
Abstract
In order to investigate the effect of severe winters on the long-term variability of benthic macrofauna communities four stations from the inner German Bight towards the Dogger Bank (North Sea) were sampled in March 1990 and from May 1995 to 2002. The stations were chosen to reflect a gradient in the hydrographic regime, temperature and organic matter supply. We initially hypothesized that the effect of severe winters on benthic communities will be more pronounced in the offshore more stable environment than in the nearshore regions. Our results clearly showed the opposite pattern. The benthic communities at the nearshore stations in the German Bight changed dramatically in species abundance and community structure after the severe winter 1995/1996. But the community structure in the German Bight returned to the previous stage within 2 years. In contrast, the offshore stations in the Oyster Ground and at the Dogger Bank remained rather unaffected by the severe winter and changed gradually during the study period. The results are discussed concerning the ecological importance of severe winters for benthic communities and the utility of our results for interpreting long-term changes.
Similar content being viewed by others


Introduction
Because of its relatively stationary habit and, thus, its incapability to avoid occasional unfavourable conditions, benthic macrofauna is regarded to be a good indicator of environmental changes in the marine environment. The spatial distribution of benthic macrofaunal communities in the southern North Sea is significantly ruled by gradients of different environmental factors such as temperature, food availability and hydrography (Heip et al. 1992; Kröncke et al. 2004). Also long-term changes in benthic communities were mainly related to winter temperatures and storm frequency (Kröncke et al. 1998, 2001; Schroeder 2003). However, episodic events such as extremely cold winters can dramatically change the diversity and structure of benthic communities, which may make the detection of long-term trends more difficult. For example, Wieking and Kröncke (2001) observed a reverse development of long-term trends in different parts of the Dogger Bank due to the cold winter 1995/1996. However, such events are characteristic features of marine areas in temperate regions.
Several investigations in the Wadden Sea and coastal areas have revealed that cold winters affect macrofaunal communities often dramatically, causing decreases in species number, abundance and biomass (Ziegelmeier 1964, 1970; Reichert and Dörjes 1980; Buchanan and Moore 1986; Beukema 1990, 1992; Beukema et al. 1996; Kröncke et al. 1998, 2001; Strasser and Pieloth 2001; Schroeder 2003; Strasser et al. 2003). Furthermore, highly successful recruitment after cold winters can result in increased biomass (Beukema 1990, 1992) indicating the importance of cold winters for the structure of littoral benthic communities.
Although Ziegelmeier (1964, 1970) and Schroeder (2003) described a negative effect of cold winters in the German Bight, only little information is available for more offshore areas of the North Sea. Wieking and Kröncke (2001) found a decrease in macrofaunal abundance on the Dogger Bank in 1996 in comparison with 1997 and 1998, which they discussed as an effect of the extreme winter 1995/1996. Several studies in the coastal areas of the southern North Sea indicate that the effects of cold winters may differ between areas. In the Dutch Wadden Sea the effect of the cold winter 1978/1979 was more evident in the lower than at the higher tidal flats (Beukema 1990). In the benthic community at Norderney (East Frisian Wadden Sea) the sublitoral was found more affected by the severe winter of 1978/1979 than the littoral (Dörjes et al. 1986). In the German Bight Schroeder (2003) found that the sublitoral macrofauna was more affected by the cold winter 1995/1996 at an offshore station in the Oyster Ground than at the nearshore stations. These results point to differently severe effects of cold winters on macrofaunal communities depending not only on abiotic parameters such as minimum temperature but also on the benthic community structure itself.
A long-term study of macrofaunal communities along a transect from the inner German Bight to the Dogger Bank, which was initiated during the ICES/IOC workshop on “Biological effects of contaminants” in March 1990 (Stebbing and Dethlefsen 1992), was analysed to evaluate the effect of the cold winter 1995/1996 on the interannual variability of the macrofaunal communities. The station transect followed gradients in temperature, organic matter supply and hydrographic conditions from near- to offshore regions (Becker et al. 1992; Reiss and Kröncke 2004, 2005).
We hypothesize that (1) the effect of cold winters on the benthic macrofauna was more pronounced in offshore than in nearshore areas where the macrofauna is adapted to a high natural variability of environmental parameters, and (2) that the recovery of the macrofaunal communities will take longer in offshore areas.
Materials and methods
Area of investigation
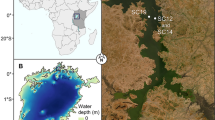
In March 1990 and May 1995–2002, four stations (Fig. 1) were sampled along a transect from the inner German Bight towards the Dogger Bank (Stebbing and Dethlefsen 1992). The depth at the four stations varied from 27 m in the inner German Bight to 43 m in the Oyster Ground (Table 1). The sediments consisted of mud at station GB2, muddy sands at stations GB5 and OG7 and fine sands at station DG9.

