiden Erlegh Lake, Lavell’s Lake and Hosehill Lake local nature reserves (Fig. 1).

The distribution of study sites in and around the Greater Reading area indicated according to broadly defined habitat types
Nest monitoring
Nest boxes were monitored from early April until final fledging in late June. Only one breeding attempt per nest box was monitored and all boxes were cleared of old nesting material prior to the breeding season. Approximately 350 potentially useable nest boxes were monitored for this study. Once a nesting attempt was found it was checked up to twice a week until fledging or confirmed failure following the British Trust for Ornithology (BTO) Nest Record Scheme (https://www.bto.org/volunteer-surveys/nrs). All surviving chicks were ringed at approximately 14 days old under BTO ringing permit C5258. Dead chicks were removed whenever found during monitoring. All nest boxes were measured to establish their internal dimensions and their locations plotted using a global positioning system (GPSMAP 60CSx; Garmin, USA). Within sites, boxes were of effectively the same design and size but there was some variation between sites which was controlled for in the mixed-models analysis by the inclusion of site as a random factor.
Nest materials and arthropod load
Successful nests were removed between 2 days and 3 weeks from the estimated fledging date and sealed in plastic bags (Moreno et al. 2009). Failed nests that had grown chicks were removed when all the chicks were found to have died or had been predated. Nests that failed at the egg stage were removed when they had apparently been abandoned for at least a month beyond the estimated hatch date and there was no evidence of adult attendance. Fully predated nests were removed when they were found, provided significant damage had not been done to the nest that had lead to the removal of material. All remaining unhatched eggs and dead chicks were disposed of during nest removal. Nests were removed during the breeding season under Natural England license 2016-23468-SCI–SCI (granted to H. J. H.). All bagged nests were stored in a cold room at ca. 5 °C for up to 4 weeks. They were then placed in Tullgren funnels equipped with 60-W bulbs to extract invertebrates. Nests were dried for 48 h then weighed to the nearest 0.01 g on an electric balance. Nest materials were then broken down into apparently natural material from the environment (plant matter and natural fibres such as animal hair) and processed anthropogenic material (primarily dyed, treated cotton and artificial materials); each element was weighed separately.
Extracted invertebrate samples from each nest were collected and stored in tubes containing 40 ml of 70% ethanol solution. Due to variation in collection time in failed and some fledged nests, invertebrate data were only collected from the successful nests removed within 3 weeks of fledging. Arthropods were sorted under a dissecting microscope (maximum magnification 100×; Nikon SMZ645; Nikon, Japan) to order and subsequently identified to lower taxonomic levels where practical, particularly in the case of likely ectoparasites. All arthropods were individually counted, with the exception of flea larvae where numbers were on rare occasions very large. In the latter situation a subsample of 25% of the material was counted, and total numbers of larvae then extrapolated from these data. Shannon diversity indices at the order level were calculated for each fledged nest examined for arthropods.
Habitat data for each collected nest were established for a 200-m radius around each nest box in ArcGIS 10.0 (ESRI 2011) using data from the Ordnance Survey Mastermap collection (EDINA, University of Edinburgh) land use data. Habitat/land use categories within 200 m of collected nests were grouped together to form four broad categories: constructed surfaces (buildings, roads, driveways and pavements); natural surfaces (trees, scrub and grassland); water bodies (primarily lakes and ponds); and private gardens (defined as garden mixed surfaces).
Analysis
All analyses were carried out within program R, version 3.3 (R Core Team 2016). Mann–Whitney U-tests were used to compare between bird species and within species between broad habitat types (garden, greenspace and rural) for productivity and nest construction. Spearman’s rank correlation was used to examine the relationship between percentage habitat cover and nest construction across all collected nests. For the subset of fledged nests collected under a standardised methodology and fully examined for arthropods, Mann–Whitney U-tests were carried out to compare between species overall and within species between broad site types for ectoparasite abundance, a measure of arthropod predator abundance (total numbers of Pseudoscorpionida, Staphylinidae and Histeridae) and arthropod diversity. For all multiple comparisons, p was automatically corrected for the false discovery rate within R.
Separate mixed-effects models for Blue Tits and Great Tits were carried out using R package lme4 (version 1.1-12; Bates et al. 2015) to examine effects of nest construction and habitat separately on the proportional usage of anthropogenic material and overall nest fate (both binomial models) along with the number of eggs laid and number of chicks fledged in failed and successful nests (both Poisson models). Predated nests were excluded from models examining nest fate and the number of chicks fledged to control for the influence of direct nest predation on breeding success (Lambrechts et al. 2016a). Separate additional mixed-effect models were run for the subset of nests examined for arthropod diversity and ectoparasite load. These examined Shannon diversity (linear model), overall flea abundance (Poisson model) and the presence/absence of Dermanyssidae mites (binomial model). To explore influences of these factors along with nest construction and habitat, the final models considered all potential factors influencing number of eggs laid and chicks fledged just for these nests.
