
Abstract
The aim of this study was to quantify the effect of bottom and suspended mussel cultures, cultured in different physical environments, on the sedimentary environmental conditions and thereby the biodiversity structure of the associated macrofaunal community. We compared two bottom cultures (Limfjorden: microtidal, wind-driven; Oosterschelde: macrotidal) and one suspended culture (Ria de Vigo in an upwelling coastal region). The sedimentary environmental conditions (mud fraction, POC, PON, phosphorus content, chl a breakdown products) were significantly elevated underneath and surrounding bottom and suspended cultures compared to culture-free sediments that were nearby and hydrodynamically similar. The relative change in environmental conditions was more pronounced in the Oosterschelde compared to Limfjorden, most likely due to differences in hydrodynamic forcing and characteristics of the mussel bed. The effect of the suspended cultures in Ria de Vigo on the surrounding sediments was influenced by local topographic and hydrodynamic conditions. The impact of mussels on the benthic community due to biodeposition was clearly seen in the community structure. The species composition changed from species which are typically present in sandy environments to more small opportunistic species, which are typically present in organically enriched sediments. The impact of bottom cultures on the benthic community due to changes in the habitat under the presence of mussels was positive, especially in the Oosterschelde where an increase in the number of epibenthic species was seen. The influence of bottom cultures on the sedimentary environment and on the macrobenthic community seems to be very local. Within the mussel site in Limfjorden, differences were detected between sites where none or almost no mussels were present with sites where mussels were very abundant.
Similar content being viewed by others


Introduction
Habitat created or modified by the physical architecture of large or spatially dominant species plays an important role in structuring communities in a variety of terrestrial, aquatic, and marine habitats. These organisms, termed foundation species by Bruno and Bertness (2001), create (facilitate) habitat for many associated species, and have been recognized as important drivers of biodiversity (Bruno et al. 2003). Jones et al. (1994, 1997) introduced the concept of ecosystem engineers for “organisms that change biotic or abiotic materials, thereby controlling availability of resources to other organisms”. Their impact and importance are likely to shift across environmental stress gradients (Crain and Bertness 2006). In extreme physical environments, ecosystem engineers will ameliorate physical stress. In physically more benign environments, ecosystem engineers will support ecosystem processes by providing competitor- or predator-free space.
A conspicuous element in shallow coastal and estuarine ecosystems is the presence and often dominance of macrobenthic suspension-feeders such as mussels and oysters. Dense populations of these bivalve filter-feeders, including both wild and cultured populations, can modify and maintain very specific habitats. Mussels like Mytilus edulis naturally occur on rocky shores and on soft sediments where they form large beds. They are attached to the substratum and to each other by byssal threads, creating an irregular surface topography. This complex epibenthic matrix provides a biogenic habitat for a large number of plant and animal species, including algae, worms, snails, crustaceans (Tsuchiya and Nishihira 1985; Dittmann 1990; Seed and Suchanek 1992; Reise 2002; Thiel and Ullrich 2002). Mussel beds control the benthic environment directly by providing habitat and indirectly by providing shelter from predation, trapping sediment and altering water flow (Gutiérrez et al. 2003). In addition, they change benthic–pelagic exchange, as they filter significant amounts of seston particles from the water column, diverting primary production and energy flow from the pelagic to the benthic (Cloern 1982). This biodeposition results in a further modification of the sedimentary habitat (Graf and Rosenberg 1997; Norkko et al. 2001). Because of the intensity of this benthic–pelagic coupling, they play an essential role in the functioning of coastal ecosystems (Alpine and Cloern 1992; Dame 1996).
Mussel beds naturally occur in many subtidal and intertidal areas around the world, but mussels are also extensively cultured. Mussel aquaculture is done by means of bottom cultures (by seeding intertidal or subtidal beds), but also by suspended cultures (using rafts or longlines), and cultures on bouchots (Smaal 2002). Being suspended into the water column, mussels from rafts or longlines do not provide direct habitat to the sedimentary environment below them, but their feeding activity results in the packaging of fine suspended material into large feces and pseudofeces that rapidly settle to the seabed. This biodeposition leads to enrichment of the sediment with organic material (Kautsky and Evans 1987), an increase in microbial activity (Kaspar et al. 1985) resulting in reduced interstitial oxygen concentrations (Tenore et al. 1982), increased sulfate reduction (Dahlbäck and Gunnarsson 1981) and increased denitrification (Kaspar et al. 1985; Christensen et al. 2003). A variety of levels of effect of shellfish farming activities on the benthic environment has been reported (Crawford et al. 2003; Newell 2004).
The EU project MaBenE aimed to develop ecological models for ecosystems dominated by benthic filter feeders, in order to optimize shellfish production in terms of yield as well as nature conservation. This requires thorough knowledge of the interplay between the physical regime and the biota in these systems. The aim of this study was to examine if bottom mussel beds or suspended cultures, maintained in different physical environments, change the bottom environmental conditions and thereby the biodiversity structure of the associated benthic macrofaunal community in a systematic and (across systems) predictable way. Therefore, we sampled very different systems that together span the range of conditions under which mussel culture may be found. Two bottom cultures (Limfjorden: microtidal, wind-driven system; Oosterschelde: macrotidal system) and one suspended culture (Ria de Vigo, an upwelling coastal region) were compared. It was hypothesized that the macrobenthic community would be altered due to the presence of mussels on the sediment surface, which creates a specific habitat that attracts other organisms, mostly epibenthic species and a variety of mobile fauna (e.g. crustaceans, echinoderms). As a result of the change in habitat structure an increase in species richness and diversity was expected. On the other hand, changes in sedimentary conditions, i.e. an increased organic flux to the sediment mainly caused by enhanced excretion of (pseudo-) feces, was expected to increase density and decrease biomass and number of endobenthic species, in accordance with general patterns of benthic response to organic enrichment (Pearson and Rosenberg 1978). This effect was expected to be stronger in a low-flow environment with direct local deposition (Limfjorden) than in a high-flow environment (Oosterschelde) where most biodeposition may be swept away with the currents. Suspended cultures, at least when they do not drop substantial numbers of mussels to the sediment (but see Grant et al. 1995) decouple the effect of habitat formation from the enrichment effect. In the nearby surroundings of suspended cultures a decrease in biotic parameter values (diversity, density and biomass) was expected, because biodeposition and therefore organic flux to the sediment was expected to be very intense.
Material and methods
Study areas
The areas selected in the MaBenE project represent some of the most important types of bottom and suspended cultures in Europe.
