
Abstract
The illusory experience of self-motion known as vection, is a multisensory phenomenon relevant to self-motion processes. While some studies have shown that neck muscle vibrations can improve self-motion parameter estimation, the influence on vection remains unknown. Further, few studies measured cybersickness (CS), presence, and vection concurrently and have shown conflicting results. It was hypothesized that 1) neck vibrations would enhance vection and presence, and 2) CS to negatively relate to presence and vection, whereas presence and vection to positively relate to one another. Thirty-two participants were visually and audibly immersed in a virtual reality flight simulator and occasional neck muscle vibrations were presented. Vection onset and duration were reported through button presses. Turning angle estimations and ratings of vection quality, presence, and CS were obtained after completion of the flights. Results showed no influence of vibrations on turning angle estimation errors, but a medium positive effect of vibrations on vection quality was found. Presence and vection quality were positively related, and no strong association between CS and presence or vection was found. It is concluded that neck vibrations may enhance vection and presence, however, from the current study it is unclear whether this is due to proprioceptive or tactile stimulation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Sometimes we experience the sensation of self-movement without physically moving. This phenomenon is referred to as vection. Vection has commonly been defined as a visually induced illusion (see Palmisano et al. 2015 for a brief overview of definitions), however, auditory (Lackner 1977; Larsson et al. 2004; Väljamäe et al. 2009), biomechanical (Ash et al. 2013; Bles et al. 1995; Bles and Kapteyn 1977), tactile (Murata et al. 2014; Tinga et al. 2018, see Kooijman et al. 2022a for a review), and olfactory (Aruga et al. 2019) cues have been used to elicit or modulate vection. The enhancement of vection by multisensory cues (e.g., Keshavarz et al. 2014; Murovec et al. 2021; Riecke et al. 2011, see Riecke et al. 2023 for a review on multimodal vection) underlines the multisensory nature of the phenomenon.
Understanding how humans process sensory information on motion can expand our knowledge on how humans perform functionally significant tasks. When we spatially orient ourselves, we utilize perspective switching which vection seems to facilitate (Riecke et al. 2015). Further understanding the processing of sensory motion information can help us understand whether vection and visually induced motion sickness (VIMS) are related, since mixed findings are reported in scientific literature (e.g., see Kuiper et al. 2019; Nooij et al. 2017) regarding their relationship. The complex relationship between vection and VIMS (see Keshavarz et al. 2015 for a review) requires further research to understand, for example, whether vection elicits VIMS. Understanding whether vection elicits VIMS can further improve methods to mitigate the negative effects VIMS can induce on virtual reality (VR) users. When VIMS is experienced in VR, it is often referred to as Cybersickness (CS; Keshavarz et al. 2015; Laessoe et al. 2023). Vection, however, appears to be a desirable sensation when using VR applications as positive correlations have been found between vection and presence (e.g., Riecke et al. 2005a; Kim et al. 2022; Kooijman et al. 2023a). In return, the fidelity of VR applications could be enhanced through vection research (Hettinger et al. 2014).
Besides these benefits in studying vection, there are some limitations in current vection literature (e.g., see Berti and Keshavarz 2020; Kooijman et al. 2022a; Väljamäe, 2009 for more information), such as the lack of an appropriate description of vection (Palmisano et al. 2015) and methodological issues surrounding the measurement of vection (see Kooijman et al. 2023b for a review). Herein, we focused on two factors in vection research which have been relatively understudied. First, the influence of proprioceptive cues on vection has received relatively little attention in literature. Second, there appears to be ample evidence on 1) the negative relationship between presence and CS (see Weech et al. 2019 for a review) and 2) the positive relationship between presence and vection (e.g., Kim et al. 2022; Kooijman et al. 2022b; Riecke et al. 2005a), however, there are few studies that have investigated the relationship between CS, presence, and vection concomitantly. The subsequent sections discuss these two points in more detail.
1.1 Proprioception, self-motion perception, and vection: the potential of neck muscle vibrations
In principle, the perception of physical self-motion originates from the integration of a variety of sensory signals, such as, visual, vestibular, and somatosensory cues (Greenlee et al. 2016). Optic flow perceived through the visual system can provide information about heading (Britten 2008; Lappe et al. 1999), whereas vestibular input presents information about linear and rotational accelerations (Britton and Arshad 2019). Further, proprioceptive signals can provide information about the movement of the body or head (Cullen and Zobeiri 2021). Together these inputs are used by the brain to differentiate between self-motion, motion of objects, and/or retinal slip due to eye or head movements (Greenlee et al. 2016). For example, neck proprioception can provide information about the movement of the head relative to the body and/or world. Since the vestibular apparatus can only sense head motion, the integration of vestibular and neck proprioceptive signals facilitate human perception of body motion separately from head motion (Cullen 2011; Mergner et al. 1991). Notably, the co-occurrence of these different sensory signals does not necessitate self-motion perception as previous research has shown that people can adequately estimate self-motion parameters from visual or vestibular signals alone (e.g., see Campos et al. 2014; Harris et al. 2000). Nonetheless, perturbing individual and combined sensory systems can provide us an understanding of how sensory signals are integrated by humans to obtain a coherent percept of self-motion.
One way to perturb the proprioceptive system is by means of muscle vibrations. During the focal vibration of muscles, primary afferents in muscle spindles are activated (Taylor et al. 2017). Muscle spindles are sensory organs that lie parallel to muscle fibers and relay the lengthening and stretching rate of muscles to the central nervous system (Kröger and Watkins 2021; Taylor et al. 2017). Neck muscle vibrations (NMVs) have been used to study the role of neck proprioception in self-motion processes, where NVMs have been shown to influence posture and spatial perception (see Jamal et al. 2020; Pettorossi and Schieppati 2014, for reviews). For example, Panichi et al. (2011) applied whole body yaw rotations to ten participants who stood on a platform in a dark room. Participants were subjected to sinusoidal or asymmetric oscillatory rotations and had to track the remembered position of a visual target in the room. In some conditions their head was fixated using a head clamp. In these fixated conditions, NVMs with a frequency of 100 Hz were applied to two different regions of participants’ neck, either to the dorsal neck muscles or the sternocleidomastoid (SCM) muscle. When NVMs were applied ipsilaterally to the SCM during asymmetric rotations, participants’ tracking error to a remembered position in the room decreased from 69° (S.D. = 21°; no vibration) to 37° (S.D. = 19°), whereas contralateral vibration increased the tracking error to 107° (S.D. = 25°). Bove et al. (2001) applied vibrational stimulation with a frequency of 80 Hz to the sternomastoid (i.e., sternocleidomastoid) muscle of nine participants while they performed a walking task. Participants walked a shorter distance and exhibited more sway when vibrations were applied while walking or just before walking compared to conditions where no vibrations were applied.
Besides affecting posture and spatial orientation, NVMs have also been shown to elicit illusions (see Taylor et al. 2017 for a review). Participants focusing on a visual target in a dark room have been found to see the target displace leftwards or rightwards when neck muscles were vibrated (Biguer et al. 1988; Taylor and McCloskey 1991). Furthermore, Lackner and Levine (1979) identified that unilateral vibrational stimulation of the sternocleidomastoid muscle elicited an illusory sensation of rotation/tilt of the head to the opposite side. Thus, it appears neck proprioception plays a significant role in self-motion perception and can elicit illusory sensory experiences. Nonetheless, the role of neck proprioception in illusory self-motion (vection) has, to the best of our knowledge, not been studied. Understanding whether perturbation of neck proprioception affects spatial orientation during vection as it would during physical self-motion can further our understanding on whether vection and self-motion share neural pathways and are integrated similarly.
Notably, proprioceptive and vibrotactile stimulation have been used in vection research independently of one another. Specifically, the role of arthrokinetic (Bles et al. 1995; Brandt et al. 1977) and podokinetic (Becker et al. 2016; Bles and Kapteyn 1977) stimulation in vection have been studied, where the passive and active movement of arms and legs have been found to elicit or enhance sensations of self-motion. In the same vein, vibrations applied to the skin (i.e., vibrotactile stimulation) of, for example, the feet, buttocks, legs, lower back and torso, ranging in frequency from 5 to 240 Hz, have been used to elicit or modulate vection (see Kooijman et al. 2022a for a review). Although these frequencies span the range of mechanoreceptors in the skin (Gardner and Kenneth 2000) and could thus provide tactile feedback, it is possible these vibrotactile cues might have had a compounded effect of cutaneous (i.e., skin) and kinaesthetic sensations. Specifically, there is evidence that proprioceptive illusions of movement (i.e., kinaesthesia) can occur in response to vibrational frequencies of approximately 80 Hz (see Roll and Vedel 1982; Seizova-Cajic et al. 2007) and lower frequencies can elicit a proprioceptive illusion of displacement (Taylor et al. 2017). The different frequencies of the vibrotactile cues used in the studies reviewed by Kooijman et al. (2022a) may have caused a stimulation of the underlying musculature, which, in turn, could have unintentionally influenced participants’ experience of vection by eliciting an illusion of displacement or movement of the affected body part.
In sum, there appears to be a link between neck proprioception and self-motion processing, however, there appears to be no study that investigated whether neck proprioception might influence vection. Herein, it is studied whether the perturbation of neck proprioception alters participants’ experience of vection.