In all mixed-effect models, individual study site (rather than broad site type) was treated as a random effect to account for the potential non-independence of nests from the same site and for the slight variation in box design between sites. In overdispersed models, individual nest identity was added as an additional random effect (Harrison 2014). Nest boxes within 200 m of each other, or on the same property/reserve, were considered to be from the same site. Model selection was carried out on the global models using delta Akaike information criteria (∆AICc) and model weights (Burnham and Anderson 2002). From the models within ∆2 AICc of the minimal model, predicted lines of best fit with 95% confidence intervals were calculated for plotting. With the exception of Shannon diversity models, the proportion of constructed surfaces was used as a measure of urbanisation. In the Shannon models the total proportion of green surfaces (natural surfaces and gardens) was found to produce more stable models and so was utilised instead, whereas the reverse was true for models considering all other dependent variables. For models considering the number of chicks fledged, clutch size was used as a fixed factor to account for any relationship between the two.
Results
Overall nests
In total 98 nests (62 Blue Tit and 36 Great Tit) were fully monitored and deconstructed. Of these at least one chick apparently fledged in 60 (35 and 25, respectively) nests, while in the other 38 (27 and 11, respectively) nests, the egg or chick stage failed. Five Blue Tit and one Great Tit nest seemingly failed due to direct predation. Breeding and nest construction parameters for collected nests at different levels of urbanisation are summarised in Table 1.
Nest composition
With Blue Tits rural nests were significantly heavier than urban greenspace nests (W = 368, p = 0.04, other comparisons p > 0.1; Table 1) while Great Tit nests showed no significant differences between habitat types (all p > 0.1; Table 1). There was no significant difference in nest box size (using interior base surface area) between broad habitat types in either species (both p > 0.1; Table 1).
Nest materials included mosses, grasses, leaves, twigs, feathers, animal hair (both domestic and wild) and anthropogenic materials, which were largely treated cotton and artificial stuffing materials. Anthropogenic material was found in 77 and 94% of Blue Tit and Great Tit nests, respectively (84% of all nests). Blue Tit nests contained proportionally less anthropogenic material and showed considerably more variation in rates of use than Great Tit nests (W = 590.5, p = 0.001; Table 1). There was no effect of habitat type on the use of anthropogenic materials for either species (p > 0.09 for all comparisons). In a logistic mixed-model regression controlling for site, neither species showed a significant association between either broad habitat type or the actual level of urbanisation as a proportion of constructed surfaces and the proportion of anthropogenic materials used.
Nest arthropod diversity and ectoparasite load
Due to the more standardised collection of nests and the loss of several invertebrate samples, only 42 fledged nests (23 Blue Tit and 19 Great Tit) from 15 different sites were explored for their arthropod diversity and numbers. Arthropods from 19 different orders were found in nests (Table 2). Parasitic Dermanyssidae mites were found in 55% of nests, so this data was treated as presence/absence data. Adult Siphonaptera (fleas) or their larvae were found in all but one nest. All adults were identified as members of the Ceratophyllus and were most likely Hen Fleas Ceratophyllus gallinae (Harper et al. 1992), with the exception of one individual Dasypsyllus gallinulae. As they are functionally alike and flea larvae could not readily be separated to species, and a number of nests contained high numbers of larvae but no adults, all fleas were combined together into a single category to form an overall measure of flea abundance in nests. Mallophaga (biting/bird lice) and Analgoidea (feather mites) were only found in single nests and so were not included in the ectoparasite analysis. Amongst the other arthropods found, potential predators of mites or fleas were identified in 52% of nests (adult predators only). Staphylinidae (Rove Beetles) were the most frequently recorded coleopterans, and these prey on other insects and mites. Other potential predators included Histeridae beetles, Pseudoscorpions (Pseudoscorpiones) and some predatory mites of the suborder Prostigmata (Table 2).
There was no difference in Shannon diversity at the order level between fledged Blue Tit and Great Tit nests (H s = 1.29 and H s = 1.28, respectively) and within species no significant difference between the broad site types (p > 0.1 in all cases; Table 3). For Blue Tit nests, the AICc-selected logistic regression mixed-effect model for factors influencing Shannon diversity found that the proportion of green surfaces within 200 m (used instead of constructed surfaces due to poor model stability) were positively associated with increased diversity [χ 2(1) = 5.34, p = 0.024, following Bates et al. 2015; Fig. 2; Table 4]. No effect was found for Great Tits.

The influence of the proportion of green surfaces (natural surfaces and gardens) on nest arthropod Shannon diversity index. Plotted line of best fit (with 95% confidence intervals) is based on predictions extracted from the linear mixed-effect model with Shannon diversity as the dependent variable. Site and nest box were random factors. The broad habitat types around the nest boxes are indicated
There was no significant difference between the overall abundance of fleas or Dermanyssidae mites in nests between bird species, or within species across habitat types (p > 0.1 in all cases; Table 3). Fleas showed a significant negative correlation with Shannon diversity overall and specifically in Blue Tits (r s = −0.37, p = 0.016 and r s = −0.42, p = 0.048, respectively). No significant correlation was found in the Dermanyssidae and there was no correlation between the abundance of mites and fleas.
