
Abstract
Drought timing determines the degree to which dry events impact ecosystems, with the ability of key processes to withstand change differing between drought periods. Findings indicate that drought timing effects vary across ecosystems, with few studies focusing on alpine grasslands. We conducted a mesocosm experiment using small grassland monoliths collected in September from the high Alps and left to overwinter at 0 °C until the experiment began in lowland Italy under late-winter outdoor conditions. Together with watered controls, we imposed three different drought treatments (zero precipitation): (1) one-month early-drought immediately after simulated snowmelt; (2) one-month mid-drought a month after melt-out; and (3) continuous two-month drought across the entire experimental period. Ecosystem responses were assessed by measuring CO2 fluxes, while vegetation responses were investigated by measuring aboveground net primary production (ANPP) of graminoids and forbs and post-harvest resprouting after one-month rehydration. We found that ecosystem respiration and gross ecosystem production (GEP) during the day were more negatively affected by mid-season drought compared to drought starting early in the season. By the end of treatments, GEP reduction under mid-season drought was similar to that of a continuous two-month drought. ANPP reduction was similar in early- and mid-drought treatments, showing a greater decrease under an enforced two-month period without precipitation. Plant resprouting, however, was only reduced in full- and mid-season drought pots, with forbs more negatively affected than graminoids. Seasonal soil moisture variation can account for these patterns: remaining winter moisture allowed almost full canopy development during the first month of the season, despite precipitation being withheld, while soil moisture depletion in the second month, resulting from higher temperatures and greater biomass, caused a collapse of gas exchange and diminished plant resprouting. Our data illustrates the importance of the timing of zero-precipitation periods for both plant and ecosystem responses in alpine grasslands.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Highlights
-
CO2 fluxes are more negatively affected by mid-season than early-season drought
-
ANPP reductions are greater under full-season drought than under early- and mid-drought
-
Plant resprouting is reduced most after mid- and full-season drought
Introduction
Many grasslands are experiencing drought stress globally (Wang and others 2019) following the intensification and increasing frequency of extreme dry events (IPCC 2021). When physiologically effective, drought—here defined as a period characterised by precipitation absence or marked rainfall deficiency relative to normal (Slette and others 2019)—can negatively affect plant growth, reducing meristem activity and plant nutrient uptake (Chaves and others 2003) through increased vapour pressure deficit and reduced soil water content. These conditions can eventually determine overall reductions in ecosystem net primary productivity, in some cases leading to plant death and reduced net carbon uptake (for example, review by Sippel and others 2018). Grassland responses, however, have been found to vary according to drought intensity, frequency, and co-occurrence with other extreme events (De Boeck and others 2016; Hoover and Rogers 2016; Zhang and others 2019a). In particular, the timing at which drought occurs has been identified as a key aspect modulating ecosystem responses to drought (Sippel and others 2018). For instance, across a 27-year study, the effect of drought on aboveground net primary production (ANPP) of a temperate, humid grassland was found to be stronger in the early growing season, declining throughout the season and being negligible in August (Craine and others 2012). Similarly, herbaceous species in a sagebrush steppe exhibited lower productivity when a 20% precipitation reduction was applied over 3 months during spring, with no effect when precipitation reduction occurred during the winter (Bates and others 2006).
Drought timing can also affect plant growth when occurring in different periods of the growing season, but effects are not always consistent. In the few studies on mesic and semi-arid grasslands, greater reductions of ANPP under later drought events were found in some cases (Denton and others 2017; Zhang and others 2020a; Hahn and others 2021), whereas a greater impact of an earlier drought was found in others (Meng and others 2019).
Temperature, soil moisture and plant-development stages have all been recognised as the main drivers of such variability, representing intrinsic inter-related features which vary not only across ecosystems but also over time, with strong inter- and intra-annual differences (De Boeck and others 2010). The level and state of these factors determine whether an ecosystem is functionally affected by drought (De Boeck and others 2011), with the composition of the plant community also driving responses, since drought impact could be more intense when occurring during a critical period for growth and reproduction of dominant species (Dietrich and Smith 2016; Meng and others 2019). In this context, it can therefore be argued that results deriving from a small group of studies focusing on temperate and semi-arid grasslands cannot be generalised to fit all herbaceous ecosystems. This is especially true for alpine grasslands, where few studies yet exist assessing the impacts of drought timing (Hu and others 2021; Zhao and others 2022).
Alpine grasslands are snow-dominated ecosystems, characterised by a short growing season with high levels of soil moisture in the period immediately after snowmelt (Williams and others 2009) and made up of plant communities mainly consisting of low-statured, perennial, herbaceous species, with fast development rates enabling them to cope with the brief snow-free period (Körner 2021). Given these characteristics, early-season drought could have a greater impact on alpine grasslands since it coincides with the most active period of plant growth (Bates and others 2006)—by mid-summer alpine species have almost completed their development (Jonas and others 2008). On the other hand, moist post-snowmelt conditions could prevent alpine plants from experiencing limitations to their growth at the beginning of the growing season in the absence of rainfall. In this scenario, a mid-summer drought could reduce plant activity and ecosystem functioning to a greater extent as a result of the concurrence of lower soil moisture, warmer temperatures and greater aboveground biomass which, in turn, increase moisture demand and, consequently, the impact of drought (Wang and others 2007).
To our knowledge, the only studies testing the effects of drought timing on high-elevation grasslands have been focused on plant phenology and water use efficiency in the Tibetan Plateau, indicating later green-up and earlier senescence under early- and mid-season drought, respectively (Hu and others 2021) and higher water use efficiency under early-season drought compared to mid-season drought (Zhao and others 2022). The reason for such a scarcity of studies could be due not only to the technical challenges of setting-up and maintaining precipitation exclusion experiments in alpine environments—difficult to reach and/or characterized by harsh conditions (Hoover and others 2018; Vorkauf and others 2021)—but also to the fact that in many alpine regions water supplies are not limiting for plants during the summer (Körner 2021). Furthermore, the majority of alpine plant life is regarded as robust “against variability and unpredictability of climatic conditions”, mainly due to the clonal life strategy characterizing the majority of species and the strong changes in environmental conditions over a short distance and short time span (Körner and Hiltbrunner 2021). This being said, experimentally reduced precipitation over the growing season has effectively been found to negatively affect alpine grasslands (Gilgen and Buchmann 2009; De Boeck and others 2016; Liu and others 2018) and drought-related wilting and plant mortality has been reported for alpine species (Griffin and Hoffmann 2012; personal obs.). Together with precipitation deficits occurring in mountain regions, especially in the Alps, over the last two decades (for example, Stephan and others 2021), not least the 2022 summer drought in Europe (Toreti and others 2022), the aforementioned findings underline the necessity to investigate whether drought seasonality could determine the extent to which ecosystem processes in the alpine life zone will be affected by dry events.