1.2 The concurrent measurement of cybersickness, presence, and vection
In vection studies that utilise VR technology to immerse participants, vection measures are mostly complemented by the recording of either presence (Kim et al. 2022; Lind et al. 2016) or CS (e.g., Palmisano et al. 2017; Pöhlmann et al. 2022, however, to the best of our knowledge, only relatively few studies have recorded vection, presence, and CS concurrently. For example, seventy-six participants in the study conducted by Luu et al. (2021) wore a head-mounted display (HMD) and viewed various optic flow patterns, which differed in speed. After cessation of the optic flow presentation, vection strength, presence, and CS were rated by the participants using different single-item numerical scales. Although no correlations between the three measures were presented in the manuscript, the authors did find that vection intensity, presence, and CS increased when optic flow speeds increased.
Thirty-eight participants in the study conducted by Fauville et al. (2021) stood (N = 20) or floated in a pool (N = 18) while they wore an HMD through which they perceived to be swimming. After immersion participants rated CS, presence, and vection. Nonetheless, the correlations between CS, presence, and vection found by the authors were below the critical value for Pearson’s r and therefore not indicative of a substantial association. Kooijman et al. (2023a) immersed twenty-nine participants in different virtual environments depicting either optic flow or a train carriage. After each trial, participants rated vection intensity, CS, and presence using single-item measures. A strong positive association was found between presence and vection in the optic flow condition without auditory feedback, whereas only vection convincingness and presence were associated in the optic flow condition with auditory feedback. Correlations between vection, presence, and CS in all other conditions were below the critical value for Pearson’s r and therefore not indicative of a substantial association. The lack of an association between CS and other measures in this study could be explained by the fact that relatively few participants reported symptoms of discomfort and the discomfort they did report was relatively mild. Additionally, the varying association between presence and vection could be because of the effect of the different virtual environments on vection and presence.
In sum, it appears there is limited evidence on the relationship between vection, presence, and CS. Studies have solely reported on self-reported measures (e.g., vection intensity), whereas the relationship between presence, CS, and the temporal characteristics of vection have not been scrutinized extensively. Furthermore, studies have used different methods to quantify CS, presence, and vection, which may hamper cross-comparability.
1.3 Goals of current study
The following study had two main goals. Based on the literature describing a link between neck muscle proprioception and self-motion control (e.g., Bove et al. 2001; Ivanenko et al. 1999; Panichi et al. 2011) and a link between vection and self-motion control (e.g., Palmisano et al. 2015; Riecke et al. 2015), we aimed to investigate whether neck muscle vibration could affect participants’ experience of vection in virtual reality. Specifically, neck muscle proprioception has shown to play a role in balance, gait, and self-motion perception (see Pettorossi and Schieppati 2014 for a review) and (neck) muscle vibrations have shown to induce various types of illusions (see Taylor et al. 2017 for a review). Thus, we aimed to investigate whether neck muscle vibrations would influence participants’ experience of vection and investigate whether neck muscle vibrations would improve participants’ estimation of self-motion parameters.
We hypothesized that synchronized neck muscle vibrations would increase participants’ vection experience, enhance participants’ estimation of self-motion parameters, and increase their level of immersion compared to no neck muscle vibrations or asynchronous neck muscle vibrations. The hypotheses would be operationalized by higher self-reported vection ratings and a quicker and prolonged vection experience, lower errors when estimating self-motion parameters, and higher self-reported ratings of virtual presence in conditions with synchronized neck muscle vibrations compared to conditions with no neck muscle vibrations or asynchronous neck muscle vibrations.
Further, we aimed to investigated how cybersickness (CS), presence, and vection are related in virtual reality. We hypothesized that CS and presence would be negatively related, CS and vection would be negatively related, and that presence and vection would be positively related.
2 Method
Deakin University Human Research Ethics Committee approved this study (2021-181) and prior to the start of the experiment written informed consent was acquired from participants. Participants were not offered any incentives (e.g., money or course credits).
2.1 Sample
Due to the absence of studies investigating the influence of muscle vibrations on vection, we estimated the sample size based on 1) the effect of vibrotactile stimulation on vection and 2) the effect of neck muscle vibration on participants’ self-motion perception. Firstly, Kooijman et al. (2022a) found from their meta-analysis that tactile vibrations had a general positive effect (r = 0.385, 95% confidence interval (CI) = [0.145, 0.626]) on participants’ vection. This general positive effect would, in a within-subject repeated-measure design, equate to a large effect (\({\eta }_{p}^{2}= {r}^{2}={ .385}^{2}= .15\), 95% C.I. = [0.02, 0.39]) of tactile vibration on vection. Secondly, the tracking error of participants in the study by Panichi et al. (2011) during no, lateral, and contralateral neck muscle vibration was 69° (S.D. = 21°), 37° (S.D. = 19°), and 107° (S.D. = 25°), respectively. The effect size from a one-way ANOVA using these summary data shows a very large effect (F(2,27) = 25.82, p < 0.001, \({\eta }_{p}^{2}= .66\)) of vibrations on participants’ spatial orientation. Based on these findings, we estimated that neck muscle vibrations would have a large effect on vection (i.e., \({\eta }_{p}^{2}> .14)\). A power-analysis (MorePower60; Campbell and Thompson 2012) showed that a minimum of 32 participants were needed to detect a large effect with 80% power in a two by three, within-subject, repeated-measures design.
Therefore, 35 (F = 6, M = 29) participants were recruited from the staff and student body of the Institute for Intelligent Systems Research and Innovation at Deakin University as well as through snowball sampling. The first two participants piloted our experiment after which we made modifications to reduce the duration of the experiment and change the characteristics of the neck muscle vibrations. From the remaining 33 participants, only the data of 32 (F = 6, M = 26) participants were retained due to some technological errors. Our final sample had a mean age of 30.5 years (SD = 6.8, min = 22, max = 45).
2.2 Materials
A desktop computer running a 64-bit version of Windows 10 Pro (version 22H2) with Intel(R) Core(TM) i9-10900 K CPU @ 3.70 GHz, NVIDIA GeForce RTX 3090 24 GB graphics card, Z490 AORUS ELITE MOBO, and 64 GB RAM @ 3200 MHz was used to run the X-Plane 12 (v12.03) flight simulator with the FlyWithLua plugin (v2.8.4). Participants were immersed into the cockpit of a Cessna Skyhawk G1000. A custom written LUA script controlled the movement of the aircraft and presented instructions and questions to the participants. The participants were presented with visual feedback via the Varjo V3 head-mounted display (HMD, Firmware Version 0.17.0.469). The integrated eye-tracker of the HMD recorded participants’ pupillary behaviour at 200 Hz using the software provided with the native software development kit (v3.8.0) of the HMD. The HMD ran using Varjo Base software (v3.8.1.9) and was tracked by two SteamVR Base Stations (v2.0). A custom version of OpenComposite was used instead of SteamVR to facilitate communication between the HMD and X-Plane. Auditory feedback was provided using the Sony WH-1000XM4 closed-back and noise-cancelling headphones. Neck stimulation was facilitated using a custom-built mechatronic set-up. A microcontroller (Arduino Uno) was running a modified version of XPLDirect,Footnote 1 which monitored X-Plane variables. The X-Plane variables were modified by the LUA script that controlled the flight simulator. The microcontroller monitored these variables via serial communication and actuated an eccentric rotating mass actuator (Model: Z7AL2B1692082) via a driver controller (Brand: MKRVRS, Model: TC78H66FTG) using pulse width modulation. The driver controller received external power through a power supply unit (Wavecom Instruments PS3005D) set at 3.3V and 1 A with over-volt and over-current protection turned on. Please see the link in the “Availability of data and materials” section for the code controlling X-Plane and the Arduino, as well as photos of the wiring of the Arduino. Additionally, participants wore a small harness underneath their clothing that contained the Equivital EQ02. Finally, participants used a Thrustmaster HOTAS™ Warthog Flight Stick to provide feedback on the onset and duration of vection during stimulus presentation. The experimental setup is shown in Fig. 1.

The experimental set-up of the study. A participant is seated in the NLR motion platform while wearing a Varjo V3 head-mounted display, Sony WH-1000XM4 closed-back and noise-cancelling headphones and the eccentric rotating mass actuators on each side of the neck. The actuators were fastened to the skin using sport tape and located on top of the sternocleidomastoid muscle approximately 6 cm below the mastoid. Participants used a HOTAS joystick to indicate their vection experience and wore the Equivital EQ02 underneath their clothing.
2.3 Design
The aircraft was programmed to fly over part of the Everglades in Florida, US, at an altitude of 25.91 m (i.e., 85 feet) with a speed of 166.68 km/h (i.e., 90 knots), and starting at 25.7860° latitude and − 81.0935° longitude facing at 90° east. The location was chosen as it is a relatively flat location with dense forestry without any clear landmark to orient to, which would be akin to a virtual environment presenting optic flow by means of a point cloud (e.g., see Churan et al. 2017, for lamellar optic flow cues). The clear weather pre-set in X-Plane was used. Figure 2A depicts the viewpoint of the participant from the cockpit of the aircraft. The navigation computer in the aircraft was turned off to prevent the participants from seeing the heading of the aircraft and the flight trajectory. Furthermore, the yoke was hidden and the compass in the cockpit was fixed to indicate ‘N’ throughout the flight to prevent the participants from knowing the flight heading.

Experimental design parameters. A. The view participants had from within the cockpit of the aircraft. B. The roll rate of the aircraft and stimulation frequency of the actuators plotted over the duration of the experiment for conditions in which the aircraft turned 50° C. The five different flightpaths. Flightpaths 1–5 indicate the path the aircraft flew when it turned − 110, − 50, 0, 50 and 110°, respectively. D. The roll rate of the aircraft and stimulation frequency of the actuators plotted over the duration of the experiment for conditions in which the aircraft turned 110°
The research design was within-subjects and comprised 3 independent variables (IVs). The first IV was the turning angle of the aircraft. The aircraft flew in a straight line for 20 s. After 20 s, the aircraft would turn either 0°, 50, or 110°, after which it would continue to fly until 80 s had passed from the start of the experimental trial. In the event the aircraft did not turn a corner, it flew straight ahead for 60 more seconds. This condition was utilized as a baseline condition.