An increase in potential predator abundance (total Pseudoscorpionida, Staphylinidae and Histeridae) was associated with a decrease in the proportion of anthropogenic nest material in the Blue Tit nests (r s = −0.52, p = 0.012; Fig. 3). There was a high outlier in the predator numbers and the correlation was still highly significant following its removal (r s = −0.55, p = 0.008; see Fig. 3). Predator abundance or presence/absence was not significantly associated with or affected by any other variables including ectoparasite abundance (all p > 0.09). No relationship was found between predator abundance or presence/absence and any other measured variable in Great Tits (p > 0.1).

The influence of the proportion of anthropogenic nest material on the abundance of predators (total abundance of Pseudoscorpionida, Staphylinidae and Histeridae) living in Blue Tit nests, fitted with a linear line of best fit. The broad habitat types around the nest boxes are indicated
In Blue Tits the minimal model for flea abundance could not be distinguished from the null model (∆AICc <2). In Great Tits increasing levels of urbanisation (constructed surfaces) led to lower flea abundance, and another model within ∆AICc 2 additionally found a significant positive relationship between the proportion of anthropogenic material (p = 0.001 and p = 0.052, respectively; Fig. 4; Table 5). No model was found containing a significant predictor following selection for the presence of Dermanyssidae mites in the nests of either species. No relationship was found for either ectoparasite type with the weight of the nests, numbers of eggs laid or number of chicks fledged.

The influence of a the proportion of constructed surfaces within 200 m of the nest box, and b the proportion of anthropogenic material comprising nests on overall flea abundance in fledged Great Tit nests. Plotted lines of best fit (with 95% confidence intervals) were based on predictions extracted from the respective mixed-effect model for these variables. Site and nest box were random factors. Broad habitat types around the nest boxes are indicated
Influences on breeding success
Overall, Blue Tits laid significantly more eggs that Great Tits (W = 1711.5, p < 0.001; Table 1) per breeding attempt, but there was no overall difference in the number of chicks they fledged, nor in the proportion of nests that fledged at least one chick. In successful nests there was no significant correlation between the number of eggs laid and the number of chicks fledged in either species (both p > 0.1). The proportion of anthropogenic material was not a significant predictor of any measure of breeding performance.
There was no significant difference in the number of eggs laid in either species across sites, but rural breeding birds fledged significantly more chicks per breeding attempt than urban greenspace and garden breeding birds for both Blue Tits (W = 368, p = 0.032 and W = 201.5, p = 0.028, respectively; Table 1) and Great Tits (W = 132, p = 0.001 and W = 39, p = 0.007, respectively; Table 1). Increasing proportions of constructed surfaces as a measure of level of urbanisation did not significantly influence the number of eggs laid, but it did lead to fewer chicks being fledged in both species (r s = −0.32, p = 0.01 and r s = −0.45, p = 0.006 for Blue Tits and Great Tits, respectively).
For Blue Tits, an increased level of urbanisation was associated with reduced overall breeding success, (p = 0.022; Fig. 5). Heavier nests were associated with more eggs being laid (p = 0.02; Fig. 6a; Table 6) and more chicks fledged (p = 0.008; Fig. 6b; Table 6). Increased urbanisation also resulted in fewer chicks fledging (p = 0.002; Fig. 6c; Table 6).

The influence of the proportion of constructed surfaces within 200 m of the nest box on the outcome of Blue Tit breeding success, where 1 indicates fledging at least one chick and 0 indicates failure. The plotted line of best fit (with 95% confidence intervals) was based on predictions extracted from the binomial mixed-effect model for nest outcome. Site and nest box were random factors. Broad habitat types around the nest boxes are indicated. Nests that failed through predation were excluded from this model

The influence of dry nest weight on a the number of eggs laid, and b the number of chicks fledged by Blue Tits. The influence of the proportion of constructed surfaces within 200 m of the nest box on the number of chicks fledged by c Blue Tits and d Great Tits. The plotted lines of best fit (with 95% confidence intervals) were based on predictions extracted from the respective mixed-effect model for these variables. Site and nest box were random factors. Broad habitat types around the nest boxes are indicated. Predated nests were excluded from the models considering the number of chicks fledged
There was an indication of a negative association between the level of urbanisation and the number of chicks fledged in Great Tits (p = 0.05; Fig. 6d), but the null model was within ∆2 AICc of that minimal model, indicating low model support. While only nests in which one or more chicks successfully fledged were studied, no evidence of an effect of arthropod or ectoparasite numbers, nest construction or level of urbanisation on the number of chicks fledged was found.
