
Abstract
Using a two-bottle choice test of short duration, we determined taste preference thresholds for sucrose, fructose, glucose, lactose, and maltose in three Western chimpanzees (Pan troglodytes verus). Further, we assessed relative preferences for these five saccharides when presented at equimolar concentrations and determined taste preference difference thresholds for sucrose, that is, the smallest concentration difference at which the chimpanzees display a preference for one of the two options. We found that the chimpanzees significantly preferred concentrations as low as 20 mM sucrose, 40 mM fructose, and 80 mM glucose, lactose, and maltose over tap water. When given a choice between all binary combinations of these five saccharides presented at equimolar concentrations of 100, 200, and 400 mM, respectively, the animals displayed significant preferences for individual saccharides in the following order: sucrose > fructose > glucose = maltose = lactose. The taste difference threshold for sucrose, expressed as Weber ratio (ΔI/I), was 0.3 and 0.4, respectively, at reference concentrations of 100 and 200 mM. The taste sensitivity of the chimpanzees to the five saccharides falls into the same range found in other primate species. Remarkably, their taste preference thresholds are similar, and with two saccharides even identical, to human taste detection thresholds. The pattern of relative taste preferences displayed by the chimpanzees was similar to that found in platyrrhine primates and to the pattern of relative sweetness as reported by humans. Taken together, the results of the present study are in line with the notion that taste sensitivity for food-associated carbohydrates may correlate positively with phylogenetic relatedness. Further, they support the notion that relative preferences for food-associated carbohydrates, but not taste difference thresholds, may correlate with dietary specialization in primates.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Comparative studies of taste perception allow us to gain insight into the mechanisms underlying the evolution of the sense of taste and the causes of between-species differences in taste performance. Primates are a particularly suitable taxon in this respect, as they comprise a large variety of dietary specializations (Fleagle 2013) and the composition of food is commonly thought to affect the taste perception of a given species (Dominy et al. 2001). Accordingly, the sense of taste has been studied with regard to its anatomy (e.g., Hofer et al. 1990; Emura 2017; Pastor et al. 2017), electrophysiology (e.g., Hellekant et al. 1996, 1997, 1998), and genetics (e.g., Go et al. 2005; Wooding et al. 2006; Li et al. 2011) in all major taxa of primates, including the hominids. Surprisingly little, in contrast, is known about taste perception in the Great Apes at the behavioral level. With regard to substances tasting sweet to humans, for example, taste preference thresholds in the chimpanzee have so far only been reported for fructose (Simmen and Charlot 2003; Remis 2006), and for the two sweet-tasting compounds of Stevia rebaudiana, stevioside and rebaudioside A (Nicklasson et al. 2018).
Chimpanzees (Pan troglodytes) include a considerable proportion of fruit into their diet, with values ranging from 55 to 75% of their annual intake, depending on study site, season of the year, and recording method (e.g., Head et al. 2011; McLennan 2013; Morgan and Sanz 2006; Watts et al. 2012; Wrangham et al. 1998). Accordingly, they are often referred to as primarily frugivorous and as ripe fruit specialists (Milton 1999; Pruetz 2006). Their food selection behavior suggests that chimpanzees may use the sweetness of fruits as a criterion for consumption (Dominy et al. 2016; Hladik and Simmen 1996).
Fruits contain a variety of soluble carbohydrates that taste sweet to humans (Kinghorn and Soejarto 1986). However, the disaccharide sucrose and its monosaccharide components fructose and glucose are usually quantitatively predominant in fruits and account for more than 90% of their total carbohydrate content (Nagy and Shaw 1980). The disaccharide lactose is the main carbohydrate in milk, the first diet in the life of mammals (Jensen 1995), and the milk of chimpanzees, for example, contains 7.4 g/100 ml of lactose (Hinde and Milligan 2011). The disaccharide maltose originates in considerable amounts from the enzymatic degradation of starch during mastication, and thus contributes to the taste sensation while feeding on starch-containing plants (Butterworth et al. 2011).
It was therefore the aim of the present study to assess the taste responsiveness of Western chimpanzees to these five food-associated carbohydrates. More specifically, we (1) determined taste preference thresholds for sucrose, fructose, glucose, lactose, and maltose in P. troglodytes verus, (2) assessed relative preferences of the chimpanzees for these five saccharides when presented at equimolar concentrations, and (3) determined taste preference difference thresholds for sucrose, that is, the smallest concentration difference for which the chimpanzees displayed a reliable difference in preference. To this end, we employed a two-bottle preference test of short duration (Richter and Campbell 1940). This test allows us to measure both absolute and relative preferences for taste substances and, at the same time, largely rules out the influence of postingestive factors on an individual’s ingestive behavior.
