
Abstract
Global environmental change has redistributed earth’s biomass and the inputs and dynamics of basal detrital resources in ecosystems, contributing to the decline of biodiversity. Yet efforts to manage detrital necromass for biodiversity conservation are often overlooked or consider only singular resource types for focal species groups. We argue there is a significant opportunity to broaden our perspective of the spatiotemporal complexity among multiple necromass types for innovative biodiversity conservation. Here, we introduce an ecosystem-scale perspective to disentangling the spatial and temporal characteristics of multiple and distinct forms of necromass and their associated biota. We show that terrestrial and aquatic ecosystems contain a diversity of necromass types, each with contrasting temporal frequencies and magnitudes, and spatial density and configurations. By conceptualising an ecosystem in this way, we demonstrate that specific necromass dynamics can be identified and targeted for management that benefits the unique spatiotemporal requirements of dependent decomposer organisms and their critical role in ecosystem biomass conversion and nutrient recycling. We encourage conservation practitioners to think about necromass quantity, timing of inputs, spatial dynamics, and to engage with researchers to deepen our knowledge of how necromass might be manipulated to exploit the distinct attributes of different necromass types to help meet biodiversity conservation goals.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Highlights
-
Necromass is a diverse resource in terrestrial and aquatic ecosystems.
-
Necromass dynamics can be differentiated by frequency, magnitude, density, and configuration.
-
Necromass quantity, timing, and spatial dynamics should be considered for biodiversity conservation.
Necromass and Global Change
Ecosystem structure and function is shaped by energy and nutrient flow exchanged between both living and dead organic matter (Odum 1962; Moore and others 2004). This dead matter, or ‘necromass’, provides basal resources to detrital food webs and a diversity of specialised biota and drives nutrient recycling processes essential to ecosystems (Swift and others 1979; Benbow and others 2019). Terrestrial resource pools have now moved significantly towards crops, livestock, and humans, and away from forests and wildlife populations (Bar-On and others 2018; Greenspoon and others 2023; Rosenberg and others 2023), therefore redistributing earth’s living biomass and resulting necromass. For example, changes to earth’s forest cover have been significant (Haddad and others 2015), and therefore, inputs of woody debris and leaf litter are similarly impacted. Populations of wild animals, fish, and livestock have been altered (Bar-On and others 2018; Greenspoon and others 2023), and the enormous change to landscapes under cultivation also has altered soil fauna biomass (Rosenberg and others 2023). We suggest that recognising the changes to necromass in ecosystems is the first step towards using these resources as a conservation tool and exploiting their distinct spatial and temporal features to benefit biodiversity.
A Wide Diversity of Necromass Underpins Ecosystems and Supports Biodiversity
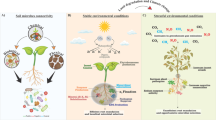
The diversity of necromass spans the full spectrum of life on earth—prokaryotic and eukaryotic, aquatic and terrestrial, autotrophic and heterotrophic (Figure 1) (Benbow and others 2019). This includes plant necromass like standing or fallen dead trees, coarse woody debris and leaf litter in terrestrial and aquatic systems (Maas and others 2021; Neumann and others 2021), bacterial and fungal necromass in soil and streams (Benbow and others 2017; Gulis and Bärlocher 2017; Wang and others 2021a), animal dung (Englmeier and others 2023), and animal carcasses in aquatic (Benbow and others 2020) and terrestrial (Barton and others 2019) ecosystems. In combination, necromass forms a significant resource pool that mirrors living biomass in its diversity, moves above and below ground and across ecosystem boundaries, and contributes significantly to total carbon and nutrient pools and turnover (Schmitz and others 2014; Benbow and others 2019; Neumann and others 2021).

Common necromass types, their generalised temporal and spatial attributes, and drivers of variability. These attributes of necromass are linked closely to the ecology of dependent species and also combine to support broader ecosystem structure and biotic heterogeneity.
There are numerous types of necromass (see Figure 1) that each support a diversity of biotic communities that have evolved to specialise on their distinct nutrient content and digestibility, or various abiotic (for example, pH) and biotic (for example, inter-specific competition) attributes (Ulyshen 2016; Charabidze and others 2021; Butterworth and others 2023a). The range of nutritional profiles of the various forms of plant and animal necromass vary widely along key axes of moisture, nitrogen, and digestibility (Carter and others 2007; Ulyshen 2016; Benbow and others 2019; Charabidze and others 2021) and in combination contribute to the evolution of distinct biota. For example, the insects found at woody debris (Ulyshen 2016) or at animal carcasses (Barton and others 2013b) display little overlap in community composition, even when focusing on a single clade (Coleoptera) within the same ecosystem (Box 1). Necromass resources therefore feed a wealth of biodiversity directly by forming the diet of necrophagous species (for example, detritivores, saprophages, scavengers), and indirectly through food webs and trophic movement of energy to higher consumers (Swift and others 1979; Moore and others 2004; Gessner and others 2010). Necromass also has further physical and structural roles in terrestrial ecosystems, such as mediating soil moisture and microclimates for plant establishment (Donath and Eckstein 2010). Yet it is the spatial and temporal attributes of necromass that is one of its most striking features, as it contributes to biodiversity through local patch dynamics, succession, and landscape heterogeneity (Benbow and others 2019; Butterworth and others 2023a). It is through these direct and indirect mechanisms and higher-order spatial and temporal processes that necromass supports individual species and drives broader community structure and heterogeneity.

The complementary role of different necromass types in supporting whole-of-ecosystem biodiversity. Beetle collections from CWD and leaf litter from forest in the Great Smoky Mountains, USA (A) found only 30% of the 216 species of beetle were shared across resources and 70% were unique to a resource (B). Collections from CWD and vertebrate carcasses in grassy eucalypt woodland near Canberra, Australia (C) found remarkably distinct beetle communities occurring at these resources (D).
Despite knowledge of the importance of spatiotemporal attributes of key necromass types for specific fauna, such as carrion and vultures (Moleon and others 2020) or coarse woody debris and saproxylic beetles (Grove and Forster 2011a, b)), frameworks that incorporate the complementary attributes of multiple necromass types on biotic communities are poorly defined. There is a significant opportunity, therefore, to build on current knowledge of the diversity among necromass types, their wide-ranging spatiotemporal attributes, and the drivers of variation in these attributes to identify research opportunities and assist decision making for conservation managers (see Table 1). Here, we synthesise concepts developed for different necromass types (for example, Kominoski and Rosemond 2012; Benbow and others 2015; Benbow and others 2019; Moleón and others 2019; Butterworth and others 2023a) to show that further insights into biodiversity dynamics are possible when thinking about multiple, distinct forms of necromass and their complementary spatiotemporal properties when scaled up across whole ecosystems.
