
Abstract
The parasite communities of the rabbitfish, Siganus rivulatus, were used to track multi-annual changes in the northern Gulf of Aqaba, Red Sea, in an environment subjected to ongoing anthropogenic impact. Parasitological data from these fish were collected from 1998 to 2000, with spring and fall samplings at three locations: at a coral reef (OBS), at a sandy beach area (NB) and at a mariculture cage farm (FF). These data were compared with data from 1995–1997 as well as data collected during 1981–1985 at the coral reef sampling site. The data analyses indicate that the ratio between heteroxenous and monoxenous parasite species declined significantly at all sites between 1995–1997 and 1998–2000. During the same period, the species richness of monoxenous parasites increased significantly at all sites. The species richness of heteroxenous parasites decreased significantly at the coral reef site, but remained steady at the other two sites. This coincided with a significant increase in the prevalence of monogeneans at the OBS and FF sites and a significant decrease in the prevalence of digeneans at the FF and NB sites. The decline in the abundance of the latter, specifically of Opisthogonoporoides sp. and Gyliauchen sp., was even more significant when compared with the 1981–1985 data. The prevalence of other gut helminths, namely the digenean Hexangium sigani and the nematodes Cucullanus sigani and Procamallanus elatensis, however, showed a significant increase over the same period. Analysis of the species richness and diversity indices of the parasite communities did not reveal conspicuous differences. These, however, did become apparent when heteroxenous and monoxenous members of particular taxa were analyzed separately. Therefore, when using parasite assemblages to detect ecological changes, it is essential to analyze not only at the community level, but also to consider separate components of particular parasitic groups.
Similar content being viewed by others


Introduction
Fish parasites with complex life histories have been used as a tool for the determination of geographical boundaries (Koie 1983, 1984; Bamber and Henderson 1985), migration routes (MacKenzie 1987; Diamant 1989b) breeding territories (MacKenzie1987) and predation behavior (Khan and Kiceniuk, 1988). The use of fish parasites as bioindicators and tags has led to their utilization as monitors of perturbations in aquatic ecosystems. While physical, chemical and even biological oceanographic parameters have constraints in time and space, fish parasites integrate and accumulate adverse effects of complex and variable stresses that influence them as well the other components of the ecosystem. Therefore, parasite community indices should arguably present an accurate and comprehensive picture on the state of an ecosystem. Various species of parasites and the parasite community as a whole have been proposed as bioindicators for environmental pollution. Changes in the prevalence and intensity of infection in certain parasite taxa (Khan 1987; Khan and Kiceniuk 1988; Khan and Payne 1997; Margolis and Cone 1997; Yeomans et al. 1997), as well as changes in the composition of parasite communities (D'Amelio and Gerasi 1997; Gelnar et al. 1997; Valtonen et al. 1997; Broeg et al 1999; Diamant et al. 1999) have been used to estimate anthropogenic impacts on aquatic environments. Relatively few studies have, however, addressed long-term changes in fish parasite communities as bioindicators for ecosystem stress (Kennedy 1985, 1993, 1997a; Koskivaara and Valtonen 1992; Kennedy et al. 1994). In addition to indices derived from population structure and diversity, the ratio between heteroxenous and monoxenous parasite species (Sh/Sm) was introduced as a parameter for adverse changes in the state of the environment (D'Amelio and Gerasi 1997; Diamant et al. 1999).
In the last two decades, the northern Gulf of Aqaba has been undergoing accelerated environmental degradation which has been associated with adverse anthropogenic impacts of urban and port development, tourism and mariculture (Fishelson 1995; Schuhmacher et al. 1995). However, this association was mostly circumstantial and not backed with biological studies. The rabbitfish (Siganus rivulatus) is one of the most conspicuous and abundant components of the coastal fish communities in the northern Red Sea. This fish was therefore chosen as a model for studies using parasites as bioindicators for monitoring the extent of anthropogenic impact on habitats in the Gulf of Aqaba (Banet 1998; Diamant et al. 1999). Initial studies on the parasites of this fish were carried in the mid 1980s and later, since 1995 (Diamant 1985, 1989a, 1989b; Diamant and Paperna 1986; Banet 1998; Diamant et al. 1999).
In the present communication, we analyze multi-annual changes in the rabbitfish parasite communities, from 1995 to 2000, compare them with those of the mid 1980s and discuss the possible causes for these changes.
Methods
Location and sampling design
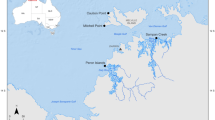
Parasitological data on S. rivulatus were collected from 1995 to 2000 at three sites along the Israeli northern part of the Gulf of Aqaba, Red Sea: FF, at the periphery of an offshore mariculture cage farm; NB, at the north-western, sandy, tourist beach next to the hotel district of Eilat, and OBS, in the western coral fringed zone (a nature reserve) near the Eilat underwater observatory (Fig. 1). Data from 1981 to 1985 were obtained from fish collected at OBS (Diamant 1985).