Study sites in the North Sea
Sampling and sample treatment
From May 1995 to 2002 sampling was carried out with RV “Senckenberg”. Samples were taken with a 0.1 m² van Veen grab, except in 1995 when a small box corer (122 cm²) was used. In March 1990, the samples were taken with two different grabs (Kröncke and Rachor 1992). At stations GB2 and GB5 a 0.1 m² van Veen grab, at stations OG7 and DG9 a 0.2 m² van Veen grab was used. All samples were sieved over 1 mm screens and fixed in 4% buffered formalin.
Sediment data
Sediment samples for the analyses of mud and sand contents as well as for total organic carbon (TOC) content analyses were taken from a separate grab. For mud contents, sediments were sieved over 63 μm mesh size.
TOC analyses were determined on freeze-dried total sediment that had been finely powdered and homogenised. An aliquot of 10–30 mg was combusted at 1010°C in a C/N analyser (Vario EL, elementar analysen) following acidification with concentrated HCl in a desiccator to remove inorganic carbonates (Hedges and Stern 1984).
Sea-surface temperatures
The Federal Maritime and Hydrographic Agency of Germany (BSH), Hamburg provided weekly data of the water surface temperature. The sea surface temperature (SST) anomalies were calculated based on the long-term average (1973–1993) per month.
Statistics
We used the PRIMER v5 program package to perform multidimensional scaling for abundance data to reveal similarities between stations (Clarke and Warwick 1994). Similarities were calculated using the Bray-Curtis coefficient. Square-root transformation was used prior to computation. Similarity percentage analysis (SIMPER) was used to analyse dominance of species. To test for a similar multivariate pattern and, thus, a similar temporal development a Spearman rank correlation was calculated using the RELATE routine of PRIMER. Non-parametric ANOVA (Kruskal–Wallis) was used to assess temporal changes in infauna parameters.
According to the different grabs used, the Hurlbert Index (ES100) was calculated for analysis of species diversity, since this index is less sample size dependent than other diversity indices or species richness (Hurlbert 1971).
Results
Sea-surface temperature
The effect of the cold winter 1995/1996 is clearly reflected in the anomalies (Fig. 2). At each station the mean SST in the coldest month (March 1996) was more than 2°C below the long-term average (1973–1993). This anomaly decreased from the German Bight towards the Dogger Bank, ranging between −3.4°C at station GB5 and −2.1 at station DG9. In the following years until 2003 the anomalies were mainly above the long-term average at each station (Fig. 2).

Deviations of mean monthly sea surface temperatures from the long-term average (1973–1993) (anomalies)
A decreasing gradient from the German Bight towards the Dogger Bank was also found for the absolute SST. SST in the coldest months (February and March) were at the offshore stations OG7 and DG9 about 2°C higher than at the nearshore stations GB2 and GB5. In contrast, for the warmest month (August) the situation was inverse, with SST 1–2°C higher in the nearshore than in the offshore region. Furthermore, the differences between the German Bight (GB2 and GB5) and the Oyster Ground (OG7) and Dogger Bank (DG9) were even more pronounced for bottom water temperatures in summer, since stations OG7 and DG9 had a stratified water column during summer while stations GB2 and GB5 had a mixed water column throughout the year (Otto et al. 1990; Becker et al. 1992; Reiss and Kröncke 2004).
Spatial distribution and temporal variability of sediments
Mud contents were highest at station GB2 (50%) and decreased towards the offshore stations with about 40% at station GB5, 20% at station OG7 and about 2% at station DG9. The intra-annual variability in mud contents was rather low at all stations, but in the Oyster Ground (OG7) a thin fluffy layer of mud was occasionally found at the top of the sediment, resulting in remarkable small-scale differences between samples in terms of surface mud content (H. Reiss, Unpublished data).
The TOC content of the sediment decreased from the German Bight towards the Dogger Bank, with 1.15 and 0.30% at stations GB2 and GB5, respectively, 0.21% at station OG7 and 0.10% at station DG9. No interannual trend in TOC content was found, but since TOC data from 1995 were lacking, no information about changes of TOC content due to the severe winter were available.
Spatial distribution of macrofaunal communities
Multidimensional scaling shows that the macrofauna at the different stations can be attributed to the following communities:
-
Stations GB2 and GB5: Nucula-nitidosa-community in the inner German Bight;
-
Station OG7: Amphiura-filiformis-community in the Oyster Ground;
-
Station DG9: Bathyporeia-elegans-Fabulina-fabula-community at the Dogger Bank.
These communities have already been described by Salzwedel et al. (1985), Kröncke and Rachor (1992), Kröncke (1992), Rachor and Nehmer (2003), Wieking and Kröncke (2003), Kröncke et al. (2004) and Reiss and Kröncke (2005). They are dominated by the species given in Table 2.