The second IV was the direction in which the aircraft turned, which was either to the left or right. Figure 2C shows a top-view of the flightpaths plotted on a Google Earth snapshot of the flight location. Left turns were indicated by using a negative sign for the degrees (e.g., − 50°). Please see the link in the “Availability of data and materials” section for a video of the left and right turning conditions.
The third IV was the use of neck vibrations, which had three levels: none, synchronized, or asynchronized. Vibrations were applied on either the left or the right sternocleidomastoid. Specifically, for each participant, two vibrators were used where a single vibrator was placed on either side of the neck. In previous studies, researchers have placed actuators on the sternocleidomastoid muscle at distances ranging between 4 to 8 cm from the mastoid (Bove et al. 2001; Panichi et al. 2011). The vibrator was placed approximately 6 cm underneath the mastoid (see insert Fig. 1). The placement of the actuator was performed as following. Participants sat in a relaxed position on a chair with their gaze facing forward and no inclination of the neck. The sternocleidomastoid was palpated to identify the muscle belly. Subsequently, the mandibular yaw was palpated to identify the angle of the mandibular (i.e., the point where the ramus and the body connect). A ruler was used to measure 4 cm from the angle of the mandibular bone along the sternocleidomastoid, which was marked with pen on sports tape that was stuck to participants’ neck. The placement of the vibrator on the soft neck tissue was chosen to minimize transmission of vibrations to the skull, which could potentially affect the visual and vestibular system (Seizova-Cajic et al. 2006). The vibrational frequency was roll-rate dependent and ranged from 43 to 100 Hz. During pilot testing, participants found that fixed frequency vibrations that were synchronous to airplane yaw movement felt unnatural and distracting. It was suggested to couple the vibrations to the roll rate to create coherence with the visual feedback, since the aircraft rolled prior to yawing. Since Lackner and Levine (1979) found that unilateral vibration of the sternocleidomastoid elicited an illusory sensation of rotation/tilt of the head, which would correspond more to visual roll than visual yaw, it was decided to couple the vibrations to roll-rate.
The vibrator was controlled by means of pulse width modulation (PWM), as can be seen from Eq. (1). The input signal to the vibrator was determined by dividing the absolute roll rate \((\dot{\emptyset })\) by the maximum roll rate \(\left( {\dot{\emptyset }_{max} = 4.307 \;{\text{deg}}/{\text{s}}} \right)\) of the aircraft during turning and multiplying this fraction by the pulse width that equated to a vibration frequency of 100 Hz \(({PWM}_{100}=180 \; {\text{Hz}})\). The resulting number was rounded to the nearest integer value and sent as an input to the driver controller to actuate the vibrator.
The timing of the vibrations depended on the moment the aircraft rolled, as shown in Fig. 2B, D for 50- and 110° turning angle, respectively. When vibrations were synchronized, they were applied on the side of the neck that was concurrent to the rolling direction of the aircraft. This means that if, from the participants’ point of view, the aircraft rolled clockwise to turn right, the right sternocleidomastoid was vibrated, and if the aircraft rolled counterclockwise, the left sternocleidomastoid was vibrated. Asynchronous vibration occurred when the neck muscle on the opposite side of the rolling direction was vibrated. For example, when the aircraft rolled clockwise to turn right, the left neck muscle was vibrated.
In total, participants were presented with a unique randomized order of the 13 (2 Turning Angles × 2 Turning Directions × 3 Neck Vibrations + 1 Straight Flight) conditions. The virtual conditions lasted in total about (13 * 80 s) / 60 s = 17.33 min.
2.4 Procedure
A member of the research team welcomed the participants and informed them about the goal of the experiment, namely, to “investigate if and how humans experience movements in virtual environments and how vibrations might affect this experience”. Participants then completed a COVID-19 screening form, were asked to read a plain language statement, and signed a consent form. After the completion of this paperwork, participants filled out a short questionnaire containing questions about demographics, gaming experience, simulator use, and current well-being. A detailed description of the questions can be found in the supplementary materials. Subsequently, the experimenter marked the location on the participants’ neck where the vibrators would be placed using sport tape (see insert Fig. 1). Next, participants were instructed on how to put on the Equivital harness on top of their clothing and then went to a private room where they attached the device underneath their clothing. The participants returned to the experiment room and the experimenter attached an auxiliary electrodermal sensor to the participants’ left hand.
Once participants were seated on the motion platform, they were shown how to use the joystick and were instructed that after each flight they were to rate a series of questions. One question required participant to rate how many degrees they perceived to have turned around the vertical axis, and this was exemplified by the experimenter in the real-world (see supplementary materials for a detailed description).
Previous research has shown that participants can estimate their physical angular displacement with reliable accuracy regardless of the angular velocity (Jürgens et al. 1999). Furthermore, neck muscle vibrations have been shown to influence participants’ performance on point-to-target tasks during physical displacement (Panichi et al. 2011), however, participants’ performance on point-to-origin tasks in VR have been found to elicit large errors (Riecke 2012). Therefore, we decided to incorporate the estimation of turning angle task used by Jurgens et al. (1999) to identify whether participants can perform this self-motion perception task in VR and whether neck vibrations influence participants performance on this task.
Once participants indicated they understood the course of the flights and the rating question, the experimenter attached the neck vibrators on the marked locations on the neck using sport tape. Vibrations of 43 Hz and 100 Hz were presented to the left and right side of participants’ neck to verify their comfort with the stimulation.
After familiarization with the equipment, the participants put on the HMD and two calibrations were performed. First, the position of the HMD was calibrated and then a 9-point calibration was used to calibrate the eye tracker. After the calibration, a 2-min baseline was recorded where participants sat quietly. After the baseline, the experimenter handed participants the noise-cancelling headphones and launched X-Plane.
A practice trial was initiated in which participants were familiarized with the tasks of the experiment. Written task instructions were presented via the HMD to prevent the experimenter from instilling a potential bias, as previous research has shown that task instruction can affect vection (Palmisano and Chan 2004). The task instructions can be found in the supplementary materials. Briefly, participants were instructed that during the flight they should indicate whether they felt that they were moving by pressing the ‘fire’ button on the joystick. If participants did not feel they were moving, they were instructed to release the ‘fire’ button or not press it at all.
The practice trial consisted of brief flight in the surrounding area of Avalon airport where the aircraft banked after 20 s and flew in a turn rotating 180° around the yaw axis. After the practice trial, the participants were asked to indicate 1) how many degrees they turned, 2) vection intensity, 3) vection convincingness, 4) CS, and 5) presence. Participants were presented with five statements and rated these verbally. The experimenter recorded the responses of participants in an Excel sheet.
The first statement regarding the perceived turning angle was rated using any number from − 180 to + 180 to indicate the degrees participants perceived they had turned leftwards (negative degrees) or rightwards (positive degrees), respectively. Participants then rated their vection experience using any number on a 0–100 scale. The statements for vection intensity and convincingness were “Please rate how strong your feeling of movement was” and “Please rate how convincing your feeling of movement was”. The wording of these statements was derived from previous research on vection terminology (Soave et al. 2021) and these rating statements have been used by the research team in a previous study (Kooijman et al. 2023a). The left and right anchors of the vection intensity and convincingness ratings were “0—not at all” and “100—extremely intense,” and “0—not at all” and “100—extremely convincing”. Furthermore, participants were prompted to rate their CS via the statement “Please rate how you felt during the flight” using the Fast Motion Sickness Scale (FMS; Keshavarz and Hecht 2011). The scale ranged from 0 to 20, where the left anchor was “0 – no sickness at all” and the right anchor was “20 – severe sickness”. Lastly, participants indicated their virtual presence by rating the statement “In the computer generated world I had a sense of 'being there'’”, which was obtained from the questionnaire developed by Slater and Usoh (1993). Participants could pick any number from 0 to 100, where the left and right anchors were “0—not at all” and “100—very much”, respectively. After completing these questions, participants were asked if they understood the tasks in the experiment and if they had any questions.
When participants indicated that they understood the course of the experiment, the experiment was initiated. Participants were presented with the same task instructions as during the practice trial and could verbally indicate that they were ready to proceed. The procedure of the practice trial was then repeated for each unique combination of IVs, where participants were presented with a virtual flight and answered questions subsequently. The presentation order of the IVs was randomized for each participant. After each trial, participants were queried whether they wanted a break or proceed to the next trial. After completing all the experimental trials, the simulator was closed.
The experimenter assisted the participants with the removal of the headphones, HMD, and the vibrators, and the experimenter offered the participants some water. Participants then removed the Equivital harness in private and, once they indicated that they were ready to proceed, a short post-experiment questionnaire was administered. Finally, after completing the questionnaire, participants were thanked for their participation and provided the opportunity to ask questions about the experiment. The total duration of the experimental procedure was about 1 h and 15 min.
2.5 Data reduction and analysis
The joystick button data and questionnaire responses were read in a custom MATLAB script (see the link in the "Availability of data and materials" section for the script and data), and time-series data were synchronized. All data reduction were performed using MATLAB whereas the statistical comparisons were performed in either SPSS (28.0.1.0. v142) or MATLAB. The data reduction steps, and the computation of dependent variables are detailed per data source. Dependent variables were computed for every trial.