A Framework to Disentangle Spatiotemporal Dynamics Within the Necromass Resource Pool
The intrinsic properties of different types of necromass give rise to a spectrum of patchy or continuous distributions (for example, dung vs. soil bacteria), and fast and slow decay dynamics (for example, small carcass vs. large woody debris). These properties have shaped the evolution and ecology of a range of consumers that form contrasting and complementary necrobiomes within terrestrial and aquatic ecosystems (Swift and others 1979; Benbow and others 2019; Butterworth and others 2023a) (Box 1). A key gap in necromass ecology, however, is a conceptual framework that effectively disentangles these spatial and temporal attributes of different forms of necromass. Such a framework is needed to allow for comparisons among necromass types, and to provide a practical way to identify features of necromass that can be manipulated to help meet conservation goals. For example, conservation managers can influence the overall quantity of necromass (for example, Manning and others 2013; Seibold and others 2016), or focus on the timing of inputs or particular spatial configurations (for example, Barton and others 2011; Moleon and others 2020). These key dimensions of necromass (quantity, timing, spatial) can affect biodiversity in ways that provide a clear and practical framework to conceptualising the dynamics of necromass in ecosystems (Figure 2).
The diverse spatiotemporal attributes of necromass resources form a spatial mosaic when viewed from a whole-of-ecosystem perspective (Figure 3A). Multiple diverse necromass resources are spread across landscapes in various spatial configurations and stages of decay, which reveals a complex array of inputs and turnover rates of necromass. It is this mosaic and its underlying temporal dynamics that we aim to disentangle, beginning with a single necromass particle, then moving to multiple particles and ultimately the whole ecosystem. The key attributes of a single necromass particle include its position in geographic space and its decay time (Figure 3A, B). Spatially, a single necromass particle will be of a particular size (for example, a small or large log) and proximity to other resources that can determine its associated biota (Harmon and others 1986; Brunet and Isacsson 2009; Moleón and others 2015). Temporally, a necromass particle decomposes over time in a predictable sequence of decay that is accompanied by the gradual release of energy into the surrounding soil and atmosphere, and via trophic pathways to decomposer organisms. The magnitude of energy release and longevity of decay is determined by the mass and nutrient content of the particle in combination with the complex external abiotic and biotic factors controlling decomposition rate (Swift and others 1979; Dawson and others 2023). Notably, larger necromass particles, like fallen timber or animal carcasses, host biotic communities that also undergo successional change through the decomposition process (Grove and Forster 2011b; Dawson and others 2022). A distinguishing feature of multiple necromass types is their contrasting set of spatial configurations and decay rates (Figure 3A, C).

Conceptual framework to disentangle whole-of-ecosystem necromass dynamics. A A whole ecosystem contains a mosaic of necromass resources in various spatial configurations and states of decay. B Each necromass particle decomposes over time in a predictable sequence of early, mid and late decay stages accompanied by the release of energy into the surrounding ecosystem (soil, atmosphere, organisms). C Different necromass resources like fallen trees, carcasses and plant litter display highly contrasting decay dynamics in terms of their magnitude and longevity of decay. D A whole-of-ecosystem spatial mosaic includes multiple necromass types with contrasting magnitudes, decay times and frequencies of input.
Spatially, distinct necromass resources vary in their particle size and configurations, ranging from small but widespread leaf litter to large but rare vertebrate carcasses that each vary in density and distribution (Figure 3A). Temporally, different necromass types will exhibit highly contrasting decay profiles characterised by a different magnitude and longevity due to their different sizes and unique composition (for example, autotroph vs. heterotroph necromass) (Benbow and others 2019) (Figure 3C). A broader view of multiple necromass types and their underlying spatial and temporal attributes reveals a mosaic of resources across a landscape (Figure 3A). It is at this scale where the full complexity of necromass dynamics has a clear role in driving species and community dynamics. Key features at this scale are the constantly shifting distribution of decomposition hotspots that each vary in magnitude and longevity, and different necromass types that enter ecosystems at a variety of frequencies (Figure 3D). This means, for example, that multiple animal carcasses will enter and cycle through an ecosystem during the more prolonged decay of a single fallen tree, thus generating multiple hotspots and hosting distinct communities corresponding to different stages of decay. Temporally, different types of necromass turnover within ecosystems at varying frequencies, and continuity in resources for dependent species therefore occurs over different timeframes when comparing across resource types.
Linking Necromass Dynamics to Conservation
Our framework highlights the diversity of dynamics among different necromass types at the whole-of-ecosystem scale, and we suggest this can guide conservation management. Particular necromass types, such as dead wood or animal carcasses, are commonly considered for conservation management due to specialised species associated with them (Sandstrom and others 2019). Yet the necromass resource pool also generates broader heterogeneity within an ecosystem through its diverse suite of contrasting magnitudes, turnover rates, densities, and distributions (Figure 2), which, in turn, supports contrasting biotic communities that have evolved to exploit the various spatiotemporal characteristics of necromass. Landscape homogenisation processes can therefore reduce necromass diversity and its ability to support diverse biota. To link this complexity to potential conservation actions, we outline five broad questions that emerge from our conceptual framework. These questions are linked to the biota, quantity, temporal attributes, spatial attributes, and methodology and are intended to guide researchers in how to make their research and findings applicable to conservation practitioners.
Which Biota are Associated with Necromass?
A primary question for conservation practitioners and researchers is ‘which biota are dependent on necromass?’. Knowledge of the biodiversity of decomposer taxa is an important first step to identify vulnerable taxa. Differences in the magnitude and frequencies by which necromass categories are inputted to ecosystems may have some utility in predicting the vulnerability of associated decomposer taxa. For example, high-amplitude and low-frequency inputs, such as large vertebrate carcasses or dead trees, can support highly specialised taxa (Bourguignon and others 2011; Zou and others 2021). It might be predicted that taxa specialised on such large and infrequent necromass may become more vulnerable as anthropogenic landscapes continue to lose large mammals (Dirzo and others 2014), large fish (Audzijonyte and others 2013), and large trees (Gilhen-Baker and others 2022), especially at large scales where dispersal ability of specialists may become constrained (Gossner and Muller 2011). Some of the most imperilled decomposers depend on larger resources. For example, the loss of adequately large carrion (for example, from extinct passenger pigeons and threatened prairie dogs) is one of the leading hypotheses about why the American burying beetle Nicrophorus americanus disappeared across most of its range in the twentieth century (Sikes and Raithel 2002). It has similarly been proposed that the loss of megaherbivores in Australia resulted in the decline of an obligate scavenger (Cryptogyps lacertosus), subsequently giving rise to raptors who scavenge facultatively (Mather and others 2022). Other examples include condors, which must fly long distances to locate suitably large carcasses (Speziale and others 2008), and a large number of insects dependent on coarse woody debris (Ulyshen and Šobotník 2018). However, it is also possible that larger organisms that depend on larger substrates have simply received more research attention than smaller organisms. For example, some taxa associated with abundant resources (that is, high frequency, low magnitude) such as plant litter or mushrooms, can also display high degrees of specialisation, such as found among mycetophagous drosophilids (Lacy 1984). Predictions about vulnerability stemming from resource dynamics could arguably go in both directions. Knowledge of the fauna associated with different necromass resources is therefore a prerequisite to identifying vulnerable taxa.