Temporal variability of macrofaunal communities
Species number, species diversity and abundance
The lowest total species number was found in the German Bight (GB5) with 111 taxa, whereas the highest total species number was found at the Dogger Bank (DG9) with 142 taxa. A similar increasing gradient from the inner German Bight towards the Dogger Bank was also found for the Hurlbert Index (ES100), which varied mostly between 15 and 20 at station GB2, and between 25 and 30 at station DG9 (Fig. 3).

Mean abundance (ind. m−2) and mean Hurlbert Index (ES100) for each station
At station GB2 the ES100 increased after the cold winter 1995/1996 from 14.3 in 1995 to 28.2 in 1997 and dropped again to 14.5 until 2000. A similar pattern was found at station OG7 in the Oyster Ground with a peak in 1997 of 30.5 (Fig. 3). At station GB5, however, an opposite pattern was observed, with a slight decrease of the ES100 after the cold winter from 24.8 in 1995 to 18.3 in 1997 (Fig. 3), followed by an increase.
At station GB2, maximum abundance was found in 1995 with 11,900 ind. m−2. After the cold winter 1995/1996 it dropped significantly to 500 ind. m−2 (ANOVA, F 1,2=25.49, P>0.05), but later on increased continuously until 2002 to about 6,000 ind. m−2. At station GB5, a similar pattern was found with a significant decrease in abundance from 1995 until 1996 (ANOVA, F 1,4=27,33, P<0.01). However, abundance in 1995 was generally lower than at station GB2. No significant change in abundance after the cold winter 1995/1996 was found at station OG7 (ANOVA, F 1,10=0.52, P<0.001), where abundance was rather uniform throughout the years with a slight increase towards 2002. At station DG9, interannual variability in abundance was rather high and primarily caused by the polychaete species Spiophanes bombyx (Table 2). Thus, the decrease in abundance in 1996, although not significant (ANOVA, F 1,10=4.6, P>0.05), may not be caused by the cold winter since similar patterns were found in later years (Figs. 3, 9).
Community structure
The MDSs (Figs. 4, 5) show that the community structure at all stations in 1990 was different from that in the other years. This difference was most pronounced at station DG9, where this year had to be excluded as an outlier (Fig. 5).

MDS plot of all four stations for the whole study period based on square root transformed abundance data; consecutive years are connected by arrows to elucidate temporal changes at the German Bight stations GB2 (dotted lines) and GB5 (solid lines)

MDS plots based on square root-transformed abundance data; consecutive years are connected by arrows
At stations GB2 and GB5 the community structure in the year 1995 differed remarkably from that in the following years 1996 and 1997 (Fig. 5). From 1998 until 2002 the community structure recovered and became more similar to the previous situation in 1995. This evident change in community structure after 1995 was not found in the Oyster Ground (OG7) and at the Dogger Bank (DG9) (Figs. 4, 5). At these stations the community structure changed gradually from 1995 until 2002. In Table 3 the results of the comparison between the similarity matrices of each station are given. Significant correlations in the changes of the community structure were found between the nearshore stations GB2 and GB5, as well as between the offshore stations OG7 and DG9, indicating a similar temporal development in both regions. However, there were no significant correlations between the German Bight stations and the offshore stations (Table 3).
The MDS combining all four stations over the whole sampling period clearly shows that the most drastic effects of the cold winter on community structure occurred at the German Bight stations GB2 and GB5 (Fig. 4). Less effects and, thus, higher interannual similarity in community structure was found at stations OG7 and at station DG9.
Species abundance
At station GB2, the bivalve Nucula nitidosa is the dominant and characteristic species of the community (Fig. 6). Abundances varied between 105 (1996) and 2,057 (2000) ind. m−2. The polychaete Owenia fusiformis occurred in highest abundances (7,171 ind. m−2) in 1995 before the cold winter as did the polychaete Scalibregma inflatum and the bivalve Abra alba. After the cold winter, these three species disappeared completely. They have been found again since 1998, but until 2002 did not return to abundances similar to 1995. The cumacean Diastylis rathkei was found between 1998 and 2000 at station GB2 with 100–553 ind. m−2, but decreased to 30 ind. m−2 in 2002. The echiurid Echiurus echiurus, an indicator of ice winters, was found in 1998 only. Juvenile ophiurids and the polychaete Nephtys hombergii occurred rather constantly in moderate to high abundance at station GB2, except of juvenile ophiurids in 2002.

Temporal changes in mean abundance (ind. m−2) of dominant and characteristic species at station GB2; no samples were taken in 2001
At station GB5, N. nitidosa was the dominant species similar to station GB2. However, in contrast to the other abundant species, it did not decrease as dramatically nor even disappeared after the cold winter (Fig. 7). Juvenile asteroids and the polychaete Lanice conchilega were the first to reappear in high abundances in 1998 or 1999, S. inflatum followed in 2000. All other species were found in lower abundances in the years after the cold winter.