Methods
Animals
We assessed taste responsiveness in two adult female and one adult male Western chimpanzees (P. troglodytes verus). The animals were 27, 33, and 48 years old at the start of the study. They were housed, together with one other individual, at Borås Zoo, Sweden, in a 750 m3 indoor exhibit, with access to a 560 m2 outdoor island with natural vegetation. We performed the tests in a smaller room adjacent to the indoor exhibit, which held three compartments in which the animals were tested separately to avoid competition and distraction. All three animals were trained to voluntarily enter the test compartments and were completely accustomed to the procedure described below (Nicklasson et al. 2018). The animals were fed fresh fruit and vegetables (e.g., apples, bananas, grapes, melons, mangoes, figs, tomatoes, cucumber, carrots, broccoli, avocado, with seasonal variations) three times per day. Commercial primate chow pellets were served once per day, eggs were served weekly, and water was provided ad libitum. The amount of food offered daily to the animals was such that leftovers were still present on the floor the next morning. Thus, it was unlikely that ravenous appetite affected the animals’ ingestive behavior.
Ethical note
The experiments reported here comply with the American Society of Primatologists’ Principles for the Ethical Treatment of Primates, and also with current Swedish laws. This study was approved by Gothenburg’s Animal Care and Use Committee (Göteborgs djurförsöksetiska nämnd, protocol #75-2016).
Taste stimuli
We used the following five saccharides: sucrose (CAS# 57-50-1), fructose (CAS# 57-48-7), glucose (CAS# 50-99-7), maltose (CAS# 6363-53-7), and lactose (CAS# 63-42-3). All substances were obtained from Sigma-Aldrich (St. Louis, MO, USA) and were of the highest available purity (≥ 99.5%).
Procedure
We used a two-bottle preference test of short duration (Richter and Campbell 1940). The animals were allowed to drink for 1 min from a pair of simultaneously presented graduated cylinders of 700 ml with metal drinking spouts. We performed between four and six of such 1-min trials per day and animal, with usually two trials in the morning, around noon, and in the afternoon, respectively.
Determination of taste preference thresholds
To determine taste preference thresholds, the animals were given the choice between tap water and defined concentrations of a saccharide dissolved in tap water. With all five saccharides, testing started at a concentration of 200 mM and proceeded in the following steps (100, 50, 20, 10 mM, etc.) until an animal failed to show a significant preference. Subsequently, they were presented with intermediate concentrations (between the lowest concentration that was preferred and the first concentration that was not) in order to determine the preference threshold value more exactly. The order in which the five saccharides were tested was the same for all three animals: (1) sucrose, (2) fructose, (3) glucose, (4) maltose, and (5) lactose.
Assessment of relative taste preferences
To assess relative preferences, the animals were given the choice between two saccharide solutions presented at equimolar concentrations. All ten possible binary stimulus combinations (e.g., sucrose vs. fructose, maltose vs. lactose, etc.) were tested. To assess whether relative preferences were stable at different concentrations, three series of tests were performed at 100, 200, and 400 mM, respectively.
Determination of taste preference difference thresholds
To determine taste preference difference thresholds for sucrose, the animals were given the choice between a reference concentration and lower concentrations of the same substance until the animals failed to show a significant preference for one of the two alternatives. To assess whether taste preference difference thresholds were stable at different concentrations, two series of tests were performed, using reference concentrations of 100 mM and 200 mM sucrose, respectively. To this end, the 100 mM sucrose reference concentration was tested against sucrose solutions of 20, 50, 60, 70, 80, and 90 mM, respectively, and the 200 mM sucrose reference concentration was tested against sucrose solutions of 20, 50, 90, 100, 120, 130, 140, and 150 mM, respectively.
In all three experiments, we presented each pair of stimuli ten times per individual animal, and the position of the stimuli was pseudo-randomized in order to counterbalance possible position preferences. Care was taken that an animal sampled both stimuli at least once during each trial. To maintain the animals’ motivation and willingness to cooperate, testing of the different stimulus combinations within an experiment did not follow a strict order but was pseudo-randomized. This was true both within a given session (morning, noon, or afternoon) and between sessions.