To this aim, not only ANPP, but also CO2 fluxes—that is, ecosystem respiration (ER), gross ecosystem production (GEP) and net ecosystem exchange (NEE)—were considered in this study, representing a non-destructive measure of vegetation and ecosystem functioning under stress. Both ER and GEP are, in fact, highly sensitive to variations in temperature and soil moisture conditions (for example, von Buttlar and others 2018) and can therefore provide insights into biological responses to drought. In addition, the drought response of these two processes determines the role of CO2 sink/source of an ecosystem. In some studies, GEP exhibited higher drought sensitivity (that is, greater reduction) compared to ER (Hoover and Rogers 2016; Liu and others 2019; Zhang and others 2019a), while in others an opposite pattern was observed (von Buttlar and others 2018). Such variable CO2 flux responses could be due to different soil moisture levels at which autotroph and heterotroph activity is limited, or even ceases, with the former exhibiting much higher water potential values (− 1.5 MPa for herbaceous species; Lambers and others 2008) than the latter (up to − 36.5 MPa; Manzoni and others 2012). While ecophysiological thresholds in response to water shortage have been investigated at a species or functional group level (Belluau and Shipley 2017; Schimel 2018), few studies have so far attempted to identify thresholds encompassing biocoenosis responses (Manzoni and others 2012). Furthermore, different responses to drought of CO2 fluxes have also been observed between drought periods, in both observational (Liu and others 2019) and experimental studies (Hao and others 2018; Zhang and others 2019b), suggesting a dependence of soil moisture thresholds on drought seasonality.
In this study, we define soil moisture thresholds as the soil moisture values at which a non-linear change in ER and GEP rates occurs. To obtain, identify, and compare soil moisture thresholds for CO2 fluxes during different drought periods, we carried out a mesocosm experiment (sensu Odum 1984) using small grassland monoliths (1.7 L) from an alpine grassland dominated by Carex curvula. ANPP of graminoids and forbs and their resprouting ability (after drought, clipping and rehydration) were also assessed to detect vegetation responses. This mesocosm experiment, performed using small monoliths to impose severe drought, was carried out after no significant effect on CO2 fluxes and ANPP was detected in situ (personal obs.) following an experimental precipitation reduction simulating a site-specific, statistically extreme rainfall amount during the snow-free period (equivalent to a 1-in-100-year drought). We hypothesised that under the current, short snow-free season, soil moisture levels in the field may still be too high to determine any detectable drought effect in the target study area (Italian Alps). The use of a restricted soil volume would, therefore, allow us to detect potential effects that drought periods might produce under a warmer climate scenario, characterised by earlier snowmelt, longer growing seasons and higher evapotranspiration, during which plant-available soil moisture pools are more likely to become exhausted.
Specifically, the following questions were addressed: (1) Do soil moisture thresholds for ER and GEP and the drought-induced, temporal dynamics of these processes depend on drought seasonality? (2) Is the extent of drought impact on CO2 fluxes, ANPP and plant resprouting dependent on drought timing and length?
Methods
Field Study Site
The field research site (46°21′14" N, 10°29′24" E) is located at about 2700 m a.s.l. in the Italian Rhaetian Alps, specifically in the Gavia valley (Stelvio National Park). The site is characterised by continuous snow cover for most of the year and a brief snow-free period of about 4 months (June–September) with mean summer air temperature at 20 cm above the ground equal to 7.5 °C and precipitation of about 380 mm. The target ecosystem is a primary alpine grassland dominated by Carex curvula All. (Figure S1), developing on siliceous bedrock, with mean ANPP of vascular plants of about 200 g m−2 and mean soil depth of 31 cm.
Experiment Design
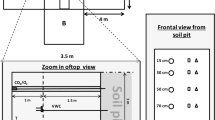
The mesocosm experiment was set-up in the University of Parma garden site (72 m a.s.l., 44°46′06" N, 10°18′53" E). In September 2019, 35 grassland monoliths were collected from the field and transplanted into pots (12 cm diameter, 15 cm depth; Figure S2a) which were then left to overwinter in an incubator for 5 months at 0 °C. At the beginning of February 2020, when air temperatures in Parma (c. 5 °C) were similar to those recorded after snowmelt in the field (Figure S3), pots were removed from the incubator and saturated with distilled water until soil water potential at field capacity (c. − 10 kPa) was reached, in order to simulate soil saturation following snowmelt (Figure S2b). Pots were then weighed and placed under a shelter made of 4 m × 6 m transparent polythene foil covering the mesocosm, used to simulate drought. To avoid soil heating, pots were settled in basins insulated with polystyrene boards and filled with expanded clay (Figure S2c). Container bases were pierced to avoid water stagnation and basins were then placed at 1.20 m from the ground (Figure S2c, d). Two pairs of halogen lamps (Figure S2c) lit the basins two hours before sunrise and after sunset to increase photosynthetic active photon flux density (PPFD, μmol m−2 s−1) and solar radiation (W m−2) and also to simulate a photoperiod similar to that experienced in the field (c. 16 h). All pots were randomly re-located twice a week to avoid bias due to pot location within the mesocosm.
Once the experiment began (8 February, day 0 of experiment), 29 pots were randomly assigned to one of four treatments—control (C, 8 replicates) and 3 drought treatments (7 replicates each) (Figure 1)—with the remaining 6 pots assigned to microclimate measurements. Control pots, together with drought treatments depending on the period, were watered twice a week with distilled water (Figure 1) in an amount simulating average daily rainfall from June to September based on the historical records of a weather station near the field site (Careser Diga, 2600 m; Figure S4). Drought treatments consisted of: (1) an early-season drought (ED: 32 days without watering after simulated snowmelt, followed by one-month re-watering); (2) a mid-season drought (MD: watering during the early-season followed by 31 days of drought); and (3) a continuous full-season drought (FD: 62 days without watering, starting as ED and ending as MD). This last treatment was added to the experiment to guarantee the identification of thresholds triggering different ER and GEP rates for drought starting early in the season and assess vegetation responses to different drought periods.

Experimental design implemented to test the effects of drought timing. The four treatments are reported together with relative sample size (number of pots) and drought length; measurement dates of CO2 fluxes, ANPP and resprouting are specified at the bottom, together with the frequency at which pots were weighed to assess soil moisture dynamics (green dots). The date of simulated snowmelt is also reported.
Mean air temperature measured in our mesocosm, as well as minimum and maximum values, were similar to those recorded in the field during the summer (Figure S3). Shelter effects on PPFD, solar radiation and air temperature were quantified by taking hourly measurements at 1.20 m aboveground both under and outside the shelter. Average daily PPFD under the shelter was 16% lower compared to values recorded outside (Figure S5a), while solar radiation was reduced by 10% (Figure S5b). Over the entire study period, mean daily air temperature recorded under the shelter was similar to that measured outside (Figure S6).
Microclimate
Soil moisture metrics, that is, volumetric water content (VWC, %) and water content in mm, were estimated based on pot volume and mass, recorded for each pot twice a week before watering. At the end of the experiment (12 May), all pots were oven-dried at 100 °C until constant mass was reached in order to quantify the amount of soil present per pot. From each of the 24 pot-mass recording dates, soil mass was subtracted to obtain pot water mass during the experiment. Water mass data in one FD pot was also used to assess the relationship between soil VWC and soil water potential (SWP, kPa) during drought (Figure S7). In addition, the top soil layer temperature (0–2 cm) was recorded hourly with thermistors in 6 pots (3 in both C and FD treatments) in order to assess soil temperature differences between the two drought periods and detect any difference in temperature due to the drought treatment imposed.