Temporal changes in mean abundance (ind. m−2) of dominant and characteristic species at station GB5
At station OG7, the ophiurid Amphiura filiformis was the dominant species. Its decrease in abundance might have been caused by the cold winter although the abundance of all other abundant species remained unaffected (Fig. 8). The bivalve Mysella bidentata, a commensal of A. filiformis, was most abundant in 1990 and 1995, but decreased continuously from 1995 to 2002. Juvenile ophiurids and the bivalve Corbula gibba increased suddenly to high abundances in 2000 and 1999, respectively. All other species occurred in rather low to moderate abundances throughout the years.

Temporal changes in mean abundance (ind. m−2) of dominant and characteristic species at station OG7
At station DG9, no decrease in abundance could be recorded after the cold winter, except perhaps for L. conchilega (Fig. 9). The amphipod Bathyporeia elegans is the dominant species of the community with continuously increasing abundance since 1996 (up to 780 ind. m−2 in 2002). Most obvious is the high interannual variability in abundance of the polychaete S. bombyx with values ranging from 22 (1990) to 6,200 (1998) ind. m−2. No clear patterns in abundance over the study period were found for other abundant species.

Temporal changes in mean abundance (ind. m−2) of dominant and characteristic species at station DG9
Discussion
A comparison of the quantitative investigations on the benthic macrofauna in the eastern North Sea in the 1920 s by Hagmeier (Hagmeier 1925; Salzwedel et al. 1985; Kröncke and Rachor 1992; Rachor and Nehmer 2003 and the present study) suggest that the spatial distribution of macrofaunal communities has remained fairly stable since the 1920s. An exception seems to be the N. nitidosa community, which in 1975 (Salzwedel et al. 1985) and 1986 (Künitzer et al. 1992) was restricted to the inner German Bight, but then expanded towards the Oyster Ground (Kröncke and Rachor 1992; Rachor and Nehmer 2003).
Changes of community structure
Irrespective of the relative stability of the spatial macrofaunal community distribution and the basic community structure on a long-term scale, species abundance and biomass can vary considerably on shorter time scales (inter- and intraannual, interdecadal). On seasonal scale, Reiss and Kröncke (2004, 2005) found that the community structure of the infauna and epifauna was more variable at the nearshore station GB5 than at the offshore stations OG7 and DG9. This correlates with a higher seasonal variability of physical factors such as temperature, stratification of water column as well as primary production in the nearshore compared to the offshore area. Schroeder (2003) detected changes in community structure at four stations in the German Bight on a decadal scale in the 1970s, 1980s and 1990s, which were also found for the macrofauna communities off the island of Norderney and at the Dogger Bank (Kröncke et al. 1998, 2001; Wieking and Kröncke 2001). These changes were mainly caused by changes in the hydroclimate of the North Sea related to the North Atlantic Oscillation (Reid and Edwards 2001).
Rachor’s stations Slt and SSd (in Schroeder 2003) are almost identical with our stations GB2 and GB5 in the German Bight, and Rachor’s station WB is close to our station OG7 in the Oyster Ground. Both data sets show similar patterns of temporal changes in communities since 1995. The most notable change in community structure during this time period was related to the extremely cold winter 1995/1996.
In contrast to our initial hypothesis that the effect of the cold winter 1995/1996 on the macrofauna should be more pronounced in offshore areas than in nearshore areas where the fauna is adapted to a high natural variability of environmental parameters, our results clearly show the opposite pattern. The offshore stations at the Dogger Bank (DG9) and the Oyster Ground (OG7) remained rather unaffected, while benthic communities at the nearshore stations in the German Bight (GB2 and GB5) changed remarkably in abundance and community structure.
Drastic decreases in macrofaunal species number, abundance and biomass after cold winters are well documented for the Wadden Sea and the German Bight (see Introduction). In the Wadden Sea, most species recovered from cold winters within 1–2 years (Beukema 1990). In the present study, we found that changes in the communities in the German Bight (GB2 and GB5) after the winter 1995/1996 lasted until 1998, when their structure became again similar to that in the early 1990s.
In contrast to nearshore communities, those at the offshore stations OG7 and DG9 did not change seriously after the cold winter. However, the community structure at the offshore stations changed gradually; in 2002 the communities at stations OG7 and especially DG9, were less similar to those in 1995 than the communities at the nearshore stations GB2 and GB5.
The winter temperature is supposed to be of major importance for long-term changes of benthic communities (Dippner and Ikauniece 2001; Kröncke et al. 2001; Schroeder 2003).