Discussion
Our work supports the view that increased urbanisation is generally associated with a reduction in the overall breeding success of wild birds (reviewed in Chamberlain et al. 2009). We were surprised to find that while Great Tits and Blue Tits differed in their use of anthropogenic materials to construct nests (Great Tit > Blue Tit), this was not associated with urbanisation, suggesting perhaps an element of choice in nesting materials beyond simply their availability. There was no difference in nest mass among habitats for Great Tits, but Blue Tit nests in rural areas were significantly heavier, and overall, heavier nests were associated with greater fledging rates. Both bird ectoparasites and nest-dwelling arthropods more generally showed evidence of declining with increasing loss of green space. However, there was a negative correlation between flea load and overall arthropod diversity in nests, suggesting that arthropods may have been predating flea larvae and/or eggs, or acting as competitors for resources. At the same time, while overall arthropod diversity and predator numbers were both negatively associated with increased proportional use of anthropogenic nesting material in nests, flea numbers conversely increased. We speculate that the reduction in the use of natural nesting materials reduces overall diversity in the arthropod nest box assemblage, which in turn reduces the effects of predation and/or competition on flea abundance.
Understanding of the role of nests has increased considerably in recent years, but the nest-building phase of breeding success is relatively understudied (Deeming and Reynolds 2015). Nests represent a major energy investment (Mainwaring and Hartley 2013) and understanding their role, particularly in the light of urbanisation, may help us to understand changes in productivity across habitat gradients and their conservation implications. In addition to finding a broad decline in reproductive output associated with urbanisation, as found elsewhere (Chace and Walsh 2006; Chamberlain et al. 2009), we also similarly found heavier nests to be linked with higher reproductive output. This concurs with a number of previous nest box studies, though not all (Lambrechts et al. 2016b), and may be linked to adult quality (Alvarez et al. 2013; Gladalski et al. 2016; Lambrechts et al. 2016a). Such differences in turn may be linked to differences in nesting behaviour and reproductive output between these species, as Blue Tits typically build proportionally heavier nests and have a higher average reproductive output per breeding attempt than the larger Great Tit (Lambrechts et al. 2014, 2015; Gladalski et al. 2016).
While the majority of nesting materials are intended to provide insulation and structure, nest components may also provide additional benefits. Aromatic plants may offer an anti-ectoparasite or even anti-bacterial function in nests (Mennerat et al. 2009a; Tomás et al. 2012; Ruiz-Castellano et al. 2016). Fewer native aromatic plants may be available at higher levels of urbanisation even though the overall plant diversity can be higher due to the planting of exotic plant species in gardens and other areas (McKinney 2008). The prevalence and importance of aromatic plants in the nests of breeding Blue Tits and Great Tits in the UK is unknown. However, Blue Tits have been found to utilise them elsewhere in their range (Mennerat et al. 2009a, b; Tomás et al. 2012). Artificial nesting materials do not offer defences against macro- or micro-parasites, and so their benefits are in terms of nest structure and insulation. Furthermore, as processed anthropogenic materials are largely artificial in origin, they will also not provide food resources for consumers (principally arthropods) and decomposers (mainly bacteria and fungi), and hence may affect the complexity of the structure of the assemblage of species (including predatory species) that inhabit the nest boxes.
We found variation in the use of anthropogenic nest components similar to that found in other studies on Blue Tit and related species here in the UK and in Europe more generally (Moreno et al. 2009; Britt and Deeming 2011; Reynolds et al. 2016) although this variation is understudied (Deeming and Mainwaring 2015). Like Reynolds et al. (2016) we found that anthropogenic material was present throughout the site types and in the majority of nests irrespective of the local level of urbanisation. Great Tit nests contained a median of 24% anthropogenic material and this varied little across habitats, suggesting that they may preferentially seek these materials when nest building, particularly when they are uncommon (Surgey et al. 2012). This may be due to a preference for the materials’ insulation properties and thus fitness benefits (Reynolds et al. 2016). In contrast, Blue Tit nests contained an overall median of 2% anthropogenic material, and while not statistically significant due to considerable variation among nests, we note that this increased to 16% in urban gardens, suggesting that Blue Tits are more opportunistic in their use of non-natural materials. This may be due to the decline of natural nesting material availability or an increase in the general availability of artificial substitutes in urban areas (Wang et al. 2009).
In contrast to Reynolds et al. (2016), we do find a possible effect of anthropogenic materials on the presence of ectoparasites and other arthropods. In Blue Tits a higher proportion of anthropogenic nest material is associated with lower arthropod diversity. While the majority of non-parasitic arthropods appeared to be opportunistic in nature, their numbers also included potential predators of flea larvae and eggs. These also appeared to decline with increasing use of anthropogenic materials. While a relationship between predator and flea numbers was not found, increased arthropod diversity was correlated with a decline in flea numbers. This may be a result of competition for resources [the consumption of adult flea frass which would otherwise be consumed by flea larvae (Tripet and Richner 1999), in a manner analogous to that found by Krištofík et al. (2017)] or predation of flea eggs and larvae.
Such changes in the degree of interspecific interactions may provide a functional explanation of the increase in flea numbers associated with an increase in the proportion of anthropogenic materials used in nests. We suggest that nests constructed with more natural materials support a more structurally diverse arthropod community (including more predators) and that this should reduce flea numbers irrespective of the level of urbanisation. Given this, it would be logical to expect birds to prefer natural materials where available. This makes the possible preference for anthropogenic materials shown by Great Tits unexpected if they are less exposed to them and more energy is expended in finding these materials (Surgey et al. 2012). As they appeared to seek these materials out regardless of overall availability in the local environment, we speculate that they may derive other advantages from utilising them.