CO 2 Fluxes
CO2 fluxes were recorded weekly (Figure 1) using a PP System EGM-4 infrared gas analyser connected to a transparent, cylindrical chamber sealed over each pot (2000 cm3 volume, 102 cm2 area). Ecosystem respiration (ER) was recorded in dark conditions by covering the chamber with an opaque sheet. Subsequently, in natural light, measurements of net ecosystem exchange (NEE) were made. Measurements lasting 60 s. were taken under clear sky conditions during the central hours of the day, prior to re-watering. Based on the ideal gas law, the change in CO2 concentration (ppm) over time was used to estimate CO2 flux rates (µmol m–2 s–1) by linear regression. Once ER rates were estimated, values were used to calculate GEP, computed by subtracting the mean ER rate from the corresponding measurement in light and fitting linear regressions. Since in the present study NEE represents CO2 fluxes going from the ecosystem into the atmosphere, negative values (GEP) indicate CO2 uptake, whereas positive values (ER) indicate CO2 release.
Vegetation Responses
Vegetation responses were estimated for all monoliths by assessing ANPP at the end of drought treatments (10 April) and within-season resprouting one month later (12 May), after clipping and rehydration of all pots (Figure 1). ANPP (g m−2) was estimated by harvesting aboveground vascular plant phytomass (that is, biomass plus attached necromass), which was divided into species, and then oven-dried at 65 °C for 48 h. ANPP was then reported for graminoids and forbs and expressed per cover unit, dividing ANPP by the proportion of vegetation cover estimated for each pot using pre-harvesting photos (Figure S8). This procedure was followed to take into account the lower cover values recorded in ED (83%) compared to the other treatments (96% in C, 96% in MD and 93% in FD) due to original plant cover in the monoliths (Figure S8).
To assess plant resprouting after clipping, the number of modules of each vascular species was counted before phytomass harvesting (a module was defined as a reiterated multicellular subunit, such as tillers for graminoids and rosettes for forbs; Table S1 for details). Phytomass was then clipped and pots were left to rehydrate under ambient precipitation for one month, after which the number of resprouting modules (that is, modules producing new leaf tissue) per species was counted. Species were then grouped into graminoids and forbs and, for all 29 pots, resprouting was reported as the percentage of modules counted after clipping and rehydration relative to the number of modules counted before harvesting.
Data Analysis
Data analysis was carried out in R (R Core Team 2020). Soil moisture thresholds triggering non-linear changes in GEP and ER rates were assessed separately for each flux and drought period onset. Specifically, threshold analysis of the first drought period included flux estimates in ED (until 11 March) and FD (until end of drought experiment), whereas analysis focused on the second period was based on MD fluxes (from 12 March to end of drought experiment). GEP estimates above 0 (10% of GEP records) were set equal to 0 and the first two measurement dates were removed from the analysis of early-drought thresholds for GEP to avoid bias due to the early phenological development of vegetation. Two-phase linear models were employed using the R package “chngpt” (Fong and others 2017). Selection of the most parsimonious model was carried out by means of the likelihood ratio test [package “lmtest” (Zeileis and Hothorn 2002)], comparing different piecewise regression models (hinge, upper hinge, segmented and stegmented models; Figure S9a) and simple linear models. To take into account the influence of hierarchical structures in data (due to repeated measurements of the same pots at fixed dates), models including different random effect structures, fitted with the package “lme4” (Bates and others 2015) were compared (by means of the AIC) with corresponding models including only the fixed component. Because the random effect structures did not improve model fitting, the random component was excluded from all models. Threshold model estimates were based on 1000 permutations and a visual check of residual patterns was performed to assess linear model assumptions. For both ER and GEP, discontinuous models (stegmented, Figure S9b) were selected as the best-fit of moisture thresholds, except for GEP under early-drought onset, for which a continuous (hinge) model was selected.
To evaluate ER, GEP and vegetation responses in the different drought treatments, pot-level normalised-to-control data was used and calculated as (\({Y}_{\mathrm{D}}-{\overline{Y} }_{\mathrm{C}})/{\overline{Y} }_{\mathrm{C}}\) where YD is the response variable measured in each drought pot and YC is the average recorded in controls. In particular, CO2 flux responses were assessed on data from the last measurement date of each drought period. One sample t-tests were performed on normalised ER, GEP and ANPP to assess differences between drought treatments and controls. Linear mixed-effect models fitted with generalised least squares through the package “nlme” (Pinheiro and others 2020) were carried out to assess the influence of different drought periods. The model employed to test CO2 flux responses included as explanatory variables the flux type (2-level predictor: ER and GEP), the drought treatment (3-level predictor: ED, MD and FD) and their interaction. To achieve homoscedasticity, three variance structures (related to flux rates, treatments and flux types) were included. For ANPP, the model included as explanatory variables the drought treatment, the plant functional type (2-level predictor: graminoids and forbs) and their interaction, with treatment variance being homogenised. Finally, the linear model for graminoid and forb resprouting included as explanatory variables the drought treatment (3-level predictor), the functional type (2-level predictor) and their interaction; the variance of treatments and functional types was homogenised to improve homoscedasticity. One sample t-tests were performed to assess differences between drought treatments and control. For CO2 fluxes, ANPP and resprouting models, Tukey post-hoc tests were carried out to assess differences among predictor levels with the package “multcomp” (Hothorn and others 2008).
Results
Microclimate
During the two months of the experiment, control (C) pots received 186 mm of water, whereas pots experiencing drought in early- (ED) and mid-season (MD) received 96 mm (after drought) and 93 mm (before drought), respectively. On day 0, soil moisture was similar in all treatments (mean ± SE): 40 ± 1% in C, 42 ± 2% in ED, 45 ± 3% in MD and 41 ± 2% in FD (Figure 2a). By the end of the first drought period, VWC in non-watered pots was 16 ± 1% (ED) and 15 ± 1% (FD), c. − 63% compared to initial values. A greater reduction was recorded at the end of the second drought period, where VWC in the one-month mid-season drought (MD) pots was on average 8 ± 1%, that is, − 81% compared to the mean value at the beginning of treatment (42 ± 2%, day 32). This reduction was similar to that measured after two months in FD pots, where VWC was as low as 5 ± 0.4% (− 88% compared to initial soil moisture). At the end of the experiment, soil moisture in re-watered ED pots was on average 23 ± 2%, showing similar values to those recorded in C pots (24 ± 1%).

a Volumetric water content (%, with corresponding soil moisture in mm) in the four treatments (C–control, ED–early-drought; MD–mid-drought; FD–full-drought) during the experiment and b cumulative water loss (mm) and soil temperature sum (°d) estimated for drought starting in the early- and mid-season (dashed lines). Dots represent average values for each measurement date; shaded areas represent SE (in (a) n = 8 for C and n = 7 for ED, MD and FD; in (b) water loss: n = 14 in early drought onset, n = 7 in mid-drought onset; soil temperature: n = 3).
