
Abstract
Sodium–glucose cotransporter 2 (SGLT2) occurs in the proximal renal tubule cells. We investigate the hepatic expression of SGLT2 and its related factors in patients with chronic liver disease. This is a retrospective human study. The liver tissues were biopsied from patients with chronic liver disease (n = 30). The expression levels of SGLT2 were evaluated by immunostaining. Furthermore, the undirected graphical model was used to identify factors associated with hepatic expression levels of SGLT2. The SGLT2 expression was observed in not only the kidney, but also the liver in immunostaining (SGLT2 intensity: kidney 165.8 ± 15.6, liver 114.4 ± 49.0 arbitrary units, P < 0.01) and immunoblotting. There was no significant difference in hepatic expression of SGLT2 in the stratified analysis according to age, sex, BMI, and the severity of the liver disease. In the undirected graphical model, SGLT2 directly interacted with various factors such as sex, fatty change, neutrophil-to-lymphocyte ratio, triglyceride, hemoglobin A1c, creatinine, and albumin (partial correlation coefficient 0.4–0.6 for sex and 0.2–0.4 for others). The expression of SGLT2 was observed in the hepatocytes of patients with chronic liver disease. The undirected graphical model demonstrated the complex interaction of hepatic expression levels of SGLT2 with gender, inflammation, renal function, and lipid/glucose/protein metabolisms.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sodium/glucose cotransporter 2 (SGLT2) is a subtype of SGLT, which mediates glucose uptake across apical cell membranes [1]. SGLT2 occurs in the luminal membrane of proximal renal tubule cells and is responsible for more than 90% of glucose reabsorption from the glomerular ultrafiltrate in normoglycemia [1]. Renal protein expression of SGLT2 is increased by 40–80% in animal models of diabetes mellitus [2]. High messenger ribonucleic acid (mRNA) and protein expressions of SGLT2 are also observed in proximal renal tubule cells isolated from patients with diabetes mellitus than in cells from healthy individuals [3]. Thus, renal SGLT2 plays a crucial role in glucose metabolism.
SGLT2 is also known to play an important role in fatty acid metabolism [4]. The liver is the central organ involved in fatty acid metabolism. The liver stores fatty acids as triglycerides in hepatocytes and metabolizes fatty acids through beta-oxidation in the mitochondria of hepatocytes [5, 6]. The expression of SGLT2 has been observed in various human hepatoma cell lines [7, 8]. In hepatoma cells, SGLT2 uptakes glucose into cells. In addition, we previously reported that SGLT2 occurs in the mitochondria and regulates fatty acid metabolism in hepatoma cells [8]. However, the expression of SGLT2 and factors associated with hepatic SGLT2 expression remain unclear in humans. Extra-renal expression of SLGT2 has been investigated and its mRNA expression has been observed in various tissues including the heart and spleen in vitro and animal studies [9, 10]. In addition, protein expression of SGLT2 has been reported to occur in patients with traumatic brain injury [11]. These previous findings suggest the possible protein expression of SGLT2 in various tissues including the heart, spleen, and brain. However, the protein expression of SGLT2 in these tissues remains unclear in humans [8, 12,13,14].
This study aimed to investigate the expression of SGLT2 in patients with chronic liver disease. We also examined the extra-renal expression of SGLT2 in the human heart, spleen, and brain tissues in humans. We further investigated the direct factors associated with the hepatic expression level of SGLT2 in patients with chronic liver disease.
Subjects and methods
Study design and ethics
This is a retrospective study. The study protocol conformed to the ethical guidelines of the 1975 Declaration of Helsinki, as reflected by the prior approval of the Institutional Review Board of Kurume University School of Medicine (#21,104). An opt-out approach was used to obtain informed consent from the patients, and personal information was protected during data collection.
Materials
The reagents were purchased from Cosmo Bio Co., Ltd. (Tokyo, Japan) unless otherwise indicated. Antibodies against SGLT2 were purchased from Proteintech Group, Inc. (24,654-1-AP, Rosemont, IL).
Human tissues
The liver tissues were obtained from patients with chronic liver disease who underwent liver biopsy (n = 30). The kidney, heart, spleen, and brain tissues were obtained from necropsy patients with no impairment in each organ (each n = 5).