A clear shift in benthic community structure in the southern North Sea occurred in the late 1980s; it was correlated to the North Atlantic Oscillation Index (NAOI) probably mediated by the rising winter SST. A period of mild winters resulted in an increase of abundance, biomass and species number of macrofauna (Kröncke et al. 1998, 2001). Dippner and Ikauniece (2001) also found that the interannual variability of macrofaunal biomass in the Gulf of Riga was mainly controlled by the climate variability during winter-time. Although the duration of our study was too short to draw conclusions about long-term trends, our results clearly show that the interannual variability of benthic communities depends on station. Despite the fact that severe winters are part of the natural variability in a temperate marine ecosystem, these events might mask long-term trends in near-shore areas or at least make their statistical detection more difficult. Thus, offshore communities might mirror large-scale long-term changes of the environment, e.g. changes in the climatic or hydrographic regime, more clearly than nearshore communities.
Changes on species level
Ziegelmeier (1970) and Schroeder (2003) found the echiurid E. echiurus to be r-selected, occupying the empty sediments after severe winters. The same occurred in our study after the winter 1995/1996 at station GB2. Echiurids were first recorded in our samples in 1998, in rather high abundance (165 ind. m−2) of big individuals, but the species disappeared again 1 year later as described also by Ziegelmeier (1970) and Schroeder (2003).
The abundance of N. nitidosa from 1996 to 2002 was similar to that described by Schroeder (2003) at Rachor’s stations Slt and SSd. Ziegelmeier (1964, 1970) found that N. nitidosa suffered less from severe winters because it is able to escape extreme temperatures due to its vagile habit. This conforms to our findings that there was only a slight decrease in abundance at station GB5 after 1995. At station GB2, however, N. nitidosa decreased in abundance (Fig. 6), although the minimum winter temperature in 1995/1996 was slightly higher at station GB2 than at station GB5. As N. nitidosa is sensitive to anoxic conditions (Rachor 1977, 1980), local hypoxia due to the decay of high amounts of dead macrofauna might explain the decrease of N. nitidosa at station GB2, although water temperature in spring is generally too low to support hypoxia. But nearly all O. fusiformis with a maximum abundance of 7,000 ind. m−2 had died during the winter 1995/1996.
Ziegelmeier (1964) mentioned that A. filiformis suffers most from severe winters and needs many years to recover. Schroeder (2003) confirmed the almost complete extinction of A. filiformis after the cold winters 1978/1979 and 1995/1996. After 1979 it took until 1991for the species to increase in abundance. This development was stopped again by the cold winter 1995/1996.
The expansion of the N. nitidosa community towards the Oyster Ground found by Kröncke and Rachor (1992) in 1990 and Rachor and Nehmer (2003) in 2000 might be a response to the low abundance of A. filiformis after cold winters. Salzwedel et al. (1985) and Künitzer et al. (1992) sampled during periods with higher abundances of A. filiformis. When ophiurid abundance is low N. nitidosa might be favored by competitive release.
An alternative explanation for the disappearance of A. filiformis from the German Bight region, and thus for the expansion of N. nitidosa might be a shift in the hydrodynamic regime in this area. This is reflected in a positive NAOI since the late 1980s. The NAO influences weather phenomena in NW Europe such as precipitation, sea surface temperature, direction and flow of currents and stability of the water column (Hurrell 1995; Planque and Taylor 1998; Reid et al. 1998). In the North Sea A. filiformis is mainly restricted to deeper regions with a stratified water column in summer (Künitzer et al. 1992). This in turn might be related to the feeding mode of this brittle star, which mainly feeds on macrofloculated organic matter, the production of which is supported by the stratification of the water column (Jones et al. 1998). However, a recent increase in strong westerly winds disturbs the stratification process and thus might reduce the food availability for A. filiformis.
In the present study, offshore stations (OG7 and DG9) were found to be less affected by the cold winter 1995/1996 than nearshore stations (GB2 and GB5). At station OG7 a slight decrease in abundance was found only for A. filiformis and M. bidentata. At station DG9 hardly any effect of the cold winter could be seen except for L. conchilega, a polychaete that is known to be very sensitive to cold temperatures (Buhr 1981; Beukema 1990).
The relatively constant community structure at the offshore station OG7 is contrary to the results of Schroeder (2003) who found the most evident changes in community structure after the cold winter 1995/1996 at Rachor’s station WB in the Oyster Ground. But these changes were based mainly on a highly successful recruitment of echinoderms in 1996, which is known to be patchy (Òlafsson et al. 1994; Thiébaut et al. 1998), and not by a decrease in abundance or number of species. Highly successful recruitment after cold winters was found for several benthic species (Dörjes et al. 1986; Beukema et al. 2001; Kröncke et al. 2001; Strasser et al. 2003). However, since most benthic species disperse via pelagic larvae, local recruitment is often decoupled from local reproduction (Bosselmann 1991; Caley et al. 1996; Eckert 2003).
Other environmental parameters
The drastic change in community structure and the decrease of species sensitive to cold temperatures at the German Bight stations after the ice winter 1995/1996 can be clearly related to the extreme winter temperatures. Nevertheless, long-term changes of benthic communities can be caused by various abiotic and biotic parameters. Most of these parameters such as changes in food supply due to sedimenting primary production, changes of the hydrographic regime or eutrophication result in more or less gradual changes of benthic communities (Frid and Hall 2001; Kröncke et al. 2001).
