
Abstract
As a conservation strategy, the South African National Biodiversity Institute (SANBI) establishes biodiversity gardens in areas with unique vegetation types that are vulnerable to extinction. The study aimed to (1) determine the vegetation cover dynamics of the Free State National Botanical Garden (FSNBG) over a 30-year period (1987–2017), focusing on different vegetation classes; (2) evaluate the ecological integrity of the Critical Biodiversity Area 1 (CBA1) vegetation using species abundance and vegetation cover; and (3) quantify potential conservation threats that may be drivers of vegetation cover changes. The “moderate vegetation cover” and “dense vegetation cover” had increased by 25.1 ha and 8.6 ha respectively in the FSNBG. Woody vegetation cover expanded significantly over the past 30-year period, suggesting “bush” encroachment. Shannon–Wiener diversity indices showed high overall plant species diversity of CBA1 vegetation type (H = 3.5), with the vegetation cover remaining high (79.6 ± 15.9%), 50 plant species no longer existing, suggesting reduced taxonomic richness. Major conservation threats included the presence of 27 alien and invasive plant species interspersed within different vegetation patches and anthropogenic habitat fragmentation in the past 19 years (i.e. covering ~ 18% of the buffer zone). We conclude that increased vegetation cover is associated with bush encroachment and we recommend interventions to reduce the population density of woody plants and establish permanent vegetation monitoring plots.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Climate change has become a major threat to biodiversity in the present century in several parts of the world, with evidence of shifts in the distribution of many biological species, particularly plants (Karger et al. 2017; Pacifici et al. 2015; Randin et al. 2009; Guisan and Thuiller 2005; Thuiller et al. 2005). Plants are primary producers and indicators of habitat change (Del Vecchio et al. 2015, 2016; Cornelissen 2011), and thus, any significant change in the local environmental conditions can be discerned from plants’ responses (Pacifici et al. 2015; Niang et al. 2014; Stankovic et al. 2011; Randin et al. 2009; Smith 1994). In South Africa, Mackellar et al. (2014) reported a downward trend in seasonal and annual total rainfall for the period 1960 to 2010, and these were evident over the central and north-eastern parts of South Africa. Additionally, they recorded a significant increase in the number of rainy days in the southern Drakensberg in the spring and summer seasons (Mackellar et al. 2014). Consistently, Rutherford and Powrie (2013) noticed a decrease in plant species richness across the rangeland biomes of South Africa, which was attributed to the reduced rainfall. Niang et al. (2014) argued that the shift in the distribution of plant species in Africa may be the result of the corresponding micro-climatic changes. The capacity of plants in finding their new niches in response to climate change may be constrained by the limitations in their ability to move from across different places (Rutherford et al. 2000).
Bush encroachment leads to the reduced ecological integrity of keystone species that drive habitat functionality (Plumptre et al. 2021; Mogashoa et al. 2021; Steffen et al. 2009). For example, bush plant encroachment has been blamed for the decline in cover of grasses and forbs in the grassland biome (see Yapi et al. 2018; Archer et al. 2017; Matsika 2008; Neke and du Plessis 2004). Mogashoa et al. (2021) note a 53% decrease of grass species richness because of the increase in bush encroachment in the semi-arid savanna grassland of South Africa. Among the factors that cause bush encroachment is the reduced precipitation events and drought conditions (Marengo et al. 2017; Botai et al. 2016; van Rooyen 2010), which place more severe physiological stress on non-woody plant species (Ward 2005; Scholes and Archer 1997). Archer et al. (2017) and Ward (2010) both report that bush encroachment is partly due to the changed local climate regime with increasing Carbon dioxide (CO2) concentrations in the atmosphere, favouring C3 woody plant growth (Sintayehu 2018; Welz 2013; Buitenwerf et al. 2012). Little et al. (2015) show that intensive burning in the Steenkampsberg Plateau in Mpumalanga results in detrimental impacts on the diversity of plant species whereby vegetation structure is changed, and species are replaced. In the Hluhluwe-IMfolozi Nature Reserve, Case and Staver (2017) observed a drastic increase over 7 years in woody cover (i.e. medium trees of 2–4 m tall) (2007–2014), caused by inappropriate land use practices such as infrequent fires and high grazing pressure.
Several studies in South Africa have shown that plant invasion poses huge threats to natural vegetation within the grassland biome (O’Connor and van Wilgen 2020; Zengeya and Wilson 2020; Yapi et al. 2018; Masocha and Dube 2018). Newly introduced plant species can out-compete natural species because of rapid growth, rapid reproduction, and can spread aggressively, in the local habitat (Yapi et al. 2018; Wong et al. 2017; Lowe et al. 2012; Rejmanek and Richardson 1996). These introduced plants displace native vegetation through competition for resources such as water, nutrients, light and space (Iponga et al. 2008; Daehler 2003). Milton et al. (2007) reported that newly introduced fleshy-fruited plants can infiltrate networks of seed dispersal and pollination in the local environment, and thus disrupt reproduction and population fitness (Traveset and Richardson 2006). Other authors have similarly shown that once a dispersal system is disrupted through the introduction of new plant species, the savannas can be transformed as native trees are suppressed and over-topped, impacting crucial perch sites and foci for directed dispersal (see Mokotjomela et al. 2013; Iponga et al. 2008). Yapi et al. (2018) studied the impact of the invasive species Acacia mearnsii in the South African Montane Grassland ecosystem and found that the grazing capacity of the area was reduced by 72%, as A. mearnsii increased, thus negatively impacting agricultural production.
It has been shown that many protected areas in South Africa also struggle to manage pressure from the loss of natural habitat through human settlement encroachment in areas adjacent to those protected areas, thus reducing their genetic diversity and persistence (Das and Angadi 2022; Taylor and Atkinson 2012; Kiringe and Okello 2007; Rosenberg et al. 1997; Haddad et al. 2015). Those protected areas become isolated as habitat corridors are impacted, leading to limited ecological processes mediated by long-distance agents (Haddad et al. 2015; de Frutos et al. 2015; SANBI 2013; Baum et al. 2004; Carey et al. 2000). Rosenberg et al. (1997) indicated that habitat corridors in the protected areas help to increase population persistence by allowing the continued exchange of genetic material between the connected populations.
The Free State National Botanical Garden (FSNBG), where this current study was based, is located within the Dry Highveld Grassland bioregion of the Grassland Biome (Mucina and Rutherford 2006; Skowno et al. 2019), which is poorly protected, with only 1.52% under formal protection, while 31.51% has been completely transformed (Carbutt et al. 2011). The Dry Highveld Grassland is threatened by various activities, including overgrazing, cultivation, mining, acid mine drainage, quarrying and urban sprawl (SANBI 2013; Mucina and Rutherford 2006). The conservation priority vegetation types—also called the Critical Biodiversity Area 1 (CBA1)—that occur in the Dry Highveld Grassland bioregion recorded in FSNBG include Gh8 Bloemfontein Karroid Shrubland and the threatened Gh5 Bloemfontein Dry Grassland (Skowno et al. 2019; Mucina and Rutherford 2006). CBA1 contains biodiversity that is either irreplaceable or near irreplaceable for meeting national conservation targets as well as high species diversity (SANBI 2017). Consequently, it is critical to understand the vegetation dynamics in the FSNBG and to assess whether the vegetation units are affected by reported threats to the broader Grassland Biome, such as fire, fragmentation and invasive extra-limital plant species—“plant species occurring outside their native habitat/biome” (Matsika 2008; Neke and du Plessis 2004). This knowledge is critical to the development and revision of the adaptive vegetation management plans of the FSNBG.
Given these challenges, the aims of the present study were (1) to determine the vegetation cover dynamics over a 30-year period (1987–2017) in the FSNBG—it was predicted that plants and thus woody vegetation cover might expand because of climate change threats such as extreme droughts (Del Vecchio et al. 2015, 2016); (2) to evaluate the CBA1’s vegetation status (i.e. species and vegetation cover) by determining the abundance of the indicator plant species and vegetation cover and (3) to document the conservation threats that may be drivers of change in vegetation cover.
Methods and material
Study site
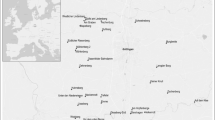
The study was conducted in the FSNBG (S 29°12′55.6″; E 26°12′41.3″), Free State Province, South Africa (Fig. 1). The FSNBG occurs in the Grassland Biome under the Dry Highveld bioregion (Mucina and Rutherford 2006; Dayaram et al 2019, SANBI 2018). The Botanical Garden covers an area of about 66.8 ha and is at an altitude between 1300 and 1400 m above sea level (Masilo 1999; Chaplin 1979), of which about 13.1 ha is transformed as a developed garden and infrastructure, 1.4 ha as national road and verges.

The location of the study area: Free State National Botanical Garden, Bloemfontein in South Africa
The vegetation structure is largely characterised by Gh 8 Bloemfontein Karroid Shrubland, Gh 5 Bloemfontein Dry Grassland, Gh 7 Winburg Grassy Shrubland and AZa 5 Highveld Alluvial Vegetation (Fig. 1; Mucina and Rutherford 2006). The Botanical Garden experiences an extremely variable continental climate, with minimum winter temperatures below 0 °C, the night temperature often falling below freezing, and frost in July. Fairly high summer temperatures frequently range between 30 and 35 °C in January (Haddad et al. 2019; Neethling and Haddad 2013). The mean annual rainfall is relatively low (584 mm year−1), with most rainfall recorded during the summer months and in early autumn.