The availability and continuity of necromass resources also play a role in supporting biodiversity in ways additional to trophic provisioning (Gibb and others 2006a; Barton and others 2013a; Moleón and others 2019). For example, necromass patchiness and ephemerality have been noted for contributing to biotic heterogeneity and community dynamics within landscapes (Barton and others 2013a; Benbow and others 2019), although landscape-scale empirical studies of this phenomenon are rare (Gossner and Muller 2011; Barton and others 2016; Mateo-Tomas and others 2019; Moleon and others 2020). Species dependent on highly ephemeral necromass that is also high amplitude and low frequency could be expected to be more vulnerable than those dependent on longer-lasting resources capable of supporting multiple generations. But this may not be the case if ephemerality selects for strong dispersal abilities. Among saproxylic insects, for example, taxa associated with the highly ephemeral phloem layer of dead trees are thought to be better dispersers and therefore less sensitive to the amount and continuity of coarse woody debris than those that utilise rotting wood (Ulyshen and Šobotník 2018). Blowflies, too, are excellent flyers and appear capable of colonising ephemeral carrion across contrasting habitat types (Barton and Evans 2017; Butterworth and others 2023b), highlighting the importance of spatial configuration of necromass resources. Patchiness or distance between resources is therefore also a critical consideration for conservation, particularly in the context of the dispersal capacities of different species (Table 1).
Distinct sets of species are found on different necromass types, as is clear in Box 1, and this means diverse necromass resources will support diverse and complementary communities. Multiple necromass types can therefore have further positive effects on biodiversity by increasing beta diversity and driving biotic heterogeneity. Many studies have demonstrated the utility of a single necromass type for studies of metacommunity theory (for example, Finn 2001; Benbow and others 2015; Abrego 2022), but we take this further by showing how we can disentangle multiple necromass dynamics and start measuring critical aspects that contribute to biotic heterogeneity at ecosystem scales. Knowing which taxa are associated with a necromass resource and how changing that resource alters those specific consumer communities is one critical step (Sandstrom and others 2019; Abrego 2022). There is also the potential of manipulating biotic complementarity among multiple resource types that is completely unexplored as a conservation tool (Box 1).
What is the Appropriate Quantity and Profile of Necromass?
Management of the overall amount of necromass resources is an important consideration in ecosystems that have experienced major disruptions to supply via extractive processes, excessive inputs, or changes to disturbance regimes. Such changes in amount are often also focused on particular particle sizes of necromass, which alters the profile or size spectrum of the necromass resource pool. Such disruptions might include firewood collection from downed trees (Manning and others 2020) or salvage logging (Thorn and others 2018), hunting or culling of large animals (Mateo-Tomás and others 2015), mass mortality events (Fey and others 2015), weed invasion (Lindsay and French, 2005), wildfire (Newsome and Spencer 2021), or livestock enterprises (Piñeiro and others 2006). This can be critical for ecosystem function if nutrient pools are significantly shifted (Subalusky and others 2017), resources for specialist decomposers are absent (Hjalten and others 2012), or the influx of resources promotes invasive pests (Spencer and others 2020). Knowledge of the magnitude of such changes is an important first step to identifying management requirements. Tools available to quantify coarse woody debris include the calculation of physical dimensions to generate volumes (Schiegg 2000b; Threlfall and others 2019; Manning and others 2020), and carrion biomass can be derived from population abundance and species body size data (Subalusky and others 2017; Barton and others 2019; Mateo-Tomás and others 2019). Mapping of live and dead biomass and use of steady-state models to derive turnover rates can be applied to either terrestrial plant litter (Gora and others 2019; Neumann and others 2021), and heterotrophic biomass in streams (Waters 1977; Benbow and others 2017; Benke and Huryn 2017; Gulis and Bärlocher 2017) (Table 1). Using these tools, researchers and managers can identify changes in quantity and develop practical actions that can address gaps in quantities or volumes to benefit biodiversity. For example, addition of coarse woody debris has been shown to benefit saproxylic insects and fungi (Ulyshen and Šobotník 2018; Sandstrom and others 2019) but observed effects appear to be stronger in temperate zones than in the subtropics (Ulyshen and Hanula 2009) and, as discussed in the next section, the amount of coarse woody debris may be less important than its temporal continuity.
Provisioning of animal carcasses can support a range of scavenger species (Moreno-Opo and others 2015; Stiegler and others 2020), and in some ecosystems with a necromass shortfall, deliberate additions can bring forward the benefits for biodiversity to bridge any production gap that might otherwise take many years (Manning and others 2013). In grasslands, the amount of grass litter influences community composition by mediating the space available for germination of forbs and grasses, and this in turn relates to management decisions about appropriate management regimes—particularly where the dynamics of necromass have been influenced by fire suppression or livestock enterprises (Piñeiro and others 2006). Beyond the effects of total necromass quantity in an ecosystem, decisions about the size profile of individual necromass particles can also have important implications for biodiversity. For instance, more species of saproxylic beetles benefit from inputs of large than small logs (Grove and Forster 2011a, b). However, many species are also specific to small-diameter wood (Jonsell and others 2007), much like various carcass sizes support different subsets of scavengers (Moleón and others 2015). Even among necromass derived from the same species, differences in size can strongly influence the composition of species associated with it (Brin and others 2011). When managing necromass quantity, therefore, a spectrum or diversity of particles should be considered.
When is the Appropriate Timing of Necromass Inputs?
There are key differences in the drivers of decomposition and succession of associated biota at contrasting necromass types (von Hoermann and others 2023). This means there is a need to maintain the input of multiple resource types at multiple decay stages (see Figure 2b) to create the ‘shifting mosaic’ in an ecosystem (Table 1). One of the biggest challenges for conservation management, therefore, is to ensure adequate continuity or availability of necromass through time. Major disruptions in continuity can result in considerable biodiversity losses. For example, differences in continuity explain why old forests can support a higher diversity of saproxylic insects than younger forests despite having no more or even less dead wood (Brin and others 2016; Traylor and others 2023). While a major pulse of resources can quickly boost the numbers of associated species (Koenig and Liebhold 2017), only with repeated inputs can such benefits be sustained long-term. Substrates with higher turnover, like carrion or dung, would need to enter an ecosystem more regularly than those with comparatively slow decay rates like dead wood. However, even regular inputs of dead trees of multiple species are needed given patterns of host-specificity and successional changes as decomposition proceeds (Ulyshen and Hanula 2010; Seibold and others 2023). A secondary consideration concerns the appropriate timing of inputs to maintain continuity. Consideration of the phenology or seasonality, frequency, and longevity of different resources can be relevant. It is variation among these attributes that generate mosaics and therefore create colonisation opportunities and continuity in breeding for dependent species and communities. Seasonality is particularly important and includes events such as increased litter fall (Nebel and others 2001), emergence of aquatic insects (Pray and others 2009), and breeding or migration events that lead to inputs of placenta or young (Quaggiotto and others 2018). In some cases species are able to track these seasonal movements in resources, including vultures following livestock (Olea and Mateo-Tomas 2009) or ravens following hunters (White 2005). In such examples, it is predictability rather than continuity that may be important (Lopez-Lopez and others 2014). In other situations, completely different sets of species use necromass resources at different times of year, as has been found for carrion and scavengers in the Australian alpine bioregion (Vandersteen and others 2023) (Figure 4 and Box 2). Climate change and shifts in rainfall, temperature, and the onset of seasons (Wang and others 2021b) will be a key challenge with the management of necromass resources. Key events linked to climate extremes such as increased storms or wildfire can result in significant inputs of necromass via windthrown trees (Peters and others 2013; Chao and others 2022) or animal mass mortalities (Newsome and Spencer 2021; Barton and others 2022). Increased interannual rainfall variability (Pendergrass and others 2017) can also have substantial impacts on necromass dynamics in grasslands. Seasons of particularly high rainfall drive prolific production of grass litter that have been shown to persist for several years and modify fauna community composition and habitat suitability (Antos and Schultz 2020).