Such a relationship between anthropogenic materials and parasite load, even if indirect, may help explain findings by others on variation in nest ectoparasite loads with different materials (Moreno et al. 2009; Tomás et al. 2012; Cantarero et al. 2013; Reynolds et al. 2016). Nevertheless, it is not clear if nest material choice and associated changes in arthropod assemblage help explain variation in breeding success associated with urbanisation. This may simply be due to food availability being the most important factor driving the reduction in fledgling success seen in urban areas (Chace and Walsh 2006; Chamberlain et al. 2009) or that the effects of parasites are frequently hidden until nestlings become highly stressed (Simon et al. 2004; Arriero et al. 2008; Bańbura et al. 2011).
We acknowledge that our sampling approach was constrained; as nests could not be removed until after fledging we cannot be certain that the biodiversity recorded in removed nests represents that present in nests when they were being used [although previous work suggests the loss of fleas should have been low (Wesołowski and Stańska 2001)]. Furthermore, Tullgren funnels may not be the most effective way of sampling adult flea abundance (Harper et al. 1992; Moreno et al. 2009), which may explain the comparatively low numbers of adult fleas found in this study compared to others that used freezing and manual deconstruction of nests to find them (Reynolds et al. 2016). Nevertheless, the counting of flea larvae does provide a measure of nest flea abundance in the absence of adults, and this approach did allow us to collect the other nest arthropods present, which for species such as mites would have been very challenging to sample using other approaches (Moreno et al. 2009).
Cavity nesters that take easily to artificial nests sites provide an important research tool for exploring the effects of various factors on breeding success (Vaugoyeau et al. 2016), but it is important to bear in mind that nest box-based studies may not be directly comparable to those on nests in natural cavities (Wesołowski and Stańska 2001; Maziarz et al. 2017). Nevertheless, in urban areas nest boxes may be the main nesting cavities available to Great and Blue Tits (Davies et al. 2009). Given the different responses detected in this study and by others of these two related species, it is important to consider that different bird species may respond differently to similar pressures (Lambrechts et al. 2015; Gladalski et al. 2016), so we must be mindful of drawing broad conclusions from single-species studies (Reynolds et al. 2016; Vaugoyeau et al. 2016). Due to the relatively poor breeding season during our fieldwork, sample numbers were lower than anticipated, which reduced the statistical power of our study. A larger/multiple year study also considering the reasons for nest failure would allow for a more considered reflection on how these factors interact to affect the breeding success of urban birds.
Being an urban adapter brings novel opportunities [e.g. supplementary food (Orros and Fellowes 2015b; Hanmer et al. 2017, in review)] and challenges [e.g. domestic cats (Thomas et al. 2014, Hanmer et al., in review]. Nest boxes provide a clear opportunity to birds when breeding sites are limited, but it is evident that increased urbanisation is associated with a decrease in the number of offspring successfully fledged in species which use nest boxes such as Blue and Great Tits (Cowie and Hinsley 1987; Chamberlain et al. 2009; this study). We examined how potential variation in nest construction may contribute to this reduction in breeding success, both directly (nesting materials affecting successful breeding) or indirectly (nests and nest boxes as homes to ectoparasites and other arthropods). Blue Tit nests in less urbanised areas tended to be heavier, with heavier nests leading to more eggs being laid and more chicks fledged, whereas no effect of urbanisation on nest weight was found in Great Tits. We found that Blue and Great Tits differ in how they utilise anthropogenic materials, with Great Tits showing higher rates of use, but no effect of urbanisation, whereas Blue Tits show some evidence of increased use in urban gardens where such material is likely to be more common. We found no evidence that the use of anthropogenic materials directly affected breeding success. However, while arthropod diversity and predator numbers declined with increasing use of anthropogenic nest materials, the number of fleas increased, suggesting that there may be indirect links between the materials used in nest construction and parasite (flea) load. Nest boxes are clearly ecological communities in their own right; we suggest that these may be more complex than they first appear, and worthy of consideration for further investigation.