Spatial data collection and image processing of vegetation
To understand the extent of vegetation-cover change over 30 years, remote sensing (RS) and geographical information system (GIS) imagery were used to analyse the spatial changes that occurred over 30 years (1987–2017) in 10-year intervals. The FSNBG is dominated by shrub and grass species, with a small portion of dense woodland. According to Hill et al. (2005), the natural expression of the growth, mortality and regeneration of trees and shrubs is measurable over a period of 10 years or more for significant trend detection, while that for grass species is measurable over a period of 5 years. High-resolution imagery was generated for the FSNBG region as an area of interest (AOI) and imported from a Google Earth shapefile in KML format (Yukhnovskyi and Zibtseva 2020). The Earth Observation System (EOS) land viewer satellite images had already undergone pre-processing to overcome image errors and remove distortions (EOS 2020). The scene search tool was used to select various satellite images of the AOI. Satellite imagery—from Thematic Mapper™ and the Operational Land Imager and Thermal Infrared Sensor (OLI/TIRS) —of the study area (path 171/row 80) was acquired for the years of interest: 1987, 1997, 2007 and 2017, with the 1987 image established as a baseline (Table 1; also see Carrasco et al. 2020 for a similar approach) (Yukhnovskyi and Zibtseva 2020; Peter et al. 2018). Extra care was taken to select images in the same season/month to increase the accuracy of the research results and the vegetation reflection data (Table 1). The cloudiness was set to 10%, with a sun elevation of more than 30°.
Because the study was focused on vegetation cover dynamics, we selected the band combination of channels that is used to obtain coverage of the vegetation indices’ characteristics. The normalised difference vegetation index (NDVI) imagery set was used to classify the Landsat satellite pattern for the 30-year period (1987–2017), and the change in the vegetation cover distribution of the study area was determined. The NDVI remote sensing method displays the healthiness and greenness (relative biomass) of the vegetation (Prabhakara et al. 2015). The NDVI measures the state of the plant’s health based on the plant’s reflection of light at certain frequencies (EOS 2020). This index takes advantage of the contrast in the characteristics of two bands from a multispectral raster dataset: the chlorophyll pigment absorptions in the red band and the high reflectivity of plant materials in the near-infrared (NIR) band (EOS 2020). For example, the Landsat™ sensor reflects both the near-infrared spectrum in band 4 and visible red (RED) in band 3 (Table 1).
The processing of NDVI indices was carried out by an automated software tool type, with the transfer of the indicators and the subsequent analysis and interpretation of the received data. The types of vegetation cover of different classes were generated in percentages or absolute units of area (m2 or ha). The NDVI EOS land viewer software provides the automatic distribution of the vegetation cover. Both supervised and unsupervised classification methods were used to classify the images in this study. The NDVI value ranges between − 1.0 and + 1.0 and reflects the healthiness of plants (greens). A low NDVI value of 0.1 and below corresponds with rocky and sandy areas or water (i.e. an NDVI below zero means no vegetation). A value from − 1 to 0.1 represent water cover, 0.1 to 0.2 represent grass cover and open soil; sparse vegetation cover values are from 0.2 to 0.4; moderate woody vegetation values are from 0.4 to 0.6; and a large value from 0.6 to 1.0 indicates dense woody vegetation (EOS 2020).
Ground-truthing the vegetation types
The vegetation units within the FSNBG were validated using descriptive methods. To evaluate the status of the vegetation type described in Mucina and Rutherford (2006), we used field surveys that used the systematic quadrat method to sample the vegetation species and number of species (Mokotjomela et al. 2009; Hill et al. 2005). The survey was conducted during the active growing season, during December 2020.
A quadrat method was used to assess whether the typical plant species still exist within the CBA1 vegetation patch (as used in Mokotjomela et al. 2009; Hill et al. 2005). A conventional frame quadrat (i.e. a mini-quadrat) of 1 m2 was employed to quantify the presence or absence of plant species in CBA1 vegetation in the set of systematically located samples. The quadrat method is recommended for visual estimation of plant cover and the presence or absence of plant species during description of vegetation in a target study area (Hill et al. 2005). The over-shoulder-toss method was used to position the mini-quadrat within the patch boundary (as used in Mokotjomela et al. 2009; Hill et al. 2005; Bullock 1997). The high ratio edge of plant species intercepted by a grid line (i.e. in or out decision) was applied (Elzinga et al. 1998). Fifty-two mini-quadrats were sampled within the CBA1 vegetation type, in which the frequency and vegetation cover of all rooted plant species (i.e. herbaceous or woody layer) were recorded.
Within the quadrat, the overall vegetation cover per quadrat (1 m2) was estimated using a modified Braun-Blanquet scale (as used in Mokotjomela et al. 2009; Hill et al. 2005). Plant species recorded in each quadrat sample were identified using existing knowledge, different plant field guides (e.g. Ulian et al. 2019; Fish et al. 2015; Van Wyk and Van Wyk 2013; Joffe and Oberhozer 2012), and a South African database (www.plantzafrica.com). When positive field identification was not possible, plant species were assigned a temporary moniker (e.g. unknown1), and both physical samples (e.g. leaves, flowers) and digital records were collected for further identification in the National Herbarium. Some species were also collected for the South African National Biodiversity Institute (SANBI) herbarium located in Pretoria for further identification. The vegetation parameters were recorded at each quadrat (e.g. vegetation percentage and abundance of plant species). A digital handheld GPS (Garmin Montana® 600) was used for recording the geographic coordinates.
Habitat ecological threats
We quantified the local threats (e.g. biological invasions and human settlement encroachment) using a direct observation method and historical information based on the most frequent threats (as reported in the study of Schulze et al. 2017) that could alter the vegetation cover of the FSNBG.
Alien and invasive plant species
A survey (i.e. a transect walk) was conducted that covered most of the gazetted areas in the Botanical Gardens. The field surveys were undertaken during the period of April 2018 to March 2019. The authoritative taxonomic descriptions were used to identify alien plants to the species level (Ulian et al. 2019; Bromilow 2018; Van Wyk and Van Wyk 2013). For small plant features (i.e. fruits and inflorescence), we used the portable pocket 100 × metal mini science microscope (Song et al. 2018). Different plant field guides (i.e. Van Wyk and Van Wyk 2013; Bromilow 2018) and a South African database (www.invasives.org.za) were used for plant species identification.
The digital handheld GPS (Garmin Montana® 600) was used to collect the individual species’ locations. The GPS gives position coordinates with an accuracy of within 1 to 10 m (Heselton 1998). The data collected each day were stored on the GPS device in keyhole markup language (KML/KMZ) tracks and imported to the Google Earth Pro app. KML/KMZ is a file format used to display geographical data in an earth browser (Marek et al. 2015). The Google Earth plot was overlaid on the Google Earth interface. Different alien and invasive plant species’ density/abundance, habitat type and area of infestation were also recorded and mapped using Google Earth Pro.
Invaded areas (i.e. both lightly and densely invaded areas) were mapped to estimate the aerial vegetation cover of alien and invasive species to guide conservation management’s efforts at prioritisation.
Human settlement encroachment
To evaluate the extent of human settlement encroachment on the FSNBG buffer zone over 19 years (2002–2021), we generated a time series analysis of habitat cover for the years 2002, 2007, 2012, 2017 and 2020 (see Ridding et al. 2020). Google Earth Pro was used to estimate the human settlement aerial cover that occurred over 19 years at 5-year intervals until 2020. A standard buffer area of 1 km for biodiversity-protected areas was applied around the perimeter (Madsen et al. 2014; Margules and Pressey 2000). This buffer area is an area that should not be disturbed as a way of protecting a specific conservation area within a protected area (Ebregt and de Grave 2000). We computed the total human settlement area as a proxy for habitat transformation by human impact (Chowdhury et al. 2018). Polygons were drawn around the anthropogenically altered landscape (i.e. built infrastructure), and the area was measured in hectares and converted into proportions (percentages). Extra care was taken in measuring between points on the built-up perimeter areas to increase the accuracy of the results. Thereafter, Google Earth automatically estimated the area size in hectares, according to specified settings.
Data analysis
Spatial data collection and image processing of vegetation
To analyse the NDVI trends during the study period (1987–2017), different proportions of vegetation cover class in hectares across the 10-year intervals were used, using the 1987 data as the baseline to quantify the potential vegetation cover change in the study site.
Ground-truthing the vegetation types
To validate the vegetation type in the FSNBG through ground-truthing, all the rooted plant species identified within the sampling quadrats were used to generate frequency as a response variable to compare species’ dominance. The plant species’ names were specified as ordinal response variables, and a descriptive statistic was applied to compare the number (i.e. the frequency) of plant species at a significance of P ≤ 0.05, using Statistical Package for Social Sciences (SPSS, version 20) software.