Map and coordinates of sampling sites at the northern Gulf of Aqaba. NB, North Beach; FF, fish farm; OBS, observatory
Parasitological data processing
Fish were trapped and kept alive in seawater tanks until they were dissected for parasitological examination (within 48 h of capture). Parasites were identified and quantified according to the protocol described in Diamant et al. (1999).
Statistical analysis
Parasite data were entered into a Microsoft Excel database, which was established to include data collected from each fish caught at the three sampling sites, and were used to calculate the parasitological indices (Diamant et al. 1999). "True" species richness was extrapolated by Y=a(1–e–bx)/b (Walter et al 1995) separately for monoxenous and heteroxenous species in order to detect differences in abundance trends between these two parasite groups. The calculation of "true" species richness and its confidence limits were done using SlideWrite Plus software. The effects of all parameters were tested using one-way ANOVA (after verifying a normal distribution). Differences between index values of the three sampling sites were compared using Tukey-Kramer multiple range test. Differences in prevalence and the relative representation of major parasite taxa were tested with χ 2 tests. All statistical procedures were carried out with JMP statistical software with the significance level set at P<0.05.
Results
From 1995 to 2000, a total of 761 fish were sampled. The length of the fish ranged from 150 to 250 mm (all specimens outside of this range were excluded from the analyses). Although the fish sampled from 1995 to 1997 were larger than those sampled from 1998 to 2000, these differences did not exceed 2 cm at any site and they have no implication regarding the fish size/age class. Condition factor values (K=100*weight/length3, see Bolger and Connolly 1989) of the fish samples from OBS from 1981 to 1985 were significantly higher than those sampled from 1995 to 2000 at all sites (Table 1). No change in the number of parasite species was noted between the 1995–1997 and 1998–2000 samples nor between the number of gut helminth species between 1981 and 1985, and between 1995 and 2000. When fish parasite communities from 1995–1997 and 1998–2000 were compared, the prevalence of infection with (heteroxenous) digeneans showed an overall decrease from 1995–1997 to 1998–2000 at FF and NB but no change at OBS. The prevalence of (monoxenous) monogeneans, on the other hand, increased significantly at FF and OBS. At the same time, a significant decrease in the prevalence of myxosporeans at OBS was also noted (Fig. 2).

Prevalence of major parasite taxa in 1995–1997 and 1998–2000 rabbitfish samples at the Red Sea sampling sites. NB, North Beach; FF, fish farm; OBS, observatory
The relative representation of members of the major parasite taxa, in relation to the whole parasite communities, in each sampling site is shown in Table 2. At OBS, the representation of each taxon was relatively uniform and no significant shifts occurred between the years 1995–1997 and 1998–2000. At FF and NB, monogenean and myxosporean infections were the most abundant. There was a significant decrease in the presence of digeneans at FF as well as a trend, though not significant, towards an increase in the abundance of nematode infections at FF and acanthocephalan infections at NB.
The mean values of parasite species richness in the 1998–2000 samples showed no significant changes from those of 1995–1997 at any of the three sites (Fig. 3).

Comparison of parasite species richness between 1995–1997 and 1998–2000 at the Red Sea sampling sites. NB, North Beach; FF, fish farm; OBS, observatory. S=no. of parasite species; D=(S–1)/ln(N) (Margalef 1951) a = S values, b = D values
The Sh/Sm ratio decreased significantly at all three sampling sites (Fig. 4a). Species richness, plotted separately for heteroxenous and for monoxenous parasites of 1995–1997 vs. 1998–2000 demonstrated a significant increase in monoxenous parasite species richness at all three sampling sites, with no change in heteroxenous parasites at FF and NB and a significant decrease at OBS (Fig. 4b, c).