2.6 Questionnaire data
The estimations of the turning angle were processed to compute the error and absolute error of the estimations. First, angle estimations that were smaller than − 180 and larger than 180 were excluded from the analysis since these values fell outside of the scale shown to participants. Next, estimation errors were computed by taking the difference between participants’ estimations and the actual turning angle (TA) of the airplane, as can be seen in Eq. (2). Absolute estimation errors were computing by taking the absolute value of the estimation error.
2.7 Joystick button data
Joystick button data were linearly interpolated to achieve a constant sampling frequency of 100 Hz. The data were then filtered using a moving average filter with a window size of (100 Hz / 60 s) = 2 samples. From the button press data, the following dependent variables were computed.
-
1. Vection onset time (VOT in seconds): The difference between the moment the trial started (ttrial,start) and the moment participants first pressed the button (i.e., the moment the button data exceeded 0; \({t}_{bd > 0}\) Riecke et al. 2005b; Kooijman et al. 2023a), as can be seen from Eq. (3). If the participant did not press the joystick button, a VOT of 80 s was used.
$$VOT= {t}_{bd\,>\,0}-{t}_{trial, start}$$(3) -
2. Vection duration (VD in seconds): The total number of samples where participants pressed the button (i.e., the button data exceeded 0; \({BD}_{i}>0\)) divided by the sampling frequency (Fs) (Kooijman et al. 2023a, b), as can be seen from Eq. (4).
$$VD=\frac{\sum_{i=1}^{n}{BD}_{i}>0}{Fs}$$(4)
A vection percentage was computed for each condition based on the number of participants from the total sample who indicated that they experienced vection by pressing the joystick button. In summary, a combination of five vection measures (i.e., intensity, convincingness, onset time, duration, percentage) were recorded to capture the different aspects of vection (Kooijman et al. 2023b).
2.8 Statistical comparisons
There has been ample criticism in scientific literature on the interpretation of test results that have a p-value higher than the alpha level as a null-effect. This criticism has emphasized the necessity to interpret results in a non-dichotomous manner (e.g., Amrhein et al. 2019; McShane et al. 2019; Wasserstein et al. 2019). Herein, we address this issue by reporting within-subject compatibility intervals alongside point estimates and standard deviations to aid readers in interpreting differences in the data. The compatibility intervals denote the range of effect sizes that, given our statistical assumptions (e.g., an alpha level of 0.05), are most compatible with our data (e.g., see Amrhein and Greenland 2022). Conversely put, compatibility intervals denote the range of plausible values surrounding the mean value that are compatible with our recorded data based on our statistical assumptions. Compatibility intervals were computed by subtracting the mean score of all conditions per participant for each variable (e.g., see Morey 2008).
Nonetheless, to further assist the readership in interpreting the differences between dependent variables, conventional inferential statistical comparisons were conducted at the level of participants. We created a linear mixed-effect model using the Maximum Likelihood method, where the intercept for each participant was random and the independent variables and their interactions were fixed effects. The formula was as follows:
A Type III Test of Fixed Effects (T3TFT) was performed using the linear mixed-effect model in SPSS to investigate the effect of turning angle (TA), turning direction (TD), and vibration (V), and their respective interactions on the dependent variables (DV). The results were discussed based on the respective effect size, which were reported using partial eta squared (\({\eta }_{p}^{2}\)). Small, medium, and large effects were identified by \({\eta }_{p}^{2}\) ≥ 0.01, \({\eta }_{p}^{2}\) ≥ 0.06, and \({\eta }_{p}^{2}\) ≥ 0.14, respectively (i.e., Cohen’s f ≥ 0.10, f ≥ 0.25, and f ≥ 0.40, respectively; Cohen 1988). Additionally, post-hoc comparisons on the estimated marginal means for each IV of the LME were conducted using SPSS. Bonferroni correction was applied for multiple comparisons. Please note, figures obtained from SPSS do not present compatibility intervals but conventional 95% confidence intervals as SPSS does not allow for the computation of compatibility intervals. Associations between subjective dependent variables were modelled using a linear mixed-effect model in MATLAB, where the slope and intercept for the independent variable was random including a random intercept for participants. The formula was as follows:
In Eq. (6), the DV and IV were the subjective dependent variables vection intensity, vection convincingness, vection onset time, vection duration, estimation error, absolute estimation error, presence, and cybersickness. The alpha level was set at 0.05, however, it was Bonferroni corrected for multiple comparisons where applicable.
3 Results
The data of 10 trials had to be excluded due to either a malfunctioning of the mechatronic set-up or X-plane software crashes. A total of 406 valid trials were retained and included in the analysis. For each condition, the number of participants associated with the dependent variable is reported. Conditions are denoted by the abbreviations of their respective independent variables (IVs), where the negative numbers indicate left turns and positive numbers indicate right turns. Furthermore, the letter “N” indicates no vibration, “S” indicates synchronous vibrations and “AS” indicates asynchronous vibrations. For example, “− 50 AS” reflects the condition where the aircraft turned leftwards by 50° and asynchronous vibrations to the neck were presented.
3.1 Vection
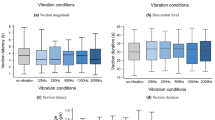
The percentage of participants reporting vection per experimental condition is shown in Fig. 3. For conditions where the aircraft turned 110°, more participants reported, on average, vection when synchronous vibration was presented compared to when no vibration was presented. Furthermore, less participants reported, on average, vection when asynchronous vibration was presented compared to no vibration and synchronous vibration. For the conditions in which the aircraft turned 50° leftwards, more participants reported, on average, vection when asynchronous vibration was presented compared to synchronous and no vibration. For 50° rightwards turning conditions, more participants reported, on average, vection when synchronous vibration was presented compared to no and asynchronous vibration. The mean, standard deviation, and compatibility interval of the vection measures are shown in Fig. 4.

Percentage of participants reporting vection per experimental condition. Grey bars indicate no vibration, blue bars indicate synchronous vibration, and red bars indicate asynchronous vibration

Mean, standard deviation, and compatibility interval of vection measures. Means (and standard deviations in parentheses) are denoted numerically for each condition next to the appropriate bar. Individual data points are marked with ‘x’. Error bars represent the compatibility intervals. Grey bars indicate no vibration, blue bars indicate synchronous vibration, and red bars indicate asynchronous vibration
3.2 Intensity
Vection intensity was, on average, the lowest in the 0 N condition, whereas it was, on average, the highest in the − 110 S condition. Intensity ratings were, on average, higher for conditions with vibrations compared to conditions without vibrations. However, the overlapping compatibility intervals for 50 and − 50 conditions suggest that there is uncertainty whether the data is compatible with an increase in vection intensity due to vibrations. For the − 110 conditions, the non-overlapping compatibility interval between no vibration and conditions with vibration suggest that there is little uncertainty whether the data is compatible with an increase in vection intensity due to vibrations. A Type III Test of Fixed Effects (T3TFT) showed a medium effect (\({\eta }_{p}^{2}\) = 0.07) of vibration on vection intensity, as indicated in Table 1. Furthermore, small (\({\eta }_{p}^{2}\) < 0.06) and negligible (\({\eta }_{p}^{2}\) < 0.01) main and interaction effects were found, however, these effects were not compatible with the rejection or confirmation of a change in vection intensity due to turning direction or turning angle.
3.3 Convincingness
Vection convincingness was, on average, the lowest in the 0 N condition, whereas it was, on average, the highest in the − 110 S condition. Convincingness ratings were, on average, higher for conditions with vibrations compared to conditions without vibrations. However, the (partially) overlapping compatibility intervals for 50 and − 50 conditions suggest that there is uncertainty whether the data is compatible with an increase in vection convincingness due to vibrations. For the − 110 conditions, the non-overlapping compatibility interval between no vibration and synchronous vibration suggest that there is little uncertainty whether the data is compatible with an increase in vection convincingness due to vibrations. A Type III Test of Fixed Effects (T3TFT) showed a medium effect (\({\eta }_{p}^{2}\) = 0.08) of vibrations on vection convincingness, as indicated in Table 1. Furthermore, small (\({\eta }_{p}^{2}\) < 0.06) and negligible (\({\eta }_{p}^{2}\) < 0.01) main and interaction effects were found, however, these effects were not compatible with the rejection or confirmation of a change in vection convincingness due to turning direction or turning angle.
3.4 Vection onset time
Vection onset time (VOT) was, on average, the highest for the 0 N condition, whereas VOT was, on average, the lowest for the condition − 110 S. VOT differed, on average, less than a second between conditions with and without vibrations when the aircraft turned 50° rightwards. However, VOT was, on average, a few seconds lower when asynchronous vibration was presented compared to synchronous and no vibration when the aircraft tuned 50° leftwards. For conditions in which the aircraft turned 110°, VOT was, on average, the lowest for synchronous vibration, followed by no vibration and asynchronous vibration, respectively. However, the (partially) overlapping compatibility intervals between conditions with and without vibration suggest that there is uncertainty whether the data is compatible with a reduction in VOT due to vibrations. A T3TFT showed small (\({\eta }_{p}^{2}\) < 0.06) and negligible (\({\eta }_{p}^{2}\) < 0.01) main and interaction effects, however, these effects were not compatible with the rejection or confirmation of a change in VOT due to turning direction, turning angle, or vibration.
3.5 Vection duration
Vection lasted, on average, the longest in the condition 110 S, whereas it lasted the shortest in the condition 50 S. For the conditions in which the aircraft turned 50°, VD was, on average, smaller for conditions with synchronous vibration compared to no vibration. Similarly, VD was, on average, smaller for the 110 S condition compared to the 110 N condition. Conversely, VD was, on average, larger for − 110S compared to − 110 N condition. Nonetheless, the (partially) overlapping compatibility intervals between conditions with and without vibration suggest that there is uncertainty whether the data is compatible with a change in VD due to vibrations. A T3TFT showed small (\({\eta }_{p}^{2}\) < 0.06) and negligible (\({\eta }_{p}^{2}\) < 0.01) main and interaction effects, however, these effects were not compatible with the rejection or confirmation of a change in VD due to turning direction, turning angle, or vibration.