The extent of the threats posed to the CBA1 vegetation type Gh8 Bloemfontein Karroid Shrubland by habitat transformation and degradation (e.g. Skowno et al. 2019; SANBI 2013) was applied to assess their overall ecological integrity. A modified generalised habitat integrity evaluation method adopted from Mokotjomela and Nombewu (2019) was used, in which a habitat with a minimum threshold of 60% habitat intactness was applied (Kleynhans et al. 2008), and used to infer the relative ecological integrity of the CBA1 vegetation type. A ‘category C’ habitat is a moderately modified habitat through some loss or alteration of the natural habitat and biota, but with its basic ecosystem function mainly unchanged (Kleynhans et al. 2008).
We calculated the Shannon–Wiener diversity index (SWDI) to investigate overall current plant species diversity with reference to the original list of plant species provided for the CBA1 vegetation type by Mucina and Rutherford (2006).
Habitat ecological threats
Alien and invasive species
The native range description at the continental level of each recorded alien and invasive species was classified and specified from different sources: books (Henderson 2001, 2020; Bromilow 2018) and a global invasive database to guide adaptive biosecurity management (Seebens et al. 2020; Mokotjomela et al. 2022). Each species was classified into its growth forms following field guides by Bromilow (2018) and Henderson (2001, 2020), and its native range, since this information provides bases for important management decisions in South Africa and growth forms can help to predict the potential impacts of species (Pyšek et al. 2012; Dorrepaal 2007; Mokotjomela et al. 2022). Alien and invasive plant species lists were classified into four categories (categories 1a, 1b, 2, and 3), according to the South African National Environmental Management: Biodiversity Act (NEM:BA, Act 10 of 2004), Alien and Invasive Species Regulations (hereafter called NEM:BA-A&IS Regulations; Department of Environmental Affairs 2020), which were revised in 2020 (No. 43735) to guide management prioritisation (Mokotjomela et al. 2022) for a brief explanation of each alien and invasive species category in the NEM:BA-A&IS regulations of 2020.
The distribution of different alien and invasive plant species was mapped using Google Map Pro software to determine the invasion spatial patterns and to guide prioritisation of their management and containment in the FSNBG.
To determine significant differences in numbers of plant species between groups within different species’ classification criteria, we used a generalised linear model fitted with Poisson error distribution and log link in SPSS software, version 20 (IBM 2011). We specified the response variable as the counts of individual plant species and independent/predictor variables were the growth forms (woody, forb, grass, etc.), continent (Africa, Asia, etc.) and NEM:BA-A&IS regulations' categories.
Human settlement encroachment
To analyse the rate of habitat transformation attributed to human settlement in the FSNBG buffer since 2002, portions of transformed (i.e. anthropogenically altered) landscape, such as the physical development of built infrastructure and plantations, were estimated in hectares for the 5-year intervals, using the 2002 data as a baseline, up to 2021. The habitat change was plotted over time, and the regression analysis was used to determine the strength of the relationship between change and time (coefficient of determination – R2).
Results
Detecting vegetation cover change: spatial data collection of vegetation
The Landsat images were taken at 10-year intervals and reflected the field where there are still signs of vegetation cover. The woody vegetation cover was seen to increase in hectares over time from 1987 to 2017 (Fig. 2). The “moderate woody vegetation cover” has expanded by 25.1 ha and the “dense woody vegetation” also expanded by 8.6 ha over a 30-year period (Fig. 2). Alternatively, the sparse vegetation cover (i.e. the reference cover change) reduced progressively by 11.07 ha from 1987 to 2017 (Fig. 2). Thus, bare soil and grass cover were found to be relatively consistent (Fig. 2).

NDVI (vegetation cover) images demonstrate the 30 years of vegetation cover dynamics of the Free State National Botanical Garden, from 1987 to 2017, in 10-year intervals (NDVI vegetation cover: A 1987, B 1997, C 2007, and D 2017). 0.6 to 1 represent dense woody vegetation cover, 0.4 to 0.6 represent moderate woody vegetation cover, 0.2 to 0.4 represent sparse vegetation cover, 0.1 to 0.2 show the grass cover with bare space, and − 1 to 0.1 represent water cover. The blue line with white dots indicates the perimeter of the FSNBG. The black lines enclosing the black stars (*) show the coverage of the CBA1 vegetation (Bloemfontein Karroid Shrubland)
Ground-truthing plant species in the Critical Biodiversity Area 1 (CBA1) vegetation type: Gh8 Bloemfontein Karroid Shrubland
The overall cover of the CBA1 vegetation type (Gh8 Bloemfontein Karroid Shrubland) was 79.6 ± 15.9% per square metre. Also, the ecological integrity determined using habitat intactness was relatively high (above the 60% threshold for habitat intactness) (sensu Mokotjomela and Nombewu 2019; Kleynhans et al. 2008).
Shannon–Wiener diversity indices showed very high overall plant species diversity (H = 3.5). A total of 55 plant species belonging to 17 families was recorded from 52 quadrats in the 22.37 ha of CBA1 vegetation type (Gh8 Bloemfontein Karroid Shrubland). The dominant plant family was Poaceae (25.6%), followed by Asteraceae (19.6%).
Based on the 77 characteristic taxa list documented by Mucina and Rutherford (2006) with the same vegetation type, 27 (35%, N = 77) plant species still existed and were identified in the field. Of the 12 reference plant species described as dominant by Mucina and Rutherford (2006), 11 species were still found in the CBA1 vegetation type at varying frequencies: Eragrostic nindensis (10.2%; N = 352); Cheilanthes eckloniana (7.1%; N = 352); Euphorbia mauritanica (4.8%; N = 352); Ruschia spinosa (4.8%; N = 352); Albuca setosa (3.7%; N = 352); Aristida diffusa (2.8%; N = 352); Somatium mustellium (2.3%; N = 352); Euryops emetrifolius (2%; N = 352); Eriocephalus ericoides (2%; N = 352); Curio radicans (2%; N = 352); and Heteropogon contortus (0.6%; N = 352) (Fig. 3).

Frequency (%) of sampled plant species within the CBA1 vegetation type (Bloemfontein Karroid Shrubland). The ✶ sign shows the reference plant species and; the
 sign indicates the subsidiary plant species as described in Mucina and Rutherford (2006); ¥ is the extra-limital plant species, and
sign indicates the subsidiary plant species as described in Mucina and Rutherford (2006); ¥ is the extra-limital plant species, and
 indicates Alien and invasive plant species
indicates Alien and invasive plant species
In addition, 16 other plant species were documented in the CBA1 vegetation patch, with Aloe grandidentata (4.3%; N = 352) being the most frequent (Fig. 5). Conversely, 25 extra-limital plant species with Anacampseros subnuda (2.5%; N = 352) and Ledebouria sp. (2.5%; N = 352; Fig. 3) being the most common and three alien and invasive plant species were recorded (i.e. dominated by Bidens bipinnata; Fig. 3).
Alien and invasive plant species
Within the 66-ha perimeter of the FSNBG, we documented a total of 27 alien and invasive plant species belonging to 16 families. The dominant plant family was Cactaceae (22.2%; N = 27), followed by Rosaceae (14.8%; N = 27) (Fig. 4A).

Classification of alien and invasive plant species recorded in the FSNBG. A Species family; B plant growth form; C South African National Environmental Management: Biodiversity Act (NEM:BA, Act 10 of 2004), Alien and Invasive Species Regulations of 2014 revised in 2020 (NEM:BA-A&IS) categories; and D continental level
There were significant differences among the various alien and invasive plant growth forms (Wald χ2 = 12.21; df = 4; p = 0.016) (Fig. 4B). Of the five plant growth forms, woody species occurred most often (58%; N = 27) (Fig. 4B).
Among the four vegetation types (Gh5 Bloemfontein Dry Grassland, Gh8 Bloemfontein Karroid Shrubland, Gh7 Winburg Grassy Shrubland, and Riparian vegetation (Aza5 Highveld Alluvial Vegetation)) conserved in the FSNBG, the Riparian vegetation was the most invaded vegetation type (n = 16), and Cestrum laevigatum (inkberry) was found to be the most invasive species.
The number of alien plant species recorded in the different NEM:BA-A&IS regulations (i.e. NEM:BA-A&IS categories 1a, 1b, 2, & 3) was not statistically different (Wald χ2 = 4.35; df = 3; p = 0.23) (Fig. 4C). Out of 27 recorded plant species, 74% (N = 27) of the species were classified under category 1b in the FSNBG, followed by 19% (N = 27) (Fig. 4C) of the species classified under category 3. Only 4% (N = 27) (Fig. 4C) of the alien and invasive species were not listed in the NEM:BA-A&IS, and there was no record of emerging plant species (category 1a).
There were also no significant differences in the numbers of alien and invasive plant species introduced from different continental levels (Wald χ2 = 4.23; df = 4; p = 0.38) (Fig. 4D). However, most of the recorded alien and invasive plant species were introduced from the Americas (64.5%; N = 27) (Fig. 4D).
Human settlement encroachment
For spatial analysis, habitat fragmentation through human settlement has been encroaching on the FSNBG buffer zone (ca 1 km) during the past 19 years. In 2002, 6.6% (45.59 ha) of the total area of the FSNBG buffer zone was covered by human settlements. Since 2002, almost 12% (73.48 ha) of the FSNBG buffer area became human-modified habitat, while this area had increased to 17.9% (119.17 ha) by 2021 (Fig. 5). Thus, there is a strong increase in human settlement encroachment coverage over time (R2 = 0.79) (Fig. 5).