References
Alvarez E, Belda EJ, Verdejo J, Barba E (2013) Variation in Great Tit nest mass and composition and its breeding consequences: a comparative study in four Mediterranean habitats. Avian Biol Res 6:39–46
Aronson MFJ, La Sorte FA, Nilon CH, Katti M, Goddard MA, Lepczyk CA, Warren PS, Williams NSG, Cilliers S, Clarkson B, Dobbs C, Dolan R, Hedblom M, Klotz S, Kooijmans JL, Kühn I, MacGregor-Fors I, McDonnell M, Mörtberg U, Pyšek P, Siebert S, Sushinsky J, Werner P, Winter M (2014) A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc R Soc B 281:20133330
Arriero E, Moreno J, Merino S, Martínez J (2008) Habitat effects on physiological stress response in nestling Blue Tits are mediated through parasitism. Physiol Bioch Zool 81:195–203
Bańbura J, Bańbura M, Glądalski M, Kaliński A, Markowski M, Michalski M, Nadolski J, Skwarska J, Zieliński P (2011) Body condition parameters of nestling Great Tits Parus major in relation to experimental food supplementation. Acta Ornithol 46:207–212
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48
Biddle LE, Goodman AM, Deeming DC (2016) Geographical effects on the mass and dimensions of finch (Fringillidae, Passeriformes) and thrush (Turdidae, Passeriformes) nests. Avian Biol Res 9:13–21
Britt J, Deeming DC (2011) First-egg date and air temperature affect nest construction in Blue Tits Cyanistes caeruleus, but not in Great Tits Parus major. Bird Study 58:78–89
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Cannon AR, Chamberlain DE, Toms MP, Hatchwell BJ, Gaston KJ (2005) Trends in the use of private gardens by wild birds in Great Britain 1995–2002. J Appl Ecol 42:659–671
Cantarero A, López-Arrabé J, Rodríguez-García V, González-Braojos S, Ruiz-De-Castañeda R, Redondo AJ, Moreno J (2013) Factors affecting the presence and abundance of generalist ectoparasites in nests of three sympatric hole-nesting bird species. Acta Ornithol 48:39–54
Chace JF, Walsh JJ (2006) Urban effects on native avifauna: a review. Landsc Urban Plan 74:46–69
Chamberlain DE, Vickery JA, Glue DE, Robinson RA, Conway GJ, Woodburn RJW, Cannon AR (2005) Annual and seasonal trends in the use of garden feeders by birds in winter. Ibis 147:563–575
Chamberlain DE, Cannon AR, Toms MP, Leech DI, Hatchwell BJ, Gaston KJ (2009) Avian productivity in urban landscapes: a review and meta-analysis. Ibis 151:1–18
Cowie RJ, Hinsley SA (1987) Breeding success of Blue Tits and Great Tits in suburban gardens. Ardea 75:81–90
Croci S, Butet A, Clergeau P (2008) Does urbanization filter birds on the basis of their biological traits? Condor 110:223–240
Davies ZG, Fuller RA, Loram A, Irvine KN, Sims V, Gaston KJ (2009) A national scale inventory of resource provision for biodiversity within domestic gardens. Biol Conserv 142:761–771
Deeming DC, Mainwaring MC (2015) Functional properties of nests. In: Deeming DC, Reynolds SJ (eds) Nests, eggs, and incubation: new ideas about avain reproduction. Oxford University Press, Oxford, pp 29–49
Deeming DC, Reynolds SJ (2015) Nests, eggs, and incubation: new ideas about avain reproduction. Oxford University Press, Oxford
Deeming DC, Mainwaring MC, Hartley IR, Reynolds SJ (2012) Local temperature and not latitude determines the design of Blue Tit and Great Tit nests. Avian Biol Res 5:203–208
Deviche P, Davies S (2014) Reproductive phenology of urban birds: environmental cues and mechanisms. In: Gil D, Brumm H (eds) Avian urban ecology: behavioural and physiological adaptations. Oxford University Press, Oxford, pp 98–115
ESRI (2011) ArcGIS desktop: release 10. Environmental Systems Research Institute, Redlands
Evans KL, Chamberlain DE, Hatchwell BJ, Gregory RD, Gaston KJ (2011) What makes an urban bird? Glob Change Biol 17:32–44
Gladalski M, Bańbura M, Kalinski A, Markowski M, Skwarska J, Wawrzyniak J, Zielinski P, Cyzewska I, Banbura J (2016) Effects of nest characteristics on reproductive performance in Blue Tits Cyanistes caeruleus and Great Tits Parus major. Avian Biol Res 9:37–43
Gregory RD, Baillie SR (1998) Large-scale habitat use of some declining British birds. J Appl Ecol 35:785–799
Hanmer HJ, Thomas RL, Fellowes MDE (2017) Provision of supplementary food for wild birds may increase the risk of local nest predation. Ibis 159:158–167
Harper GH, Marchant A, Boddington DG (1992) The ecology of the Hen Flea Ceratophyllus gallinae and the Moorhen Flea Dasypsyllus gallinulae in nestboxes. J Anim Ecol 61:317–327
Harrison XA (2014) Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ 2:e616
Harrison TJE, Smith JA, Martin GR, Chamberlain DE, Bearhop S, Robb GN, Reynolds SJ (2010) Does food supplementation really enhance productivity of breeding birds? Oecologia 164:311–320