Shifting use of carrion by vertebrate scavengers across seasons. A Total number of vertebrate scavenging and investigation, based on replicated monitoring of 15 carcasses per season in the Australian Alps. Higher scavenging in spring was linked to ravens (Corvus sp.) (B), and in winter to brushtail possums (Trichosurus vulpecula). Modified after (Vandersteen and others 2023).
Where is the Appropriate Location of Necromass?
Complexity in the spatial attributes of necromass resources will affect biotic heterogeneity within ecosystems, just as habitat structural complexity drives animal diversity (Tews and others 2004; Loke and Chisholm 2022). Approaches to measuring spatial complexity can include the density of necromass types and their spatial configuration, but also variation in their density and configuration (Loke and Chisholm 2022; Butterworth and others 2023a). The simplification of ecosystems and landscapes, and resulting biotic homogenisation, is a global phenomenon, with documented negative outcomes for biodiversity (Gossner and others 2016; Wang and others 2021c). Maintaining or increasing spatial complexity among multiple necromass types is therefore part of addressing this problem. For example, returning coarse woody debris to improve ground-layer habitat structural complexity in grassy eucalypt woodlands in south-eastern Australia has had positive effects on insect and reptile biota (Barton and others 2011; Manning and others 2013), but the geographic distance among woody debris also is critical to compositional turnover (Barton and others 2009) and therefore broader ecosystem heterogeneity. The spatial complexity and connectivity of coarse woody debris is important for the richness of saproxylic invertebrates (Schiegg 2000a; Gibb and others 2006b). Because local conditions strongly impact the suitability of dead wood to many saproxylic taxa, a diversity of inputs that cover a range of conditions will have the greatest benefits to biodiversity (Table 1). This is because spatial complexity of necromass can affect organism perception of resource quality. For example, whether woody debris is sun-exposed or shaded is known to affect the composition of saproxylic beetle communities (Vodka and others 2009; Müller and others 2020; Vogel and others 2021). For avian scavengers of carrion, the spatial predictability of necromass can be important for more social or dominant species of vulture (Cortés-Avizanda and others 2016) or gulls feeding on coastal resources (Quaggiotto and others 2018).
What Method of Necromass Management can be Implemented?
The method by which necromass is added to or removed from a system can affect the quality or suitability of the resource for various species. Regarding inputs of woody necromass, trees killed through natural processes such as fire or beetle attack support a greater diversity of colonising species than those killed through artificial means such as girdling (Shea and others 2002). Moreover, standing and fallen trees support distinct invertebrate communities and exogenous factors such as fire and sun exposure can further influence which species colonise woody debris (Vindstad and others 2020). Decisions on how vertebrate necromass is added can have similarly important implications. Research shows that vertebrate carcasses with traumatic injuries (for example, broken skin) decay more rapidly and support different scavenger communities compared with intact carcasses (Mann and others 1990; Cairncross and others 2022). The seasonality or distribution of hunting remains has also been shown to affect the diversity and structure of scavenger communities (Selva and Fortuna 2007). A guiding principle that emerges from such examples is that the dynamics of necromass will be most advantageous to biodiversity when it closely mimics natural processes (Table 1). In human-modified landscapes where ‘natural’ processes are altered, management might be in ways that fit ecological and evolutionary requirements of species. For instance, scavenger conservation was incorporated into European sanitary regulations for managing animal by-products such as livestock carcasses (Regulation (EC) No 1069/2009) by considering ‘the natural consumption patterns’, that is, the unpredictable way in which carrion appears in natural ecosystems (Mateo-Tomás and others 2019). An understanding of the diversity of resources, their dependent taxa, and naturally occurring quantities, temporalities, and distributions would be greatly helpful for managers to deal with necromass for conservation.
Implications and Conclusions
Ecosystems globally have experienced changes in necromass resource dynamics via harvesting and extractive practices that have reduced inputs, or via invasive species, unmanaged populations and mass mortality events that have increased inputs of some necromass types (for example, Sandstrom and others 2019; Bartel and others 2023). Such disturbances can affect biodiversity dependent on necromass as well as broader ecosystem structure and function, but the complex dynamics of necromass makes it difficult to disentangle the key spatial and temporal dimensions of these resources that are able to be managed. To address this challenge, we have introduced a conceptual framework that unpacks the distinct temporal and spatial attributes of contrasting necromass types within ecosystems. Multiple necromass types complement each other to support distinct species and biotic communities, thus benefitting biodiversity at the whole-of-ecosystem scale. A guiding principle that emerges from our framework is that the dynamics of necromass will be most advantageous to biodiversity when it closely mimics natural processes. In modified ecosystems, knowledge of natural processes and their variability might be limited, and experimental or adaptive management interventions could be appropriate to build cause-and-effect relationships between necromass dynamics and biodiversity outcomes (Table 1). ‘Backcasting’ could also be necessary to establish what quantities, for example, were present in an ecosystem prior to modification (Manning and others 2006). We encourage an experimental approach to necromass management more broadly as a way to improve knowledge of necromass-dependent biota, and to test how the quantity, timing, and spatial arrangements of multiple necromass types can produce positive biodiversity change.
References
Abrego N. 2022. Wood-inhabiting fungal communities: Opportunities for integration of empirical and theoretical community ecology. Fungal Ecology 59:101112.
Antos M, Schultz NL. 2020. Climate-mediated changes to grassland structure determine habitat suitability for the critically endangered plains-wanderer (Pedionomus torquatus). Emu - Austral Ornithology 120:2–10.
Audzijonyte A, Kuparinen A, Gorton R, Fulton EA. 2013. Ecological consequences of body size decline in harvested fish species: positive feedback loops in trophic interactions amplify human impact. Biology Letters 9:20121103.
Bar-On YM, Phillips R, Milo R. 2018. The biomass distribution on Earth. Proceedings of the National Academy of Sciences (USA) 115:6506–6511.