3.6 Post-hoc comparisons
Figure 5 shows the estimated marginal mean and 95% confidence interval for each of the vection measures per independent variable. Vection intensity and convincingness were, on average, lower for conditions without vibration compared to conditions with vibration. VOT was, on average, lower for synchronous vibration compared to no vibration followed by asynchronous vibration. However, VD was, on average, higher for no vibration, followed by synchronous and asynchronous vibration, respectively. Post-hoc comparisons of the estimated marginal means are denoted in Table 2. The positive effect of synchronous and asynchronous vibration on vection intensity and convincingness ranged from small (\(0.2 \le d<0.5\)) to very small (\(0.0 \le d<0.2\)). However, asynchronous vibration appeared to reduce vection convincingness compared to synchronous vibration (d = 0.14). Post-hoc comparisons between conditions with and without vibrations shown in Fig. 4 are reported in Table S1 in the supplementary material. Furthermore, Table 2 shows a very small effect of turning angle on VD, where VD was larger for 110° turning angles compared to 50° turning angles.

Estimated marginal means, standard error, and 95% confidence interval of vection measures per independent variable. Means (and standard error in parentheses) are denoted numerically for each factor next to the appropriate bar. Error bars represent 95% confidence intervals
3.7 Estimation of turning angle
Figure 6 depicts the mean, standard deviation, and compatibility interval of turning angle estimations. Figure 6 (left) denotes the raw estimation error whereas Fig. 6 (right) depicts the absolute estimation error. The smallest raw estimation error was, on average, in the condition 0 N, whereas the largest was, on average, in the condition 110 N. For conditions where the aircraft turned 50° to the right, the estimation error was, on average, smaller for synchronous vibration compared to no vibration and asynchronous vibration. or conditions where the aircraft turned 50° to the left, the estimation error was, on average, larger for conditions with vibration compared to conditions without vibration. For conditions where the aircraft turned 110° rightwards, estimation errors were, on average smaller for condition with vibrations compared to conditions without vibration, however, the opposite trend is noticeable for the conditions where the aircraft turned 110° leftwards. Nonetheless, the (partially) overlapping compatibility intervals between conditions with and without vibration suggest that there is uncertainty whether the data is compatible with a change in estimation error due to vibrations. A T3TFT showed small (\({\eta }_{p}^{2}\) < 0.06) and negligible (\({\eta }_{p}^{2}\) < 0.01) main and interaction effects as can be seen in Table 1. However, these effects were not compatible with the rejection or confirmation of a change in estimation error due to turning direction, turning angle, or vibration.

Mean, standard deviation, and compatibility interval of turning angle estimations. Means (and standard deviations in parentheses) are denoted numerically for each condition next to the appropriate bar. Individual data points are marked with ‘x’. Error bars represent the compatibility intervals. Grey bars indicate no vibration, blue bars indicate synchronous vibration, and red bars indicate asynchronous vibration
The smallest absolute estimation error was, on average, in the condition 0 N, whereas the largest was, on average, in the condition 110 N. For conditions where the aircraft turned 50° to the right, the absolute estimation error was, on average, larger when asynchronous vibration was presented compared to synchronous and no vibration. Conversely, for conditions where the aircraft turned 50° leftwards the absolute estimation error was, on average, larger when synchronous vibration was presented compared to asynchronous and no vibration. For conditions in which the aircraft turned 110°, absolute estimation errors were, on average, lower for conditions with vibrations compared to conditions without. The largest difference can be observed between 110 N and 110 S. Nonetheless, the (partially) overlapping compatibility intervals between conditions with and without vibration suggest that there is uncertainty whether the data is compatible with a change in absolute estimation error due to vibration.
A T3TFT showed a moderate effect (\({\eta }_{p}^{2}\) = 0.12) of turning angle on absolute estimation error. Furthermore, small (\({\eta }_{p}^{2}\) < 0.06) and negligible (\({\eta }_{p}^{2}\) < 0.01) main and interaction effects were found, however, these effects were not compatible with the rejection or confirmation of a change in absolute estimation error due to turning direction or vibration.
3.8 Post-hoc comparisons
Figure 7 shows the estimated marginal mean and 95% confidence interval for each of the estimation errors per independent variable. Raw estimation errors were, on average, the smallest for synchronous vibration followed by no vibration and asynchronous vibration, respectively. Absolute estimation errors were, on average, smaller for synchronous and asynchronous vibration compared to no vibration. The largest difference was between 50 and 110° turning conditions for the absolute estimation errors where errors were, on average, smaller for 50° compared to 110° turning conditions. Post-hoc comparisons of the estimated marginal means, denoted in Table 2, show that a small difference (d = 0.38) was found between left and right turning conditions for the absolute estimation error. Post-hoc comparisons between conditions with and without vibrations shown in Fig. 6 are reported in Table S2 in the supplementary material.

Estimated marginal mean, standard error, and 95% confidence interval of turning angle estimations per independent variable. Means (and standard error in parentheses) are denoted numerically for each factor next to the appropriate bar. Error bars represent 95% confidence intervals
3.9 Presence and cybersickness
Figure 8(left) depicts the mean, standard deviation, and compatibility interval of presence ratings. Presence was, on average, the lowest in the 0 N condition, whereas it was, on average, the highest in the − 110 S condition. Presence was, on average, lower for conditions without vibration compared to conditions with vibration. However, the non-overlapping compatibility interval between conditions with and without vibration for the 50° and the − 110° turning conditions suggest that here is little uncertainty whether the data is compatible with an increase in presence due to vibrations. The largest difference can be observed between − 110 N and − 110 S. In all conditions presence ratings for asynchronous vibration were, on average, larger compared to no vibration.
A T3TFT showed a medium effect (\({\eta }_{p}^{2}\) = 0.10) of vibration on presence. Furthermore, small (\({\eta }_{p}^{2}\) < 0.06) and negligible (\({\eta }_{p}^{2}\) < 0.01) main and interaction effects were found, however, these effects were not compatible with the rejection or confirmation of a change in presence due to turning angle or direction.
Twenty-three participants reported no sickness (i.e., 0) as their strongest rating. Four participants individually reported a maximum FMS rating of 1, 3, 4 and 7, whereas four other participants reported a maximum FMS rating of 10. One participant reported a maximum FMS rating of 15. The five participants who reported an FMS rating of 10 and higher indicated they wanted to continue the experiment after they had a break. Figure 8 (right) depicts the mean, standard deviation, and compatibility interval of Cybersickness (CS) ratings. On average, CS was the lowest in the 0 N condition and highest in the 110 AS condition. Except for the conditions where the aircraft turned 110° leftwards, the addition of synchronous vibration, on average, reduced CS ratings compared to conditions without vibration. For the conditions in which the aircraft turned rightwards, CS ratings were, on average, higher with asynchronous vibration compared to no vibration. Nonetheless, the (partially) overlapping compatibility intervals between conditions with and without vibration suggest that there is uncertainty whether the data is compatible with a change in cybersickness due to vibration. A T3TFT showed small (\({\eta }_{p}^{2}\) < 0.06) and negligible (\({\eta }_{p}^{2}\) < 0.01) main and interaction effects, however, these effects were not compatible with the rejection or confirmation of a change in cybersickness due to turning direction, turning angle, or vibration.

Mean, standard deviation, and compatibility interval of presence and cybersickness ratings. Means (and standard deviations in parentheses) are denoted numerically for each condition next to the appropriate bar. Individual data points are marked with ‘x’. Error bars represent the compatibility intervals. Grey bars indicate no vibration, blue bars indicate synchronous vibration, and red bars indicate asynchronous vibration
3.10 Post-hoc comparisons
Figure 9 shows the estimated marginal mean and 95% confidence interval for presence and cybersickness per independent variable. Presence was, on average, the smallest for no vibration followed by synchronous and asynchronous vibration, respectively. Cybersickness was, on average, the smallest for synchronous vibration followed by asynchronous and no vibration, respectively. Post-hoc comparisons of the estimated marginal means, denoted in Table 2, show that a small difference was found between no vibration and synchronous vibration (d = 0.34), as well as no vibration and asynchronous vibration (d = 0.20), which suggests that presence increased due to vibration. Additionally, a small difference was found between synchronous and asynchronous vibration (d = 0.14), which suggest that asynchronous vibration reduced presence compared to synchronous vibration. Post-hoc comparisons between conditions with and without vibrations shown in Fig. 8 are reported in Table S2 in the supplementary material.

Estimated marginal mean, standard error, and 95% confidence interval of presence and cybersickness per independent variable. Means (and standard error in parentheses) are denoted numerically for each factor next to the appropriate bar. Error bars represent 95% confidence intervals
3.11 Relationship between cybersickness, presence, and vection
Table 3 shows the estimates of the random effect covariate between subjective dependent variables obtained through a linear mixed-effect model. Presence and 1) vection intensity, and 2) vection convincingness were positively associated. The trends between CS and 1) presence and 2) vection measures was not strong enough conclude an association between CS and presence or vection. Table S3 in the supplementary materials denotes the repeated-measure correlations derived via the method proposed by Bakdash and Marusich (2017), whereas Table S5 denotes Spearman’s rank correlations between subjective dependent variables averaged across experimental conditions. Both methods (i.e., repeated-measure and Spearman’s rank) present similar trends as the results shown in Table 3.