Hypoxia can lead to changes of benthic communities, which are similar to those caused by extreme cold winters. However, periods of hypoxia in the German Bight were mainly detected in the 1980s, possibly enhanced by eutrophication and hydrographic conditions. Hypoxia, for instance, led to an increased mortality of the benthic fauna in the German Bight in the years 1981–1983 (Dyer et al. 1983; von Westernhagen et al. 1986; Niermann 1990). During the 1990s oxygen concentrations below 4 mg/l were recorded only in 1994 (van Beusekom et al. 2003). In contrast to the changes in benthic communities recorded in this study, which persisted for at least 2 years, changes after severe hypoxia events in the 1980s were only short-lived and communities recovered quickly due to the survival of most species in adjacent areas (van Beusekom et al. 2003). Nevertheless, severe winters can lead to an accumulation of dead biomass in the sediment, which in turn can enhance the development of hypoxia events besides other factors such as primary production and water temperature (Günther and Niesel 1999; Böttcher 2003). However, since in this study sampling took place always in spring, the effects of hypoxia on communities, which normally happen during late summer, would be detected in the following year at the earliest.
Another factor, which can severely affect benthic communities is the disturbance of the seabed due to fishing activities. It is supposed to have caused a long-term shift of benthic communities from long-lived to more opportunistic species in many areas of the North Sea (Craeymeersch 1994; van Santbrink and Bergman 1994; Kaiser and Spencer 1996; Rumohr and Kujawski 2000; Frid and Hall 2001). The German Bight is one of the most intensively fished areas of the North Sea, where mainly beam trawling is carried out (Rijnsdorp et al. 1998; Reiss and Kröncke 2004). The drastic changes of benthic communities in the German Bight after 1995 did probably not result from an increase in fishing effort. The station GB2 is situated within the “plaice box”, a partially closed area for large beam trawlers established in 1989, whereas the station GB5 is situated outside the box. Thus, fishing effort probably differed markedly between these two stations. Nevertheless, the community changes at stations GB2 and GB5 were significantly similar, pointing to a large-scale impact.
References
Becker GA, Dick S, Dippner JW (1992) The hydrography of the german bight. Mar Ecol Prog Ser 91:9–18
Beukema JJ (1990) Expected effects of changes in winter temperatures on benthic animals living in soft sediments in coastal North Sea areas. In: Beukema JJ (ed) Expected effects of climatic change on marine coastal ecosystems. Kluwer Academic Publisher, Dordrecht, MA
Beukema JJ (1992) Expected changes in the Wadden Sea benthos in a warmer world: lessons from periods with mild winters. Neth J Sea Res 30:73–79
Beukema JJ, Dekker R, Essink K, Michaelis H (2001) Synchronized reproductive success of the main bivalve species in the Wadden Sea: causes and consequences. Mar Ecol Prog Ser 211:143–155
Beukema JJ, Essink K, Michaelis H (1996) The geographic scale of synchronized patterns in zoobenthos populations as a key underlying factors: climatic or man-induced. ICES J Mar Sci 53:964–971
Bosselmann A (1991) Recruitment and postlarval growth of some macrozoobenthos species in the German Bight. Meeresforschung 33:141–158
Böttcher ME (2003) Schwarze Flecken und Flächen im Wattenmeer. In: Lozán JL, Rachor E, Reise K, Sündermann J, von Westernhagen H (eds) Warnsignale aus Nordsee und Wattenmeer. Wissenschaftliche Auswertungen, Hamburg
Buchanan JB, Moore JJ (1986) Long-term studies at a benthic station off the coast off Northumberland. Hydrobiologia 142:121–127
Buhr K-J (1981) Auswirkungen des kalten Winters 1978/79 auf das Makrobenthos der Lanice-Siedlung im Weser-Ästuar. Ver Inst Meeresforsch Bremerh 19:115–131
Caley MJ, Carr MH, Hixon MA, Hughes TP, Jones GP, Menge BA (1996) Recruitment and the local dynamics of open marine populations. Ann Rev Ecol Syst 27:477–500
Clarke KR, Warwick RM (1994) Change in marine communities: an approach to statistical analysis and interpretation. Plymouth Marine Laboratory, UK
Craeymeersch JA (1994) Effects of a 4m beam trawl on the benthic fauna of the Belgian continental shelf (Flemish Banks). In: de Groot SJ, Lindeboom HJ (eds) Environmental impact of bottom gears on benthic fauna in relation to natural resources management and protection of the North Sea. NIOZ-Rapport 1994 -11 Rivo-DLO report C026/94