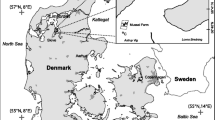
Limfjorden (1,575 km2, 7.1 km3), in the north of Denmark, is an open sound in connection with the North Sea in the west and Kattegat in the east (Fig. 1). It is a shallow system (mean depth 4.5 m) with a salinity that ranges from 22 to 32. There is a constant intrusion of high saline water from the North Sea and an input from the Kattegat of low saline water. The freshwater input from the surrounding area measures 2.7 km³ per year. Water temperatures in Limfjorden average around 2–3°C in winter, and 15–17°C in summer. Limfjorden is a micro-tidal system with a tidal amplitude of 0.1–0.2 m; the main physical driving forces are wind and solar radiation (Wiles et al. 2006). The center of Limfjorden opens into a large shallow basin, Løgstør Bredning (depth 5.8 m), where the field sites of the present study were located (Fig. 1). Limfjorden is a eutrophic water body affected by nutrient input from the surrounding watershed. This results in high primary production rates of up to 1,000 mg C m−2 per day in summer (Dolmer and Frandsen 2002) which supports a high level of biomass of benthic suspension-feeders. Mussel fishery in Limfjorden is based primarily on wild populations of M. edulis. Mussels are gathered by dredging; this is done once in approx. 2.5 years, because it takes that time for the minimum legal size of 4.5 cm to be reached (Dolmer et al. 1999). Natural recruitment occurs in existing mussel beds and is sufficient to maintain exploitation of the wild beds.

Map of Limfjorden (Denmark) showing the position of the mussel site (filled circle) and the control site (open circle)
The Oosterschelde is a macrotidal system with an average depth of 9 m, a tidal range of 3.25 m, and a surface area of 350 km2, of which 30% is tidal flats (Fig. 2). Due to the construction of a storm-surge barrier and two compartment dams (finalized in 1986), the estuary has changed into a sea inlet, and is nowadays characterized by a low freshwater river discharge (10 m3 s−1), high salinity (>30), relatively high water transparency (>2 m on average), long water residence time (10–50 days in the western part, 150 to >200 days in the eastern part) and low inorganic nutrient concentrations. Current velocities in the Oosterschelde can reach up to 1 m/s in the main gullies. The Oosterschelde estuary has an extensive bottom culture of mussels on cultivation plots. Mussel spat and half-grown mussels (M. edulis) are imported from the Wadden Sea and further cultivated on lease sites. Culture plots are mostly situated on the banks of tidal channels, from the intertidal range to 10–15 m below low tide level. The cultivation time of mussels is 14–33 months.

Map of the Oosterschelde (The Netherlands) showing the different mussel culture plots (left). The right figure shows a digital terrain model of the field site (measured by a multi-beam survey by GKSS). The depths are given in m below normal chart datum (RD/NAP). The two upper panels show the bed profiles across the observational sites. The profile origin is in the north
Ria de Vigo is one of the four Rias Baixas on the NW of the Iberian Peninsula (Fig. 3). Here the northern limit of the NW Africa upwelling system occurs; winds interact with the coastal topography to generate upwelling–downwelling dynamics on the continental shelf. The upwelling season occurs on average from March to September, when northerly winds prevail, but strength and frequency of upwelling events during this season vary strongly from year to year (Figueiras et al. 2002). Production of phytoplankton depends on the upwelling of nutrients with the cold deep seawater, which is advected into the bay. The high primary production, based on these nutrients, allows intensive suspended mussel cultures (Mytilus galloprovincialis) in the Rias. The Rias support a high density of mussel floating rafts made of crossed wooden laths (Figueiras et al. 2002). Rafts are 500 m2 (25 × 20 m) with on average 500 hanging ropes 12 m long each.

Map of Ria de Vigo (Spain) showing the different mussel polygons (black polygons, left). Right detailed map of the outer polygons showing the transects sampled for the characterization of the benthic environment. Each transect consisted of six stations (see text for more information)
Field sites and benthic sampling
To investigate the benthic environment in Limfjorden, Oosterschelde and Ria de Vigo samples were collected on 1 June 2003, 28 April 2004 and 28 July 2004, respectively.
In Limfjorden, sampling was done in Løgstør Bredning (Fig. 1). Two sites were identified: one area over mussel beds and the other, which was 950 m to the southeast, served as control site without mussels. Water depth at the mussel site was 7 m, and at the control site 5 m. Tidal currents at both sites are <0.1 m s−1. Hydrographic conditions, such as current speed, wave impact on the bottom, eddy viscosity and stratification conditions were very similar between the sites (Wiles et al. 2006; Riethmüller, pers. comm.). Samples were taken randomly within an area of 50 × 50 m in the mussel site and the control site. At both sites 25 samples were taken with a box-corer. Because of the low coverage, box cores taken in the mussel site sometimes did not contain any mussels. This allowed us to look also to small spatial scale differences, by dividing the mussel site further into samples with mussels (m-site, n = 16) and samples with no or 1 individual mussel (nm-site, n = 9).
In the Oosterschelde sampling was done on the eastern slope of the ‘Brabants Vaarwater’ channel (Fig. 2). The mussel site was in a mussel culture plot of 4.5 ha area that was seeded with 5,000 tons fresh weight of mussels in December 2003. The average water depth was around 6 m. The control site was located about 300 m south of the mussel site, with a water depth of around 8 m. The bed morphology here was characterized by sand ripples transverse to the tidal flows with a wavelength of 10 m and amplitude of 0.5 m. Maximal tidal currents were observed during flood with a max. velocity of 0.9 m s−1 at both sites. The water column was thoroughly mixed. Vertical velocity gradients and eddy viscosity were slightly different between sites, due to the increased bottom roughness at the mussel site. Thus, the only detected differences in hydrography between the sites were part of the mussel effect (Riethmüller et al., in preparation). Ten box core samples were taken randomly within an area of 50 × 50 m at both sites.
In Ria de Vigo, rafts are organized in large polygons containing tens to hundreds of rafts. Sampling was done in a polygon in the outer part of the Ria, close to the northern shore in an embayment near the village of Cangas (Fig. 3). The area is characterized by alternating bays and headlands that reach out below the water surface to the central axis of the Ria. Three polygons with 68, 35 and 55 rafts are situated here. The arrangement of the rafts is in five parallel rows, with each row 100 m apart from the next. Three transects were laid out between rafts in the eastern polygon (mussel site) (Fig. 3). The reference samples were taken outside the polygons but at similar depths. For this purpose, two transects were laid out west of the polygons (reference site). In each transect, six box-core samples were taken at regular distances (100 m). For the mussel site, the first and sixth sample were located just outside the polygon, the other four samples were within the polygon, in between the rafts. The water depths ranged between 12 and 34 m. In all transects, depth increased towards the south. Hydrographic conditions, measured with a roving ADCP, were similar between the reference and mussel sites (Simpson, pers. comm.).
Sampling was done by different types of box cores in each study site. The surface areas of the samples taken at Limfjorden, Oosterschelde and Vigo were 0.0117, 0.0774 and 0.019 m2, respectively. From each box core a sediment sample of the upper cm (lump sample of three replicates) was collected with a syringe (1 cm ∅) for sediment and pigment analysis. These samples were immediately frozen and kept in the dark. The remaining of the core was sieved on board (mesh size 1 mm) and fixed with formaldehyde.
Additional video tracks were made to record the distribution of the mussels and/or presence of large epibenthic animals. In Limfjorden, this was done by a diver who videoed transects along pre-laid outlines with markers every 50 cm.
Laboratory analysis
The macrofauna was determined to the lowest taxonomic level possible. Species were classified into feeding groups based on food resource and feeding mechanism: subsurface deposit feeders (SSDF), surface deposit feeders (SDF), suspension feeders (SF), predators and omnivores. Biomass of small species was estimated by first establishing, using a selection of individuals, a regression line of wet weight to ash free dry weight (AFDW). For all other individuals wet weight was determined and converted to AFDW using the regression line. To estimate biomass of large or broken animals the AFDW was measured.