Evapotranspiration differed across the two drought periods (Figure 2b), with lower values when drought started in the early-season (1.05 mm d−1 water loss rate) compared to mid-season onset (1.57 mm d−1). In addition, evapotranspiration rates were not constant within each period, being higher during the earlier, moister part: pots experiencing an early-drought onset exhibited evapotranspiration rates of 1.30 mm d−1 and 0.88 mm d−1 in the first and the second half of the one-month drought period, respectively; the same pattern was seen in pots under mid-season drought, with evapotranspiration rates of 2.73 mm d−1 and 0.93 mm d−1 in the first and the second half of the one-month drought, respectively. During the second month of drought, evapotranspiration further declined, with rates of 0.67 mm d−1 and 0.39 mm d−1 in the first and the second half of the second month of drought, respectively.
Top soil layer temperature recorded in C was higher in the second part of the experiment (mid-season) compared to the first part (Figure 2b): cumulative temperature between the start and end of each drought period was 339°d and 414°d for early- and mid-season, respectively (corresponding to a difference of c. 3 K in mean daily temperature between the two drought periods). Soil temperature was not affected by drought, with C and FD pots exhibiting similar values over the entire study period (Figure S10).
Soil Moisture Thresholds for ER and GEP
A schematic diagram to support result interpretation is provided in Supplemental Material (Figure S9b). Soil moisture thresholds for ER and GEP were reached under drier soil conditions when zero precipitation was imposed from the beginning of the growing season rather than one month later (Table 1, Figure 3). Under early-drought onset, ER rate significantly declined at 12% VWC (corresponding to − 59 kPa SWP, Figure S7), whereas GEP rate ceased at 7% VWC (− 1234 kPa SWP); soil moisture levels were reached after 37 and 52 days without precipitation, respectively (Figure 2a). Under mid-drought onset, the threshold for ER (Table 1, Figure 3) was reached at 22% VWC (− 19 kPa SWP), while for GEP it was reached at 14% VWC (− 40 kPa SWP); soil moisture levels were reached after 10 and 18 days without precipitation, respectively (day 42 and day 50 in Figure 2a).

Two-phase linear models of soil moisture thresholds (volumetric water content, %) for ecosystem respiration (ER) and gross ecosystem production (GEP) in μmol m–2 s–1. Thresholds are reported for drought starting in the early season (blue) and mid-season (red) and respective soil moisture values are indicated by the dashed lines; shaded areas represent standard errors of threshold estimates.
ER and GEP Response to Drought
At the end of the respective drought periods, CO2 fluxes were significantly reduced in all drought treatments compared to C (Figure 4). ER exhibited a 52% and 72% reduction in ED (t6 = − 8.21, p < 0.001) and MD pots (t6 = − 22.23, p < 0.001), respectively, and a 94% reduction in FD (t6 = − 61.31, p < 0.001). On the other hand, GEP was reduced by 80% in ED pots (t6 = − 12.63, p < 0.001) and by 99% in MD (t6 = − 273.38, p < 0.001) and FD (t6 = − 1749.00, p < 0.001). The extent of CO2 flux reduction to drought differed across treatments (F2,36 = 395.06, p < 0.001) and between ER and GEP (F1,36 = 123.86, p < 0.001). When comparing the two drought periods, the reduction of ER and GEP rates was more pronounced under mid-season drought, while GEP rates showed a greater reduction than ER rates across all drought treatments. Similar GEP reduction was found in MD and FD pots, while ER rates in MD were higher than those in FD, indicating different drought sensitivity of CO2 fluxes to diverse drought treatments (treatment × flux type interaction: F2,36 = 69.65, p < 0.001).

Response to drought of ecosystem respiration (ER) and gross ecosystem production (GEP) estimated at the end of each drought treatment: 11 March for ED (early-drought, n = 7) and 07 April for MD (mid-drought, n = 7) and FD (full-drought, n = 7). Null values of normalised CO2 fluxes correspond to control data, whereas negative values indicate rate reductions. All values for each treatment are reported (coloured dots), as well as their mean (black dots). Asterisks indicate significant differences from controls following the one sample t-test (***p < 0.001), while different letters indicate significant differences between drought-treatments (post-hoc Tukey contrast, p < 0.05).
Vegetation Responses: ANPP and Resprouting
ANPP was significantly reduced in all drought treatments compared to C (Figure S11). Graminoids exhibited significant ANPP reductions (Figure 5a) in ED (− 31%; t6 = − 3.36, p = 0.015) and FD (− 41%; t6 = − 12.05, p < 0.001), but not in MD (t6 = − 1.40, p = 0.210). On the other hand, forbs exhibited significantly lower ANPP in all drought treatments when compared to controls: ED (− 28%; t6 = − 5.32, p = 0.002), MD (− 21%; t6 = − 2.65, p = 0.038), and FD (− 56%; t6 = − 12.09, p < 0.001).

a Response to drought of ANPP and b within-season plant resprouting after clipping estimated for both plant functional types (PFTs) and for each treatment: ED (early-drought, n = 7), MD (mid-drought, n = 7) and FD (full-drought, n = 7). Null values of normalised ANPP and plant resprouting correspond to control data, whereas negative values indicate reductions. In (a) all values for each treatment are reported (coloured dots), as well as their mean (black dots), with asterisks indicating differences from controls (one-sample t-test: n.s. not significant, *p < 0.05, **p < 0.01, ***p < 0.001). In (b) all values for each treatment are reported, together with means ± 1 SE for recovery (vertical bars) and soil moisture (horizontal bars). For both graphs, different letters indicate significant differences between groups (post-hoc Tukey contrast, p < 0.05).
The extent of ANPP reduction to drought significantly differed across treatments (F2,38 = 10.31, p < 0.001), with the greatest reduction in FD. Forbs exhibited a more pronounced decline in ANPP under drought compared to graminoids (F1,38 = 4.22, p = 0.047), but no significant interaction between functional type and treatment was found. Given such a consistent effect of drought treatments on functional types, similar reductions across treatments were also found for total ANPP, where reduction compared to C was equal to 30% in ED, 19% in MD and 49% in FD (Figure S11).
Plant resprouting (Figure 5b) differed between drought treatments (F2,36 = 32.24, p < 0.001) and functional types (F1,36 = 7.31, p = 0.010), with forbs exhibiting lower resprouting values than graminoids. Treatment effects were not consistent between the two functional types (significant interaction term: F2,36 = 4.05, p = 0.026).
In ED pots, graminoids exhibited full resprouting after clipping, with values equivalent to C (t6 = 1.99, p = 0.094), whereas forb resprouting was slightly reduced compared to C (t6 = − 2.49, p = 0.047); no significant difference was found between graminoids and forbs. Lower resprouting levels compared to C were found for both graminoids and forbs in MD (graminoids: t6 = − 3.29, p = 0.016; forbs: t6 = − 9.51, p < 0.001) and FD (graminoids: t6 = − 6.18, p = 0.001; forbs: t6 = − 167.78, p < 0.001). In MD, forbs exhibited lower resprouting levels than graminoids (− 74% and − 22%, respectively, compared to C). The same pattern was also found in FD, with both functional types showing the lowest resprouting levels compared to controls (− 99% for forbs and − 62% for graminoids). The relationship between resprouting (normalised to C) and minimum soil moisture recorded in each pot during the experiment (Figure 5b) highlights the different responses of graminoids and forbs to declining VWC during drought periods.