Comparison of mean Sh/Sm (a), monoxenous (b) and heteroxenous (c) parasite species richness between 1995–1997 and 1998–2000 at the Red Sea sampling sites. NB, North Beach; FF, fish farm; OBS, observatory
"True" species richness curves for all parasite species did not vary significantly from 1995–1997 to 1998–2000. Monoxenous parasite "true" species richness curves, on the other hand, increased significantly in all three sampling sites (Fig. 5). Similar curves plotted for heteroxenous parasites revealed some decline, but this trend was not statistically significant (Fig. 6).

Cumulative curves of monoxenous parasites "true" species richness extrapolated by Y=a(1–e–bx)/b (Walther et al. 1995) between 1995–1997 and 1998–2000 in each of the Red Sea sampling sites. FF, fish farm (a); NB, North Beach (b); OBS, observatory (c). Significant differences in the values of variables a, b at OBS and b at NB)

Comparison of heteroxenous parasite "true" species richness extrapolated by Y=a(1–e–bx)/b (Walther et al. 1995) between 1995–1997 and 1998–2000 in each sampling site. FF, fish farm (a); NB, North Beach (b); OBS, observatory (c) with no significant differences in the values of variables a, b
Shannon-Weiner diversity values increased significantly at OBS and the Janion index of evenness increased significantly at FF when comparing 1995–1997 with 1998–2000, while values of the same indices from the other sampling sites showed no significant change (Fig. 7).

Comparison of diversity index values between 1995–1997 and 1998–2000 at the Red Sea sampling sites. NB, North Beach; FF, fish farm; OBS, observatory. a: Diversity H′=–Σ(PilnPi); b: evenness J=H′/Hmax ( where Hmax=ln[S])
Fish sampled at OBS during 1995–2000 (1990s), when compared with those from 1981–1985 (1980s) showed a significant decline in the abundance of two gut digeneans, Opisthogonoporoides sp. and Gyliauchen sp. (Figs. 8, 9). Their mean intensity (number of worms per infected fish) decreased significantly over this period, from 28.1 to 4.6 and 27.2 to 1.5, respectively. When the prevalence of these two species was considered, Opisthogonoporoides sp. displayed a minor, albeit consistent over time, decrease (49% to 43.5%) while Gyliauchen sp. showed a dramatic decrease, from 36.3% to only 1.7%.

Comparison of prevalence (a) and intensity (b) of gut helminth infections between the 1990s (1995–2000) and 1980s (1981–1985) at OBS. Abbreviations: Opist, Opisthogonoporoides sp.; Hexan, Hexangium sigani; Gylia, Gyliauchen sp.; Procam, Procamallanus elatensis; Cucu Cucullanus sigani; Scler Sclerocollum rubrimaris

Changes in the mean intensity of infection of Opisthogonoporoides sp. and Gyliauchen sp. over time
On the other hand, the prevalence of other rabbitfish gut helminths, namely the digenean Hexangium sigani and the nematodes Cucullanus sigani and Procamallanus elatensis, showed a significant increase over the last 15 years (Fig. 8b).
The model demonstrated that the sampling effort required for reaching the "true" species richness of the habitat was considerably lower during the 1980s than in the recent samplings (Fig. 10).