Heeb P, Kölliker M, Richner H (2000) Bird–ectoparasite interactions, nest humidity, and ectoparasite community structure. Ecology 81:958–968
Jones EL, Leather SR (2012) Invertebrates in urban areas: a review. Eur J Entomol 109:463–478
Krištofík J, Darolová A, Hoi C, Hoi H (2017) Housekeeping by lodgers: the importance of bird nest fauna on offspring condition. J Ornithol 158:245–252
Lambrechts MM, Demeyrier V, Fargevieille A, Giovannini P, Lucas A, Marrot P, Midamegbe A, Perret P, Charmantier A, Doutrelant C, Gregoire A (2014) Great Tits build shallower nests than Blue Tits. Avian Biol Res 7:251–254
Lambrechts MM, Blondel J, Dubuc-Messier G, Marrot P, de Franceschi C, Perret P, Doutrelant C, Gregoire A, Charmantier A (2015) Great Tits build shallower nests than Blue Tits in an insular oak-dominated habitat mosaic. Avian Biol Res 8:117–121
Lambrechts MM, Blondel J, de Franceschi C, Doutrelant C (2016a) Nest size is positively correlated with fledging success in Corsican Blue Tits (Cyanistes caeruleus) in an insular oak-dominated habitat mosaic. J Ornithol 158:1–8
Lambrechts MM, Marrot P, Fargevieille A, Giovannini P, Lucas A, Demeyrier V, Midamegbe A, Perret P, Grégoire A, Charmantier A, Doutrelant C (2016b) Nest size is not closely related to breeding success in Blue Tits: a long-term nest-box study in a Mediterranean oak habitat. Auk 133:198–204
Mainwaring MC, Hartley IR (2013) The energetic costs of nest building in birds. Avian Biol Res 6:12–17
Mainwaring MC, Deeming DC, Jones CI, Hartley IR (2014) Adaptive latitudinal variation in Common Blackbird Turdus merula nest characteristics. Ecol Evol 4:851–861
Mason CF (2006) Avian species richness and numbers in the built environment: can new housing developments be good for birds? Biodivers Conserv 15:2365–2378
Maziarz M, Broughton RK, Wesołowski T (2017) Microclimate in tree cavities and nest-boxes: implications for hole-nesting birds. For Ecol Manage 389:306–313
McKinney ML (2008) Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst 11:161–176
Mennerat A, Mirleau P, Blondel J, Perret P, Lambrechts MM, Heeb P (2009a) Aromatic plants in nests of the Blue Tit Cyanistes caeruleus protect chicks from bacteria. Oecologia 161:849–855
Mennerat A, Perret P, Lambrechts MM (2009b) Local individual preferences for nest materials in a passerine bird. PLoS One 4:e5104
Merino S, Potti J (1995) Mites and blowflies decrease growth and survival in nestling Pied Flycatchers. Oikos 73:95–103
Moreno J, Merino S, Lobato E, Ruiz-De-Castaneda R, Martinez-De la Puente J, Del Cerro S, Rivero-De Aguilar J (2009) Nest-dwelling ectoparasites of two sympatric hole-nesting passerines in relation to nest composition: an experimental study. Ecoscience 16:418–427
Newton I (1998) Population limitation in birds. Academic Press, London
Office for National Statistics (2013) Neighbourhood statistics. 2011 census data. HM Government
Oppliger A, Richner H, Christe P (1994) Effect of an ectoparasite on lay date, nest-site choice, desertion, and hatching success in the Great Tit (Parus major). Behav Ecol 5:130–134
Orros ME, Fellowes MDE (2015a) Widespread supplementary feeding in domestic gardens explains the return of reintroduced Red Kites Milvus milvus to an urban area. Ibis 157:230–238
Orros ME, Fellowes MDE (2015b) Wild bird feeding in a large UK urban area: characteristics and estimates of energy input and individuals supported. Acta Ornithol 50:43–58
Plummer KE, Bearhop S, Leech DI, Chamberlain DE, Blount JD (2013) Winter food provisioning reduces future breeding performance in a wild bird. Sci Rep 3:2002
R Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Remeš V, Krist M (2005) Nest design and the abundance of parasitic Protocalliphora blow flies in two hole-nesting passerines. Ecoscience 12:549–553
Reynolds SJ, Davies CS, Elwell E, Tasker PJ, Williams A, Sadler JP, Hunt D (2016) Does the urban gradient influence the composition and ectoparasite load of nests of an urban bird species? Avian Biol Res 9:224–234
Robb GN, McDonald RA, Chamberlain DE, Reynolds SJ, Harrison TJE, Bearhop S (2008) Winter feeding of birds increases productivity in the subsequent breeding season. Biol Lett 4:220–223
Ruiz-Castellano C, Tomas G, Ruiz-Rodriguez M, Martin-Galvez D, Soler JJ (2016) Nest material shapes eggs’ bacterial environment. PLoS One 11:e0148894
Shaw LM, Chamberlain D, Evans M (2008) The House Sparrow Passer domesticus in urban areas: reviewing a possible link between post-decline distribution and human socioeconomic status. J Ornithol 149:293–299
Simon A, Thomas D, Blondel J, Perret P, Lambrechts MM (2004) Physiological ecology of Mediterranean Blue Tits (Parus caeruleus L.): effects of ectoparasites (Protocalliphora spp.) and food abundance on metabolic capacity of nestlings. Physiol Biocheml Zool 77:492–501