A bar graph of human settlement encroachment measured in hectares from 2002 to 2021, with habitat change estimated at 5-year intervals
Discussion
Detecting vegetation cover change
In support of our study prediction, the remote sensing and GIS’s investigation on vegetation cover of the FSNBG showed that the woody vegetation cover has expanded over a period of 30 years (i.e. “moderate woody vegetation cover” has expanded by 25.1 ha, and similarly, the “dense woody vegetation” also expanded by 8.6 ha) with the likely driver being the woody species encroachment. It is possible that the observed bush encroachment is also a result of the absence of the biophysical disturbances that are essential for ecological succession in the grassland biome (Eldridge et al. 2011). For example, the exclusion of fire as an essential component of successional changes in the grass communities of the FSNBG vegetation could result in the recruitment of woody species (Neary and Leonard 2020; Hare et al. 2020; Gordijn 2010; Van Rooyen 2010; Trollope 1974) since periodic fires suppress woody plants’ recruitment in the grasslands (Van Rooyen 2010; Le Maitre and Midgley 1992; Trollope and Tainton 1986). In addition, the encroachment of woody plants in the study could be attributed to the absence of browsers that suppress the population growth in palatable woody species (Van Rooyen 2010). It has been shown that trampling by animals could also strongly suppress the woody vegetation cover in savannas (Oliveras and Malhi 2016). This finding is consistent with the report that woody vegetation in grasslands and savannas has increased worldwide over the past 100–200 years (Archer et al. 2017).
The woody state could be attributed to a combination of factors, including severe drought conditions (Jardim et al. 2022; Karakani et al. 2021; Marengo et al. 2017; Botai et al. 2016; Van Rooyen 2010) and the rise of CO2 concentrations in the atmosphere that are induced by global climate change (Archer et al. 2017; Ward 2010) and which enhance the growth of woody species (Sintayehu 2018; Welz 2013; Buitenwerf et al. 2012) and suppress the herbaceous plant cover (Kgosikoma and Mogotsi 2013). Previous studies have shown that drought conditions favour the growth of woody species since the roots of grasses can extract water only from the topsoil layer, while woody species have access to deeper water sources (Ward et al. 2013; Van Rooyen 2010; Walker and Noy-Meir 1982). These differences are likely to increase the negative impacts of the tree-grass interactions in the grassland biome and result in increased bush encroachment (Ward 2005). Several studies have consistently noted that parts of the Grassland Biome in South Africa have undergone extensive modification and has seen a fundamental shift from its original state to resemble savanna vegetation (see Little et al. 2015; Matsika 2008; Neke and du Plessis 2004).
In addition, several studies have demonstrated that the woody vegetation cover in South Africa can be associated with high propagule pressure of native and alien fleshy-fruited woody plant species that are consumed and dispersed by frugivorous birds (Vukeya et al. 2022), such as reported for the savanna habitat by Milton et al. (2007). Therefore, we argue that the observed changes in vegetation cover, which indicate an increase in woody species’ cover between 1987 and 2017, can be partly attributed to the substantial seed rain associated with resident frugivorous bird species and colonial wintering birds (Vukeya et al. 2020; Vukeya et al. 2022). It has been shown that bird species’ gut treatment of the seeds promotes the germination of the local woody plant species (Mokotjomela et al. 2021; Vukeya et al. 2021). Woody cover encroachment reportedly jeopardises grassland biodiversity and transforms grasslands and savannas into shrublands and woodlands (Archer et al. 2001, 2017). Indeed, Mogashoa et al. (2021) found that an increase in tree encroachment decreased grass species’ richness by 53% in South African savannas. Since bush encroachment represents a conservation threat (Archer et al. 2001, 2017), there is an urgent need to mitigate the potential impacts of encroaching woody species on the vegetation types preserved in the FSNBG.
Ground-truthing the vegetation types
Of critical importance in the conservation of vegetation in the FSNBG is the Gh8 Bloemfontein Karroid Shrubland, which is listed in national conservation priorities (SANBI 2017). The validation of ecological integrity showed that the Bloemfontein Karroid Shrubland still has excellent vegetation cover (79.6 ± 15.9%), possibly due to effective protection by the FSNBG. Termansen et al. (2019) indicated that vegetation cover reflects both the natural habitat health and the adaptive management responses. For example, Hesp (1991) reported that excellent vegetation cover reduces splash erosion by intercepting rainfall, decreasing the overland flow and improving the infiltration of precipitation into the soil. In addition, the vegetation intactness was above the threshold of 60%, suggesting partial vegetation loss, although the representative habitat functionality is still maintained (i.e. the category C ecological integrity measured in Mokotjomela and Nombewu 2019; Kleynhans et al. 2008). Indeed, most of the indicator plant species (i.e. 11 out of 12) (Mucina and Rutherford 2006) were still occurring and could be supporting the patch functionality. However, the study found that 50 of the 77 other plant species of Bloemfontein Karroid Shrubland (Mucina and Rutherford 2006) were lost, thus confirming the recorded partial loss of ecological integrity. Two major climatic drivers that might account for the reported change include extreme frost and drought in the Free State Province (Jardim et al. 2022; Abubakar et al. 2020; Basto et al. 2018; Tilman and Haddi 1992). Tilman and Haddi (1992) found that the above-ground living plant mass of grassland vegetation decreased by an average of 47% during an extreme drought condition in 1988; if were to reoccur, it may lead to an increased rate of species’ local extinctions. It is possible that the lost plant species could have been partly replaced by the 25 recorded extra-limital plant species that are currently colonising the Bloemfontein Karroid Shrubland (Begon et al. 1990). Among the species, Anacampseros subnuda and Ledebouria sp. were found to be dominant: A. subnuda has thus far been recorded in the Gm11 Rand Highveld Grassland of Mpumalanga and Gauteng, South Africa (Von Staden and de Castro 2016).
Impacts of alien and invasive plant species
The record of 27 alien and invasive species (of which 13 were woody species) in the study site highlights the vulnerability of the Grassland Biome to the negative impacts of alien species (O’Connor and Van Wilgen 2020; Yapi et al. 2018; Matsika 2008). Many alien trees and shrubs have threatened biodiversity in South African (Roura-Pascual et al. 2009). For example, it was reported that the estimated extent and impact of the Black wattle (Acacia mearnsii) invasion was higher on the Grassland Biome’s surface water runoff in South Africa than that of other plant species (Van Wilgen and Wilson 2018; Henderson and Wilson 2017; Roura-Pascual et al. 2009). Of the 27 plant species, inkberry (Cestrum laevigatum) was found to be the most invasive species in the Riparian vegetation, possessing a huge seed bank that could worsen its invasion risk to wetland biodiversity. Inkberry is known to prevent the regeneration of native species with its dense stands, and it is highly toxic to some wildlife (Weber 2003). This has the possibility to lead native species to their extinction (Winter et al. 2009). Inkberry also has attractive flowers and non-seasonal fruiting patterns that may disrupt native mutualism by attracting native pollinators and seed dispersal agents (Le Roux et al. 2020; Mokotjomela et al. 2013; Traveset and Richardson 2006), subsequently decoupling native mutualist partner organisms. This decoupling may consequently enhance the transformation of the preserved vegetation types in the FSNBG.
We noted that the dominance of the Cactaceae family already has had negative impacts elsewhere and has been targeted for management in South Africa since 1995 (Van Wilgen et al. 2020; Novoa et al. 2015, 2019; Henderson and Wilson 2017; Walters et al. 2011). Masocha and Dube (2018) reported that cactus invasions may threaten native biodiversity in many countries. In South Africa, for example, Dean and Milton (2000) and Du Toit (1942) both reported that about 900,000 ha were abandoned by many landowners in the 1940s owing to the thicket invasion of Opuntia ficus-indica (sweet prickly pear). The problem could be exacerbated by highly improved germination of seeds consumed with fruits and defecated by birds as long-distance dispersal agents (Mokotjomela et al. 2021).
We found many management priority alien species (i.e. category 1b plant species in the NEM:BA-A&IS) (74%, N = 27) and argue that these invaders could also lead to economic loss in the FSNBG. Indeed, Van Wilgen et al. (2020) indicated that, since 1995, the South African government has spent R15 billion on alien plant control operations (e.g. the Working for Water programme) across South Africa. The finding that 4% (N = 27) of the alien species were not listed in NEM:BA-A&IS categories highlights the urgent need for further research and risk analysis into alien taxa to outline the likelihood of invasion, potential impacts in the ecosystem, and to determine options for management needs (Kumschick et al. 2020; IUCN 2020; Keller and Kumschick 2017; Blackburn et al. 2014). Some species are not listed, for example, because of the limited capacity in documenting alien species (Bouchard et al. 2015; Irlich et al. 2017; Mokotjomela et al. 2022). The South American native plant species were dominant, which supports the report by Turbelin et al. (2017) indicating that South America is the main source of the global spread of alien and invasive species through either deliberate or unintentional introduction by human-mediated activities for economic development (e.g. the agricultural industry) and horticultural purposes (e.g. ornamental plants) (Seebens et al. 2015; Mokotjomela et al. 2022).