Comparison of parasites "true" species richness extrapolated by Y=[a*(1–e–bx)]/b (Walther et al. 1995) of S. rivulatus gut helminths between 1981–1985 and 1995–2000 at OBS (values of variables a and b are significantly different)
Discussion
The Gulf of Aqaba is fringed by a highly developed, diverse coral reef, which supports a broad variety of marine life. The existence of this delicate, complex ecosystem depends on the stability of the oligotrophic conditions, which have typically prevailed in this water body (Fishelson 1995). The Gulf of Aqaba was a near-pristine area when the city of Eilat was founded in 1949. In recent years, studies have provided evidence of coastal degradation in the region (Fishelson 1995; Schuhmacher et al. 1995). There are three main sources of organic pollution for the reported increased eutrophication in the gulf: urban and maritime sewage, mariculture cage farms and phosphate loading berths (Silverman et al. 2001). Abelson et al. (1999) demonstrated that particulate matter from point sources at Eilat's North Beach reached the remote coral reefs to the south, both as bed-load and suspension-load particles. Such shifting of organic matter with currents may promote phytoplankton blooms and a corresponding increase in zooplankton and zoobenthos, such as copepods, polychaetes and other species, all potential intermediate hosts of fish parasites. However, there is no direct quantitative evidence to confirm the above predictions.
The most important change between 1995–1997 and 1998–2000 is probably the significant decline in the heteroxenous/monoxenous parasite species ratios. While monoxenous parasite abundance increased at all sampling stations, heteroxenous species showed a marked decline only at OBS. This trend was unmistakably demonstrated by the cumulative model that extrapolates "true" species richness of the habitat as a function of sample size (Walther et al. 1995). Our basic assumption was that in a disturbed environment heteroxenous parasites are less likely than monoxenous parasites to complete their life cycles, either due to direct adverse effects on their free living stages, or as an indirect consequence of the elimination of their intermediate hosts (Paperna 1997). Previous studies have proposed and applied Sh/Sm as a tool for monitoring the state of an ecosystem (D'Amelio and Gerasi 1997; Diamant et al. 1999). The transition with time in the relationship between heteroxenous and monoxenous parasites, with a more conspicuous increase in monoxenous species abundance than the decline in heteroxenous species, agrees with the reports of Khan and Thulin (1991), Gelnar et al. (1997) and Bagg and Valtonen (1996) which associate an increase in monoxenous species abundance, monogeneans in particular, with increased perturbation of the habitat. Chronic exposure to organic pollution and contaminated sediment has been shown also to enhance monoxenous ciliate infections, in particular trichodinids (Yeomans 1997; Broeg et al. 1999). The suggested indirect effect (physiological or immunosuppressive) on the hosts under such circumstances (Khan and Thulin 1991) still warrants experimental validation.
We found between 1995–1997 and 1998–2000 a consistent decrease in the abundance of digenetic trematodes at FF and NB as well as their relative representation in the community at FF. Similarly, the digeneans Gyliauchen sp. and Opisthogonoporoides sp. declined significantly in abundance between the 1980s and 1990s. The abundance of other gut helminths—the digenean H. sigani and the nematodes C. sigani and P. elatensis—increased, while the abundance of the acanthocephalan Sclerocollum rubrimaris maintained steady levels. The life cycle details of all these helminths are as yet unknown, and we may consider them only in broad terms, i.e. implicating molluscs as intermediate hosts of digeneans and amphipods, copepods and possibly also annelids for nematodes and acanthocephalans. S. rivulatus apparently becomes infected when ingesting these and other small invertebrates associated with macroalgae or sea-grass (Lundsberg and Lipkin 1979; Diamant 1989a).