Bartel SL, Stephenson T, Crowder DW, Jones ME, Storfer A, Strickland MS, Lynch L. 2023. Global change influences scavenging and carrion decomposition. Trends in Ecology & Evolution.
Barton PS, Evans MJ. 2017. Insect biodiversity meets ecosystem function: differential effects of habitat and insects on carrion decomposition. Ecological Entomology 42:364–374.
Barton PS, Manning A, Gibb H, Lindenmayer DB, Cunningham SA. 2009. Conserving ground-dwelling beetles in an endangered woodland community: Multi-scale habitat effects on assemblage diversity. Biological Conservation 142:1701–1709.
Barton PS, Manning AD, Gibb H, Wood JT, Lindenmayer DB, Cunningham SA. 2011. Experimental reduction of native vertebrate grazing and addition of logs benefit beetle diversity at multiple scales. Journal of Applied Ecology 48:943–951.
Barton PS, Cunningham SA, Lindenmayer DB, Manning AD. 2013a. The role of carrion in maintaining biodiversity and ecological processes in terrestrial ecosystems. Oecologia 171:761–772.
Barton PS, Cunningham SA, Macdonald BCT, Mcintyre S, Lindenmayer DB, Manning AD. 2013b. Species traits predict assemblage dynamics at ephemeral resource patches created by carrion. PLoS ONE 8:e53961.
Barton PS, Evans MJ, Foster CN, Pechal JL, Bump JK, Quaggiotto MM, Benbow ME. 2019. Towards quantifying carrion biomass in ecosystems. Trends in Ecology & Evolution 34:950–961.
Barton PS, McIntyre S, Evans MJ, Bump JK, Cunningham SA, Manning AD. 2016. Substantial long-term effects of carcass addition on soil and plants in a grassy eucalypt woodland. Ecosphere 7.
Barton PS, Reboldi A, Bonat S, Mateo-Tomás P, Newsome TM. 2022. Climate-driven animal mass mortality events: is there a role for scavengers? Environmental Conservation in press.
Benbow ME, Barton PS, Ulyshen MD, Beasley JC, DeVault TL, Strickland MS, Tomberlin JK, Jordan HR, Pechal JL. 2019. Necrobiome framework for bridging decomposition ecology of autotrophically and heterotrophically derived organic matter. Ecological Monographs 89:e01331.
Benbow ME, Pechal JL, Mohr RM. 2015. Community and landscape ecology of carrion. Benbow ME, Tomberlin JK, Tarone AM editors. Carrion Ecology, Evolution, and Their Applications: CRC Press, p151–185.
Benbow ME, Pechal JL, Ward AK. 2017. Chapter 9—Heterotrophic Bacteria Production and Microbial Community Assessment. Hauer FR, Lamberti GA editors. Methods in Stream Ecology, Volume 1 (Third Edition). Boston: Academic Press, p161–176.
Benbow ME, Receveur JP, Lamberti GA. 2020. Death and decomposition in aquatic ecosystems. Frontiers in Ecology and Evolution 8.
Benke AC, Huryn AD. 2017. Chapter 35—Secondary Production and Quantitative Food Webs. Lamberti GA, Hauer FR editors. Methods in Stream Ecology (Third Edition): Academic Press, p235–254.
Bourguignon T, Šobotník J, Lepoint G, Martin J-M, Hardy OJ, Dejean A, Roisin Y. 2011. Feeding ecology and phylogenetic structure of a complex neotropical termite assemblage, revealed by nitrogen stable isotope ratios. Ecological Entomology 36:261–269.
Brin A, Bouget C, Brustel H, Jactel H. 2011. Diameter of downed woody debris does matter for saproxylic beetle assemblages in temperate oak and pine forests. Journal of Insect Conservation 15:653–669.
Brin A, Valladares L, Ladet S, Bouget C. 2016. Effects of forest continuity on flying saproxylic beetle assemblages in small woodlots embedded in agricultural landscapes. Biodiversity and Conservation 25:587–602.
Brunet J, Isacsson G. 2009. Restoration of beech forest for saproxylic beetles-effects of habitat fragmentation and substrate density on species diversity and distribution. Biodiversity and Conservation 18:2387–2404.
Butterworth NJ, Benbow ME, Barton PS. 2023a. The ephemeral resource patch concept. Biological Reviews 98:697–726.
Butterworth NJ, Wallman JF, Johnston NP, Dawson BM, Sharp-Heward J, McGaughran A. 2023b. The blowfly Chrysomya latifrons inhabits fragmented rainforests, but shows no population structure. Oecologia 201:703–719.
Cairncross RJ, Barton PS, Bonat S, Crowther MS, Dickman CR, Vandersteen J, Newsome TM. 2022. The predatory impacts of invasive European wasps on flies are facilitated by carcasses with open wounds. Food Webs 31.
Carter D, Yellowlees D, Tibbett M. 2007. Cadaver decomposition in terrestrial ecosystems. Naturwissenschaften 94:12–24.
Chao KJ, Liao PS, Chen YS, Song GZM, Phillips OL, Lin HJ. 2022. Very low stocks and inputs of necromass in wind-affected tropical forests. Ecosystems 25:488–503.
Charabidze D, Trumbo S, Grzywacz A, Costa JT, Benbow ME, Barton PS, Matuszewski S. 2021. Convergence of social strategies in carrion breeding insects. Bioscience 71:1028–1037.
Cortés-Avizanda A, Blanco G, DeVault TL, Markandya A, Virani MZ, Brandt J, Donázar JA. 2016. Supplementary feeding and endangered avian scavengers: benefits, caveats, and controversies. Frontiers in Ecology and the Environment 14:191–199.
Dawson BM, Wallman JF, Evans MJ, Barton PS. 2022. Insect abundance patterns on vertebrate remains reveal carrion resource quality variation. Oecologia 198:1043–1056.
Dawson BM, Ueland M, Carter DO, McLntyre D, Barton PS. 2023. Bridging the gap between decomposition theory and forensic research on postmortem interval. International Journal of Legal Medicine 98:697–726.
Dirzo R, Young HS, Galetti M, Ceballos G, Isaac NJB, Collen B. 2014. Defaunation in the anthropocene. Science 345:401–406.
Donath TW, Eckstein RL. 2010. Effects of bryophytes and grass litter on seedling emergence vary by vertical seed position and seed size. Plant Ecology 207:257–268.
Englmeier J, Mitesser O, Benbow ME, Hothorn T, von Hoermann C, Benjamin C, Fricke U, Ganuza C, Haensel M, Redlich S, Riebl R, Botero SR, Rummler T, Steffan-Dewenter I, Stengel E, Tobisch C, Uhler J, Uphus L, Zhang J, Muller J. 2023. Diverse effects of climate, land use, and insects on dung and carrion decomposition. Ecosystems 26:397–411.
Ferro ML, Gimmel ML, Harms KE, Carlton CE. 2012. Comparison of the Coleoptera communities in leaf litter and rotten wood in Great Smoky Mountains National Park, USA. Insecta Mundi 259:1–58.