Impact of human settlement encroachment
The spatial assessment of human settlement encroachment has shown that the integrity of the natural vegetation in the FSNBG buffer area (ca. 1 km) has been decreasing in the past 19 years, which suggests an increasing conservation threat to local biodiversity. Increases in natural habitat loss may disrupt ecological processes, such as the pollination and seed dispersal that is mediated by long-distance agents visiting the FSNBG (Haddad et al. 2015; SANBI 2013; Carey et al. 2000). It has been shown that many protected areas in South Africa also struggle to manage pressure from the loss of natural habitat through human settlement encroachment in areas adjacent to those protected areas (Taylor and Atkinson 2012; Kiringe and Okello 2007). Since the FSNBG is also a protected area, the destruction of biodiversity corridors leads to the isolation of small populations, thus reducing their genetic diversity and persistence (Reisch et al. 2017; Haddad et al. 2015; de Frutos et al. 2015; SANBI 2013; Rosenberg et al. 1997). Rosenberg et al. (1997) indicated that habitat corridors in the protected areas help to increase population persistence by allowing the continued exchange of genetic material between the connected populations. Our results, therefore, indicate an urgent need for conservation and restoration measures to protect habitat corridors and to help maintain ecosystem health in the FSNBG since they would preserve the CBA1 vegetation type.
Based on the above findings, the vegetation management implications of this study’s findings are:
-
1.
Population of trees/shrubs should be reduced in the open grassland in order to reduce their ecological competition pressure on grasses.
-
2.
Biophysical disturbances (i.e. fires and browsers) that are essential for ecological succession in the grassland biome should be used as part of routine management in the garden and possibly in the buffer zone.
-
3.
A regulatory monitoring system to detect vegetation cover change should be established.
-
4.
The management of the environment should be adapted so as to attract dispersers and pollinators to offset the impact of habitat fragmentation.
Conclusions
Vegetation cover provides important indicator of environmental change (Del Vecchio et al. 2015, 2016; Cornelissen 2011), and in many year of the implementation of the conservation gardens strategy in South Africa, none of the existing studies has evaluated the vegetation cover dynamics in SANBI national gardens such as FSNBG. We have shown that contrary to our study prediction, the “moderate vegetation cover” has expanded by 25.1 ha, and similarly, the “dense vegetation” also increased by 8.6 ha over a period of 30 years with a possible driver being the woody species encroachment. Since all gardens are fenced to protect rare biodiversity and reduce property vandalism, we speculated that the absence of biophysical disturbances such as fire and mammal browsers that trample the habitat and lead to an excellent community recruitment in the Grassland Biome (Neary and Leonard 2020), might have led to increased population recruitment of the woody species in FSNBG. In addition, the high propagule pressure from native fleshy-fruited woody plant species, which are consumed and dispersed by localised bird species observed in the study site (Vukeya et al. 2020), is a likely driver of the reported bush encroachment in the FSNBG. This may have been exacerbated by the infiltration by 13 alien woody plant species and eight alien succulent species interspersed within the different vegetation patches.
That the CBA1 patch was above the accepted vegetation intactness threshold of 60% (Mokotjomela and Nombewu 2019; Kleynhans et al. 2008), suggests that this patch still possesses representative habitat functionality and desired conservation value. We have argued that the ecological integrity of the CBA1 vegetation type could also have been reduced by several biodiversity conservation threats, including alien plant invasion, which reportedly transform natural habitats (Van Wilgen et al. 2020). Although the neighbouring ecological support vegetation areas (ESAs) were found to be in good condition for the vegetation in CBAs, they may also be experiencing some pressures that indirectly affected integrity of CBA1s, and this requires some further focused research. We contend that regular monitoring of vegetation and control of native tree populations in the study site are needed for proper conservation planning as recommended by Jardim et al. (2022) and Carrasco et al. (2020). Such knowledge could help offset the potential negative impacts of the woody plant encroachment in FSNBG and thus guide adaptive management of the protected unique biodiversity.
Change history
08 June 2023
A Correction to this paper has been published: https://doi.org/10.1007/s10113-023-02087-2
References
Abubakar HB, Newete WS, Scholes MC (2020) Drought characterization and trend detection using the reconnaissance drought index for Setsoto Municipality of the Free State Province of South Africa and the impact on maize yield. Water 12:2993. https://doi.org/10.3390/w12112993
Archer SR, Boutton T, Hibbard K (2001) Trees in grasslands: biogeochemical consequences of woody plant expansion. In: Schulze E, Harrison S, Heimann M, Holland E, Lloyd J, Prentice I, Schimel D (eds) Global biogeochemical cycles in the climate system, p 47 Academic Press, San Diego. https://doi.org/10.1016/B978-012631260-7/50011-X
Archer SR, Andersen EM, Predick KI, Schwinning S, Steidl RJ et al (2017) Woody plant encroachment: causes and consequences. In: Briske D (ed) Rangeland systems. Springer Series on Environmental Management. Springer, Cham. https://doi.org/10.1007/978-3-319-46709-2
Basto S, Thompson K, Grime JP, Fridley JD, Calhim S et al (2018) Severe effects of long-term drought on calcareous grassland seed banks. Clim Atmos Sci 1(1). https://doi.org/10.1038/s41612-41017-a40007-41613
Baum KA, Haynes KJ, Dillemut F, Cronin JT (2004) The matrix enhances the effectiveness of corridors and stepping stones. Ecology 85(10):2671–2676. https://doi.org/10.1890/04-0500
Begon M, Harper JL, Townsend CR (1990) Ecology: individuals, populations, and communities. Blackwell Scientific, Oxford
Blackburn TM, Essl F, Evans T, Hulme PE, Jeschke JM, Kühn I et al (2014) A unified classification of alien species based on the magnitude of their environmental impacts. PLoS Biol 12(5):e1001850. https://doi.org/10.1371/journal.pbio.1001850
Botai CM, Botai JO, Dlamin LC, Zwane NS, Phaduli E (2016) Characteristics of droughts in South Africa: a case study of Free State and North West Provinces. Water 8(10):439. https://doi.org/10.3390/w8100439
Bouchard EH, Little LE, Miller CML, Rundell SM, Vlodaver EM et al (2015) Undeclared baggage: do tourists act as vectors for seed dispersal in fynbos protected areas? Koedoe 57(1):Art. #1323, 9 pages. https://doi.org/10.4102/koedoe.v57i1.1323
Bromilow C (2018) Problem plants and alien weeds of Southern Africa. Fourth edition. Pretoria: Briza Publications, Pretoria: South Africa
Buitenwerf R, Bond WJ, Stevens N, Trollope W (2012) Increased tree densities in South African savannas: > 50 years of data suggests CO2 as a driver. Glob Chang Biol 18(2):675–684. https://doi.org/10.1111/j.1365-2486.2011.02561.x
Bullock J (1997) Plants. In: Sutherland WJ (ed) Ecological census techniques: a handbook. Cambridge University, Cambridge, pp 186–213
Carbutt C, Tau M, Stephens A, Escott B (2011) The conservation status of temperate grasslands in southern Africa. The Grassland Society of Southern Africa 11(1):17–22
Carey C, Dudley N, Stolton S (2000) Squandering parasite? The importance and vulnerability of the world’s protected areas. World Wide Fund for Nature International, Gland, Switzerland
Carrasco RA, Pinheiro MMF, Junior JM, Cicerelli RE, Silva PA et al (2020) Land use/land cover change dynamics and their effects on land surface temperature in the western region of the state of São Paulo, Brazil. Reg Environ Chang 20(3):1–12. https://doi.org/10.1007/s10113-020-01664-z
Case MF, Staver AC (2017) Fire prevents woody encroachment only at higher-than-historical frequencies in a South African savanna. J Appl Ecol 54:955–962. https://doi.org/10.1111/1365-2664.12805
Chaplin PJ (1979) Orange Free State Botanic Gardens. Volkskas (Ltd)
Chowdhury PKR, Bhaduri BL, McKee JJ (2018) Estimating urban areas: new insights from very high-resolution human settlement data. Remote Sens Appl: Soc Environ 10:93–103. https://doi.org/10.1016/j.rsase.2018.03.002
Cornelissen T (2011) Climate change and its effects on terrestrial insects and herbivory patterns. Neotrop Entomol 40(2):155–163. https://doi.org/10.1590/S1519-566X2011000200001
Daehler C (2003) Performance comparisons of co-occurring native and alien invasive plants: Implications for conservation and restoration. Annu Rev Ecol Evol Syst 34:183–211. https://www.jstor.org/stable/30033774
Das S, Angadi DP (2022) Land use land cover change detection and monitoring of urban growth using remote sensing and GIS techniques: a micro-level study. GeoJournal 87(3):2101–2123. https://doi.org/10.1007/s10708-020-10359-1
Dayaram A, Harris LR, Grobler BA, Van der Merwe S, Powrie LW, et al. (2019) Vegetation map of South Africa, Lesotho and Swaziland 2018: a description of changes since 2006. Bothalia 49(1):a2452. https://doi.org/10.4102/abc.v49i1.2452
De Frutos A, Navarro T, Pueyo Y, Alados CL (2015) Inferring resilience to fragmentation induced changes in plant communities in a semi-arid Mediterranean ecosystem. PLoSoNE 10(3):e0118837. https://doi.org/10.1371/journal.pone.0118837
Dean WRJ, Milton SJ (2000) Directed dispersed of Opuntia species in the Karoo, South Africa: are crows the responsible agents? J Arid Environ 45:305–314. https://doi.org/10.1006/jare.2000.0652
Del Vecchio S, Prisco I, Acosta ATR, Stanisci A (2015) Changes in plant species composition of coastal dune habitats over a 20-year period. AoB Plants 7:plv018. https://doi.org/10.1093/aobpla/plv018
Del Vecchio S, Slaviero A, Fantinato E, Buffa G (2016) The use of plant community attributes to detect habitat quality in coastal environments. AoB Plants 8:plw040. https://doi.org/10.1093/aobpla/plw040.