Data analysis
For each animal, we recorded the amount of liquid consumed from each bottle, summed it for the ten trials with a given stimulus combination, converted it to percentages (relative to the total amount of liquid consumed from both bottles), and took 66.7% (i.e., 2/3 of the total amount of liquid consumed) as the criterion of preference. We chose this rather conservative criterion for reasons of comparability of data as the same criterion had been used in previous studies on sweet-taste responsiveness with other primate species (Laska 1996, 1997, 2000; Laska et al. 1996, 1998, 1999a, b, 2001; Nicklasson et al. 2018; Wielbass et al. 2015), and in order to avoid misinterpretation due to a too liberal criterion. Additionally, we performed binomial tests, and regarded an animal as significantly preferring one of the two stimuli if it reached the criterion of 66.7% and consumed more from the bottle containing the preferred stimulus in at least eight out of ten trials (binomial test, P < 0.05).
Thus, we defined taste preference threshold as the lowest concentration at which the animals met both criteria mentioned above. Preliminary analyses of the data indicated that there were no systematic differences in choice behavior and liquid consumption between the first and the second presentation of a session, or between the morning, the noon, and the afternoon session, respectively. Intraindividual variability of the amount of liquid consumed across the ten trials with a given stimulus combination was low and averaged less than 20%. Thus, a theoretically possible bias in the overall preference score due to excessive drinking in aberrant trials did not occur. All data are reported as mean values ± SD.
Taste preference difference threshold values were expressed as Weber ratios (ΔI/I). They are based on the Weber–Fechner law of psychophysics and commonly used to quantify the just noticeable difference (JND) between different concentrations of the same stimulus (Fischer et al. 1965).
Results
Taste preference thresholds
Taste preference thresholds of the three Western chimpanzees were found to be 20 mM for sucrose, 40 mM for fructose, and 80 mM for glucose, maltose and lactose, respectively (Fig. 1). All animals failed to show a significant preference for the lowest concentrations presented, suggesting that the preference for higher concentrations was indeed based on the chemical nature of the stimuli. In most cases, interindividual variability was low for both sub- and suprathreshold concentrations tested and with only two exceptions all three animals either reached the criterion of preference (> 66.7% of total consumption, plus binomial test, P < 0.05) with a given stimulus combination or all three animals failed to do so.

Mean taste responses (± SD) of three Western chimpanzees to aqueous solutions of sucrose, fructose, glucose, maltose, and lactose tested against tap water. Each data point represents the mean value of ten trials of 1 min per animal. The dotted horizontal lines at 66.7% and at 50% indicate the criterion of preference and the chance level, respectively
Relative taste preferences
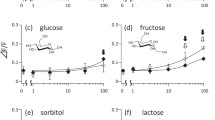
When given the choice between two aqueous saccharide solutions presented at equimolar concentrations of 100 mM, the chimpanzees significantly preferred sucrose over all other saccharides. Further, they showed a non-significant trend for preferring fructose over the other three saccharides (Fig. 2, upper panel). At 200 mM, the chimpanzees significantly preferred sucrose and fructose over glucose, lactose, and maltose (Fig. 2, middle panel). At 400 mM, the chimpanzees only displayed a significant preference for sucrose over lactose. However, they also showed a non-significant trend for preferring sucrose over glucose and maltose, and for preferring fructose over glucose, maltose, and lactose (Fig. 2, lower panel). Interindividual variability was low and with only few exceptions all three animals either reached the criterion of preference (> 66.7% of total consumption, plus binomial test, P < 0.05) with a given stimulus combination or all three animals failed to do so.

Relative taste preferences of three Western chimpanzees when given the choice between two aqueous solutions presented at equimolar concentrations of 100, 200, and 400 mM, respectively. Each bar represents the mean preference (± SD) from ten trials of 1 min per individual for the saccharide on the left side relative to the saccharide on the right. A white circle indicates a significant preference according to the criterion > 66.7% of total amount of liquid consumed. Asterisks indicate a significant preference according to a two-tailed binomial test, with P < 0.05 (one asterisk) and P < 0.01 (two asterisks). The fat horizontal lines at 66.7% and at 50% indicate the criterion of preference and the chance level, respectively
Taste preference difference thresholds
When given the choice between different concentrations of sucrose, the chimpanzees significantly preferred the reference concentration of 100 mM over concentrations up to 70 mM, and they significantly preferred the reference concentration of 200 mM over concentrations up to 120 mM (Fig. 3). Accordingly, the taste preference difference threshold for sucrose, expressed as Weber ratio (ΔI/I), was 0.3 at a reference concentration of 100 mM, and 0.4 at a reference concentration of 200 mM. Here, too, interindividual variability was low and with only few exceptions all three animals either reached the criterion of preference (> 66.7% of total consumption, plus binomial test, P < 0.05) with a given stimulus combination or all three animals failed to do so.