Fey SB, Siepielski AM, Nussle S, Cervantes-Yoshida K, Hwan JL, Huber ER, Fey MJ, Catenazzi A, Carlson SM. 2015. Recent shifts in the occurrence, cause, and magnitude of animal mass mortality events. Proceedings of the National Academy of Sciences of the United States of America 112:1083–1088.
Finn JA. 2001. Ephemeral resource patches as model systems for diversity-function experiments. Oikos 92:363–366.
Gessner MO, Swan CM, Dang CK, McKie BG, Bardgett RD, Wall DH, Hattenschwiler S. 2010. Diversity meets decomposition. Trends in Ecology & Evolution 25:372–380.
Gibb H, Hjälten J, Ball JP, Atlegrim O, Pettersson RB, Hilszczanski J, Johansson T, Danell K. 2006a. Effects of landscape composition and substrate availability on saproxylic beetles in boreal forests: a study using experimental logs for monitoring assemblages. Ecography 29:191–204.
Gibb H, Pettersson RB, Hjalten J, Hilszczanski J, Ball JP, Johansson T, Atlegrim O, Danell K. 2006b. Conservation-oriented forestry and early successional saproxylic beetles: responses of functional groups to manipulated dead wood substrates. Biological Conservation 129:437–450.
Gilhen-Baker M, Roviello V, Beresford-Kroeger D, Roviello GN. 2022. Old growth forests and large old trees as critical organisms connecting ecosystems and human health. A review. Environmental Chemistry Letters 20:1529–1538.
Gora EM, Kneale RC, Larjavaara M, Muller-Landau HC. 2019. Dead wood necromass in a moist tropical forest: stocks, fluxes, and spatiotemporal variability. Ecosystems 22:1189–1205.
Gossner MM, Lewinsohn TM, Kahl T, Grassein F, Boch S, Prati D, Birkhofer K, Renner SC, Sikorski J, Wubet T, Arndt H, Baumgartner V, Blaser S, Bluthgen N, Borschig C, Buscot F, Diekotter T, Jorge LR, Jung K, Keyel AC, Klein AM, Klemmer S, Krauss J, Lange M, Muller J, Overmann J, Pasalic E, Penone C, Perovic DJ, Purschke O, Schall P, Socher SA, Sonnemann I, Tschapka M, Tscharntke T, Turke M, Venter PC, Weiner CN, Werner M, Wolters V, Wurst S, Westphal C, Fischer M, Weisser WW, Allan E. 2016. Land-use intensification causes multitrophic homogenization of grassland communities. Nature 540: 266.
Gossner MM, Muller J. 2011. The influence of species traits and q-metrics on scale-specific beta-diversity components of arthropod communities of temperate forests. Landscape Ecology 26:411–424.
Greenspoon L, Krieger E, Sender R, Rosenberg Y, Bar-On YM, Moran U, Antman T, Meiri S, Roll U, Noor E, Milo R. 2023. The global biomass of wild mammals. Proceedings of the National Academy of Sciences 120:e2204892120.
Grove SJ, Forster L. 2011a. A decade of change in the saproxylic beetle fauna of eucalypt logs in the Warra long-term log-decay experiment, Tasmania. 1. Description of the fauna and seasonality patterns. Biodiversity and Conservation 20:2149–2165.
Grove SJ, Forster L. 2011b. A decade of change in the saproxylic beetle fauna of eucalypt logs in the Warra long-term log-decay experiment, Tasmania. 2. Log-size effects, succession, and the functional significance of rare species. Biodiversity and Conservation 20:2167–2188.
Gulis V, Bärlocher F. 2017. Chapter 10 - Fungi: Biomass, Production, and Community Structure. Hauer FR, Lamberti GA editors. Methods in Stream Ecology, Volume 1 (Third Edition). Boston: Academic Press, p177–192.
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, Lovejoy TE, Sexton JO, Austin MP, Collins CD, Cook WM, Damschen EI, Ewers RM, Foster BL, Jenkins CN, King AJ, Laurance WF, Levey DJ, Margules CR, Melbourne BA, Nicholls AO, Orrock JL, Song D-X, Townshend JR. 2015. Habitat fragmentation and its lasting impact on Earth’s ecosystem. Science Advances 1:e1500052.
Harmon ME, Franklin JF, Swanson FJ, Sollins P, Gregory SV, Lattin JD, Anderson NH, Cline SP, Aumen NG, Sedell JR, Lienkaemper GW, Cromack JK, Cummins KW. 1986. Ecology of coarse woody debris in temperate ecosystems. Advances in Ecological Research 15:133–302.
Hjalten J, Stenbacka F, Pettersson RB, Gibb H, Johansson T, Danell K, Ball JP, Hilszczanski J. 2012. Micro and macro-habitat associations in saproxylic beetles: implications for biodiversity management. PLoS ONE 7.
Jonsell M, Hansson J, Wedmo L. 2007. Diversity of saproxylic beetle species in logging residues in Sweden—comparisons between tree species and diameters. Biological Conservation 138:89–99.
Koenig WD, Liebhold AM. 2017. A decade of emerald ash borer effects on regional woodpecker and nuthatch populations. Biological Invasions 19:2029–2037.
Kominoski JS, Rosemond AD. 2012. Conservation from the bottom up: forecasting effects of global change on dynamics of organic matter and management needs for river networks. Freshwater Science 31:51–68.
Lacy RC. 1984. Predictability, toxicity, and trophic niche breadth in fungus-feeding Drosophilidae (Diptera). Ecological Entomology 9:43–54.
Lindsay EA, French K. 2005. Litterfall and nitrogen cycling following invasion by Chrysanthemoides monilifera ssp rotundata in coastal Australia. Journal of Applied Ecology 42:556–566.
Loke LHL, Chisholm RA. 2022. Measuring habitat complexity and spatial heterogeneity in ecology. Ecology Letters 25:2269–2288.
Lopez-Lopez P, Garcia-Ripolles C, Urios V. 2014. Food predictability determines space use of endangered vultures: implications for management of supplementary feeding. Ecological Applications 24:938–949.
Maas GCB, Sanquetta CR, Marques R, Machado SD, Sanquetta MNI, Dalla Corte AP, Schmidt LN. 2021. Combining sample designs to account for the whole necromass carbon stock in brazilian atlantic forest. Journal of Sustainable Forestry 40:639–655.
Mann RW, Bass WM, Meadows L. 1990. Time since death and decomposition of the human body: variables and observations in case and experimental field studies. Journal of Forensic Sciences 35:103–111.
Manning AD, Lindenmayer DB, Fischer J. 2006. Stretch goals and backcasting: approaches for overcoming barriers to large-scale ecological restoration. Restoration Ecology 14:487–492.
Manning AD, Fischer J, Felton A, Newell B, Steffen W, Lindenmayer DB. 2009. Landscape fluidity—a unifying perspective for understanding and adapting to global change. Journal of Biogeography 36:193–199.