Discussion
Soil Moisture Thresholds for ER and GEP
Soil moisture thresholds for CO2 fluxes—indicators of a non-linear decrease in ER and GEP rates under drought—occurred at higher moisture levels when drought was applied later during the growing season, that is, one-month after snowmelt. In particular, under mid-drought CO2 fluxes showed non-linear declines at soil volumetric water content (VWC) of 22% for ER and 14% for GEP, a soil moisture reduction of approx. half compared to initial values (− 52% and − 69% of VWC for ER and GEP, respectively). In contrast, under early-drought onset, CO2 flux thresholds occurred at 12% VWC for ER and 7% for GEP, a soil moisture reduction of 71% and 83%, respectively, compared to initial VWC.
To our knowledge, no field data is currently available for a comparison with the soil moisture thresholds for CO2 fluxes identified in the present study, although there are reports of greater reductions in photosynthesis, plant growth and heterotrophic respiration rates following late-season drought, explained by initial soil moisture, temperature and ecosystem development stage at the time of drought (De Boeck and others 2011; Vitra and others 2019; Yang and others 2020). The experimental approach implemented in this study allows the exclusion of a role for initial soil moisture, since monoliths exposed to mid-season drought were kept moist until the beginning of the zero-precipitation period. As the two one-month drought periods in this study began with similar soil moisture levels, our thresholds would suggest that CO2 emission and uptake processes in high-elevation grasslands are potentially more affected by a mid-season water deficit.
The warmer temperatures and the full canopy development characterising the middle of the growing season are intrinsic features of the alpine ecosystem, which cannot be disentangled from the impact of drought timing. At the same time, these are pivotal factors driving the temporal dynamics of CO2 fluxes, promoting higher evapotranspiration rates and, consequently, more rapid soil moisture depletion (Wang and others 2007; De Boeck and others 2011).
Furthermore, results indicate that soil moisture thresholds not only differ for ER and GEP but are also subject to seasonal dynamics, probably related to the different components involved (autotrophs and heterotrophs) and their phenology. In fact, ER rates exhibited non-linear reductions at higher soil moisture levels compared to GEP, initially suggesting ER was more negatively affected by decreasing water availability. However, null values were never recorded for ER, as opposed to GEP. Higher soil moisture thresholds for ER could be a result of soil drying out in the upper layer (Borken and Matzner 2009 and references therein), where the majority of plant roots and microbial communities are found in these grasslands (Grabherr 1989).
Regarding seasonal dynamics, patterns of GEP reduction differed according to drought timing: under mid-drought onset, GEP exhibited an abrupt, discontinuous reduction; under early-drought onset, a continuous GEP decline was recorded, until CO2 uptake completely ceased. Although the abrupt reduction could depend on plant wilting under mid-drought (noted in mid-drought pots, personal observ.), the continuous decrease in GEP could be explained by aboveground biomass reduction (Noormets and others 2008) and/or earlier leaf senescence (De Boeck and others 2016; Stampfli and others 2018) which, in turn, could have led to lower stomatal conductance and assimilation capacity (Zhang and others 2020b).
CO 2 Flux Responses to Drought at the End of Each Drought Period
Data gathered at the end of each drought period for ER and GEP showed that the reduction rate was greater under mid-drought, confirming results from the threshold analysis. In particular, GEP reduction at the end of the one-month mid-drought equalled that found in the most extreme two-month drought treatment. Precipitation exclusion studies in the field are usually subject to treatment–control covariation which can alter the magnitude of the treatment, making it difficult to assess short-term ecological responses (Hoover and others 2018). Therefore, by comparing two drought periods of equal length, our results overcome the limitations of both observational and experimental field studies, thus supporting the conclusion that CO2 fluxes in the investigated grassland are more sensitive to mid-season drought. This result would also seem to apply to net ecosystem exchange (NEE, Figure S12) which was strongly reduced compared to controls after c. 3 weeks of mid-season drought, whereas no change in NEE rates was observed under drought in the early season. It should be noted, however, that this observation is only valid for the central hours of the day: overall NEE (beyond the scope of this study) substantially depends on CO2 fluxes occurring over a 24-h period throughout the growing season and, in these high-elevation ecosystems, on ER dynamics occurring under snow cover.
Interestingly, results revealed that GEP had declined more than ER by the end of each dry event, in line with previous findings under both natural climate extremes (Schwalm and others 2012; Liu and others 2019) and experimental conditions (Hoover and Rogers 2016). However, when considering normalised CO2 fluxes recorded in drought treatments (Figure S13) during the entire experiment, ER in drying pots fell below control rates at higher soil moisture levels (38% VWC) compared to levels for GEP (30% VWC). Soil drying of the top layer and consequent reduction of microbial activity could once more explain this result.
ANPP and Plant Resprouting Under Early- Versus Mid-season Drought
A similar reduction of ANPP was found in both drought-timing treatments, although a tendency towards a greater reduction of total ANPP was noted in the early treatment (− 30% under early-drought vs − 19% under mid-drought). Despite early-drought monoliths being watered for one month after the zero-precipitation period, it appears that vegetation was not able to achieve control values, a result in line with findings showing that negative effects on ANPP are greater during the most active growth period (Darenova and others 2017; Lemoine and others 2018). Not surprisingly, the greatest ANPP reduction (− 49%) was found in the most extreme (two-month) drought. Interestingly, ANPP reduction was less pronounced than that recorded for CO2 fluxes. Because ANPP integrates plant growth over a longer period than the experimentally imposed one-month drought, primary production would appear to be more stable when experiencing short-term water shortage.
Notably, resprouting levels differed between the two drought-timing treatments: plants experiencing zero precipitation in the first part of the growing season exhibited a full within-season resprouting (that is, vegetative growth after drought and clipping), as opposed to lower resprouting levels under mid-drought, where values were similar to those of a two-month period without precipitation. This reduced recovery could be due to faster soil moisture depletion recorded under mid-drought, with a higher amount of plant biomass exposed to stressful conditions—plant biomass is related to the extent to which climate extremes could impact plant communities (Wang and others 2007; van Ruijven and Berendse 2010). Higher evapotranspiration due to warmer temperatures and full canopy development caused leaf wilting under mid-drought which resulted either from unsatisfied water demand or insufficient time available for plants to adapt to the rate of soil drying (Volaire 2018).
As expected, forbs were more drought-sensitive than graminoids, confirming results found in previous experimental studies (for example, Winkler and others 2016; Rosbakh and others 2017) and observations following the 2015 and 2018 dry summers in the Central Alps (Körner 2021). In the current study, graminoid responses were mainly driven by the sedge Carex curvula, a species found to have a strong tolerance to dehydration, which exhibited fully turgid, green leaves 8 days after the end of a 44-day experimentally imposed desiccation (Körner 2021 and reference therein). On the other hand, forb responses were mainly driven by (1) snowbed species (Alchemilla pentaphyllea and Soldanella pusilla), which thrive in areas where the snow-pack provides water at the beginning of the short snow-free period, and/or (2) species with a relatively large leaf area (Lentodon helveticus and Ligusticum mutellina). Such characteristics account for the increased sensitivity to low water availability of these species (Peterson and Billings 1982; De Boeck and others 2016).