Taste preference difference thresholds at two different reference concentrations of sucrose (100 and 200 mM) in three Western chimpanzees. Each data point represents the mean value (± SD) of ten trials of 1 min per animal. The dotted horizontal lines at 66.7% and at 50% indicate the criterion of preference and the chance level, respectively. Please note that, for reasons of readability of the graph, the x-axis displays concentrations on a linear rather than on a logarithmic scale
Discussion
The results of the present study show that Western chimpanzees have a well-developed sensitivity for food-associated saccharides. Further, they show that the five carbohydrates differ in their attractiveness and thus in their relative sweetness and that P. troglodytes verus has a well-developed ability to distinguish between different concentrations of sucrose.
Taste preference thresholds
The concentrations of sucrose, fructose, and glucose found in ripe tropical fruits are markedly higher than the taste preference thresholds of the chimpanzees found in the present study (Nagy and Shaw 1980). Thus, these saccharides should contribute substantially to the taste sensation in chimpanzees while feeding on fruits. This, in turn, supports the notion that chimpanzees may indeed use the sweetness of fruits as a criterion for consumption (Dominy et al. 2016; Hladik and Simmen 1996). Similarly, the lactose content of chimpanzee milk of 7.4 g/100 ml (Hinde and Milligan 2011), corresponding to a concentration of 217 mM, is markedly higher compared to their taste preference threshold of 80 mM found here. This suggests that infant chimpanzees experience sweet taste from their very first meal of breast milk on. Some authors even hypothesize this early exposure to the sweet taste of lactose in breast milk to be the reason for the “sweet tooth”, that is, the predilection for sweet taste in primates (Ramirez 1990; Beauchamp and Mennella 2009). Although it is not possible to reliably determine the concentration of maltose that is produced via the enzymatic degradation of starch during mastication, chimpanzees are known to produce salivary α-amylase (Perry et al. 2007) and are thus, unlike some other primate species, able to enzymatically convert starch to maltose. This, in turn, makes it likely that maltose also contributes to sweet-taste perception in chimpanzees while feeding on starch-containing fruits.
The taste preference threshold value of 40 mM for fructose determined in the present study compares favorably with the results of the only two previous studies which assessed sweet-taste sensitivity in chimpanzees with carbohydrates so far: Simmen and Charlot (2003) reported a taste preference threshold value of 40 mM for fructose in two chimpanzees, and Remis (2006) a value of 50 mM in four chimpanzees. This suggests that our results are robust, despite the limited number of animals used.
The taste sensitivity of P. troglodytes verus for the five saccharides tested here falls into the range of threshold values reported in other species of nonhuman primates that have been tested previously with the same or a similar method (two-bottle preference test of short duration) (Table 1). The pattern of sweet-taste sensitivity found in the chimpanzees, with the threshold value for sucrose being lowest among the five saccharides tested, and the threshold value for fructose being lower than that for glucose, is also consistent with the majority of nonhuman primates tested so far.
It is interesting to note that the taste preference thresholds reported here with chimpanzees are also similar, and in two cases (fructose and glucose) even identical to the taste detection threshold values obtained with human subjects (Table 1). This is not trivial, as the sophisticated signal detection methods employed in taste tests with human subjects are known to yield lower threshold values compared to the simple two-bottle preference test used with nonhuman primates, which provides only a conservative approximation of a species’ taste sensitivity (Spector 2003). The finding of similar sweet-taste thresholds in chimpanzees and humans as well as the similarity in the pattern of sweet-taste sensitivity in P. troglodytes verus and Homo sapiens across the five saccharides supports the notion that phylogenetic relatedness may correlate positively with taste sensitivity for food-associated carbohydrates (Glaser et al. 1995; Nofre et al. 1996).