Manning AD, Wood J, Cunningham RB, McIntyre S, Shorthouse DJ, Gordon IJ, Lindenmayer DB. 2011. Integrating research and restoration: a long-term experiment in south-eastern Australia. Australian Zoologist 35:663–648.
Manning AD, Cunningham RB, Lindenmayer DB. 2013. Bringing forward the benefits of coarse woody debris in ecosystem recovery under different levels of grazing and vegetation density. Biological Conservation 157:204–214.
Manning AD, Cunningham RB, Tongway D, Lindenmayer DB. 2020. Woodlands and woody debris: Understanding structure and composition to inform restoration. PLoS ONE 15.
Mateo-Tomás P, Olea PP, Moleón M, Vicente J, Botella F, Selva N, Viñuela J, Sánchez-Zapata JA. 2015. From regional to global patterns in vertebrate scavenger communities subsidized by big game hunting. Diversity and Distributions 21:913–924.
Mateo-Tomás P, Olea PP, López-Bao JV, González-Quirós P, Peón P. 2019. Different criteria for implementing sanitary regulations lead to disparate outcomes for scavenger conservation. Journal of Applied Ecology 56:500–508.
Mateo-Tomas P, Olea PP, Selva N, Sanchez-Zapata JA. 2019. Species and individual replacements contribute more than nestedness to shape vertebrate scavenger metacommunities. Ecography 42:365–375.
Mather EK, Lee MSY, Worthy TH. 2022. A new look at an old Australian raptor places Taphaetus lacertosus de Vis 1905 in the Old World vultures (Accipitridae: Aegypiinae). Zootaxa 5168:1–23.
Moleón M, Sanchez-Zapata JA, Sebastian-Gonzalez E, Owen-Smith N. 2015. Carcass size shapes the structure and functioning of an African scavenging assemblage. Oikos 124:1391–1403.
Moleon M, Selva N, Sanchez-Zapata JPA. 2020. The components and spatiotemporal dimension of carrion biomass quantification. Trends in Ecology & Evolution 35:91–92.
Moleón M, Selva N, Quaggiotto MM, Bailey DM, Cortés-Avizanda A, DeVault TL. 2019. Carrion availability in space and time. Carrion ecology and management: Springer, p23–44.
Moore JC, Berlow EL, Coleman DC, de Ruiter PC, Dong Q, Hastings A, Johnson NC, McCann KS, Melville K, Morin PJ, Nadelhoffer K, Rosemond AD, Post DM, Sabo JL, Scow KM, Vanni MJ, Wall DH. 2004. Detritus, trophic dynamics and biodiversity. Ecology Letters 7:584–600.
Moreno-Opo R, Trujillano A, Arredondo A, Gonzalez LM, Margalida A. 2015. Manipulating size, amount and appearance of food inputs to optimize supplementary feeding programs for European vultures. Biological Conservation 181:27–35.
Müller J, Ulyshen M, Seibold S, Cadotte M, Chao A, Bässler C, Vogel S, Hagge J, Weiß I, Baldrian P, Tláskal V, Thorn S. 2020. Primary determinants of communities in deadwood vary among taxa but are regionally consistent. Oikos 129:1579–1588.
Nebel G, Dragsted J, Vega AS. 2001. Litter fall, biomass and net primary production in flood plain forests in the Peruvian Amazon. Forest Ecology and Management 150:93–102.
Neumann M, Turner J, Lewis T, McCaw L, Cook G, Adams MA. 2021. Dynamics of necromass in woody Australian ecosystems. Ecosphere 12:e03693.
Newsome TM, Spencer EE. 2021. Megafires attract avian scavenging but carcasses still persist. Diversity and Distributions 28:515–528.
Odum EP. 1962. Relationships between structure and function in the ecosystem. Japanese Journal of Ecology 12:108–118.
Olea PP, Mateo-Tomas P. 2009. The role of traditional farming practices in ecosystem conservation: the case of transhumance and vultures. Biological Conservation 142:1844–1853.
Pendergrass AG, Knutti R, Lehner F, Deser C, Sanderson BM. 2017. Precipitation variability increases in a warmer climate. Scientific Reports 7:17966.
Pereira LM, Owen-Smith N, Moleón M. 2014. Facultative predation and scavenging by mammalian carnivores: Seasonal, regional and intra-guild comparisons. Mammal Review 44:44–55.
Peters EB, Wythers KR, Bradford JB, Reich PB. 2013. Influence of disturbance on temperate forest productivity. Ecosystems 16:95–110.
Piñeiro G, Paruelo J, Oesterheld M. 2006. Potential long-term impacts of livestock introduction on carbon and nitrogen cycling in grasslands of Southern South America. Global Change Biology 12:1267–1284.
Pray CL, Nowlin WH, Vanni MJ. 2009. Deposition and decomposition of periodical cicadas (Homoptera: Cicadidae: Magicicada) in woodland aquatic ecosystems. Journal of the North American Benthological Society 28:181–195.
Quaggiotto MM, Barton PS, Morris CD, Moss SEW, Pomeroy PP, McCafferty DJ, Bailey DM. 2018. Seal carrion is a predictable resource for coastal ecosystems. Acta Oecologica-International Journal of Ecology 88:41–51.
Rosenberg Y, Bar-On YM, Fromm A, Ostikar M, Shoshany A, Giz O, Milo R. 2023. The global biomass and number of terrestrial arthropods. Science Advances 9: eabq4049.
Sandstrom J, Bernes C, Junninen K, Lohmus A, Macdonald E, Muller J, Jonsson BG. 2019. Impacts of dead wood manipulation on the biodiversity of temperate and boreal forests. A systematic review. Journal of Applied Ecology 56:1770–1781.
Schiegg K. 2000a. Are there saproxylic beetle species characteristic of high dead wood connectivity? Ecography 23:579–587.
Schiegg K. 2000b. Effects of dead wood volume and connectivity on saproxylic insect species diversity. Ecoscience 7:290–298.
Schmitz OJ, Raymond PA, Estes JA, Kurz WA, Holtgrieve GW, Ritchie ME, Schindler DE, Spivak AC, Wilson RW, Bradford MA, Christensen V, Deegan L, Smetacek V, Vanni MJ, Wilmers CC. 2014. Animating the carbon cycle. Ecosystems 17:344–359.
Seibold S, Bässler C, Baldrian P, Reinhard L, Thorn S, Ulyshen MD, Weiß I, Müller J. 2016. Dead-wood addition promotes non-saproxylic epigeal arthropods but effects are mediated by canopy openness. Biological Conservation 204:181–188.
Seibold S, Weisser WW, Ambarlı D, Gossner MM, Mori AS, Cadotte MW, Hagge J, Bässler C, Thorn S. 2023. Drivers of community assembly change during succession in wood-decomposing beetle communities. Journal of Animal Ecology 92:965–978.
Selva N, Fortuna MA. 2007. The nested structure of a scavenger community. Proceedings of the Royal Society b: Biological Sciences 274:1101–1108.