Dippner JW, Ikauniece A (2001) Long-term zoobenthos variability in the Gulf of Riga in relation to climate variability. J Mar Syst 30:155–164
Dörjes J, Michaelis H, Rhode B (1986) Long term studies of macrozoobenthos in intertidal and shallow subtidal habitats near the island of Norderney (East Frisian coast, Germany). Hydrobiologia 142:217–232
Dyer MF, Fry WG, Fry PD, Cranmer GJ (1983) Benthic regions within the North Sea. J Mar Biol Assoc UK 63:683–693
Eckert GL (2003) Effects of the planktonic period on marine population fluctuations. Ecology 84:372–383
Frid C, Hall S (2001) Ecological quality objectives for benthic communities: if we protect the habitat do we need to do more. ICES CM 2001/T:06:15
Günther C-P, Niesel V (1999) Effects of the ice winter 1995/96. In: Dittmann S (ed) The Wadden Sea ecosystem. Springer, Berlin
Hagmeier A (1925) Vorläufiger Bericht über die vorbereitenden Untersuchungen der Bodenfauna der Deutschen Bucht mit dem Petersen-Bodengreifer. Ber Dt Wiss Komm Meeresforsch B. NF 1:247–272
Hedges JI, Stern JH (1984) Carbon and nitrogen determination of carbonate-containing solids. Limnol Oceanogr 29:657–663
Heip C, Basford D, Craeymeersch J, Dewarumez JM, Dörjes J, de Wilde P, Duineveld G, Eleftheriou A, Herman PMJ, Niermann U, Kingston P, Künitzer A, Rachor E, Rumohr H, Soetaert K, Soltwedel T (1992) Trends in biomass, density and diversity of North Sea macrofauna. ICES J Mar Sci 49:13–22
Hurlbert SH (1971) The non-concept of species diversity: a critique and alternative parameters. Ecology 52:577–586
Hurrell JW (1995) Decadal trends in the North Atlantic oscillation: regional temperatures and precipitation. Science 269:676–679
Jones SE, Jago CF, Bale AJ, Chapman D, Howland RJM, Jackson J (1998) Aggregation and resuspension of suspended particulate matter at a seasonally stratified site in the southern North Sea: physical and biological controls. Cont Shelf Res 18:1283–1309
Kaiser MJ, Spencer BE (1996) The effects of beam-trawl disturbance on infaunal communities in different habitats. J Anim Ecol 65:348–358
Kröncke I (1992) Macrofauna standing stock of the Dogger Bank. A comparison III. 1950–1954 versus 1985–1987. A final summary. Helgol Meeresunters 46:137–169
Kröncke I, Dippner JW, Heyen H, Zeiss B (1998) Long-term changes in macrofaunal communities off Norderney (East Frisia, Germany) in relation to climate variability. Mar Ecol Prog Ser 167:25–36
Kröncke I, Rachor E (1992) Macrofauna investigations along a transect from the inner German Bight towards the Dogger Bank. Mar Ecol Prog Ser 91:269–276
Kröncke I, Stoeck T, Wieking G, Palojärvi A (2004) Relationship between structural and trophic aspects of microbial and macrofaunal communities in different areas of the North Sea. Mar Ecol Prog Ser 282:13–31
Kröncke I, Zeiss B, Rensing C (2001) Long-term variability in macrofauna species composition off the island of Norderney (East Frisia, Germany) in relation to changes in climatic and environmental condition. Senckenb Marit 31:65–82
Künitzer A, Basford D, Craeymeersch JA, Dewarumez JM, Dörjes J, Duineveld GCA, Eeleftheriou A, Heip C, Herman P, Kingston P, Niermann U, Rachor E, Rumohr H, de Wilde PAJ (1992) The benthic infauna of the North Sea: species distribution and assemblages. ICES J Mar Sci 49:127–143
Niermann U (1990) Makrobenthos der südöstlichen Nordsee: Fluktuationen in den Jahren 1984–1988. PhD, University of Hamburg, Germany
Òlafsson EB, Peterson CH, Ambrose WGJ (1994) Does recruitment limitation structure populations and communities of macroinvertebrates in marine soft sediments: the relative significance of pre- and post-settlement processes. Oceanogr Mar Biol Ann Rev 32:65–109
Otto L, Zimmerman JTF, Furnes GK, Mork M, Saetre R, Becker G (1990) Review of the physical oceanography of the North Sea. Neth J Sea Res 26:161–238
Planque B, Taylor AH (1998) Long-term changes in zooplankton and the climate of the North Atlantic. ICES J Mar Sci 55:644–654
Rachor E (1977) Faunenverarmung in einem Schlickgebiet in der Nähe Helgolands. Helgol wiss Meeresunters 30:633–651
Rachor E (1980) The inner German Bight—an ecologically sensitive area as indicated by the bottom fauna. Helgol Meeresunters 33:522–530
Rachor E, Nehmer P (2003) Erfassung und Bewertung ökologisch wertvoller Lebensräume in der Nordsee, Bundesamt für Naturschutz