The observed decline in the populations of the digeneans Gyliauchen sp. and Opisthogonoporoides sp. could be linked to the decline in the abundance of molluscs in the reef habitat (see Fishelson 1995). The observed increase in the digenean H. sigani and the two nematode species could be attributed to a possible rise in the abundance of benthic or planktonic invertebrates acting as intermediate hosts. In summary, the remarks of Kennedy (1997b) are as relevant as ever and well worth repeating here. At present, we lack knowledge on parasite life histories, on the effect of different sources of pollution on each stage of their life cycle and on the effects on their intermediate hosts. What we can do is record changes and differences in populations and communities; but we are still incapable of unequivocally relating them to any specific causal factors.
Extreme conditions may have different, sometimes even conflicting impacts on diverse taxa of parasites. Under such circumstances, overall community indices may incorporate opposite trends and possibly fail to track transitions that have actually taken place in the composition of the community. Accordingly, species richness and diversity indices of the complete parasite communities demonstrated only some differences between the 1995–1997 and 1998–2000 sampling periods. The present results are in agreement with those of Gelnar et al. (1997), who investigated the diversity of freshwater fish parasites in relation to pollution. These authors showed a succession of species with no change in diversity indices, a significant decrease in the community's species richness. Therefore, we emphasize that in order to pinpoint ecological changes, it is essential to analyze parasite assemblages not only at the community level, but also to carefully consider components of particular parasitic groups.
References
Abelson A, Shteinman B, Fine M, Kaganovski S (1999) Mass transport from pollution sources to remote coral reef in Eilat (Gulf of Aqaba, Red Sea). Mar Pollut Bull 38:25–29
Bagg A, Valtonen ET (1996) Experimental study on the influence of paper and pulp mill effluent on the gill parasite communities of roach (Rutilus rutilus). Parasitology 112:499–508
Bamber RN, Henderson PA (1985) Diplostomiasis in sand smelt Atherina presbyter Cuvier from the fleet, Dorset and its use as population indicator. J Fish Biol 26:223–229
Banet A (1998) Parasite community structure of the rabbitfish Siganus rivulatus as an early warning biomarker for ecological degradation of marine habitats in the Gulf of Aqaba, (Hebrew text, English summary). MSc thesis, Faculty of Agriculture, Hebrew University of Jerusalem, Jerusalem
Bolger T, Connolly PL (1989) The selection of suitable indices for the measurements and analysis of fish condition. J Fish Biol 34:171–182
Broeg K, Zander S, Diamant A, Koerting W, Kruener G, Paperna I, Westernhagen H von (1999) The use of fish metabolic, pathological and parasitological indices in pollution monitoring. I. North Sea. Helgol Mar Res 53:171–194
D'Amelio S, Gerasi L (1997) Evaluation of environmental deterioration by analysing fish parasite biodiversity and community structure. Parassitologia 39:237–242
Diamant A (1985) Biology of the parasites of Siganus spp.(Teleosti:Siganidae) from the northern Red Sea and eastern Mediterranean coasts of Israel (in Hebrew, English Summery). PhD thesis, Hebrew University of Jerusalem, Jerusalem
Diamant A (1989a) Ecology of the acanthocephlan Sclerocollum rubrimaris Schmidt and Paperna (1978) (Rhadinorhynchidae:Gorgorhynchinae) from wild populations of rabbitfish (genus Siganus) in the northern Red Sea. J Fish Biol 34:387–398
Diamant A (1989b) Lessepsian migrants as hosts: a parasitological assessment of rabbitfish Siganus luridus and Siganus rivulatus (Siganidae) in their original and new zoogeographical regions. In: Spanier E, Steinberger Y, Luria M (eds) Environmental quality and ecosystems stability, IV–B. Environmental quality. ISEEQS, Jerusalem, pp 187–194
Diamant A, Paperna I (1986) The parasites of wild Red Sea rabbitfish Siganus spp. (Pisces: Siganidae) as potential pathogens in mariculture. In:Vivares CP, Bonami J-R, Jaspers E (eds) Pathology in marine aquaculture. Eur Aqua Soc Spec Publ 9, Bredene, Belgium, pp 71–83
Diamant A, Banet A, Paperna I, Westernhagen H von, Broeg K, Kruener G, Koerting W, Zander S (1999) The use of fish metabolic, pathological and parasitological indices in pollution monitoring. II. The Red Sea and Mediterranean. Helgol Mar Res 53:195–208
Fishelson L (1995) Elat (Gulf of Aqaba) littoral-life on the red line of degradation. Isr J Zool 41:43–55
Gelnar M, Sebelova S, Dusek L, Koubkova B, Jurajda P, Zahradkova S (1997) Biodiversity of parasites in freshwater environment in relation to pollution. Parassitologia 39:189–200