Sediment grain size was analyzed by laser diffraction, using the Malvern Mastersizer. Particulate organic carbon (POC) and nitrogen (PON) of the sediments were analyzed by a Carlo Erba elemental analyzer, type NA-1500. The phosphorus content of the sediments was determined by using Perkin Elmer ICP-OES, type Optima 3300DV. Photopigments were extracted from the freeze-dried sediment by adding 10 ml 90% acetone, placing it in a ball mill for 20 s and centrifuging for 5 min at 1,500 rpm. The supernatant was analyzed by HPLC following Jeffrey et al. (1997). Sediment chlorophyll a concentrations were determined as a relative measure of photosynthetic biomass. The phaeopigments phaeophytin a-like and phaeophorbide a-like pigments were used as indication for the presence of degradation products (chlorophyll derivatives) due to the feeding activity and digestive processes of the mussels (Hawkins et al. 1986).
Statistical analysis
The hypothesis underlying this study is that mussels, by biodeposition and habitat structuring, have predictable effects on the rest of the benthic community, regardless of type of culture or specific habitat characteristics. Therefore, we chose to replicate ecosystems, spanning a variety of mussel-dominated systems, rather than spend our efforts to replication within each of the systems. Within each of the systems our approach can be called pseudoreplicated since only a single mussel site was compared with a single control site. However, these sites were carefully selected to be as close to each other as practically possible (some distance is required to assure that no effects are transferred horizontally) and hydrographically similar. In addition, we demonstrated this similarity with extensive measurements. Overall, our study is composed of replicate sets of sites that, to our best knowledge, only differ in the presence of mussels. Any consistent trends in the difference between mussel and control sites can, therefore, be ascribed to the presence of mussels.
Within systems, we used Student’s t tests to test for differences in sediment properties and macrofaunal characteristics between the mussel sites and reference sites. The assumptions of homogeneity of variances and normality were confirmed using Levene’s and Kolmogorov–Smirnov tests. Total abundance and total biomass and abundances and biomass of macrofauna species were log transformed prior to analysis. Across systems, we rely on qualitative comparison because too many details differed in the sampling protocols (e.g. size of the box corers, influence of depth in one of the systems, species composition and species richness) to allow for a practical formal analysis.
Similarities and differences in macrofaunal communities were explored using non-metric multidimensional scaling (n-MDS), based on Bray-Curtis similarity indices on double square root, unstandardized data. Stress values indicate how well the solution (two-dimensional MDS plot) reflects the similarities among samples. Values <0.1 are good and <0.2 are useful (Clarke 1993). Overall and pair-wise comparisons for significant differences in macrofaunal composition between habitats were made using analysis of similarity (ANOSIM). SIMPER analyses were used to determine the percent of similarity of samples and the particular taxa responsible for differences between groups. Multivariate analyses and diversity measures were analyzed with the program Primer 5.0 (Clarke 1993).
Results
Characteristics of the sediment mussel matrix
The Limfjorden mussel site was characterized by a very patchy distribution of the mussel M. edulis (Fig. 4). Patch size ranged from small clumps of a few cm to large beds of mussels of >25 m. Based on the video transects the average coverage was estimated to be 27%. Average length of the mussels was 22.26 ± 0.32 mm. The length frequency distribution shows that mainly mussels of a single cohort are present (2-year old), with a few individuals of an older cohort (Fig. 5). Of the 25 box cores taken, nine contained none or only one individual of M. edulis. Average density of M. edulis was 3,911 ± 717 ind. m−2 (max. 10,000 ind. m−2); average biomass was 300 g AFDW m−2 (max. 1,011 g AFDW m−2).

Mussel characteristics at the three study sites. Left reconstruction of a 3.5 m of the Limfjorden mussel bed, based on video tracking (videoing done by Jens Larsen). Horizontal lines mark distances of 0.5 m. Right upper photo of the Oosterschelde mussel bed showing the dens coverage of mussels (Photo by Jens Larsen). Right lower photo of a raft culture in Vigo showing the ropes hanging down in the water column (Photo by Jens Larsen)

Length frequency distribution of the mussels observed at the field sites in Limfjorden (left) and Oosterschelde (right)
The mussel site in the Oosterschelde was in a mussel culture plot of 4.5 ha area that was seeded with 5,000 tons fresh weight of mussels M. edulis in December 2003. Based on the video transects a coverage of >75% was observed (Fig. 4); box cores always contained mussels. Mussels had on average a length of 55.1 ± 0.66 mm; all belonged to a single cohort (Fig. 5). Average density of M. edulis was 616 ± 139 ind. m−2 (max. 1,525 ind. m−2). Average biomass was 615 g AFDW.m−2 (max. 1,315 g AFDW m−2).
Biogeochemical characteristics of the seafloor sediments
A significant effect of the presence of the mussels on grain size distribution and mud content (fraction <63 μm) was observed in all three study areas (Table 1; Fig. 6). In Limfjorden median grain size at the mussel site was 1.3× lower than at the control site. No mud was observed at the control site, whereas at the mussel site mud content averaged 13 ± 1.2%. At the small spatial scale within the mussel site, samples containing mussels had a significantly (p < 0.01) lower median grain size and higher mud content than non-mussel samples (Table 1). In the Oosterschelde, differences between the mussel site and the control site were more pronounced compared to Limfjorden, with a ~3× lower median grain size at the mussel site and a mud content of 41 ± 5.7% compared to zero (Table 1). In Ria de Vigo, the situation is more complicated as the sediment grain size changed along the depth gradient, but on average a higher mud content and lower median grain size was found along the transects in the mussel site than along the transects in the control site (Fig. 6). The difference is most expressed at the largest depths.

Median grain size (μm), mud content (%), POC (organic carbon, %) and PON (nitrogen, %), chl a and phaeophytin a observed along the transects running through the mussel polygon (3 transects, 18 samples) and the reference site (2 transects, 12 samples) in Ria de Vigo. Lines represent linear regressions (full line mussel site; broken line reference site)
The chemical properties of the sediment showed similar trends as the trends observed for grain size and mud content, with significantly higher contents of POC, PON and phosphorus at the mussel sites than in the control sites (Table 1). In Limfjorden, values were 2 to 2.5 times higher at the mussel site and C:N ratio did not differ significantly between sites. At the small spatial scale within the mussel site, we observed also a significant difference (p < 0.01) with the m-site (samples with mussels) having a 1.5 to 2.5 times higher POC, PON and phosphorus content than the nm-site (samples without mussels). In the Oosterschelde larger differences were observed (factor 20, 15 and 50 for POC, PON and phosphorus, respectively; Table 1). C:N ratio was significantly higher at the mussel site. In Ria de Vigo POC, PON and phosphorus content varied with depth as did mud content (Fig. 6). Highest absolute POC, PON and phosphorus concentrations in our study were found in Vigo.