4 Discussion
This study investigated the potential enhancement of vection and self-motion parameter estimation through vibrational cues. Additionally, the relationship between CS, presence, and vection in virtual reality was studied. The data of thirty-two participants were analysed who were immersed in a flight simulator wherein they were a passenger in a small aircraft that flew specific patterns autonomously. Participants pressed a button on a joystick to indicate their experience of vection during the flights. Furthermore, after each flight, participants estimated how many degrees they had turned around the vertical axis, and rated vection intensity, convincingness, presence, and CS. Our study had two main aims, which were subdivided into the following two sections where our findings are discussed in more detail.
4.1 The effect of vibrations on vection, self-motion parameter estimations, and presence
In line with our expectations, it was found that neck muscle vibrations appeared to increase participants’ experience of vection and presence. Specifically, a medium effect of vibration on vection intensity, vection convincingness, and presence was found through inferential statistics, and for some conditions compatible increments in vection intensity, vection convincingness, and presence ratings were evident from non-overlapping compatibility intervals when vibrations were presented compared to when they were not. Post-hoc comparisons showed that synchronous and asynchronous vibrations elicited more intense and convincing vection and more immersion compared to no vibration, and vection was more convincing and more immersive when synchronous vibration was presented compared to asynchronous vibration.
Our findings are in line with previous research on proprioceptive influence on the sensation of ego-motion; Bles et al. (1995) found that a higher percentage of participants reported (congruent) self-motion sensations when the direction of illusory ego-motion elicited by hand-movements matched the movement direction of the sled they sat on, compared to when the illusory sensation by hand-movements was opposed to the movement direction of the sled. Similarly, Seno et al. (2013) found that when participants made breaststroke movements whilst viewing optic flow patterns, vection was enhanced when the illusory direction of the breaststroke movements was congruent to the illusory direction of the optic flow compared to when they were incongruent to each other. Furthermore, Seno et al. (2011) found that when the direction of treadmill locomotion was incongruent to the direction of optic flow, vection degraded.
The positive impact of neck muscle vibrations on vection in the current study can, in part, be explained by multisensory integration. To achieve a valid, coherent percept of the environment, the brain integrates, or segregates, multisensory sensory cues (Shams and Beierholm 2011; Van Atteveldt et al. 2014). When sensory signals are temporally correlated and compatible to each other, optimal sensory integration occurs (Parise and Ernst 2016). However, it is also possible that the integration process breaks down when a conflict between sensory cues is substantial enough (Greenlee et al. 2016), as exemplified by the findings from the studies mentioned above.
Nonetheless, it should be noted that our findings are not direct evidence that the neck vibrations caused an activation of the muscle spindles and therefore elicited a proprioceptive effect on vection. Firstly, no pre-tests were conducted herein that measured the known proprioceptive effect of neck muscle vibration on pointing behaviour (Han and Lennerstrand 1999; Panichi et al. 2011) or gait/locomotion (e.g., see Bove et al. 2001). Secondly, a substantial effect of vibration on self-motion parameter estimations was not found in the current study. Thus, it could be possible that merely the tactile sense was activated by the vibration of the neck, which caused the enhancement of vection. There is ample evidence in literature on the enhancement of vection through congruent vibrotactile stimulation to other sensory cues (Matsuda et al. 2021; see Kooijman et al. 2022a for a review).
Furthermore, our results show differing trends of synchronous and asynchronous vibration depending on the turning direction and angle. For example, the percentage of participants experiencing vection when synchronous vibration was presented was higher compared to asynchronous vibration during 50° rightward turns whereas more participants experienced vection when asynchronous vibration was presented compared to synchronous vibration during 50° leftward turns (see Fig. 3). Similarly, vection intensity rating were higher for asynchronous compared to synchronous vibrations during 110° rightward turns whereas the opposite was evident for 110° leftward turns. Although the differences are small, future research could further scrutinise whether parameters of curvilinear vection affect participants’ experience of illusory self-motion and how multisensory cues modulate the experience.
In sum, our results indicate that neck vibrations can enhance vection and presence, however, it is uncertain whether this effect can be attributed to proprioceptive or tactile stimulation.
4.2 The relationship between cybersickness, presence, and vection
In line with our expectations and consistent with previous research (Riecke et al. 2005a; Kim et al. 2022; Kooijman et al. 2023a), our results show a positive association between presence and vection. However, the association was mainly found for measures on the quality of vection (i.e., intensity, convincingness, see Kooijman et al. 2023b) compared to measure on the temporal characteristics of vection. The difference in strength between presence and 1) vection ratings and 2) temporal vection measures could, in part, be explained by the aspects of vection that the vection measures capture. For example, vection onset time (VOT) and vection duration (VD) predominantly capture the occurrence and extent of vection, respectively, whereas intensity and convincingness ratings capture the quality of participants’ vection experience (Kooijman et al. 2023b). Although 1) measures capturing the temporal characteristics and the quality of the vection experience tend to correlate well with one another (e.g., Kooijman et al. 2023a) and 2) even though efforts have been made to understand and model the relationship between the measures capturing the different components of vection (e.g., Seno et al. 2017), vection studies do not always identify consistently similar effects across the different measures (e.g., Keshavarz et al. 2019; Seya et al. 2015; Yahata et al. 2021). Thus, it may be that the quality of participants’ vection experience may play more of a role in the evaluation of virtual presence compared to the moment when and how long vection lasted for. Nonetheless, more research is needed to fully understand how the different components of vection might relate to participants’ experience of presence.
Our results, particularly the lack of an effect of CS on the relationship between vection and presence, could partially be attributed to the limited number of participants who experienced pronounced discomfort during our study. Specifically, five participants reported an FMS rating over 10 for a single condition after which they were forced to have a break. Moreover, these participants all volunteered to continue the experiment after the symptoms had dissipated from taking a break. Therefore, this break may have avoided any change in vection and presence due to discomfort that could have occurred otherwise if participants did not have a break.
4.3 Limitations, implications, and outlook
First, our study was limited by our sample size. We expected large positive effects of vibrations on vection, but we found moderate negative effects of vibration on vection intensity, vection convincingness, and presence. Although our study had a large enough sample to detect large effects (\({\eta }_{p}^{2}\) = 0.14), our study was not powerful enough to detect medium (\({\eta }_{p}^{2}\) = 0.06) effects. A post-hoc power calculation (MorePower60; Campbell and Thompson 2012) showed that to adequately detect medium effects, an additional 46 participants (i.e., 78 participants in total) were required to achieve 80% power. As such, caution should be exerted when interpreting these results as they may be due to a Type II Error. Nonetheless, our results theoretically imply that 1) the introduction of an additional stimulus through an unperturbed sensory modality during stimulus presentation could enhance participants’ vection and immersion when applied congruently in time and amplitude to another sensory modality, and 2) offering breaks to participants who experience CS likely does not alter their experience of vection and presence.
Future research could employ a pre-test using neck muscle vibration to ascertain a proprioceptive activation of the neck muscles occurs. Moreover, future studies could consider avoiding offering breaks to participants who experience CS to identify how changes in vection and presence might occur due to CS.
5 Conclusion
Herein, it was studied whether neck muscle vibrations could enhance illusory self-motion experienced in virtual reality during a virtual flight. Based on our findings, it was identified that vibrations applied to the neck can enhance vection and virtual presence. However, due to a lack of pre-test, it cannot be concluded whether the enhancement was due to tactile or proprioceptive stimulation. Furthermore, presence and the subjective quality of vection were found to be positively associated, however, presence and the temporal characteristics of vection did not present a cohesive association. Additionally, no strong associations between CS and presence or vection were found, which was likely due to the lack of strongly fluctuating stimuli that could induce nausea used in the current study. Future research on vection employing muscle vibrations should include rigorous pre-tests to ascertain proprioceptive activation.
Availability of data and materials
The study reported in this article has not been registered. The data and source code are accessible via: https://doi.org/10.26187/deakin.25000730.