Relative taste preferences
The food-associated saccharides used here are known to differ in their stimulating efficiency, that is, some of them (e.g., sucrose) are perceived by humans as sweeter than others (e.g., lactose) when presented at equimolar concentrations (Pfaffmann et al. 1971). Differences in ligand affinity to the mammalian sweet-taste receptor have been identified as the underlying molecular mechanism of this phenomenon (Chandrashekar et al. 2006). It is commonly agreed that differences in the attractiveness of sweet-tasting substances reflect differences in stimulating efficiency. This notion is supported by electrophysiological findings in nonhuman primates including chimpanzees (Scott et al. 1991; Plata-Salaman et al. 1993; Hellekant et al. 1997). Therefore, the relative taste preferences found here can be considered as an approximation of the relative sweetness as perceived by the chimpanzees. The pattern of relative taste preference found in the present study in Western chimpanzees (sucrose > fructose > glucose = maltose = lactose) is identical or at least largely similar to that found in previous studies with squirrel monkeys, spider monkeys, and black-and-white ruffed lemurs, and also similar to the pattern of relative sweetness as reported by human subjects (Table 2). All five species clearly prefer sucrose over the other saccharides tested when presented at equimolar concentrations. Interestingly, pigtail macaques differ markedly in their relative taste preferences for food-associated saccharides. Unlike the other primates tested so far, but similar to rats, Macaca nemestrina clearly prefers maltose over sucrose. It has been hypothesized that rodents such as rats, and possibly also macaques, may have an additional taste receptor for starch-derived polysaccharides that also responds to maltose, the disaccharide building-block of starch, but not to sucrose (Bachmanov and Beauchamp 2007). The diet of rats and pigtail macaques is known to contain a high proportion of starch, either in the form of seeds or in the form of starch-containing underground storage organs such as roots, tubers, and bulbs (Caldecott 1986). The diet of other primates such as chimpanzees, in contrast, usually does not contain significant quantities of starch. The present results are in line with this hypothesis. Thus, the present findings lend further support to the notion that relative preferences for food-associated carbohydrates may correlate with dietary specialization in primates.
Taste preference difference thresholds
For species which at least partly meet their energy requirements by using easily metabolizable carbohydrates, it should be particularly important to be able to evaluate the sugar content of potential food items. In addition to a sufficiently high sensitivity allowing for the detection of sugars in food, such species should have a well-developed ability to distinguish between different sugar concentrations in order to optimize their energy yield as the carbohydrate content and thus the nutritive value of fruits may change markedly during maturation (Kinghorn and Soejarto 1986). The food selection behavior of chimpanzees supports this notion, as they have been observed to regularly base their decisions about consumption or rejection of a fruit on the taste sensation they experience during the first bite (Hladik and Simmen 1996). The taste preference difference thresholds of the chimpanzees for sucrose found in the present study are in the same range as those reported in squirrel monkeys, but higher than those in spider monkeys and olive baboons (Table 3). This finding is somewhat unexpected, as some studies suggest that the ability to distinguish between different concentrations of sweet-tasting food constituents should correlate positively with the degree of frugivory (Laska et al. 1999a, b). Spider monkeys, for example, are clearly more frugivorous than squirrel monkeys and Ateles geoffroyi also displays lower Weber ratios, that is, a higher ability to discriminate between sucrose concentrations than Saimiri sciureus. However, such a correlation might only hold true for platyrrhine primates, but not for catarrhine primates, as these two taxa have also been found to differ with regard to other aspects of sweet-taste perception, e.g., the ability to detect certain types of proteins that taste sweet to humans (Glaser et al. 1978, 1992, 1996).
The taste difference threshold values for sucrose reported in human studies show considerable variation and a general tendency for lower values compared to the chimpanzees studied here (see Table 3). Whereas the former is probably due to the fact that the human studies employed different methods (e.g., triangle or paired-comparison tests) and threshold criteria, the latter can probably be ascribed to the fact that signal detection methods yield lower threshold values compared to simple two-bottle preference tests (Spector 2003). In this context, it should be mentioned that all animal data reported in Table 3 are based on the same method, that is, two-bottle preference tests of short duration. The findings of the present study do not support the notion that dietary specialization would systematically affect taste preference difference thresholds.
Taken together, the results of the present study are in line with the notion that taste sensitivity for food-associated carbohydrates may correlate positively with phylogenetic relatedness. Further, they support the notion that relative preferences for food-associated carbohydrates, but not taste preference difference thresholds, may correlate with dietary specialization in primates.