The chl a contents on average did not differ between the mussel bed and the control site in Limfjorden, whereas in the Oosterschelde chl a was ~40× higher at the mussel site (Table 1). In both Limfjorden and Oosterschelde, phaeopigment contents were significantly higher at the mussel site. At the small spatial scale within the mussel site in Limfjorden, we observed significantly higher chl a and phaeopigment contents at the m-site compared to the nm-site. In Vigo, compensating for depth, both chl a and (especially) phaeophytin a contents were higher at the mussel than at the control sites (Fig. 6).
Macrobentic diversity and community structure
Species richness and diversity
In Limfjorden, 23 species were observed at the mussel site; taxonomic groups included Annelida (74%, 17 sp.) and Mollusca (17%, 4 sp.), together with one species of echinoderms and one species of ascidians. Fourteen of these species were not found at the control site. At the control site, 19 species were identified; also here Annelida (63%, 12 sp.) and Mollusca (26%, 5 sp.) dominated, together with one species of echinoderms and one species of sea anemones. Ten species were not found at the mussel site. Epifaunal species were very rare at both sites: two species at the mussel site and one at the control site respectively. This was confirmed by the video transects. From these images we observed regularly the sea slug Philine aperta and the gastropod Hinia reticulata, and occasionally individuals of the crab Carcinus maenas and the sea star Asterias rubens. Species richness, diversity and evenness were similar at both sites (Table 2). At the small scale, however, within the mussel site a higher species richness was observed in the samples with mussels present, but with a significantly lower diversity and evenness (Table 3). In the m-site and the nm-site in total 20 and 12 species respectively were found.
In the Oosterschelde 54 species were identified at the mussel site; taxonomic groups included Annelida (58%, 31 sp.), Crustacea (26%, 13 sp.) and Mollusca (8%, 7 sp.), together with two species of echinoderms and one unidentified species of sea anemone. Forty of the identified species were not found at the control site. At that site 28 species were identified: Annelida (52%, 15 sp.), Crustacea (18%, 6 sp.), and Mollusca (15%, 4 sp.), together with two species of echinoderms. Thirteen species were not found at the mussel site. The number of epibenthic species was much higher in the mussel site compared to the control site (17 and 8 species, respectively). The video transects showed A. rubens and C. maenas as common species at the mussel site. Species richness, as well as diversity, was significantly higher at the mussel than at the control site, but evenness was lower (Table 2).
In Ria de Vigo a diverse macrofaunal community was observed, and not all individuals were determined at species level. In the mussel site 159 taxa were identified in the three transects (18 samples): Polychaeta (57%), Crustacea (21%), Mollusca (6%) and Echinodermata (6%) dominated. Sixty-three of the identified taxa were not found at the control site. In the control site, 152 taxa were identified in the two transects (12 samples). The same phyla dominated. Sixty-eight taxa were not found at the mussel site. The species richness in the reference site was 31.4 ± 2.26 per sample and in the mussel site 25.6 ± 2.18, but no significant difference was observed between the two sites, because of the large variability. Diversity and evenness were significantly higher in the reference site. No significant relation with depth was observed.
Abundance and biomass
In Limfjorden and Oosterschelde, the total mean abundance and biomass were significantly higher at the mussel site than at the control site, but this was completely attributed to the presence of the mussels (Table 2). Without mussels the mean abundance and biomass were significantly lower at the mussel site than at the control site in Limfjorden, whereas in the Oosterschelde the opposite (but non-significant) was observed. At the small spatial scale in Limfjorden, within the mussel site, the abundance and biomass at the m-site were significantly higher than at the nm-site (Table 3), but this difference was not significant anymore when omitting the mussels.
In Oosterschelde, endofaunal abundance in the mussel site (504 ind. m−2) was higher compared to the control site (357 ind. m−2), but endofaunal biomass was significantly lower; epifaunal biomass on the other hand was significantly higher in the mussel bed (Fig. 7).

Endofaunal biomass (left) and epifaunal biomass (right) in the mussel site and the control site in the Oosterschelde
In Ria de Vigo, total abundance did not differ significantly between the reference site and the mussel site (Table 4). Total biomass at the mussel site was significantly lower than at the reference site, but a large variability was observed because of the dominance of a few large individuals. No significant relation with depth was observed.
Feeding types
In Limfjorden, SF dominated the mussel site in density and biomass. At the control site, SSDF (mainly Heteromastus filiformis) numerically dominated, SF (Ensis) dominated in terms of biomass. The number of SSDF (H. filiformis) was significantly lower at the mussel site compared to the control site, whereas the number of SDF was higher. At the small spatial scale within the mussel site, the m-site had higher densities of SDF and predators than the nm-site. Small SDF like Polydora sp. and Syllidia armata and small predators like Harmothoe sp. only occurred at the m-site and not at the nm-site (Table 3).
In the Oosterschelde, SF dominated the mussel site in terms of density and biomass. At the control site SSDF (Scoloplos armiger) and predators (Nephtys cirrosa) numerically dominated. The biomass of SSDF was significantly lower at the mussel site, whereas endofaunal SDF biomass was higher. The biomass of epifaunal predators and omnivores was much higher at the mussel site than at the control site (Fig. 7).
In Ria de Vigo small polychaetes numerically dominated at the mussel site, mainly SDF (51%, Prinospio sp., Aphelochaeta marioni, Ampharete sp.) and SSDF (31%, H. filiformis) (Table 4). Predators (9%) and especially SF (3%) were of minor importance. SDF (47%, Spio sp., Prinospio sp., A. marioni, Paraonis fulgens) also dominated at the reference site. SSDF (19%, H. filiformis) were less dominant than at the mussel site, whereas SF (15%, e.g. Ampelisca sp., Actiniaria, Ophiothrix fragilis) and predators (13%, e.g. Lumbrineris sp., Sphaerosyllis sp.) were more abundant. Biomass at both sites was dominated by a few large individuals of SF, e.g. Acanthocardia tuberculata and Cucumaria frondosa at the mussel site, and Cucumaria elongata, C. frondosa, and Dosinia exolata at the reference site.
Benthic community structure
The MDS ordination analysis for Limfjorden showed a clear separation of samples between the control and mussel sites. This separation remained after removal of M. edulis from the data matrix (Fig. 8, ANOSIM p < 0.01). Species that were responsible for the significant dissimilarity (66%, without mussels) were the polychaetes H. filiformis and S. armiger (characteristic for control site) and Nereis sp. (mainly Nereis succinea), Harmothoe sp., S. armata, Polydora sp. and Capitella capitata (characteristic for mussel site; Table 2). The polychaete Pectinaria koreni, common at both sites, showed significantly higher densities at the mussel site. In addition, the samples from the control site were more similar within the site (average similarity 71%) than samples from the mussel site (similarity 52%). The community structure inside the mussel site had a rather large variability. This was to a large extent explained by the significant difference in community structure between the m- and nm-site (Fig. 8, ANOSIM p < 0.01).