References
Amrhein V, Greenland S (2022) Rewriting results in the language of compatibility. Trends Ecol Evol 37(7):567–568. https://doi.org/10.1016/j.tree.2022.02.001
Amrhein V, Greenland S, McShane B (2019) Scientists rise up against statistical significance. Nature 567(7748):305–307. https://doi.org/10.1038/d41586-019-00857-9
Aruga A, Bannai Y, Seno T (2019) Investigation of the Influence of scent on self-motion feeling by vection. Int J Inform Soc 11(2):65–73
Ash A, Palmisano S, Apthorp D, Allison RS (2013) Vection in depth during treadmill walking. Perception 42(5):562–576. https://doi.org/10.1068/p7449
Bakdash JZ, Marusich LR (2017) Repeated measures correlation. Front Psychol. 8:456. https://doi.org/10.3389/fpsyg.2017.00456
Becker W, Kliegl K, Kassubek J, Juergens R (2016) Podokinetic circular vection: characteristics and interaction with optokinetic circular vection. Exp Brain Res 234:2045–2058. https://doi.org/10.1007/s00221-016-4604-x
Berti S, Keshavarz B (2020) Neuropsychological approaches to visually-induced vection: an overview and evaluation of neuroimaging and neurophysiological studies. Multisens Res 34(2):153–186. https://doi.org/10.1163/22134808-bja10035
Biguer B, Donaldson IML, Hein A, Jeannerod M (1988) Neck muscle vibration modifies the representation of visual motion and direction in man. Brain 111(6):1405–1424. https://doi.org/10.1093/brain/111.6.1405
Bles W, Kapteyn TS (1977) Circular vection and human posture I. Does the proprioceptive system play a role? Agressol Rev Int Physio-Biol Pharmacol Appl Effets L’agress 18(6):325–328
Bles W, Jelmorini M, Bekkering H, de Graaf B (1995) Arthrokinetic information affects linear self-motion perception. J Vestib Res 5(2):109–116. https://doi.org/10.3233/VES-1995-5202
Bove M, Diverio M, Pozzo T, Schieppati M (2001) Neck muscle vibration disrupts steering of locomotion. J Appl Physiol 91(2):581–588. https://doi.org/10.1152/jappl.2001.91.2.581
Brandt T, Büchele W, Arnold F (1977) Arthrokinetic nystagmus and ego-motion sensation. Exp Brain Res 30(2–3):331–338. https://doi.org/10.1007/BF00237260
Britten KH (2008) Mechanisms of self-motion perception. Annu Rev Neurosci 31:389–410. https://doi.org/10.1146/annurev.neuro.29.051605.112953
Britton Z, Arshad Q (2019) Vestibular and multi-sensory influences upon self-motion perception and the consequences for human behavior. Front Neurol 10:63. https://doi.org/10.3389/fneur.2019.00063
Campbell JI, Thompson VA (2012) MorePower 6.0 for ANOVA with relational confidence intervals and Bayesian analysis. Behav Res Methods 44:1255–1265. https://doi.org/10.3758/s13428-012-0186-0
Campos JL, Butler JS, Bülthoff HH (2014) Contributions of visual and proprioceptive information to travelled distance estimation during changing sensory congruencies. Exp Brain Res 232:3277–3289. https://doi.org/10.1007/s00221-014-4011-0
Churan J, Paul J, Klingenhoefer S, Bremmer F (2017) Integration of visual and tactile information in reproduction of traveled distance. J Neurophysiol 118(3):1650–1663. https://doi.org/10.1152/jn.00342.2017
Cohen J (1988) Statistical power analysis for the behavioral sciences, 2nd edn. Academic Press
Cullen KE (2011) The neural encoding of self-motion. Curr Opin Neurobiol 21(4):587–595. https://doi.org/10.1016/j.conb.2011.05.022
Cullen KE, Zobeiri OA (2021) Proprioception and the predictive sensing of active self-motion. Curr Opin Physiol 20:29–38. https://doi.org/10.1016/j.cophys.2020.12.001
Fauville G, Queiroz A, Woolsey ES, Kelly JW, Bailenson JN (2021) The effect of water immersion on vection in virtual reality. Sci Rep 11(1):1–13. https://doi.org/10.1038/s41598-020-80100-y
Gardner EP, Kenneth OJ (2000) Chapter 23: Touch. In: Kandel ER, Schwartz JH, Jessell TM (eds) Principles of neural science. McGraw-Hill, pp 498–529
Greenlee MW, Frank SM, Kaliuzhna M, Blanke O, Bremmer F, Churan J, Cuturi LF, MacNeilage PR, Smith AT (2016) Multisensory integration in self motion perception. Multisens Res 29(6–7):525–556. https://doi.org/10.1163/22134808-00002527
Han Y, Lennerstrand G (1999) Changes of visual localization induced by eye and neck muscle vibration in normal and strabismic subjects. Graefes Arch Clin Exp Ophthalmol. 237:815–823. https://doi.org/10.1007/s004170050318
Harris LR, Jenkin M, Zikovitz DC (2000) Visual and non-visual cues in the perception of linear self motion. Exp Brain Res 135:12–21. https://doi.org/10.1007/s002210000504
Hettinger L, Schmidt-Daly TN, Jones DL, Keshavarz B (2014) Illusory self-motion in virtual environments. In: Stanney KM, Hale KS (eds) Handbook of virtual environments: design, implementations, and applications, 2nd edn. CRC Press, pp 435–465
Ivanenko YP, Grasso R, Lacquaniti F (1999) Effect of gaze on postural responses to neck proprioceptive and vestibular stimulation in humans. J Physiol 519(1):301–314
Jamal K, Leplaideur S, Leblanche F, Raillon AM, Honoré T, Bonan I (2020) The effects of neck muscle vibration on postural orientation and spatial perception: a systematic review. Neurophysiol Clin 50(4):227–267. https://doi.org/10.1016/j.neucli.2019.10.003
Jürgens R, Boß T, Becker W (1999) Estimation of self-turning in the dark: comparison between active and passive rotation. Exp Brain Res 128:491–504. https://doi.org/10.1007/s002210050872
Keshavarz B, Hecht H (2011) Validating an efficient method to quantify motion sickness. Hum Factors 53(4):415–426. https://doi.org/10.1177/0018720811403736
Keshavarz B, Hettinger LJ, Vena D, Campos JL (2014) Combined effects of auditory and visual cues on the perception of vection. Exp Brain Res 232:827–836. https://doi.org/10.1007/s00221-013-3793-9
Keshavarz B, Riecke BE, Hettinger LJ, Campos JL (2015) Vection and visually induced motion sickness: How are they related? Front Psychol 6:472. https://doi.org/10.3389/fpsyg.2015.00472
Keshavarz B, Philipp-Muller AE, Hemmerich W, Riecke BE, Campos JL (2019) The effect of visual motion stimulus characteristics on vection and visually induced motion sickness. Displays 58:71–81. https://doi.org/10.1016/j.displa.2018.07.005
Kim J, Charbel-Salloum A, Perry S, Palmisano S (2022) Effects of display lag on vection and presence in the Oculus Rift HMD. Virt Real. https://doi.org/10.1007/s10055-021-00570-x
Kooijman L, Asadi H, Mohamed S, Nahavandi S (2022a) A systematic review and meta-analysis on the use of tactile stimulation in vection research. Atten Percept Psychophys 84(1):300–320. https://doi.org/10.3758/s13414-021-02400-3
Kooijman L, Asadi H, Mohamed S, Nahavandi S (2023a) A virtual reality study investigating the train illusion. R Soc Open Sci 10(4):221622. https://doi.org/10.1098/rsos.221622
Kooijman L, Berti S, Asadi H, Nahavandi S, Keshavarz B (2023b) Measuring vection: a review and critical evaluation of different methods for quantifying illusory self-motion. Behav Res Methods. https://doi.org/10.3758/s13428-023-02148-8
Kooijman L, Asadi H, Mohamed S, Nahavandi S (2022b) A virtual reality study investigating the effect of cybersickness on the relationship between vection and presence across environments with varying levels of ecological relevance. In 2022 15th international conference on human system interaction (HSI). IEEE, pp 1–8. https://doi.org/10.1109/HSI55341.2022.9869507
Kröger S, Watkins B (2021) Muscle spindle function in healthy and diseased muscle. Skeletal Muscle 11(1):1–13. https://doi.org/10.1186/s13395-020-00258-x
Kuiper OX, Bos JE, Diels C (2019) Vection does not necessitate visually induced motion sickness. Displays 58:82–87. https://doi.org/10.1016/j.displa.2018.10.001
Lackner JR (1977) Induction of illusory self-rotation and nystagmus by a rotating sound-field. Aviat Space Environ Med 48:129–131
Lackner JR, Levine MS (1979) Changes in apparent body orientation and sensory localization induced by vibration of postural muscles: vibratory myesthetic illusions. Aviat Space Environ Med 50(4):346–354
Laessoe U, Abrahamsen S, Zepernick S, Raunsbaek A, Stensen C (2023) Motion sickness and cybersickness–sensory mismatch. Physiol Behav 258:114015. https://doi.org/10.1016/j.physbeh.2022.114015
Lappe M, Bremmer F, van den Berg AV (1999) Perception of self-motion from visual flow. Trends Cogn Sci 3(9):329–336. https://doi.org/10.1016/S1364-6613(99)01364-9
Larsson P, Västfjäll D, Kleiner M (2004) Perception of self-motion and presence in auditory virtual environments. In Proceedings of seventh annual workshop presence, Vol 2004, pp 252–258
Lind S, Thomsen L, Egeberg M, Nilsson N, Nordahl R, Serafin S (2016, March). Effects of vibrotactile stimulation during virtual sandboarding. In 2016 IEEE Virtual Reality (VR) (pp. 219–220). IEEE. https://doi.org/10.1109/VR.2016.7504732
Luu W, Zangerl B, Kalloniatis M, Kim J (2021) Effects of stereopsis on vection, presence and cybersickness in head-mounted display (HMD) virtual reality. Sci Rep 11(1):12373. https://doi.org/10.1038/s41598-021-89751-x
Matsuda Y, Nakamura J, Amemiya T, Ikei Y, Kitazaki M (2021) Enhancing virtual walking sensation using self-avatar in first-person perspective and foot vibrations. Front Virt Real 2:26. https://doi.org/10.3389/frvir.2021.654088
McShane BB, Gal D, Gelman A, Robert C, Tackett JL (2019) Abandon statistical significance. Am Stat 73(sup1):235–245. https://doi.org/10.1080/00031305.2018.1527253
Mergner T, Siebold C, Schweigart G, Becker W (1991) Human perception of horizontal trunk and head rotation in space during vestibular and neck stimulation. Exp Brain Res 85(2):389–404. https://doi.org/10.1007/BF00229416