In general, these outcomes suggest that (1) measurements of recovery, based on resprouting or other data, are necessary, since ANPP used alone only assesses within-season vegetation responses and does not provide a complete picture of future drought impact, and (2) the different extent to which drought timing acts on functional types in the short term could translate into different times of recovery and/or the triggering of accelerated vegetation dynamics. Lagged effects may occur when changes in plant and soil characteristics take place, including reduction in carbohydrate reserves in plants, changes in soil microbial communities, higher soil nitrogen availability following drought, and plant death (van der Molen and others 2011; Sippel and others 2018). Considering results from the present study, alpine grasslands could, for instance, completely recover ANPP after early-drought stress in a relatively short period (1–2 years after drought, De Boeck and others 2018), whereas legacy effects from a mid-season dry period may last longer if plant death occurs. In addition, plants subject to mid-season water deficit may not have the time to recover during the late season (De Boeck and others 2011), because in this period alpine grasslands are already moving towards senescence (Erschbamer and Winkler 1995).
Implications and Limitations of the Study
With drought events becoming more frequent and drought-induced wilting being observed at high elevations (Griffin and Hoffmann 2012; Körner 2021; Vorkauf and others 2021), it is worth understanding whether soil moisture thresholds estimated in the present study occur in the field. A similar exercise for Carex curvula grasslands was carried out by Körner (2021), who calculated the likelihood of soil moisture depletion based on field data, using average evapotranspiration and water storage of different types of soil at different depth. The author estimated 24 bright days (with an average evapotranspiration of 4 mm d−1) for moisture depletion to occur—calculations were made for soils with a depth of 25 cm and high humus content, 10% rock debris and initial soil moisture of 95 mm. In our mesocosm, with monoliths characterised by a similar soil type, but a depth of 15 cm, we found that it took almost 2 months to reach near-to-null soil moisture values when zero-precipitation was imposed immediately after snowmelt, with only one month necessary to reach similar soil moisture levels when no water was supplied in mid-season. In addition, our results indicate that soil moisture depletion occurred at a different speed (1) within each drought period, with higher water loss in the earlier, moister days, and (2) between drought periods, with greater evapotranspiration in mid-season.
In addition, by measuring non-linear changes in CO2 fluxes in relation to soil moisture, we were able to detect the number of days without precipitation necessary for physiologically effective soil moisture depletion to occur: only 10 and 18 days were required for mid-season drought to produce significant alterations of ER and GEP, respectively, while threshold values were reached a month after a dry event starting in the early season. During the last two decades, the average length of the longest rainfall-free period between June and August was 7 days, with a maximum of 15 days (mean 10 days and max. 17 days for days with < 1 mm precipitation; Figure S14). In addition, the good relationship between volumetric water content derived from water mass and that registered by probes (Figure S15) allows us to compare soil moisture thresholds for ER and GEP with VWC measured in the field where monoliths were collected (Figure S16). Records show that it is unlikely, though not impossible, for threshold values to occur under natural conditions in the 0–7.6 cm soil profile: the lowest soil moisture in the field (12.7% VWC), below the thresholds for ER and GEP (VWC of 22% and 14%, respectively) were, in fact, recorded in the 2022 mid-season, which was characterised by extreme heat and drought conditions (Toreti and others 2022).
With regard to the limitations of this study, problems connected with methods for CO2 flux estimates must first be acknowledged, as these can be biased by, for example, overestimation of daytime ER (and hence of GEP) (Heskel and others 2013) or artefact effects due to the use of a closed chamber, such as modified soil and air temperatures (Kutzbach and others 2007) or altered atmospheric pressure (Brændholt and others 2017). This bias could represent a serious issue when estimating ecosystem C balance. In this study, however, CO2 flux data was only used for treatment comparison, so we believe that methodological issues do not modify the main outcomes of our analysis.
A second limitation of our study derives from the use of small grassland monoliths that, while guaranteeing the identification of thresholds, are unable to capture the complexity of natural ecosystems (Kreyling and others 2014). On the one hand, small monolith volume enables the water supply to be limited, simulating conditions that could occur in a warmer climate scenario. On the other hand, pots are confined environments with altered biotic interactions and size-related biological constraints, such as reduced soil resources and root growth (Poorter and others 2012 and references therein). Together with drought duration, rooting depth and physical soil proprieties determine the physiologically effectiveness of drought and under natural conditions a small fraction of roots of alpine plants can reach deeper layers, thus securing a water supply even under superficial soil desiccation (Körner 2021). Consequently, it is likely that both response magnitude and non-linear changes in CO2 fluxes in our mesocosm are greater than in the field. Despite these limitations, we believe our results confirm the importance of drought timing for key processes in high-elevation ecosystems and provide useful insights for the interpretation of field results.
Conclusion
Experimentally enforced drought seasonality influences the degree to which alpine grasslands are affected by dry events in the short-term. Drought occurring in mid-season is more likely to determine non-linear reduction of ER and GEP, plant wilting, and a reduction of within-season plant resprouting, resulting from faster soil moisture depletion due to warmer temperatures and a fully developed canopy. A dry event in the early season could, on the other hand, reduce ANPP in these ecosystems, although it seems unlikely that in this period soil moisture will reach threshold values for CO2 fluxes or induce plant wilting. Further research is needed to address the long-term implications of drought on alpine grasslands, but we conclude that timing is an important feature which should be factored in when assessing drought effects on ecosystems.
Data Availability
Data is available as Supplemental Material.
References
Bates D, Maechler M, Bolker B, Walker S. 2015. Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48.
Bates JD, Svejcar T, Miller RF, Angell RA. 2006. The effects of precipitation timing on sagebrush steppe vegetation. J Arid Environ 64:670–697.
Belluau M, Shipley B. 2017. Predicting habitat affinities of herbaceous dicots to soil wetness based on physiological traits of drought tolerance. Ann Bot 119:1073–1084.
Borken W, Matzner E. 2009. Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Glob Chang Biol 15:808–824.
Brændholt A, Larsen KS, Ibrom A, Pilegaard K. 2017. Overestimation of closed-chamber soil CO2 effluxes at low atmospheric turbulence. Biogeosciences 14:1603–1616.
Breinl K, Di Baldassarre G, Mazzoleni M, Lun D, Vico G. 2020. Extreme dry and wet spells face changes in their duration and timing. Environ Res Lett 15:074040.
Chaves MM, Maroco JP, Pereira JS. 2003. Understanding plant responses to drought—from genes to the whole plant. Funct Plant Biol 30:239–264.
Craine JM, Nippert JB, Elmore AJ, Skibbe AM, Hutchinson SL, Brunsell NA. 2012. Timing of climate variability and grassland productivity. PNAS 109:3401–3405.
Darenova E, Holub P, Krupkova L, Pavelka M. 2017. Effect of repeated spring drought and summer heavy rain on managed grassland biomass production and CO2 efflux. J Plant Ecol 10:476–485.