Dorrepaal E (2007) Are plant growth-form-based classifications useful in predicting northern ecosystem carbon cycling feedbacks to climate change? J Ecol 95:1167–1180. https://doi.org/10.1111/j.1365-2745.2007.01294.x
Du Toit R (1942) The spread of prickly pear in the Union. Farming in South Africa 17:300–304
Ebregt A, de Grave P (2000) Buffer zones and their management: policy and best practices for terrestrial ecosystems in developing countries. Theme Studies Series 5. EC-LNV and IAC, Wageningen
Eldridge DJ, Bowker MA, Maestre FT, Roger E, Reynolds JF et al (2011) Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol Lett 14:709–722. https://doi.org/10.1111/j.1461-0248.2011.01630.x
Elzinga CL, Saizer DW, Willoughby JW (1998) Measuring and monitoring plant populations. Bureau of Land Management, Denver, CO. BLM Technical Reference 1730–1. BLM/RS/ST-98/005+1730
EOS Homepage. https://eos.com/landviewer, last accessed: 2020/02/24
Fish L, Mashau AC, Moeaha MJ, Nembudani MT (2015) Identification guide to southern African grasses. An identification manual with keys, descriptions and distributions. Strelitzia 36. South African National Biodiversity Institute, Pretoria
Gordijn PJ (2010) The role of fire in bush encroachment in Ithala Game Reserve. Master of Science in the Discipline of Botany. University of KwaZulu-Natal. http://hdl.handle.net/10413/10776
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecol Lett 8(9):993–1009. https://doi.org/10.1111/j.1461-0248.2005.00792.x
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A et al (2015) (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystem. Sci Adv 1:e1500052. https://doi.org/10.1126/sciadv.1500052
Haddad CR, Jager LJC, Foord SH (2019) Habitats and cardinal directions are key variables structuring spider leaf litter assemblages under Searsia lancea. Pedobiologia-J Soil Ecol 73:10–19. https://doi.org/10.1016/j.pedobi.2019.01.002
Hare ML, Xu X, Wang Y, Gedda AI (2020) The effects of bush control methods on encroaching woody plants in terms of die-off and survival in Borana rangelands, southern Ethiopi. Pastoralism 10:16. https://doi.org/10.1186/s13570-020-00171-4
Henderson L, Wilson JRU (2017) Changes in the composition and distribution of alien plants in South Africa: an update from the Southern African Plant Invaders Atlas. Bothalia 47(2):a2172. https://hdl.handle.net/10520/EJC-7270550e0
Henderson L (2001) Alien weeds and invasive plants. A complete guide to declared weeds and invaders in South Africa. Plant Protection Research Institute South Africa
Henderson L (2020) Invasive alien plants in South Africa. Agriculture Research Council. ISBN: 978–0–620–86146–5
Heselton RR (1998) Elevation effects on GPS positional accuracy. Master of Science in Geography. Virginia Polytechnic Institute and State University
Hesp PA (1991) Ecological processes and plant adaptations on coastal dunes. J Arid Environ 21:165–191. https://doi.org/10.1016/S0140-1963(18)30681-5
Hill D, Fasham M, Tucker G, Shewry M, Shaw P (2005) Handbook of biodiversity methods. Cambridge University Press
Iponga DM, Milton SJ, Richardson DM (2008) Superiority in competition for light: a crucial attribute defining the impact of the invasive alien tree Schinus molle (Anacardiaceae) in South African savanna. J Arid Environ 72:612–623. https://doi.org/10.1016/j.jaridenv.2007.10.001
Irlich UM, Potgieter L, Stafford L, Gaertner M (2017) Recommendations for municipalities to become compliant with national legislation on biological invasions. Bothalia 47:a2156. https://doi.org/10.4102/abc.v47i2.2156
IUCN (2020) IUCN EICAT Categories and criteria. The Environmental Impact Classification for Alien Taxa (EICAT), 1st edn. IUCN, Gland, Switzerland and Cambridge, UK
Jardim AMDRF, Araújo Júnior GDN, Silva MVD, Santos AD, Silva JLBD et al (2022) Using remote sensing to quantify the joint effects of climate and land use/land cover changes on the Caatinga Biome of Northeast Brazilian. Remote Sensing 14(8):1911. https://doi.org/10.3390/rs14081911
Joffe P, Oberhozer T (2012) Creative gardening with indigenous plants: a South African guide. 2 edition. Briza
Karakani EG, Malekian A, Gholami S, Liu J (2021) Spatiotemporal monitoring and change detection of vegetation cover for drought management in the Middle East. Theoret Appl Climatol 144(1):299–315. https://doi.org/10.1007/s00704-021-03543-x
Karger DN, Conrad O, Böhner J, Kawohl T, Kreft H et al (2017) Data descriptor: climatologies at high resolution for the earth’s land surface areas. Sci Data 4:170122. https://doi.org/10.1038/sdata.2017.122
Keller RP, Kumschick S (2017) Promise and challenges of risk assessment as an approach for preventing the arrival of harmful alien species. Bothalia 47(2):1–8. a2136. https://doi.org/10.4102/abc.v47i2.2136
Kgosikoma OE, Mogotsi K (2013) Understanding the causes of bush encroachment in Africa: the key to effective management of savanna grasslands. Tropical Grasslands-Forrajes Tropicals 1:215–219. https://uknowledge.uky.edu/igc/22/2-12/3.
Kiringe JW, Okello MM (2007) Threats and their relative severity to wildlife protected areas of Kenya. Appl Ecol Environ Res 5(2):49–62. https://doi.org/10.15666/aeer/0502_049062
Kleynhans CJ, Louw MD, Graham M (2008) River reach demarcation, delineation and suitability. In: Department of Water Affairs and Forestry, South Africa. 2008. Comprehensive Reserve Determination Study for Selected Water Resources (Rivers, Groundwater and Wetlands) in the Inkomati Water Management Area, Mpumalanga. Sabie and Crocodile Systems: Resource Unit Delineation: Prepared by Water for Africa, authored by Louw, MD. Report no. 26/8/3/10/12/006
Kumschick S, Wilson JRU, Foxcroft LC (2020) A framework to support alien species regulation: The Risk Analysis for Alien Taxa (RAAT). NeoBiota 62:213–239. https://doi.org/10.3897/neobiota.62.51031
Le Maitre DC, Midgley JJ (1992) Plant reproductive ecology. In: Cowling RM (ed) The ecology of Fynbos: nutrients, fire and diversity. Oxford University Press, Cape Town, pp 135–174
Little IT, Hockey PAR, Jansen R (2015) Impacts of fire and grazing management on South Africa’s moist highland grasslands: a case study of the Steenkampsberg Plateau, Mpumalanga, South Africa. Bothalia 45(1):15. https://doi.org/10.4102/abc.v45i1.178
Lowe MR, Wu W, Peterson MS, Brown-Peterson NJ, Slack WT et al (2012) Survival, growth and reproduction of non-native Nile Tilapia II: fundamental niche projections and invasion potential in the Northern Gulf of Mexico. PLoS ONE 7(7):e41580. https://doi.org/10.1371/journal.pone.0041580
Mackellar N, New M, Jack C (2014) Observed and modelled trends in rainfall and temperature for South Africa: 1960–2010. S Afr J Sci 110(7/8):Art. #2013–0353, 13 pages. https://doi.org/10.1590/sajs.2014/20130353
Madsen T, Schipperij J, Christiansen LB, Nielsen TS, Troelsen J (2014) Developing suitable buffers to capture transport cycling behavior. Front Public Health 2(61):1–8. https://doi.org/10.3389/fpubh.2014.00061
Marek L, Tuček P, Pászto V (2015) Using geovisual analytics in Google Earth to understand disease distribution: a case study of campylobacteriosis in the Czech Republic (2008–2012). Int J Health Geogr 14:7. https://doi.org/10.1186/1476-072X-14-7
Marengo JA, Torres RR, Alves LM (2017) Drought in Northeast Brazil—past, present, and future. Theoret Appl Climatol 129(3):1189–1200. https://doi.org/10.1007/s00704-016-1840-8
Margules CR, Pressey RL (2000) Systematic conservation planning. Nature 405:243–253. https://doi.org/10.1038/35012251
Masilo T (1999) The development of ISO 14001-based environmental management in the Free State National Botanical Garden. Environmental Management, University Free State
Masocha M, Dube T (2018) Global terrestrial biomes at risk of cacti invasion identified for four species using consensual modelling. J Arid Environ 156:77–86. https://doi.org/10.1016/j.jaridenv.2018.05.006
Matsika R (2008) Land-cover change: threats to the grassland biome of South Africa. Master of science in resource conservation biology University of the Witwatersrand
Milton SJ, Wilson JRU, Richardson DM, Seymour CL, Dean WRJ et al (2007) Invasive alien plants infiltrate bird-mediated shrub nucleation processes in arid savanna. J Ecol. https://doi.org/10.1111/j.1365-2745.2007.01247.x
Mogashoa R, Dlamini P, Gxasheka M (2021) Grass species richness decreases along a woody plant encroachment gradient in a semi-arid savanna grassland, South Africa. Landsc Ecol. https://doi.org/10.1007/s10980-020-01150-1
Mokotjomela TM, Nombewu N (2019) Potential benefits associated with implementation of the national biodiversity economy strategy in the Eastern Cape Province, South Africa. S Afr Geogr J. https://doi.org/10.1080/03736245.2019.1670233
Mokotjomela TM, Schwaibold U, Pillay N (2009) Does the ice rat Otomys sloggetti robertsi contribute to habitat change in Lesotho? Acta Oecol 35:437–443. https://doi.org/10.1016/j.actao.2009.01.004