References
Bachmanov AA, Beauchamp GK (2007) Taste receptor genes. Annu Rev Nutr 27:389–414
Beauchamp GK, Mennella JA (2009) Early flavor learning and its impact on later feeding behavior. J Pediatr Gastroenterol Nutr 48:S25–S30
Berg HW, Filipello F, Hinreiner E, Webb AD (1955) Evaluation of thresholds and minimum difference concentrations for various constituents of wines. I. Water solutions of pure substances. Food Technol 9:23–26
Brosvic GM, Slotnick BM (1986) Absolute and intensity-difference taste thresholds in the rat: evaluation of an automated multichannel gustometer. Physiol Behav 38:711–717
Butterworth PJ, Warren FJ, Ellis PR (2011) Human α-amylase and starch digestion: an interesting marriage. Starch 63:305–405
Caldecott JO (1986) An ecological and behavioural study of the pig-tailed macaque. Contributions to Primatology, vol 21. Karger, Basel
Chandrashekar J, Hoon MA, Ryba NJP, Zuker CS (2006) The receptors and cells for mammalian taste. Nature 444:288–294
Dominy NJ, Lucas PW, Osorio D, Yamashita N (2001) The sensory ecology of primate food perception. Evol Anthropol 10:171–186
Dominy NJ, Yeakel JD, Bhat U, Ramsden L, Wrangham RW, Lucas PW (2016) How chimpanzees integrate sensory information to select figs. Interface Focus 6:20160001
Emura S (2017) Morphology of the lingual papillae in the chimpanzee. Okajimas Folia Anat Jpn 94:115–118
Fischer R, Griffin F, Archer RC, Zinsmeister SC, Jastram PS (1965) Weber ratio in gustatory chemoreception: an indicator of systemic (drug) reactivity. Nature 207:1049–1053
Fleagle JG (2013) Primate adaptation and evolution, 3rd edn. Academic Press, San Diego
Geldard FA (1972) The sense of taste. In: Geldard FA (ed) The Human Senses. Wiley, New York, pp 480–524
Gilmore MM, Murphy C (1989) Aging is associated with increased Weber ratios for caffeine, but not for sucrose. Percept Psychophys 46:555–559
Glaser D (1986) Geschmacksforschung bei Primaten. Vj Naturforsch Ges Zürich 131:92–110
Glaser D, Hellekant G, Brouwer JN, van der Wel H (1978) The taste responses in primates to the proteins thaumatin and monellin and their phylogenetic implications. Folia Primatol 29:56–63
Glaser D, van der Wel H, Brouwer JN, Dubois GE, Hellekant G (1992) Gustatory responses in primates to the sweetener aspartame and their phylogenetic implications. Chem Senses 17:325–335
Glaser D, Tinti JM, Nofre C (1995) Evolution of the sweetness receptor in primates. I. Why does alitame taste sweet in all prosimians and simians, and aspartame only in Old World simians? Chem Senses 20:573–584
Glaser D, Tinti JM, Nofre C (1996) Gustatory responses of non-human primates to dipeptide derivatives or analogues, sweet in man. Food Chem 56:313–321
Go Y, Satta Y, Takenaka O, Takahata N (2005) Lineage-specific loss of function of bitter taste receptor genes in humans and nonhuman primates. Genetics 170:313–326
Head JS, Boesch C, Makaga L, Robbins MM (2011) Sympatric chimpanzees (Pan troglodytes troglodytes) and gorillas (Gorilla gorilla gorilla) in Loango National Park, Gabon: dietary composition, seasonality, and intersite comparisons. Int J Primatol 32:755–775
Hellekant G, Ninomiya Y, Dubois GE, Danilova V, Roberts TW (1996) Taste in chimpanzees. III: labeled-line coding in sweet taste. Physiol Behav 65:191–200
Hellekant G, Ninomiya Y, Danilova V (1997) Taste in chimpanzees. II: single chorda tympani fibers. Physiol Behav 61:829–841
Hellekant G, Ninomiya Y, Danilova V (1998) Taste in chimpanzees. I: the summated response to sweeteners and the effect of gymnemic acid. Physiol Behav 60:469–479
Hinde K, Milligan LA (2011) Primate milk: proximate mechanisms and ultimate perspectives. Evol Anthropol 20:9–23
Hladik CM, Simmen B (1996) Taste perception and feeding behavior in nonhuman primates and human populations. Evol Anthropol 5:58–71
Hofer HO, Meinel W, Sauer E (1990) Comparative anatomical studies of the tongue of Pan troglodytes (Blumenbach, 1799) and other primates. I. The chimpanzee tongue. Gegenbaurs Morph Jahrb 136:455–492