De Boeck HJ, Dreesen FE, Janssens IA, Nijs I. 2010. Climatic characteristics of heat waves and their simulation in plant experiments. Glob Chang Biol 16:1992–2000.
De Boeck HJ, Dreesen FE, Janssens IA, Nijs I. 2011. Whole-system responses of experimental plant communities to climate extremes imposed in different seasons. New Phytol 189:806–817.
De Boeck HJ, Bassin S, Verlinden M, Zeiter M, Hiltbrunner E. 2016. Simulated heat waves affected alpine grassland only in combination with drought. New Phytol 209:531–541.
De Boeck HJ, Hiltbrunner E, Verlinden M, Bassin S, Zeiter M. 2018. Legacy effects of climate extremes in alpine grassland. Front Plant Sci 9:1586.
Denton EM, Dietrich JD, Smith MD, Knapp AK. 2017. Drought timing differentially affects above- and belowground productivity in a mesic grassland. Plant Ecol 218:317–328.
Dietrich JD, Smith MD. 2016. The effect of timing of growing season drought on flowering of a dominant C4 grass. Oecologia 181:391–399.
Erschbamer B, Winkler J. 1995. Shoot and leaf demography of Carex curvula ssp. curvula and Carex curvula ssp. rosae in the Central Alps. J Veg Sci 6:593–598.
Fong Y, Huang Y, Gilbert PB, Permar SR. 2017. chngpt: threshold regression model estimation and inference. BMC Bioinform 18:454–460.
Gilgen AK, Buchmann N. 2009. Response of temperate grasslands at different altitudes to simulated summer drought differed but scaled with annual precipitation. Biogeosci Discuss 6:5217–5250.
Grabherr G. 1989. On community structure in high alpine grasslands. Vegetatio 83(1–2):223–227.
Griffin PC, Hoffmann AA. 2012. Mortality of Australian alpine grasses (Poa spp.) after drought: species differences and ecological patterns. J Plant Ecol 5:121–133.
Hahn C, Lüscher A, Ernst-Hasler S, Suter M, Kahmen A. 2021. Timing of drought in the growing season and strong legacy effects determine the annual productivity of temperate grasses in a changing climate. Biogeosciences 18:585–604.
Hao Y, Zhang H, Biederman JA, Li L, Cui X, Xue K, Jianqing D, Wang Y. 2018. Seasonal timing regulates extreme drought impacts on CO2 and H2O exchanges over semiarid steppes in Inner Mongolia, China. Agric Ecosyst Environ 266:153–166.
Heskel MA, Atkin OK, Turnbull MH, Griffin KL. 2013. Bringing the Kok effect to light: a review on the integration of daytime respiration and net ecosystem exchange. Ecosphere 4:1–14.
Hoover DL, Rogers BM. 2016. Not all droughts are created equal: the impacts of interannual drought pattern and magnitude on grassland carbon cycling. Glob Chang Biol 22:1809–1820.
Hoover DL, Wilcox KR, Young KE. 2018. Experimental droughts with rainout shelters: a methodological review. Ecosphere 9:e02088.
Hothorn T, Bretz F, Westfall P. 2008. Simultaneous inference in general parametric models. Biom J 50:346–363.
Hu G, Gao Q, Ganjurjav H, Wang Z, Luo W, Wu H, Li Y, Yan Y, Gornish SE, Schwartz MW, Wan Y, Li Y. 2021. The divergent impact of phenology change on the productivity of alpine grassland due to different timing of drought on the Tibetan Plateau. Land Degrad Dev 32:4033–4041.
IPCC 2021. Masson-Delmotte V, Zhai P, Pirani A, Connors SL, Péan C, Berger S, Caud N, Chen Y, Goldfarb L, Gomis MI, Huang M, Leitzell K, Lonnoy E, Matthews JBR, Maycock TK, Waterfield T, Yelekçi O, Yu R, Zhou B, editors. Climate change 2021: the physical science basis. Contribution of Working Group I to the sixth assessment report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, in press.
Jonas T, Rixen C, Sturm M, Stoeckli V. 2008. How alpine plant growth is linked to snow cover and climate variability. J Geophys Res Biogeosci 113:G03013.
Körner C. 2021. Alpine plant life: functional plant ecology of high mountain ecosystems. Berlin/Heidelberg: Springer.
Körner C, Hiltbrunner E. 2021. Why is the alpine flora comparatively robust against climatic warming? Diversity 13:383.
Kreyling J, Jentsch A, Beier C. 2014. Beyond realism in climate change experiments: gradient approaches identify thresholds and tipping points. Ecol Lett 17:125-e1.
Kutzbach L, Schneider J, Sachs T, Giebels M, Nykänen H, Shurpali NJ, Martikainen P, Alm J, Wilmking M. 2007. CO2 flux determination by closed-chamber methods can be seriously biased by inappropriate application of linear regression. Biogeosciences 4:1005–1025.
Lambers H, Chapin FS III, Pons TL. 2008. Plant physiological ecology. New York: Springer.
Lemoine NP, Griffin-Nolan RJ, Lock AD, Knapp AK. 2018. Drought timing, not previous drought exposure, determines sensitivity of two shortgrass species to water stress. Oecologia 188:965–975.
Liu H, Mi Z, Lin LI, Wang Y, Zhang Z, Zhang F, Wang H, Liu L, Zhu B, Guangmin C, Zhao X, Sanders NJ, Classen AT, Reich PB, He J-S. 2018. Shifting plant species composition in response to climate change stabilizes grassland primary production. PNAS 115:4051–4056.
Liu P, Zha T, Jia X, Black TA, Jassal RS, Ma J, Bai Y, Wu Y. 2019. Different effects of spring and summer droughts on ecosystem carbon and water exchanges in a semiarid shrubland ecosystem in Northwest China. Ecosystems 22:1869–1885.
Manzoni S, Schimel JP, Porporato A. 2012. Responses of soil microbial communities to water stress: results from a meta-analysis. Ecology 93:930–938.
Meng B, Shi B, Zhong S, Chai H, Li S, Wang Y, Henry HAL, Ma JY, Sun W. 2019. Drought sensitivity of aboveground productivity in Leymus chinensis meadow steppe depends on drought timing. Oecologia 191:685–696.
Noormets A, McNulty SG, DeForest JL, Sun G, Li Q, Chen J. 2008. Drought during canopy development has lasting effect on annual carbon balance in a deciduous temperate forest. New Phytol 179:818–828.
Odum EP. 1984. The mesocosm. BioScience 3:558–562.
Peterson KM, Billings WD. 1982. Growth of alpine plants under controlled drought. Arct Alp Res 14:189–194.
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team. 2020. nlme: linear and nonlinear mixed effects models. R package version 3.1–149.
Poorter H, Bühler J, van Dusschoten D, Climent J, Postma JA. 2012. Pot size matters: a meta-analysis of the effects of rooting volume on plant growth. Funct Plant Biol 39:839–850.
R Core Team. 2020. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Rosbakh S, Leingärtner A, Hoiss B, Krauss J, Steffan-Dewenter I, Poschlod P. 2017. Contrasting effects of extreme drought and snowmelt patterns on mountain plants along an elevation gradient. Front Plant Sci 8:1478.