Mokotjomela TM, Musil CF, Esler KJ (2013) Frugivorous birds visit fruits of emerging alien shrub species more frequently than those of native shrubs species in the South African Mediterranean climate region. S Afr J Bot 86:73–78. https://doi.org/10.1016/j.sajb.2013.02.004
Mokotjomela TM, Nemurangoni T, Mundalamo T, Jaca TP, Kuhudzai AG (2022) The value of dump sites for monitoring biological invasions in South Africa. Biol Invasions 24(4):971–986. https://doi.org/10.1007/s10530-021-02683-5
Mokotjomela TM, Thabethe V, Downs C (2021) Comparing germination metrics of Opuntia ficus-indica and O. robusta between two sets of bird species (Pied Crows and smaller species). Acta Oecol 110:103676. https://doi.org/10.1016/j.actao.2020.103676
Mucina L, Rutherford MC (eds) (2006) The vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19, South African National Biodiversity Institute, Pretoria
Neary DG, Leonard JM (2020) Effects of fire on grassland soils and water: a review. In: Kindomihou, Valentin Missiako (ed) Grasses and grassland aspects. IntechOpen. Online: https://www.intechopen.com/books/grasses-and-grassland-aspects/effects-of-fire-on-grassland-soils-and-water-a-review
Neethling JA, Haddad CR (2013) Arboreal spider assemblages associated with four tree species in the Grassland Biome of central South Africa (Arachnida: Araneae). Trans R Soc S Afr 68(2):123–131. https://hdl.handle.net/10520/EJC137981
Neke KS, du Plessis MA (2004) The threat of transformation: quantifying the vulnerability of grassland in South Africa. Conserv Biol 18(2):466–477. https://doi.org/10.1111/j.1523-1739.2004.00157.x
Niang I, Ruppel OC, Abdrabo MA, Essel A, Lennard C et al (2014) Africa. In: Barros VR, CB Field, DJ Dokken, MD Mastrandrea, KJ Mach et al (eds) Climate change 2014: impacts, adaptation, and vulnerability. Part B: Regional aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. United Kingdom and New York, NY, USA: Cambridge University Press, Cambridge:1199–1265
Novoa A, Brundu G, Day MD, Deltoro V, Essl F et al (2019) Global actions for managing cactus invasions. Plants MDPI 8:421. https://doi.org/10.3390/plants8100421
Novoa A, Le Roux JJ, Robertson MP, Wilson JRU, Richardson DM (2015) Introduced and invasive cactus species: a global review. AoB Plants 7:plu078. https://doi.org/10.1093/aobpla/plu078
O’Connor T, van Wilgen BW (2020) The impact of invasive alien plants on rangelands in South Africa. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 457–486. https://doi.org/10.1007/978-3-030-32394-3_16
Oliveras I, Malhi Y (2016) Many shades of green: the dynamic tropical forest-savannah transition zones. Philos Trans R Soc Lond B Biol Sci 371(1703):20150308. https://doi.org/10.1098/rstb.2015.0308
Pacifici M, Foden WB, Visconti P, Watson JEM, Butchart SHM et al (2015) Assessing species vulnerability to climate change. Nat Clim Chang 5:215–225. https://doi.org/10.1038/nclimate2448
Peter A, Mujuru M, Dube T (2018) An assessment of land cover changes in a protected nature reserve and possible implications on water resources, South Africa. Phys Chem Earth, Part A/B/C 107:86–96. https://doi.org/10.1016/j.pce.2018.09.005
Plumptre AJ, Baisero D, Belote T, Vázquez-Domínguez E, Faurby S et al (2021) Where might we find ecologically intact communities? Front For Glob Chang. https://doi.org/10.3389/ffgc.2021.626635
Prabhakara K, Ahively WD, McCarty GW (2015) Evaluating the relationship between biomass, percent groundcover and remote sensing indices across six winter cover crop fields in Maryland, United State. Int J Appl Earth Obs Geoinf 39:88–102. https://doi.org/10.1016/j.jag.2015.03.002
Pyšek P, Jarosik V, Hulme PE, Pergl J, Hejda M et al (2012) A global assessment of invasive plant impacts on resident species, communities and ecosystems: the interaction of impact measures, invading species’ traits and environment. Glob Change Biol. https://doi.org/10.1111/j.1365-2486.2011.02636.x
Randin CF, Engler R, Normand S, Zappa M, Zimmermann NE et al (2009) Climate change and plant distribution: local models predict high-elevation persistence. Glob Change Biol 15(6):1557–1569. https://doi.org/10.1111/j.1365-2486.2008.01766.x
Reisch C, Schmidkonz S, Meier K, Schöpplein Q, Meyer C et al (2017) Genetic diversity of calcareous grassland plant species depends on historical landscape configuration. Reisch et al. BMC Ecol 17:19. https://doi.org/10.1186/s12898-017-0129-9
Rejmanek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77(6):1655–1661. https://doi.org/10.2307/2265768
Ridding LE, Watson SCL, Newton AC, Rowland CS, Bullock JM (2020) Ongoing, but slowing, habitat loss in a rural landscape over 85 years. Landscape Ecol 35:257–273. https://doi.org/10.1007/s10980-019-00944-2
Rosenberg DK, Noon BR, Meslow EC (1997) Biological corridors: form, function, and efficacy. Bioscience 47:677–687. https://doi.org/10.2307/1313208
Roura-Pascual N, Richardson DM, Krug R, Brown A, Chapman RA et al (2009) Ecology and management of alien plant invasions in South African fynbos: accommodating key complexities in objective decision- making. Biol Conserv 142(8):1595–1604. https://doi.org/10.1016/j.biocon.2009.02.029
Le Roux JJ et al (2020) Biotic interactions as mediators of biological invasions: insights from South Africa. In: van Wilgen B, Measey J, Richardson D, Wilson J, Zengeya T (eds) Biological Invasions in South Africa. Invading Nature - Springer Series in Invasion Ecology, vol 14. Springer, Cham. https://doi.org/10.1007/978-3-030-32394-3_14
Rutherford M, Midgley GF, Bond WJ, Powrie LW, Roberts R et al (2000) Plant biodiversity: vulnerability and adaptation assessment (Tourism, D.O.E.a.A. Trans.). In: Kiker G (ed) Climate change impacts in southern Africa. Report to the National Climate Change Committee
Rutherford MC, Powrie LW (2013) Impacts of heavy grazing on plant species richness: a comparison across rangeland biomes of South Africa. S Afr J Bot 87:146–156. https://doi.org/10.1016/j.sajb.2013.03.020
SANBI (2013) Grazing and burning guidelines: managing grasslands for biodiversity and livestock production. Compiled by Lechmere-Oertel, R.G. South African National Biodiversity Institute, Pretoria
SANBI (2017) Technical guidelines for CBA Maps: Guidelines for developing a map of critical biodiversity areas & ecological support areas using systematic biodiversity planning. In: Driver A, Holness S, Daniels F (eds) Pretoria: South African National Biodiversity Institute
SANBI (2018) National Vegetation Map Project (VEGMAP): 2018 Vegetation Map. South African National Biodiversity Institute. http://bgis.sanbi.org/Projects/Detail/208. Accessed 20 July 2022
Scholes RJ, Archer SR (1997) Tree-grass interactions in savannas. Annu Rev Ecol Syst 28:517–44. https://www.jstor.org/stable/2952503
Schulze K, Knight K, Coad L, Geldmann J, Leverington F et al (2017) An assessment of threats to terrestrial protected areas. Conserv Lett 11:e12435. https://doi.org/10.1111/conl.12435
Seebens H, Essl F, Dawson W, Fuentes N, Moser D, Pergl J, Pyšek P, van Kleunen M, Weber E, Winter M, Blasius B (2015) Global trade will accelerate plant invasions in emerging economies under climate change. Glob Chang Biol 21(11):4128–4140. https://doi.org/10.1111/gcb.13021
Seebens H, Bacher S, Blackburn TM, Capinha C, Dawson W et al (2020) Projecting the continental accumulation of alien species through to 2050. Glob Change Biol 00:1–13. https://doi.org/10.1111/gcb.15333
Sintayehu DW (2018) Impact of climate change on biodiversity and associated key ecosystem services in Africa: a systematic review. Ecosyst Health Sustain 4(9):225–239. https://doi.org/10.1080/20964129.2018.1530054
Skowno AL, Poole CJ, Raimondo DC, Sink KJ, Van Deventer H et al (2019) National Biodiversity Assessment 2018: the status of South Africa’s ecosystems and biodiversity. Synthesis Report. South African National Biodiversity Institute, an entity of the Department of Environment, Forestry and Fisheries, Pretoria. pp 1–214
Smith RIL (1994) Vascular plants as bioindicators of regional warming in Antarctica. Oecologia 99:322–328. https://doi.org/10.1007/BF00627745
Song J, Moon BC, Choi G, Yang S (2018) Morphological identification of Lepidii seu descurainiae semen and adulterant seeds using microscopic analysis. Appl Sci 8:2134. https://doi.org/10.3390/app8112134
Stankovic D, Krstic B, Orlovic S, Trivan G, Poljak PL et al (2011) Woody plants and herbs as bioindicators of the current condition of the natural environment in Serbia. J Med Plants Res 5(15):3507–3512. E at http://www.academicjournals.org/JMPR
Steffen W, Burbidge AA, Hughes L, Kitching R, Lindenmayer D et al (2009) Australia’s biodiversity and climate change: a strategic assessment of the vulnerability of Australia’s biodiversity to climate change. A report to the Natura Resource Management Ministerial Council commissioned by the Australian Government. CSIRO Publishing
Taylor SJ, Atkinson D (2012) ‘Delivering community benefits acts as insurance for the survival of small protected areas such as the Abe Bailey Nature Reserve, South Africa’. Koedoe 54(1):Art. #1043, 9 pages. https://doi.org/10.4102/koedoe.v54i1.1043.