MDS ordination diagrams for Limfjorden (stress 0.14, top), Oosterschelde (stress 0.08, middle) and Ria de Vigo (stress 0.21, bottom). The MDS shows the differences in community structure of the macrobenthos between the mussel site and control (reference) site. In Limfjorden, the mussel site is further split up into nm-site = samples without mussels and m-site = samples with mussels
In the Oosterschelde, the MDS (excl. M. edulis) indicated that the community structure of the control and mussel site significantly differed from each other (Fig. 8, ANOSIM p < 0.01). Species responsible for the significant dissimilarity (91%) were S. armiger, N. cirrosa and Magelona papillicornis (characteristic for control site) and Oligochaeta, Pygospio elegans, Streblospio shrubsoli, C. capitata and Harmothoe sp. (characteristic for mussel site; Table 2). Additionally, several epibenthic species were only observed in the mussel site (e.g. the amphipods Amphilochus neapolitanus, Abludomelita obtusata, Microdeutopus anomalus, Microprotopus maculatus and the decapods Athanas nitescens, C. maenas, Pinnotheres pisum; the latter living symbiotically in mussel shells).
In Ria de Vigo, the community structure significantly differed between the two sites (Fig. 8, ANOSIM p < 0.05), but both sites showed a large variability. The community of the mussel site was numerically dominated by H. filiformis and Prinospio sp. These two species also occurred at the reference site, but in lower abundances. Other species contributing to the dissimilarity between the mussel site and the reference site were Ampharete sp., Oligochaeta, C. capitata (characteristic for the mussel site) and Spio sp. and Ampelisca sp. (characteristic for the reference site; Table 4).
Discussion
Our study suggests that the presence of mussel cultures (both bottom and suspended cultures) change the sediment environment and thereby alter the biodiversity of the macrobenthic communities compared to nearby and otherwise very similar mussel-free areas. Comparisons of the sedimentary environment and associated benthic macrofauna of mussel bed and non-mussel bed locations may be confounded by location effects. In particular, different areas may experience different hydrodynamic conditions. In our study, we carefully checked (and measured) these conditions to reduce these differences as much as possible. However, we did not sample all possible conditions in each of these systems exhaustively. For that reason, we place emphasis in the interpretation on those patterns that are consistent across systems and culture practices, rather than on impacts of mussel cultures within each of the systems.
Sediment and physico-chemical characteristics
In all three study sites, the sedimentary environment was changed by the presence of mussels. Underneath or in the surrounding areas of the mussel beds and suspended raft cultures grain size was smaller and particulate organic carbon, nitrogen and phosphorus contents were higher than in nearby, culture-free sediments (with similar hydrodynamic conditions). Similar findings were reported for natural mussel beds (Ragnarsson and Raffaelli 1999), commercial bottom-cultured mussel beds (Smith and Shackley 2004) and suspended cultures (Chamberlain et al. 2001).
For mussel beds the combined effect of sediment trapping and biodeposition leads to high sedimentation rates. Biodeposition in beds of suspension-feeding bivalves results from the active filter feeding by the bivalves, which leads to non-digested material being excreted to the sediment surface as feces and pseudo-feces (Kautsky and Evans 1987; Norkko et al. 2001). Bivalves affect the structure of the sediment–water interface (Graf and Rosenberg 1997), resulting in high local deposition rates (Dame 1993) and enrichment of sediments in C and N (Kautsky and Evans 1987). In all three systems, the effect on the sedimentary environment was local and more or less limited to the mussel bed or its direct surroundings (see also Beadman et al. 2004). This local effect was very well demonstrated in Limfjorden, where within the mussel site we observed significant differences between patches of mussels and the bare sediment in between. A mussel transplant experiment in the intertidal by Ragnarsson and Raffaelli (1999) also showed a significant increase in silt content between mussel transplant plots and nearby (2 m) control plots.
The change of the sedimentary environment in the presence of a mussel bed was more pronounced in the Oosterschelde than in Limfjorden. Firstly, the control site had coarser sediment in the hydrodynamically more dynamic Oosterschelde, leading to larger differences. Secondly, aerial cover and total biomass of mussels was higher in Oosterschelde than in Limfjorden, enhancing both the biodeposition rate and the degree of cover (and protection against resuspension) of the sediment. Thirdly, feeding conditions and concentration of suspended sediment may have differed and this may also explain differences in amount of biodeposition. Finally, the age of the mussel bed may also influence its effect on the sedimentary environment. However, the age of the beds studied in Limfjorden (2 years) and Oosterschelde (1.5 years) were rather similar and this effect cannot be studied from our data.
Many studies have indicated that the primary environmental impact of bivalve suspended cultures is increased sedimentation due to biodeposition. Sedimentation rates have been reported to be 1.3–5.5 times higher compared to culture-free areas (Dahlbäck and Gunnarsson 1981; Hatcher et al. 1994; Grant et al. 1995; Callier et al. 2006; Giles et al. 2006, pers. observ; this study). The elevated POC, PON, phosphorus, and phaeo concentrations in the surficial sediments in the raft area in Ria de Vigo are consistent with measurements of enhanced organic input elsewhere (Dahlbäck and Gunnarsson 1981; Chamberlain et al. 2001; Stenton-Dozey et al. 2001; Christensen et al., 2003; Hartstein and Rowden 2004; Giles and Pilditch 2006; Giles et al. 2006). The observation of a strong increase in the phaeopigment concentration further suggests that a majority of the phytosynthetic pigments in the mussel-affected sediments were depositional chlorophyll degradation products derived from mussel fecal and pseudofecal materials (Mirto et al. 2000; Christensen et al. 2003; Giles et al. 2006).
Our hypothesis that biodepositional effects of mussels would be smaller in hydrodynamically rough circumstances than in calm conditions is rejected by the comparison between Oosterschelde and Limfjorden. Apparently (pseudo)fecal material is also deposited nearby the mussel bed under a strong current regime as in Oosterschelde. In this system, the physical structure of the (dense) bed can be expected to cause skimming flow, leading to protected conditions in the bed. Moreover, the strong hydrodynamic forces lead to much higher suspended matter concentrations in the water and thus increase the biodeposition rates of mussels as compared to the quiet clearer waters of Limfjorden.
Also in suspended culture, the distance over which an impact of mussels is measured is, in general, relatively small. Mussel farm impacts were confined to less than 50 m from the farm boundary in several studies (Mattsson and Lindén 1983; Chamberlain et al. 2001; Hartstein and Stevens 2005). The degree of environmental impact is directly related to the system’s ability to disperse the organic material originating from the mussel cultures (Chamberlain et al. 2001; Newell 2004), which is strongly linked to the local hydrodynamics (Hartstein and Rowden 2004; Hartstein and Stevens 2005). Hartstein and Stevens (2005) observed a significant effect of biodeposits on sediments underneath mussel farms in sheltered sites, reducing to natural levels approximately 30–50 m from the farm site, whereas a more exposed site showed no sign of any mussel deposits in sediments beneath or up to 200 m from the farm boundary. For this exposed site, they concluded that there is at time sufficient energy to re-suspend mussel biodeposits and to disperse these deposits over a wide enough area thereby leaving little impact on the natural sediment. In both sites of Ria de Vigo (polygon and reference) it was seen that the physico-chemical parameters showed a decrease in grain size and chl a and an increase in the other sediment parameters (mud content, POC, PON, phosphorus content, phaeo pigments) with increasing depth. This interaction with depth can be explained by the natural lateral transport of sediments to the drainage channel and by wave action in the more shallow stations that will disturb the sediments. The decrease in chl a with increasing depth can be further explained by the reduced light penetration with increasing depth. On top of this natural gradient, the mussel farm contributed to local organic enrichment through the enhanced biodeposition.