Shea PJ, Laudenslayer WF, Ferrell G, Borys R. 2002. Girdles versus bark beetle-created ponderosa pine snags: Utilization by cavity-dependent species and differences in decay rate and insect diversity. Service UF editor. Proceedings of the Symposium on the Ecology and Management of Dead Wood in Western Forests, p145–153.
Shorthouse DJ, Iglesias D, Jeffress S, Lane S, Mills P, Woodbridge G, McIntyre S, Manning AD. 2012. The ‘making of’ the Mulligans Flat—goorooyarroo experimental restoration project. Ecological Management & Restoration 13:112–125.
Sikes DS, Raithel CJ. 2002. A review of hypotheses of decline of the endangered american burying beetle (Silphidae: Nicrophorus americanus Olivier). Journal of Insect Conservation 6:103–113.
Spencer EE, Barton PS, Ripple WJ, Newsome TM. 2020. Invasive European wasps alter scavenging dynamics around carrion. Food Webs 24:e00144.
Speziale KL, Lambertucci SA, Olsson O. 2008. Disturbance from roads negatively affects Andean condor habitat use. Biological Conservation 141:1765–1772.
Stiegler J, Von Hoermann C, Muller J, Benbow ME, Heurich M. 2020. Carcass provisioning for scavenger conservation in a temperate forest ecosystem. Ecosphere 11:e03063.
Subalusky AL, Dutton CL, Rosi EJ, Post DM. 2017. Annual mass drownings of the Serengeti wildebeest migration influence nutrient cycling and storage in the Mara River. Proceedings of the National Academy of Sciences of the United States of America 114:7647–7652.
Swift MJ, Heal OW, Anderson JM. 1979. Decomposition in terrestrial ecosystems. Oxford: Blackwell Scientific Publications.
Tews J, Brose U, Grimm V, Tielbörger K, Wichmann MC, Schwager M, Jeltsch F. 2004. Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. Journal of Biogeography 31:79–92.
Thorn S, Bässler C, Brandl R, Burton PJ, Cahall R, Campbell JL, Castro J, Choi C-Y, Cobb T, Donato DC, Durska E, Fontaine JB, Gauthier S, Hebert C, Hothorn T, Hutto RL, Lee E-J, Leverkus AB, Lindenmayer DB, Obrist MK, Rost J, Seibold S, Seidl R, Thom D, Waldron K, Wermelinger B, Winter M-B, Zmihorski M, Müller J. 2018. Impacts of salvage logging on biodiversity: a meta-analysis. Journal of Applied Ecology 55:279–289.
Threlfall CG, Law BS, Peacock RJ. 2019. Benchmarks and predictors of coarse woody debris in native forests of eastern Australia. Austral Ecology 44:138–150.
Traylor CR, Ulyshen MD, McHugh JV. 2023. Forest age drives saproxylic beetle biodiversity in the southeastern United States. Biological Conservation 285:110238.
Ulyshen MD. 2016. Wood decomposition as influenced by invertebrates. Biological Reviews 91:70–85.
Ulyshen MD, Hanula JL. 2009. Responses of arthropods to large-scale manipulations of dead wood in loblolly pine stands of the southeastern United States. Environmental Entomology 38:1005–1012.
Ulyshen MD, Hanula JL. 2010. Patterns of saproxylic beetle succession in loblolly pine. Agricultural and Forest Entomology 12:187–194.
Ulyshen MD, Šobotník J. 2018. An introduction to the diversity, ecology, and conservation of saproxylic Insects. Ulyshen MD editor. Saproxylic Insects: Diversity, Ecology and Conservation. Cham: Springer International Publishing, p1–47.
Vandersteen J, Fust C, Crowther MS, Smith M, Viola B, Barton P, Newsome TM. 2023. Carcass use by mesoscavengers drives seasonal shifts in Australian alpine scavenging dynamics. Wildlife Research 50:1031–1045.
Vindstad OPL, Birkemoe T, Ims RA, Sverdrup-Thygeson A. 2020. Environmental conditions alter successional trajectories on an ephemeral resource: a field experiment with beetles in dead wood. Oecologia 194:205–219.
Vodka S, Konvicka M, Cizek L. 2009. Habitat preferences of oak-feeding xylophagous beetles in a temperate woodland: implications for forest history and management. Journal of Insect Conservation 13:553–562.
Vogel S, Bussler H, Finnberg S, Müller J, Stengel E, Thorn S. 2021. Diversity and conservation of saproxylic beetles in 42 European tree species: an experimental approach using early successional stages of branches. Insect Conservation and Diversity 14:132–143.
von Hoermann C, Benbow ME, Rottler-Hoermann AM, Lackner T, Sommer D, Receveur JP, Bassler C, Heurich M, Muller J. 2023. Factors influencing carrion communities are only partially consistent with those of deadwood necromass. Oecologia 201:537–547.
Wang BR, An SS, Liang C, Liu Y, Kuzyakov Y. 2021a. Microbial necromass as the source of soil organic carbon in global ecosystems. Soil Biology & Biochemistry 162.
Wang J, Guan Y, Wu L, Guan X, Cai W, Huang J, Dong W, Zhang B. 2021b. Changing lengths of the four seasons by global warming. Geophysical Research Letters 48: e2020GL091753.
Wang SP, Loreau M, de Mazancourt C, Isbell F, Beierkuhnlein C, Connolly J, Deutschman DH, Dolezal J, Eisenhauer N, Hector A, Jentsch A, Kreyling J, Lanta V, Leps J, Polley HW, Reich PB, van Ruijven J, Schmid B, Tilman D, Wilsey B, Craven D. 2021c. Biotic homogenization destabilizes ecosystem functioning by decreasing spatial asynchrony. Ecology 102.
Waters TF. 1977. Secondary production in inland waters. Advances in Ecological Research 10:91–164.
White C. 2005. Hunters ring dinner bell for ravens: experimental evidence of a unique foraging strategy. Ecology 86:1057–1060.
Zou D, Tian S, Zhang T, Zhuoma N, Wu G, Wang M, Dong L, Rossiter SJ, Zhao H. 2021. Vulture genomes reveal molecular adaptations underlying obligate scavenging and low levels of genetic diversity. Molecular Biology and Evolution 38:3649–3663.
Acknowledgements
We thank Mike Ferro for kindly providing photographs of the coarse woody debris and leaf litter shown in Figure 2. PSB would like to thank Eric Benbow and Maldwyn Evans for their many stimulating discussions about necromass dynamics. PMT was funded in part by the Regional Government of Asturias (GRUPIN grant IDI/2021/000075). The authors declare no conflict of interest.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions: PB conceived the study, analysed the data, and wrote the first draft of the manuscript. NS, NB, MU, PMT and TN contributed key materials and edits to the manuscript.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Barton, P.S., Schultz, N., Butterworth, N.J. et al. Disentangling Ecosystem Necromass Dynamics for Biodiversity Conservation. Ecosystems 27, 544–558 (2024). https://doi.org/10.1007/s10021-024-00907-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-024-00907-y