Termansen M, Chapman DS, Quinn CH, Fraser EDG, Jin N et al (2019) Modelling land use dynamics in socio-ecological systems: a case study in the UK uplands. In: Bohan DA, Dumbrell AJ (eds) Resilience in complex socio-ecological system. Advances in Ecological Research 60:125–152. https://doi.org/10.1016/bs.aecr.2019.03.002
Thuiller WL, Lavorel S, Araujo MB, Sykes MT, Prentice IC (2005) Climate change threats to plant diversity in Europe. Proc Natl Acad Sci USA 102:8245–8250
Tilman D, Haddi A (1992) Drought and biodiversity in grasslands. Oecologia 89(2):257–264. https://doi.org/10.1007/BF0031722
Traveset A, Richardson DM (2006) Biological invasions as disruptors of plant reproductive mutualisms. Trends Ecol Evol 21(4):208–216. https://doi.org/10.1016/j.tree.2006.01.006
Trollope WSW (1974) Role of fire in preventing bush encroachment in the Eastern Cape. Proceedings of the Annual Congresses of the Grassland Society of Southern Africa 9(1):67–72. https://doi.org/10.1080/00725560.1974.9648722
Trollope WSW, Tainton NM (1986) Effect of fire intensity on the grass and brush components of the Eastern cape thornveld. Afr J Range Forage Sci 3(2). https://doi.org/10.1080/02566702.1986.9648030
Turbelin AJ, Malamud BD, Francis RA (2017) Mapping the global state of invasive alien species: patterns of invasion and policy responses. Glob Ecol Biogeogr 26:78–92. https://doi.org/10.1111/geb.12517
Ulian T, Flores C, Lira R, Mamatsharaga A, Mogotsi KK et al (eds) (2019) Wild plants for a sustainable future: 110 Multipurpose species. Royal Botanic Garden, Kew
Van Rooyen N (2010) Veld management principles and procedures. In: Bothma JP, du Toit JG (eds) Game Ranch Management, 5th. Van Schaik Publishers: 779–847
Van Wilgen BW, Wilson JR (eds) (2018) The status of biological invasions and their management in South Africa in 2017. South African National Biodiversity Institute, Kirstenbosch and DST-NRF Centre of Excellence for Invasion Biology, Stellenbosch
Van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (2020) Biological invasions in South Africa: an overview. In: van Wilgen B, Measey J, Richardson D, Wilson J, Zengeya T (eds) Biological invasions in South Africa. Invading Nature - Springer Series in Invasion Ecology vol 14. Springer, Cham. https://doi.org/10.1007/978-3-030-32394-3_13
Van Wyk B, van Wyk P (2013) Field guide to trees of Southern Africa. 2nd ed. Struik Nature, Cape Town
Von Staden L, de Castro A (2016) Anacampseros subnuda Poelln. subsp. lubbersii (Bleck) Gerbaulet. National Assessment: Red List of South African Plants version 2020.1. Accessed on 2020/12/13
Vukeya LR, Mokotjomela TM, Malebo NJ, Oke S (2021) Interspecific competition in germination of bird-dispersed seeds in a habitat with sparse tree vegetation in South Africa. Bot Stud 62(10):1–18. https://doi.org/10.1186/s40529-021-00317-6
Vukeya LR, Mokotjomela TM, Malebo NJ, Oke S (2022) Seed dispersal phenology of encroaching woody species in the Free State National Botanical Garden, South Africa. Afr J Ecol 00:1–13. https://doi.org/10.1111/aje.13013
Vukeya LR, Mokotjomela TM, Malebo NJ, Oke S (2020) A 40 years’ vegetation cover dynamics in the Free State National Botanical Garden, South Africa. 46th Annual conference of the South African Association of Botanists (SAAB) 7-10 January 2020 QwaQwa Campus, University of the Free State.https://doi.org/10.13140/RG.2.2.34212.30083
Walker BH, Noy-Meir I (1982) Aspects of the stability and resilience of savanna ecosystems. In: Huntley BJ, Walker BH (eds) Ecology of tropical savannas. Springer, Berlin: 556–590. https://doi.org/10.1007/978-3-642-68786-0_26
Walters M, Figueiredo E, Crouch NR, Winter PJD, Smith GF et al (2011) ‘Naturalised and invasive succulents of Southern Africa’, ABC Taxa 11 91–217. http://hdl.handle.net/20.500.12143/274
Ward D (2005) Do we understand the causes of bush encroachment in African savannas? Afr J Range Forage Sci 22:101–105. https://doi.org/10.2989/10220110509485867
Ward D (2010) A resource ratio model of the effects of changes in CO2 on woody plant invasion. Plant Ecol 209:147–152. https://doi.org/10.1007/s11258-010-9731-z
Ward D, Wiegand K, Getzin S (2013) Walter’s two-layer hypothesis revisited: back to the roots! Oecologia 172:617–630. https://doi.org/10.1007/s00442-012-2538-y
Weber E (2003) Invasive plant species of the world: a reference guide to environmental weeds. CABI Publishing
Welz A (2013) The surprising role of CO2 in changes on the African Savanna. Yale School of Forestry & Environmental Studies: Yale Environment 360
Winter M, Schweiger O, Klotz S, Nentwig W, Andriopoulos P et al (2009) Plant extinctions and introductions lead to phylogenetic and taxonomic homogenization of the European flora. PNAS 106(51):21721–21725. https://doi.org/10.1073/pnas.0907088106
Wong L, Grainger TN, Start D, Gilbert B (2017) An invasive herbivore structures plant competitive dynamic. Biol Let 13:20170374. https://doi.org/10.1098/rsbl.2017.0374
Yapi TS, O’farrelL PJ, Dziba LE, Esler KJ (2018) Alien tree invasion into a South African montane grassland ecosystem: impact of Acacia species on rangeland condition and livestock carrying capacityInt J Biodivers Sci, Ecosyst Serv Manag 14(1):105 116https://doi.org/10.1080/21513732.2018.1450291
Yukhnovskyi V, Zibtseva O (2020) Green space trends in small towns of Kyiv region according to EOS Land Viewer – a case study. J For Sci 66(6):252–263. https://doi.org/10.17221/142/2019-JFS
Zengeya TA, Wilson JR (eds) (2020) The status of biological invasions and their management in South Africa in 2019. pp.71. South African National Biodiversity Institute, Kirstenbosch and DSI-NRF Centre of Excellence for Invasion Biology, Stellenbosch. https://doi.org/10.5281/zenodo.3947613
Acknowledgements
We thank the South African National Biodiversity Institute (SANBI) Free State National Botanical Garden (FSNBG) for the use of the study site. The Central University of Technology (CUT) and Centre for Invasion Biology (CIB) are acknowledged for the support provided for the mapping of alien and invasive species in the FSNBG and the development of a management plan. We thank Mr Leslie Powrie for providing insightful comments on the MS draft.
Funding
Funding support for this study was provided by the South African National Biodiversity Institute (SANBI) and the Central University of Technology (CUT) (Free State’s Research Grants and Scholarship). The South African Department of Forestry, Fisheries and the Environment (DFFE) are also thanked for partial funding, noting that this publication does not necessarily represent the views or opinions of DFFE or its employees.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Communicated by Xiangzheng Deng
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised: In this article, "Gh7 Bloemfontein Karroid Shrubland" must be changed to "Gh8 Bloemfontein Karroid Shrubland".
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vukeya, L.R., Mokotjomela, T.M., Malebo, N.J. et al. The vegetation cover dynamics and potential drivers of habitat change over 30 years in the Free State National Botanical Garden, South Africa. Reg Environ Change 23, 24 (2023). https://doi.org/10.1007/s10113-022-02024-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10113-022-02024-9