Compared to bottom cultures it was expected that the changes in sediments under a suspended culture would be larger, because of the much higher biomass of mussels per m2 surface in a suspended culture. This higher biomass leads to a higher biodeposition, and resuspension of biodeposits from intertidal or shallow-water bivalve populations is more likely than those from bivalves living in either deeper water or grown in suspended aquaculture systems, where the underlying sediments are isolated from frequent disturbance by wave action. Most parameters (e.g. mud content, POC, PON) were indeed several times higher in mussel site sediments in Ria de Vigo compared to Oosterschelde and Limfjorden, but highest values in the reference site stations also exceeded those from the reference sites in Oosterschelde and Limfjorden. The different hydrodynamic conditions make direct comparisons among the three study areas difficult.
Benthic macrofauna response: mussel beds as habitat
In both Limfjorden and Oosterschelde, we observed a significant change in the macrobenthos. A few, mainly intertidal studies have demonstrated significant effects of the presence of mussel beds on macrobenthic communities (Dittmann 1990; Ragnarsson and Raffaelli 1999; Beadman et al. 2004; Commito et al. 2005), although the strength and direction of these effects differ among studies. Dolmer (2002) suggested a positive relationship between mussel abundance and the number of associated species due to the complex substratum by the mussels, but Beadman et al. (2004) found that increasing the abundance of mussels decreased infaunal diversity and abundance. Different processes and mechanisms will influence the presence of the associated benthic macrofauna in mussel beds. The relative importance of each mechanism will determine the (combined) outcome of the ecosystem engineering effect of the mussels.
Firstly, the complex epibenthic mussel matrix provides a biogenic habitat that offers a habitat for epibenthic, hard substrate species, as well as shelter and predator refuge for mobile epibenthos (Seed and Suchanek 1992; Reise 2002; Gutiérrez et al. 2003). Therefore, it was hypothesized that mussel beds will increase the epifaunal biodiversity compared to the surrounding bare sediments. In our study this hypothesis was clearly supported for the Oosterschelde, where the number of (mainly mobile) epibenthic species and their biomass significantly increased in the mussel bed. In Limfjorden, epibenthic species were very rare overall and showed no increase in the mussel bed. Some polychaete species however, like Harmothoe, Pholoe and Polydora, were only present in the mussel bed in Limfjorden, and these species can be considered as epibenthic species, as they live in between the mussel matrix (Dittmann 1990). These species also occurred within the mussel matrix in the Oosterschelde. Increased epifaunal diversity in mussel beds is observed in several studies, both in the intertidal (Dittmann 1990; Ragnarsson and Raffaelli 1999; Asmus 1987) and the subtidal (Seed and Suchanek 1992; Saier 2002). Some of these species (e.g. crabs C. maenas and seastar A. rubens which increased in both Oosterschelde and Limfjorden) are predators of the mussels themselves and are thus also attracted by the mussels as prey.
Secondly, the biodeposition caused by the bivalves will locally enrich the sediment (see above). For endobenthos living conditions will tend to deteriorate with increasing intensity and persistence of the organic (bio)deposition, following the empirical model of Pearson and Rosenberg (1978). High rates of biodeposition may result in increased microbial activity and strongly reducing conditions in the sediments, leading to endobenthic communities with low diversity, dominated by opportunistic species such as capitellids and oligochaetes. Several authors observed a shift in the endobenthic community from one dominated by polychaetes in non-mussel areas to one dominated by oligochaetes in mussel beds (Commito and Boncavage 1989; Dittmann 1990; Ragnarsson and Raffaelli 1999), or at least a decline in polychaetes but not in oligochaetes in the presence of mussels (Beadman et al. 2004). In particular, the abundance of Tubificoides benedii (Commito and Boncavage 1989; Commito et al. 2005) and C. capitata (Ragnarsson and Raffaelli 1999) in mussel beds have been attributed to their tolerance for organically enriched, anoxic sediment. In the Oosterschelde, where the mussel bed was very dense, we also observed a shift from polychaetes to oligochaetes, with a dominance of Oligochaetes and C. capitata. In Limfjorden, with a more patchy mussel bed, C. capitata only occurred in the mussel patches, but Oligochaeta did not increase in abundance compared to the control site. On the other hand, several species (e.g. free living species like Scoloplos armiger and Nephtys cirrosa) were absent from the mussel sites or severely reduced in numbers in comparison to the reference sites.
Thirdly, the activity of the mussels themselves might influence the settlement of other species. Woodin (1976) predicted that dense suspension-feeding bivalves reduce the probability of successful larval settlement by any larvae, including their own. Several authors have debated this hypothesis, and showed that infaunal species, especially those that form cocoons, brood, fragment asexually, or disperse at large post-larval stages may be relatively more abundant in mussel beds than species with planktonic larval dispersal (Commito and Boncavage 1989; Dittmann 1990; Thiel and Ullrich 2002). A good example are Oligochaeta that reproduce by means of cocoons. Dolmer (2002), in contrast to the above hypothesis, suggested that the higher spatial complexity of mussel beds enhances larval settlement of several species.
Both in Limfjorden and Oosterschelde, the associated fauna in mussel beds constituted only about 3–4% of the total biomass. This is also observed in other studies (e.g. Asmus 1987). The biomass of the associated fauna within mussel beds often does not exceed that of fauna in surrounding habitats (Asmus 1987; this study). Thus, mussel beds do not necessarily enhance biomass (or production) of associated fauna but provide a habitat for particular species that otherwise could not exist in the surrounding unengineered soft-bottom environments (see also Crooks 2002).
Benthic macrofauna response: suspended mussel farming impacts
The feeding activity of suspended bivalve suspension feeders results in enhanced organic (bio)deposition, without however affecting the habitat structure. It was, therefore, hypothesized that it would mostly affect the endobenthos, promoting opportunistic species.
Overall, this hypothesis was confirmed in our study. Diversity significantly decreased under the mussel culture in Ria de Vigo, and also species richness, although not significantly, showed an average decrease. Total abundance slightly increased in the mussel site, and total biomass significantly decreased. The variability within each site was large, but the between variability was larger and showed significantly different benthic communities between the mussel and reference sites. Other studies on the influence of suspended bivalve culture on the benthic macrofauna show varying results. Most studies, including this study, observed a reduced macrobenthic species richness and diversity under suspended mussel cultures (Tenore et al. 1982; Mattsson and Lindén 1983; Kaspar et al. 1985; Stenton-Dozey et al. 2001). Callier et al. (2007) also observed a decrease under lines with mussels at least 1-year old (1+), whereas sites under lines with mussels less than 1-year old showed the greatest number of species compared to reference and 1+ sites, suggesting that some species have benefited from the moderate organic loading from the 0+ mussels. Most studies, including our own, observed a shift in the benthic community structure, but the analysis of trophic structure revealed different outcomes among different studies. In Ria the Vigo, we observed a shift towards small surface deposit feeders, as was also observed in the bottom cultures. Mattsson and Lindén (1983) observed higher abundances of deposit feeders at mussel culture sites compared to reference sites. Others, sampling right underneath rafts or longlines, observed a dominance of predators and carnivores that profit from mussel drop-off (Grant et al. 1995; Stenton-Dozey et al. 1999). Kaspar et al. (1985) indicated that the organic debris underneath suspended cultures had a smothering impact on filter feeders.