Jensen RG (1995) Handbook of milk composition. Academic Press, New York
Kinghorn AD, Soejarto DD (1986) Sweetening agents of plant origin. Crit Rev Plant Sci 4:79–120
Laing DG, Prescott J, Bell GA, Gillmore R, James C, Best DJ, Allen S, Yoshida M, Yamazaki K (1993) A cross-cultural study of taste discrimination with Australians and Japanese. Chem Senses 18:161–168
Laska M (1994) Taste difference thresholds for sucrose in squirrel monkeys (Saimiri sciureus). Folia Primatol 63:144–148
Laska M (1996) Taste preference thresholds for food-associated sugars in the squirrel monkey (Saimiri sciureus). Primates 37:91–95
Laska M (1997) Taste preferences for five food-associated sugars in the squirrel monkey (Saimiri sciureus). J Chem Ecol 23:659–672
Laska M (2000) Gustatory responsiveness to food-associated sugars and acids in pigtail macaques Macaca nemestrina. Physiol Behav 70:495–504
Laska M, Carrera Sanchez E, Rodriguez Rivera JA, Rodriguez Luna E (1996) Gustatory thresholds for food-associated sugars in the spider monkey (Ateles geoffroyi). Am J Primatol 39:189–193
Laska M, Carrera Sanchez E, Rodriguez Luna E (1998) Relative taste preferences for food-associated sugars in the spider monkey (Ateles geoffroyi). Primates 39:91–96
Laska M, Scheuber HP, Carrera Sanchez E, Rodriguez Luna E (1999a) Taste difference thresholds for sucrose in two species of nonhuman primates. Am J Primatol 48:153–160
Laska M, Schüll E, Scheuber HP (1999b) Taste preference thresholds for food-associated sugars in baboons, Papio hamadryas anubis. Int J Primatol 20:25–34
Laska M, Kohlmann S, Scheuber HP, Hernandez Salazar LT, Rodriguez Luna E (2001) Gustatory responsiveness to polycose in four species of nonhuman primates. J Chem Ecol 27:1997–2011
Li X, Bachmanov AA, Maehashi K, Li W, Lim R, Brand JG, Beauchamp GK, Reed DR, Thai C, Floriano WB (2011) Sweet taste receptor gene variation and aspartame taste in primates and other species. Chem Senses 36:453–475
Lundgren B, Pangborn RM, Barylko-Pikielna N, Daget N (1976) Difference taste thresholds for sucrose in water and in orange juice: an interlaboratory study. Chem Senses 2:157–176
McBride RL (1983) A JND-scale/category-scale convergence in taste. Percept Psychophys 34:77–83
McLennan MR (2013) Diet and feeding ecology of chimpanzees (Pan troglodytes) in Bulindi, Uganda: foraging strategies at the forest-farm interface. Int J Primatol 34:585–614
Milton K (1999) Nutritional characteristics of wild primate foods: do the diets of our closest living relatives have lessons for us? Nutrition 15:488–498
Morgan D, Sanz C (2006) Chimpanzee feeding ecology and comparisons with sympatric gorillas in the Goualougo Triangle, Republic of Congo. In: Homann G, Robbins MM, Boesch C (eds) Feeding ecology in apes and other primates. Cambridge University Press, Cambridge, pp 97–122
Nagy S, Shaw PE (1980) Tropical and subtropical fruits: composition, nutritive values, properties and uses. Avi Pub Co, Westport
Nicklasson S, Sjöström D, Amundin M, Roth D, Hernandez Salazar LT, Laska M (2018) Taste responsiveness to two steviol glycosides in three species of nonhuman primates. Curr Zool 64:63–68
Nishi E, Tsutsui K, Imai H (2016) High maltose sensitivity of sweet taste receptors in the Japanese macaque (Macaca fuscata). Sci Rep 6:39352
Nofre C, Tinti JM, Glaser D (1996) Evolution of the sweetness receptor. II. Gustatory responses of non-human primates to nine compounds known to be sweet in man. Chem Senses 21:747–762
Pastor JF, Barbosa M, De Paz FJ, San José I, Levanti M, Potau JM, Vega JA, Cabo R (2017) Ultrastructure of lingual papillae in common chimpanzees (Pan troglodytes) foetus, newborn and adult specimens. Anat Histol Embryol 46:431–438
Perry GH, Dominy NJ, Claw KG, Lee AS, Fiegler H, Redon R, Werner J, Villanea FA, Mountain JI, Misra R, Carter NP, Lee C, Stone AC (2007) Diet and the evolution of human amylase gene copy number variation. Nat Genet 39:1256–1260