Morey RD (2008) Confidence intervals from normalized data: a correction to Cousineau (2005). Tutor Quant Methods Psychol 4(2):61–64. https://doi.org/10.20982/tqmp.01.1.p042
Murata K, Seno T, Ozawa Y, Ichihara S (2014) Self-Motion perception induced by cutaneous sensation caused by constant wind. Psychology 5(15):1777. https://doi.org/10.4236/psych.2014.515184
Murovec B, Spaniol J, Campos JL, Keshavarz B (2021) Multisensory effects on illusory self-motion (vection): the role of visual, auditory, and tactile cues. Multisens Res 34(8):869–890. https://doi.org/10.1163/22134808-bja10058
Nooij SAE, Pretto P, Oberfeld D, Hecht H, Bülthoff HH (2017) Vection is the main contributor to motion sickness induced by visual yaw rotation: implications for conflict and eye movement theories. PLoS ONE 12(4):1–19. https://doi.org/10.1371/journal.pone.0175305
Palmisano S, Chan AY (2004) Jitter and size effects on vection are immune to experimental instructions and demands. Perception 33(8):987–1000. https://doi.org/10.1068/p5242
Palmisano S, Allison RS, Schira MM, Barry RJ (2015) Future challenges for vection research: definitions, functional significance, measures, and neural bases. Front Psychol 6:193. https://doi.org/10.3389/fpsyg.2015.00193
Palmisano S, Mursic R, Kim J (2017) Vection and cybersickness generated by head-and-display motion in the Oculus Rift. Displays 46:1–8. https://doi.org/10.1016/j.displa.2016.11.001
Panichi R, Botti FM, Ferraresi A, Faralli M, Kyriakareli A, Schieppati M, Pettorossi VE (2011) Self-motion perception and vestibulo-ocular reflex during whole body yaw rotation in standing subjects: the role of head position and neck proprioception. Hum Mov Sci 30(2):314–332. https://doi.org/10.1016/j.humov.2010.10.005
Parise CV, Ernst MO (2016) Correlation detection as a general mechanism for multisensory integration. Nat Commun 7(1):11543. https://doi.org/10.1038/ncomms11543
Pettorossi VE, Schieppati M (2014) Neck proprioception shapes body orientation and perception of motion. Front Hum Neurosci 8:895. https://doi.org/10.3389/fnhum.2014.00895
Pöhlmann KMT, Föcker J, Dickinson P, Parke A, O’Hare L (2022) The relationship between vection, cybersickness and head movements elicited by illusory motion in virtual reality. Displays 71:102111. https://doi.org/10.1016/j.displa.2021.102111
Riecke BE, Feuereissen D, Rieser JJ, McNamara TP (2015) More than a cool illusion? Functional significance of self-motion illusion (circular vection) for perspective switches. Front Psychol 6:1174. https://doi.org/10.3389/fpsyg.2015.01174
Riecke B, Murovec B, Campos J, Keshavarz B (2023) Beyond the eye: multisensory contributions to the sensation of illusory self-motion (Vection). Multisens Res. https://doi.org/10.1163/22134808-bja10112
Riecke BE, Schulte-Pelkum J, Avraamides MN, von der Heyde M, Bulthoff HH (2005a) Scene consistency and spatial presence increase the sensation of self-motion in virtual reality. In: Proceedings of the 2nd symposium on applied perception in graphics and visualization, pp 111–118. https://doi.org/10.1145/1080402.1080422
Riecke BE, Schulte-Pelkum J, Caniard F, Bulthoff HH (2005b) Towards lean and elegant self-motion simulation in virtual reality. In IEEE Proceedings. VR 2005. Virtual Reality, 2005. IEEE, pp 131–138. https://doi.org/10.1109/VR.2005.1492765
Riecke BE, Feuereissen D, Rieser JJ, McNamara TP (2011) Spatialized sound enhances biomechanically-induced self-motion illusion (vection). In Proceedings of the SIGCHI conference on human factors in computing systems. pp 2799–2802. https://doi.org/10.1145/1978942.1979356
Riecke BE (2012) Are left-right hemisphere errors in point-to-origin tasks in VR caused by failure to incorporate heading changes?. In International conference on spatial cognition. Springer Berlin Heidelberg, Berlin , pp. 143–162
Roll JP, Vedel JP (1982) Kinaesthetic role of muscle afferents in man, studied by tendon vibration and microneurography. Exp Brain Res 47(2):177–190. https://doi.org/10.1007/BF00239377
Seizova-Cajic T, Sachtler WB, Curthoys IS (2006) Eye movements cannot explain vibration-induced visual motion and motion aftereffect. Exp Brain Res 173:141–152. https://doi.org/10.1007/s00221-006-0373-2
Seizova-Cajic T, Smith JL, Taylor JL, Gandevia SC (2007) Proprioceptive movement illusions due to prolonged stimulation: reversals and aftereffects. PLoS ONE 2(10):e1037. https://doi.org/10.1371/journal.pone%3c.0011037
Seno T, Ito H, Sunaga S (2011) Inconsistent locomotion inhibits vection. Perception 40(6):747–750. https://doi.org/10.1068/p7018
Seno T, Funatsu F, Palmisano S (2013) Virtual swimming—breaststroke body movements facilitate vection. Multisens Res 26(3):267–275. https://doi.org/10.1163/22134808-00002402
Seno T, Sawai KI, Kanaya H, Wakebe T, Ogawa M, Fujii Y, Palmisano S (2017) The oscillating potential model of visually induced vection. i-Perception 8(6):2041669517742176. https://doi.org/10.1177/2041669517742176
Seya Y, Shinoda H, Nakaura Y (2015) Up-down asymmetry in vertical vection. Vision Res 117:16–24. https://doi.org/10.1016/j.visres.2015.10.013
Shams L, Beierholm U (2011) Chapter 13: Humans’ multisensory perception, from integration to segregation, follows Bayesian inference. In: Trommershauser J, Kording K, Landy MS (eds) Sensory cue integration. Oxford University Press, United Kingdom, pp 251–262
Slater M, Usoh M (1993) Representations systems, perceptual position, and presence in immersive virtual environments. Presence Teleoperat Virt Environ 2(3):221–233. https://doi.org/10.1162/pres.1993.2.3.221
Soave F, Padma Kumar A, Bryan-Kinns N, Farkhatdinov I (2021) Exploring terminology for perception of motion in virtual reality. In: Designing interactive systems conference 2021. Association for Computing Machinery, pp 171–179. https://doi.org/10.1145/3461778.3462064
Taylor JL, McCloskey DI (1991) Illusions of head and visual target displacement induced by vibration of neck muscles. Brain 114(2):755–759. https://doi.org/10.1093/brain/114.2.755
Taylor MW, Taylor JL, Seizova-Cajic T (2017) Muscle vibration-induced illusions: review of contributing factors, taxonomy of illusions and user’s guide. Multisens Res 30(1):25–63. https://doi.org/10.1163/22134808-00002544
Tinga AM, Jansen C, van der Smagt MJ, Nijboer TCW, van Erp JBF (2018) Inducing circular vection with tactile stimulation encircling the waist. Acta Physiol (oxf) 182:32–38. https://doi.org/10.1016/j.actpsy.2017.11.007
Väljamäe A (2009) Auditorily-induced illusory self-motion: a review. Brain Res Rev 61(2):240–255. https://doi.org/10.1016/j.brainresrev.2009.07.001
Väljamäe A, Larsson P, Västfjäll D, Kleiner M (2009) Auditory landmarks enhance circular vection in multimodal virtual reality. J Audio Eng Soc 57(3):111–120
Van Atteveldt N, Murray MM, Thut G, Schroeder CE (2014) Multisensory integration: flexible use of general operations. Neuron 81(6):1240–1253. https://doi.org/10.1016/j.neuron.2014.02.044
Wasserstein RL, Schirm AL, Lazar NA (2019) Moving to a world beyond “p < 0.05.” Am Stat 73:1–19. https://doi.org/10.1080/00031305.2019.1583913
Weech S, Kenny S, Barnett-Cowan M (2019) Presence and cybersickness in virtual reality are negatively related: a review. Front Psychol 10:158. https://doi.org/10.3389/fpsyg.2019.00158
Yahata R, Takeya W, Seno T, Tamada Y (2021) Hot wind to the body can facilitate vection only when participants walk through a fire corridor virtually. Perception 50(2):154–164. https://doi.org/10.1177/0301006620987087
Acknowledgements
This research was supported by the Australian Research Council (ARC) (Project ID: DE210101623) and the Institute for Intelligent Systems Research and Innovation. The authors would like to thank Dr. Navid Mohajer for his advice on several sensors and actuators. The authors would like to acknowledge that this research was conducted on the land of the Wadawurrung people of the Kulin Nation, the traditional owners of the lands, and we would like to pay respect to their elders past, present, and emerging.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. Lars Kooijman and Camilo Gonzalez Arango both received support from Deakin University Postgraduate Research Scholarship (DURPS), the Australian Research Council (ARC) (Project ID: DE210101623) as well as from the Institute for Intelligent Systems Research and Innovation (IISRI). Houshyar Asadi received support from the Australian Research Council (ARC) (Project ID: DE210101623) as well as from the Institute for Intelligent Systems Research and Innovation (IISRI).
Author information
Authors and Affiliations
Contributions
Conceptualization: all authors; Methodology: LK, HA and CGA; Software: LK; Validation Verification: HA, CGA; Formal Analysis: LK; Investigation: LK; Resources: HA, SM, and SN; Data Curation: LK and HA; Writing – Original Draft: LK; Writing – Review & Editing: all authors; Visualization: LK; Supervision: HA, SM, and SN; Project Administration: HA and SN; Funding Acquisition: HA and SN.
Corresponding author
Ethics declarations
Conflict of interest
We have no conflict of interest to disclose.
Ethical approval
Deakin University Human Research Ethics Committee approved this study (2021-181).
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kooijman, L., Asadi, H., Gonzalez Arango, C. et al. Investigating the influence of neck muscle vibration on illusory self-motion in virtual reality. Virtual Reality 28, 76 (2024). https://doi.org/10.1007/s10055-024-00951-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10055-024-00951-y