Several studies mention species-specific responses and one of the common examples showing increased abundance under mussel rafts is the capitellid worm C. capitata. Christensen et al. (2003) showed that the dissimilarity between farm-affected stations and a reference station was primarily due to the enhancement of small surface deposit-feeding polychaetes, such as C. capitata and Prinospio spp. On the other hand, large, bioturbating species, such as burrowing bivalves and crustaceans, and a dense population of suspension-feeding ophiuroids, were displaced, leading to important implications for the sediment bioturbating capacity (Christensen et al. 2003). These patterns were also observed by e.g. Mattsson and Lindén (1983), and Hartstein and Rowden (2004).
Impact of “foundation species” along stress gradients
It is generally assumed that ‘foundation species’ or ecosystem engineers have the highest impact in extreme physical environments (Crain and Bertness 2006), where they will ameliorate physical stress. In less stressful conditions, their effect may be reduced to the provision of competitor- or predator-free space. In mussels, these roles seem to largely coincide: alleviation of physical stress and provision of predator-free space are both a consequence of the habitat structure provided by mussel beds. For epibenthic species this has a clear effect, especially in hydrodynamically rough conditions as in Oosterschelde. It may explain why the effect on epibenthos was much more pronounced there than in Limfjorden. The biodeposition by mussel beds, however, is not related directly to hydrodynamic stress (see “Discussion”). Rather, it creates chemical stress and selects for stress-resistant endobenthic species without apparently promoting the use of these (spatially concentrated) organic resources in animal production. Thus, ecosystem engineering in this case cannot solely be viewed as a stress-ameliorating process, nor is it uniquely linked to physical stress gradients.
Conclusions
The presence of Mytilus cultures was shown to have significant effects on the sedimentary environment and the benthic macrofauna associated with these cultures. Although the three study areas in this study differed considerably in type of mussel culture and locality (e.g. hydrodynamic conditions), we observed some generalities that could be attributed to the presence and activity (biodepostion) of the mussels: (1) the sedimentary environment showed an increase in POC, PON, phosphorus and phaeopigment concentrations in the presence of mussel cultures, (2) infaunal macrobenthic community structure significantly changed from species which are typically present in sandy environments to small opportunistic SDF species, which are typically present in organically enriched sediments. The consistency of the patterns observed at all three study areas, which encompassed a range of environmental conditions and mussel culturing practices, does increase the generality of our findings. Additionally, our observations suggest that effects are rather locally and restricted to the nearby environment of the mussel cultures.
On the other hand, the present study also illustrates the habitat-forming effect of mussel beds. It is clear that the engineered habitat provides a resource that is not available in the unengineered environment. This has a positive effect on the biodiversity (through an increase in epibenthic diversity), which is not observed in sediments near suspended cultures. However, when comparing the biodiversity effects of bottom and suspended cultures, one should also bear in mind that the mussel ropes of suspended cultures are a habitat on itself that also attracts many epifaunal and infaunal species (Murray et al. 2007).
References
Alpine AE, Cloern JE (1992) Trophic interactions and direct physical effects control phytoplankton biomass and production in an estuary. Limnol Oceanogr 37:946–955
Asmus H (1987) Secondary production of an intertidal mussel bed community related to its storage and turnover compartments. Mar Ecol Prog Ser 39:251–266
Beadman HA, Kaiser MJ, Galanidi M, Shucksmith R, Willozs RI (2004) Changes in species richness with stocking density of marine bivalves. J Appl Ecol 41:464–475
Bruno JF, Bertness MD (2001) Habitat modification and facilitation in benthic marine communities. In: Bertness MD, Gaines SD, Hay ME (eds) Marine community ecology. Sinauer Associates, Sunderland, pp 201–218
Bruno JF, Stachowicz JJ, Bertness MD (2003) Inclusion of facilitation into ecological theory. Trends Ecol Evolut 18:119–125
Callier MD, Weise AM, McKindsey CW, Desrosiers G (2006) Sedimentation rates in a suspended mussel farm (Great-Entry Lagoon, Canada): biodeposit production and dispersion. Mar Ecol Prog Ser 322:129–141
Callier MD, McKindsey CW, Desrosiers G (2007) Multi-scale spatial variations in benthic sediment geochemistry and macrofaunal communities under a suspended mussel culture. Mar Ecol Prog Ser 348:103–115
Chamberlain J, Fernandes TF, Read P, Nickell TD, Davies IM (2001) Impacts of biodeposits from suspended mussel (Mytilus edulis L.) culture on the surrounding surficial sediments. J Mar Sci 58:411–416
Christensen PB, Glud RN, Dalsgaard T, Gillespie P (2003) Impacts of longline mussel farming on oxygen and nitrogen dynamics and biological communities of coastal sediments. Aquaculture 218:567–588
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Aust J Ecol 18:117–143
Cloern JE (1982) Does the benthos control phytoplankton biomass in South San Francisco Bay? Mar Ecol Prog Ser 9:191–202
Commito JA, Boncavage EM (1989) Suspension-feeders and coexisting infauna: an enhancement counterexample. J Exp Mar Biol Ecol 125:33–42
Commito JA, Celano EA, Celico HJ, Como S, Johnson CP (2005) Mussels matter: polstlarval dispersal dynamics altered by a spatially complex ecosystem engineer. J Exp Mar Biol Ecol 316:133–147
Crain CM, Bertness MD (2006) Ecosystem engineering across environmental gradients: Implications for conservation and management. Bioscience 56:211–218
Crawford CM, Macleod CKA, Mitchell IM (2003) Effects of shellfish farming on the benthic environment. Aquaculture 224:117–140
Crooks JA (2002) Characterizing ecosystem-level consequences of biological invasions: the role of ecosystem engineers. Oikos 97:153–166
Dahlbäck B, Gunnarsson LÅH (1981) Sedimentation and sulphate reduction under mussel culture. Mar Biol 63:269–275
Dame RF (1993) Bivalve filter feeders in estuarine and coastal processes. NATO ASI series G: ecological sciences 33. Springer, Heidelberg
Dame RF (1996) Ecology of marine bivalves: an ecosystem approach. CRC Press, Boca Raton, 254 pp
Dittmann S (1990) Mussel beds—amensalism or amelioration for intertidal fauna. Helgolander Meeresuntersuchungen 44:335–352
Dolmer P (2002) Mussel dredging: impact on epifauna in Limfjorden, Denmark. J Shellfish Res 21:529–537
Dolmer P, Frandsen RP (2002) Evaluation of the Danish mussel fishery: suggestions for an ecosystem management approach. Helgoland Mar Res 56:13–20