Kennedy CR (1985) Interaction of fish and parasite populations: to perpetuate or pioneer? In: Rollinson D, Anderson RM (eds) Ecology and genetics of host-parasite interactions. Linn Soc Symp Ser 11:1–20
Kennedy CR (1993) The dynamics of intestinal helminth communities in eels Anguilla anguilla in a small stream: long-term changes in richness and structure. Parasitology 107:71–78
Kennedy CR (1997a) Long-term and seasonal changes in composition and richness of intestinal helminth communities in eels Anguilla anguilla of an isolated English river. Folia Parasitol 44:27–273
Kennedy CR (1997b) Freshwater fish parasites and environmental quality: an overview and caution. Parassitologia 39:249–254
Kennedy CR, Wyatt RJ, Starr K (1994) The decline and natural recovery of an unmanaged coarse fishery in relation to changes in land use and attendant eutrophication. In: Cowx IG (ed) Rehabilitation of fresh water fisheries. Fishing News Books, Oxford, pp 366–375
Khan RA (1987) Effects of chronic exposure to petroleum hydrocarbons on two species of marine fish infected with hemoprotozoan Trypanosoma murmanensis. Can J Zool 65:2703–2709
Khan RA, Kiceniuk JV (1988) Effect of petroleum aromatic hydrocarbons on monogeneids parasitizing Atlantic cod, Gadus morhua (L.). Bull Environ Contam Toxicol 41:94–100
Khan RA, Payne JF (1997) A multidisciplinary approach using several biomarkers, including a parasite, as indicators of pollution: a case history from a paper mill in Newfoundland. Parassitologia 39:183–188
Khan RA, Thulin J (1991) Influence of pollution on parasite of aquatic animals. Adv Parasitol 30:201–238
Koie M (1983) Digenetic trematodes from Limanda limanda (L.) (Osteichthyes, Pleuronectidae) from Danish adjacent waters, with special reference to their life histories. Ophelia 22:201–228
Koie M (1984) Digenetic trematodes from Gadus morhua (L.) (Osteichthyes, Gadidae) from Danish adjacent waters, with special reference to their life histories. Ophelia 23:195–222
Koskivaara M, Valtonen ET (1992) Dactylogyrus (Monogenea) communities on the gills of roach in three lakes in central Finland. Parasitology 104:263–272
Lundberg B, Lipkin Y (1979) Natural food of the herbivorous rabbitfish (Siganus spp.) in the northern Red Sea. Bot Mar 22:173–181
MacKenzie K (1987) Long-term changes in the prevalence of two helminth parasites (Cestoda: Trypanorhyncha) infecting marine fish. J Fish Biol 31:83–88
Marcogliese DJ, Cone DK (1997) Parasite communities as indicators of ecosystem stress. Parassitologia 39:227–232
Margalef R (1958) Information theory in ecology. Gen Syst 3:36–71
Paperna I (1997) Fish parasites as indicators of environmental quality—introductory remarks. Parassitologia 39:168
Schuhmacher H, Kroll DK, Reinicke GB (1995) Long-term fluctuation of coral communities at Aqaba and on Sanguine-Atoll (northern and central Red Sea) over more than a decade. Beitr Palaeontol 20:89–97
Silverman J, Lazar B, Erez J (2001) A long-term metabolic performance record of the nature reserve reef in Eilat. Abstracts of the 33rd anniversary conference of the Heinz Steinitz Marine Biology Laboratory, Eilat pp 135–137
Valtonen ET, Holmes JC, Koskivaara M (1997) Eutrophication, pollution and fragmentation: effects on the parasite communities in roach (Rutilus rutilus) and perch (Perca fluviatilis) in four lakes in central Finland. Can J Aquat Fish Sci 54:572–585
Walther BA, Cotgreave P, Price RD, Gregory RD, Clayton DH (1995). Sampling effort and parasite species richness. Parasitol Today 11:306–310
Yeomans WE, Chubb JC, Sweeting RA (1997) Use of protozoan communities for pollution monitoring. Parassitologia 39:201–212
Acknowledgements
The field and laboratory assistance of Ayana Banet and Yariv Shtupler throughout different stages of this study is gratefully acknowledged. This study was supported by the German-Israeli Cooperation in Marine Science (MARS), Project Biological Indicators of Natural and Man-made Changes in Marine and Coastal Waters, BMBF-Code 03F0200A coordinated by G.K.S.S. Geesthacht and the Israel Ministry of Sciences and Arts.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H. von Westernhagen, A. Diamant
Rights and permissions
About this article
Cite this article
Dzikowski, R., Paperna, I. & Diamant, A. Multi-annual changes in the parasite communities of rabbitfish Siganus rivulatus (Siganidae) in the Gulf of Aqaba, Red Sea. Helgol Mar Res 57, 228–235 (2003). https://doi.org/10.1007/s10152-003-0139-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-003-0139-1