Pfaffmann C, Bartoshuk LM, McBurney DH (1971) Taste psychophysics. In: Autrum H, Beidler L, Acree T (eds) Handbook of sensory physiology. Part 2, vol 4. Springer, Berlin, pp 75–101
Plata-Salaman CR, Scott TR, Smith-Swintosky VL (1993) Gustatory neural coding in the monkey cortex: the quality of sweetness. J Neurophysiol 69:482–493
Pruetz JD (2006) Feeding ecology of savannah chimpanzees at Fongoli, Senegal. In: Homann G, Robbins MM, Boesch C (eds) Feeding ecology in apes and other primates. Cambridge University Press, Cambridge, pp 161–182
Ramirez I (1990) Why do sugars taste good? Neurosci Biobehav Rev 14:125–134
Remis MJ (2006) The role of taste in food selection by African apes: implications for niche separation and overlap in tropical forests. Primates 47:56–64
Richter CP, Campbell KH (1940) Taste thresholds and taste preferences of rats for five common sugars. J Nutr 20:31–46
Schutz HG, Pilgrim FJ (1957) Differential sensitivity in gustation. J Exp Psychol 54:41–48
Sclafani A, Mann S (1987) Carbohydrate taste preference in rats: glucose, sucrose, maltose, fructose and polycose compared. Physiol Behav 40:563–568
Scott TR, Giza BK (1987) A measure of taste intensity discrimination in the rat through conditioned taste aversions. Physiol Behav 41:315–320
Scott TR, Plata-Salaman CR, Smith VL, Giza BK (1991) Gustatory neural coding in the monkey cortex: stimulus intensity. J Neurophysiol 65:76–86
Simmen B (1994) Taste discrimination and diet differentiation among New World primates. In: Chivers DJ, Langer P (eds) The digestive system in mammals: food, form, and function. Cambridge University Press, Cambridge, pp 150–165
Simmen B, Charlot S (2003) A comparison of taste thresholds for sweet and astringent-tasting compounds in great apes. C R Biol 326:449–455
Simmen B, Hladik CM (1998) Sweet and bitter taste discrimination in primates: scaling effects across species. Folia Primatol 69:129–138
Simmen B, Josseaume B, Atramentowicz M (1999) Frugivory and taste responses to fructose and tannic acid in a prosimian primate and a didelphid marsupial. J Chem Ecol 25:331–346
Spector AC (2003) Psychophysical evaluation of taste function in nonhuman mammals. In: Doty RL (ed) Handbook of olfaction and gustation, 2nd edn. Marcel Dekker, New York, pp 861–879
Sunderland G, Sclafani A (1988) Taste preferences of squirrel monkeys and bonnet macaques for polycose, maltose, and sucrose. Physiol Behav 43:685–690
van Gemert LJ (2011) Flavour thresholds. Compilations of flavour threshold values in water and other media, 2nd edn. Oliemans Punter & Partner, Utrecht
Wagner MW, Green KF, Manley MB (1965) Paired comparison method for measurement of sugar preference in squirrel monkeys. Science 148:1473–1474
Watts DP, Potts KB, Lwanga JS, Mitani JC (2012) Diet of chimpanzees (Pan troglodytes schweinfurthii) at Ngogo, Kibale National Park, Uganda, 1. Diet composition and diversity. Am J Primatol 74:114–129
Wielbass A, Amundin M, Laska M (2015) Gustatory responsiveness of black-and-white ruffed lemurs (Varecia variegata variegata) to food-associated sugars. Int J Primatol 36:460–472
Wooding S, Bufe B, Grassi C, Howard MT, Stone AC, Vazquez M, Dunn DM, Meyerhof W, Weiss RB, Bamshad MJ (2006) Independent evolution of bitter-taste sensitivity in humans and chimpanzees. Nature 440:930–934
Wrangham RW, Conklin-Brittain NL, Hunt KD (1998) Dietary response of chimpanzees and cercopithecines to seasonal variation in fruit abundance. I. Antifeedants. Int J Primatol 19:949–970
Acknowledgements
The primate caretakers at Borås Zoo are gratefully acknowledged for their help and support with the animals.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Norlén, E., Sjöström, D., Hjelm, M. et al. Taste responsiveness of Western chimpanzees (Pan troglodytes verus) to five food-associated saccharides. Primates 60, 29–39 (2019). https://doi.org/10.1007/s10329-018-0697-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-018-0697-0