Reichert A, Dörjes J (1980) Die Bodenfauna des Crildumersieler Watts (Jade, Nordsee) und ihre Veränderung nach dem Eiswinter 1978/79. Senckenb Marit 12:213–245
Reid PC, Edwards M (2001) Long-term changes in the fishery, pelagos and benthos of the North Sea. Senckenb Marit 31:107–115
Reid PC, Edwards M, Hunt HG, Warner J (1998) Phytoplankton change in the North Atlantic. Nature 391:546
Reiss H, Kröncke I (2004) Seasonal variability of epibenthic communities in different areas of the southern North Sea. ICES J Mar Sci 61:882–905
Reiss H, Kröncke I (2005) Seasonal variability of infaunal community structures in three areas of the North Sea under different environmental conditions. East Coast Shelf Sci 65:253–274
Rijnsdorp AD, Buys AM, Storbeck F, Visser EG (1998) Micro-scale distribution of beam trawl effort in the southern North Sea between 1993 and 1996 in relation to the trawling frequency of the sea bed and the impact on benthic organisms. ICES J Mar Sci 55:403–419
Rumohr H, Kujawski T (2000) The impact of trawl fishery on the epifauna of the southern North Sea. ICES J Mar Sci 57:1389–1394
Salzwedel H, Rachor E, Gerdes D (1985) Benthic macrofauna communities in the German Bight. Ver Inst Meeresforsch Bremerh 20:199–267
Schroeder A (2003) Community dynamics and development of soft bottom macrozoobenthos in the German Bight (North Sea) 1969–2000. PhD, University of Bremen, Germany
Stebbing ARD, Dethlefsen V (1992) Introduction to the Bremerhaven workshop on biological effects of contaminants. Mar Ecol Prog Ser 91:1–8
Strasser M, Dekker R, Essink K, Günther C-P, Jaklin S, Kröncke I, Madsen PB, Michaelis H, Vedel G (2003) How predictable is high bivalve recruitment in the Wadden Sea after severe winter? J Sea Res 49:47–57
Strasser M, Pieloth U (2001) Recolonization pattern of the polychaete Lanice conchilega on an intertidal sand flat following the severe winter of 1995/96. Helg Mar Res 55:176–181
Thiébaut E, Lagadeuc Y, Olivier F, Dauvin JC, Retière C (1998) Do hydrodynamic factors affect the recruitment of marine invertebrates in a macrotidal area? Hydrobiologia 375/376:165–176
van Beusekom J, Brockmann U, Elbrächter M, Pätsch J, Wiltshire K (2003) Die Bedeutung und die Gefahr von Algenblüten in Wattenmeer und Nordsee. In: Lozán JL, Rachor E, Reise K, Sündermann J, von Westernhagen H (eds) Warnsignale aus Nordsee und Wattenmeer. Wissenschaftliche Auswertungen, Hamburg
van Santbrink JW, Bergman MJN (1994) Direct effects of beam trawling on macrofauna in a soft bottom area in the southern North Sea. In: de Groot SJ, Lindeboom HJ (eds) Environmental impact of bottom gears on benthic fauna in relation to natural resources management and protection of the North Sea. NIOZ-Rapport 1994 -11 Rivo-DLO report C026/94
von Westernhagen H, Hickel W, Bauerfeind E, Niermann U, Kröncke I (1986) Sources and effects of oxygen deficiencies in the south-eastern North Sea. Ophelia 26:457–473
Wieking G, Kröncke I (2001) Decadal changes in macrofauna communities on the Dogger Bank caused by large-scale climate variability. Senckenb Marit 31:125–141
Wieking G, Kröncke I (2003) Macrofauna communities of the Dogger Bank (central North Sea) in the late 1990s: spatial distribution, species composition and trophic structure. Helg Mar Res 57:34–46
Ziegelmeier E (1964) Einwirkungen des kalten Winters 1962/63 auf das Makrobenthos im Ostteil der Deutschen Bucht. Helgol wiss Meeresunters 10:276–282
Ziegelmeier E (1970) Über das Massenvorkommen verschiedener makrobenthaler Wirbelloser während der Wiederbesiedlungsphase nach Schädigung durch “katastrophale” Umwelteinflüsse. Helgol wiss Meeresunters 21:9–20
Acknowledgements
We thank the captains and crews of the RV “Senckenberg” for their help with sampling throughout the years. We are also grateful to Christa Rensing, students and people doing their community service, for their help at sea and with sorting samples. The Federal Maritime and Hydrographic Agency, Hamburg, are acknowledged for providing temperature data.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.D. Franke
Rights and permissions
About this article
Cite this article
Reiss, H., Meybohm, K. & Kröncke, I. Cold winter effects on benthic macrofauna communities in near- and offshore regions of the North Sea. Helgol Mar Res 60, 224–238 (2006). https://doi.org/10.1007/s10152-006-0038-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-006-0038-3