Dolmer P, Kristensen PS, Hoffmann E (1999) Dredging of blue mussels (Mytilus edulis L.) in a Danish sound: stock sizes and fishery-effects on mussel population dynamic. Fish Res 40:73–80
Figueiras FG, Labarta U, Reiriz MJF (2002) Coastal upwelling, primary production and mussel growth in the Rias Baixas of Galicia. Hydrobiologia 484:121–131
Giles H, Pilditch CA (2006) Effects of mussel (Perna canaliculus) biodeposit decomposition on benthic respiration and nutrient fluxes. Mar Biol 150:261–271
Giles H, Pilditch CA, Bell DG (2006) Sedimentation from mussel (Perna canaliculus) culture in the Firth of Thames, New Zealand: impacts on sediment oxygen and nutrient fluxes. Aquaculture 261:125–140
Graf G, Rosenberg R (1997) Bioresuspension and biodeposition: a review. J Mar Syst 11:269–278
Grant J, Hatcher A, Scott DB, Pocklingtion P, Schafer CT, Winter GV (1995) A multidisciplinary approach to evaluating impacts of shellfish aquaculture on benthic communities. Estuaries 18:124–144
Gutiérrez JL, Jones CG, Strayer DL, Iribarne OO (2003) Mollusks as ecosystem engineers: the role of shell production in aquatic habitats. Oikos 101:79–90
Hartstein ND, Rowden AA (2004) Effect of biodeposits from mussel culture on macroinvertebrate assemblages at sites of different hydrodynamic regime. Mar Env Res 57:339–357
Hartstein ND, Stevens CL (2005) Deposition beneath long-line mussel farms. Aquac Eng 33:192–213
Hatcher A, Grant J, Schofield B (1994) Effects of suspended mussel culture (Mytilus spp.) on sedimentation, benthic respiration and sediment nutrient dynamics in a coastal bay. Mar Ecol Prog Ser 115:219–235
Hawkins AJS, Bayne BL, Mantoura RFC, Llewellyn C (1986) Chlorophyll degradation and absorption throughout the digestive system of the blue mussel Mytilus edulis L. J Exp Mar Biol Ecol 96:213–223
Jeffrey SW, Mantoura RFC, Wright SW (1997) Phytoplankton pigments in oceanography, 1 edn. United Nations International
Jones CG, Lawton JH, Shachak M (1994) Organisms as ecosystem engineers. Oikos 69:373–386
Jones CG, Lawton JH, Shachak M (1997) Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78:1946–1957
Kaspar HF, Gillespie PA, Boyer IC, MacKenzie AL (1985) Effects of mussel aquaculture on the nitrogen cycle and benthic communities in Kenepura Sound, Marlbrough Sounds, New Zealand. Mar Biol 85:127–136
Kautsky N, Evans S (1987) Role of biodeposition by Mytilus edulis in the circulation of matter and nutrients in a Baltic coastal ecosystem. Mar Ecol Prog Ser 38:201–212
Mattsson J, Lindén O (1983) Benthic macrofauna succession under mussels, Mytilus edulis L. (bivalvia), cultured on hanging long-lines. Sarsia 68:97–102
Mirto S, La Rosa T, Danovaro R, Mazzola A (2000) Microbial and meiofaunal response to intensive mussel-farm biodeposition in coastal sediments of the Western Mediterranean. Mar Pollut Bull 40:244–252
Murray LG, Newell CR, Seed R (2007) Changes in the biodiversity of mussel assemblages induced by two methods of cultivation. J Shellfish Res 26:153–162
Newell RIE (2004) Ecosystem influences of natural and cultivated populations of suspension-feeding bivalve mollusks: a review. J Shellfish Res 23:51–61
Norkko A, Hewitt JE, Thrush SF, Funnell GA (2001) Benthic–pelagic coupling and suspension-feeding bivalves: linking site specific sediment flux and biodeposition to benthic community structure. Limnol Oceanogr 46:2067–2072
Pearson TH, Rosenberg R (1978) Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr Mar Biol A Rev 16:229–311
Ragnarsson SA, Raffaelli D (1999) Effects of the mussel Mytilus edulis L. on the invertebrate fauna of sediments. J Exp Mar Biol Ecol 241:31–43
Reise K (2002) Sediment mediated species interactions in coastal waters. J Sea Res 48:127–141
Saier B (2002) Subtidal and intertidal mussel beds (Mytilus edulis L.) in the Wadden Sea: diversity differences of associated epifauna. Helgol Mar Res 56:44–50
Seed R, Suchanek TH (1992) Population and community ecology of Mytilus. In: Gosling E (ed) The mussel Mytilus: ecology, physiology, genetics and culture. Elsevier, Amsterdam, pp 87–169
Smaal AC (2002) European mussel cultivation along the Atlantic coast: production status, problems and perspectives. Hydrobiologica 484:89–98
Smith J, Shackley SE (2004) Effects of a commercial mussel Mytilus edulis lay on a sublittoral, soft sediment benthic community. Mar Ecol Prog Ser 282:185–191
Stenton-dozey JME, Jackson LF, Busby AJ (1999) Impact of mussel culture on macrobenthic community structure in Saldanha Bay, South Africa. Mar Pollut Bull 39:357–366
Stenton-dozey JME, Probyn T, Busby AJ (2001) Impact of mussel (Mytilus galloprovincialis) raft-culture on benthic macrofauna, in situ oxygen uptake, and nutrient fluxes in Saldanha Bay, South Africa. Can J Fish Aquat Sci 58:1021–1031
Tenore KR et al (1982) Coastal upwelling in the Rias Bajos, n.w. Spain, contrasting benthic regimes of the Rias de Arosa and de Muros. J Mar Res 40:701–772
Thiel M, Ullrich N (2002) Hard rock versus soft bottom: the fauna associated with intertidal mussel beds on hard bottoms along the coast of Chile, and considerations on the functional role of mussel beds. Helgol Mar Res 56:21–30
Tsuchiya M, Nishihira M (1985) Islands of Mytilus as a habitat for small intertidal animals: effect of island size on community structure. Mar Ecol Prog Ser 25:71–81
Wiles PJ, van Duren LA, Haëse C, Larsen J, Simpson JH (2006) Stratification and mixing in the Limfjorden in relation to mussel culture. J Mar Syst 60:129–143
Woodin SA (1976) Adult-larval interactions in dense infaunal assemblages: patterns of abundance. J Mar Res 34:25–41
Acknowledgments
We want to thank the local authorities of the different study sites, the mussel farmer organization OPMEGA in Vigo and the mussel farmer Mr. Padmos in the Oosterschelde for their collaboration. Special thanks to the crews of RV Genetica, RV Luctor and RV Mytilus for their help with the sampling. Special thanks to Jens Larsen for providing the underwater photos and for the diving in Limfjorden. This work was funded by the EU project MaBenE (EVK3-CT-2002-00071). The authors acknowledge the support by the MarBEF Network of Excellence ‘Marine Biodiversity and Ecosystem Functioning’ (RMP MARENG) which is funded by the Sustainable Development, Global Change and Ecosystems Programme of the European Community’s Sixth Framework Programme (GOCE-CT-2003-505446). This publication is contribution number MPS-08055 of MarBEF and NIOO-KNAW publication number 4451.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by K. Reise.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Ysebaert, T., Hart, M. & Herman, P.M.J. Impacts of bottom and suspended cultures of mussels Mytilus spp. on the surrounding sedimentary environment and macrobenthic biodiversity. Helgol Mar Res 63, 59–74 (2009). https://doi.org/10.1007/s10152-008-0136-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-008-0136-5