Schimel JP. 2018. Life in dry soils: effects of drought on soil microbial communities and processes. Annu Rev Ecol Evol Syst 49:409–432.
Schwalm CR, Williams CA, Schaefer K, Baldocchi D, Black TA, Goldstein AH, Law BE, Oechel WC, Paw KTU, Scott RL. 2012. Reduction in carbon uptake during turn of the century drought in western North America. Nat Geosci 5:551–556.
Sippel S, Reichstein M, Ma X, Mahecha MD, Lange H, Flach M, Frank D. 2018. Drought heat and the carbon cycle: a review. Curr Clim Chang Rep 4:266–286.
Slette IJ, Post AK, Awad M, Even T, Punzalan A, Williams S, Smith MD, Knapp AK. 2019. How ecologists define drought, and why we should do better. Glob Chang Biol 25:3193–3200.
Stampfli A, Bloor JM, Fischer M, Zeiter M. 2018. High land-use intensity exacerbates shifts in grassland vegetation composition after severe experimental drought. Glob Chang Biol 24:2021–2034.
Stephan R, Erfurt M, Terzi S, Žun M, Kristan B, Haslinger K, Stahl K. 2021. An inventory of Alpine drought impact reports to explore past droughts in a mountain region. Nat Hazards Earth Syst Sci 21:2485–2501.
Toreti A, Bavera D, Acosta Navarro J, Cammalleri C, de Jager A, Di Ciollo C, Hrast Essenfelder A, Maetens W, Magni D, Masante D, Mazzeschi M, Niemeyer S, Spinoni J. 2022. Drought in Europe August 2022. Luxembourg: Publications Office of the European Union.
van der Molen MK, Dolman AJ, Ciais P, Eglin T, Gobron N, Law BE, Meir P, Peters W, Phillips OL, Reichstein M, Chen T, Dekker SC, Doubkova M, Friedl MA, Jung M, van den Hurk BJJM, de Jeu RAM, Kruijt B, Ohta T, Rebel KT, Plummer S, Seneviratne SI, Sitch S, Teuling AJ, van der Werf GR, Wang G. 2011. Drought and ecosystem carbon cycling. Agric For Meteorol 151:765–773.
van Ruijven J, Berendse F. 2010. Diversity enhances community recovery, but not resistance, after drought. J Ecol 98:81–86.
Vitra A, Deleglise C, Meisser M, Risch AC, Signarbieux C, Lamacque L, Delzon S, Buttler A, Mariotte P. 2019. Responses of plant leaf economic and hydraulic traits mediate the effects of early-and late-season drought on grassland productivity. AoB Plants 11:plz023.
Volaire F. 2018. A unified framework of plant adaptive strategies to drought: crossing scales and disciplines. Glob Chang Biol 24:2929–2938.
von Buttlar J, Zscheischler J, Rammig A, Sippel S, Reichstein M, Knohl A, Jung M, Menzer O, Arain MA, Buchmann N, Cescatti A, Gianelle D, Kiely G, Law BE, Magliulo V, Margolis H, McCaughey H, Merbold L, Migliavacca M, Montagnani L, Oechel W, Pavelka M, Peichl M, Rambal S, Raschi A, Scott RL, Vaccari FP, van Gorsel E, Varlagin A, Wohlfahrt G, Mahecha MD. 2018. Impacts of droughts and extreme-temperature events on gross primary production and ecosystem respiration: a systematic assessment across ecosystems and climate zones. Biogeosciences 15:1293–1318.
Vorkauf M, Kahmen A, Körner C, Hiltbrunner E. 2021. Flowering phenology in alpine grassland strongly responds to shifts in snowmelt but weakly to summer drought. Alp Bot 131:73–88.
Wang Y, Yu S, Wang J. 2007. Biomass-dependent susceptibility to drought in experimental grassland communities. Ecol Lett 10:401–410.
Wang Q, Yang Y, Liu Y, Tong L, Zhang QP, Li J. 2019. Assessing the impacts of drought on grassland net primary production at the global scale. Sci Rep 9:1–8.
Williams CJ, McNamara JP, Chandler DG. 2009. Controls on the temporal and spatial variability of soil moisture in a mountainous landscape: the signature of snow and complex terrain. Hydrol Earth Syst Sci 13:1325–1336.
Winkler DE, Chapin KJ, Kueppers LM. 2016. Soil moisture mediates alpine life form and community productivity responses to warming. Ecology 97:1553–1563.
Yang Z, Wei Y, Fu G, Song H, Li G, Xiao R. 2020. Asymmetric effect of increased and decreased precipitation in different periods on soil and heterotrophic respiration in a semiarid grassland. Agric For Meteorol 291:108039.
Zeileis A, Hothorn T. 2002. Diagnostic checking in regression relationships. R News 2:7–10.
Zhang F, Quan Q, Ma F, Tian D, Zhou Q, Niu S. 2019a. Differential responses of ecosystem carbon flux components to experimental precipitation gradient in an alpine meadow. Funct Ecol 33:889–900.
Zhang H, Yu H, Zhou C, Zhao H, Qian X. 2019b. Aboveground net primary productivity not CO2 exchange remain stable under three timing of extreme drought in a semi-arid steppe. PLoS ONE 14:e0214418.
Zhang J, Miao Y, Zhang T, Wei Y, Qiao X, Miao R, Wang D, Yang Z. 2020a. Drought timing and primary productivity in a semiarid grassland. Land Degrad Dev 31:2185–2195.
Zhang R, Zhao X, Zuo X, Degen AA, Li Y, Liu X, Luo Y, Qu H, Lian J, Wang R. 2020b. Drought-induced shift from a carbon sink to a carbon source in the grasslands of Inner Mongolia, China. Catena 195:104845.
Zhao T, Zhang Y, Zhang T, Xu M, Zhu J, He Y, Yu G. 2022. Drought occurrence and time-dominated variations in water use efficiency in an alpine meadow on the Tibetan Plateau. Ecohydrology 15:e2360.
Acknowledgements
We thank the Stelvio National Park for authorizing research activity at the field site. We thank G. Andreoli, F. Arici, T. L. Bacchilega, M. Boschetti, P. Chiudinelli, M. Gaudenzi, M. M. Maniscalco and M. Timco for support in the field work. We also thank the anonymous reviewers of earlier drafts of this manuscript for their helpful comments towards the manuscript improvement. We thank Rebecca Whittingham for the English revision of the manuscript.
Funding
Open access funding provided by Università degli Studi di Parma within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest
Additional information
Author Contributions: All the authors planned the research and participated in the experimental set-up and data collection. TGWF and MC performed data analyses. TGWF led the writing of the manuscript, and all authors contributed to revisions.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Forte, T.G., Carbognani, M., Chiari, G. et al. Drought Timing Modulates Soil Moisture Thresholds for CO2 Fluxes and Vegetation Responses in an Experimental Alpine Grassland. Ecosystems 26, 1275–1289 (2023). https://doi.org/10.1007/s10021-023-00831-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-023-00831-7