Suárez-Rodríguez M, López-Rull I, Macías Garcia C (2013) Incorporation of cigarette butts into nests reduces nest ectoparasite load in urban birds: new ingredients for an old recipe? Biol Lett 9:20120931
Suárez-Rodríguez M, Montero-Montoya R, Macías Garcia C (2017) Anthropogenic nest materials may increase breeding costs for urban birds. Front Ecol Evol 5:4
Surgey J, Feu CRD, Deeming DC (2012) Opportunistic use of a wool-like artificial material as lining of Tit (Paridae) nests. Condor 114:385–392
Thomas RL, Baker PJ, Fellowes MDE (2014) Ranging characteristics of the Domestic Cat (Felis catus) in an urban environment. Urban Ecosyst 17:911–921
Tomás G, Merino S, Moreno J, Morales J (2007) Consequences of nest reuse for parasite burden and female health and condition in Blue Tits, Cyanistes caeruleus. Anim Behav 73:805–814
Tomás G, Merino S, Martínez-de la Puente J, Moreno J, Morales J, Lobato E, Rivero-de Aguilar J, del Cerro S (2012) Interacting effects of aromatic plants and female age on nest-dwelling ectoparasites and blood-sucking flies in avian nests. Behav Process 90:246–253
Townsend AK, Barker CM (2014) Plastic and the nest entanglement of urban and agricultural Crows. PLoS One 9:e88006
Tripet F, Richner H (1999) Dynamics of Hen Flea Ceratophyllus gallinae subpopulations in Blue Tit nests. J Ins Behav 12:159–174
Tryjanowski P, Baraniak E, Bajaczyk R, Gwiazdowicz DJ, Konwerski S, Olszanowski Z, Szymkowiak P (2001) Arthropods in nests of the Red-backed Shrike (Lanius collurio) in Poland. Belg J Zool 131:69–74
UN (2011) World population prospects: the 2010 revision. United Nations, Department of Economic and Social Affairs, Population Division, New York
Vaugoyeau M, Adriaensen F, Artemyev A, Bańbura J, Barba E, Biard C, Blondel J, Bouslama Z, Bouvier J-C, Camprodon J, Cecere F, Charmantier A, Charter M, Cichoń M, Cusimano C, Czeszczewik D, Demeyrier V, Doligez B, Doutrelant C, Dubiec A, Eens M, Eeva T, Faivre B, Ferns PN, Forsman JT, García-del-Rey E, Goldshtein A, Goodenough AE, Gosler AG, Grégoire A, Gustafsson L, Harnist I, Hartley IR, Heeb P, Hinsley SA, Isenmann P, Jacob S, Juškaitis R, Korpimäki E, Krams I, Laaksonen T, Lambrechts MM, Leclercq B, Lehikoinen E, Loukola O, Lundberg A, Mainwaring MC, Mänd R, Massa B, Mazgajski TD, Merino S, Mitrus C, Mönkkönen M, Morin X, Nager RG, Nilsson J-Å, Nilsson SG, Norte AC, Orell M, Perret P, Perrins CM, Pimentel CS, Pinxten R, Richner H, Robles H, Rytkönen S, Senar JC, Seppänen JT, Pascoal da Silva L, Slagsvold T, Solonen T, Sorace A, Stenning MJ, Tryjanowski P, von Numers M, Walankiewicz W, Møller AP (2016) Interspecific variation in the relationship between clutch size, laying date and intensity of urbanization in four species of hole-nesting birds. Ecol Evol 6:5907–5920
Votier SC, Archibald K, Morgan G, Morgan L (2011) The use of plastic debris as nesting material by a colonial seabird and associated entanglement mortality. Mar Poll Bull 62:168–172
Wang Y, Chen S, Blair RB, Jiang P, Ding P (2009) Nest composition adjustments by Chinese Bulbuls Pycnonotus sinensis in an urbanized landscape of Hangzhou (E China). Acta Ornithol 44:185–192
Wesołowski T, Stańska M (2001) High ectoparasite loads in hole-nesting birds—a nestbox bias? J Avian Biol 32:281–285
Wiebe KL (2011) Nest sites as limiting resources for cavity-nesting birds in mature forest ecosystems: a review of the evidence. J Field Ornithol 82:239–248
Acknowledgements
This work was supported by a studentship funded by the Keith Duckworth Trust and Songbird Survival. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the paper. The datasets collected for the current study are available from the corresponding author on request. Many thanks to all the volunteers for access to their nest boxes, Colin Prescott and Tim Alexander for allowing the direct monitoring of their respective nest boxes, Alejandra Perotti for identifying the mites and Chris Foster for providing assistance with general invertebrate identification. Additional thanks to Dave Coles and Tim Ball for providing additional rural nest samples from their nest box monitoring schemes at Beale Wildlife Park and Gardens and Hose Hill Lake Nature Reserve, respectively.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bairlein.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Hanmer, H.J., Thomas, R.L., Beswick, G.J.F. et al. Use of anthropogenic material affects bird nest arthropod community structure: influence of urbanisation, and consequences for ectoparasites and fledging success. J Ornithol 158, 1045–1059 (2017). https://doi.org/10.1007/s10336-017-1462-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-017-1462-7