Data collection
The variables were retrospectively reviewed using medical records. The following data were collected at the time of liver biopsy: age, sex, body mass index (BMI), red blood cell count, hemoglobin level, white blood cell count, neutrophil and lymphocyte ratio (NLR), platelet count, fasting blood glucose level, hemoglobin A1c (HbA1c) level, and serum levels of aspartate aminotransferase (AST), alanine aminotransferase (ALT), lactate dehydrogenase, gamma-glutamyl transpeptidase (GGT), alkaline phosphatase, cholinesterase, albumin, total bilirubin, prothrombin activity, total cholesterol, high-density lipoprotein (HDL)-cholesterol, triglyceride, blood urea nitrogen, and creatinine. We also calculated FIB-4 Index. In addition, the presence of hepatic steatosis and liver cirrhosis were evaluated by histological findings assessed by a board-certified pathologist (J.A.).
Immunohistochemistry for SGLT2
Liver specimens were fixed in 10% formalin and embedded in paraffin. Consecutive 4-μm sections were stained with hematoxylin. Immunohistochemical staining was performed on paraffin-embedded tissues. Immunohistochemical staining was performed using SGLT2 (PGI 24,654-1-AP, Proteintech, Tokyo, Japan) and BOND-III (Leica Microsystems, Newcastle, UK). Proximal renal tubule cells of the kidney were used as a positive control for SGLT2.
Image acquisition and processing
All samples were scanned at 40 × magnification using the Nano Zoomer (Hamamatsu Photonics K.K. Hamamatsu, Japan), which is a slide scanning system [15]. All images were saved in Tagged Image File Format files and processed using the FIJI software (version 2.0.0-rc-49/1.51d). Then, SGLT2 staining was separated from nuclear staining according to the color tone by using a color deconvolution plugin in FIJI [16]. This plugin separates the histological stains in the input image into different channels by applying the built-in H-3,3'-diaminobenzidine (DAB) vector to generate two images: color 1 (hematoxylin) and color 2 (DAB [SGLT2]). The SGLT2 channel was converted into an 8-bit grayscale image. Identical operations were applied to each image as follows: the background was subtracted by binarization. Binarization was performed using the default settings for the local thresholding algorithms (i.e. radius = 15 pixels, parameter 1 = 0, and parameter 2 = 0) [17].
Measurement of SGLT2 staining intensity in the tissue
We measured the SGLT2 staining intensity by the oval region of interest. Three independent researchers measured the intensity thrice times in a tissue image with no clinical information (D.N., T.T., and T.K). The mean intensity of the nine measurements was used in the analysis.
Classification of age and BMI
We classified all patients into the younger and older groups, or low and high BMI groups according to the median age (median age, 69.5 [59.0–79.3] years) and BMI (median 23.6, [20.9–24.9] years), respectively.
Classification of the high and low SGLT2 groups
We classified all patients into the high and low SGLT2 groups according to the median hepatic expression level of SGLT2 (median SGLT2, 114 [73.2 -162.5]).
Immunoblotting analysis
We examined the expression of SGLT2 by immunoblotting using kidney, normal liver, heart, spleen, and brain lysates. These lysates were purchased from Pro Sci inc (PSC 1706-02, 1718-01, 1301, 1306, and 1731-01, Poway, CA).
Immunoblotting was performed using the anti-SGLT2 antibody (sc-393350, Santa Cruz Biotechnology, Dallas, TX) as previously described [8].
An undirected graphical model
An undirected graphical model was employed to explore the complex interaction between SGLT2 and the variables in graphs as previously described [18]. Two variables were connected by a line if the partial correlation coefficient was ≥0.2. An undirected graphical model was performed using JUSE-Stat Works/V5 (The Institute of Japanese Union of Scientists & Engineers, Tokyo, Japan).
Statistics
Data are expressed as numbers or means ± standard errors. Differences between the two groups were analyzed using the Mann–Whitney U test. All statistical analyses were conducted by a biostatistician (M.K.).
Results
Patients’ characteristics
The patients’ characteristics are summarized in Table 1. The median age was 69.5 years and the male ratio was 66.7% (Table 1). The median BMI was 23.6 kg/m2. Hepatitis C virus was the major etiology of chronic liver disease followed by hepatitis B virus and non-virus. The prevalence of fatty liver and liver cirrhosis was 10.0% and 36.7% of all subjects, respectively (Table1).
In the biochemical examination, the median serum levels of AST, ALT, alkaline phosphatase, and GGT were within reference values. However, the median level of albumin and platelet count was lower than the reference value and the median FIB-4 index was 3.0. The median levels of blood urea nitrogen, creatine, estimated glomerular filtration rate, HbA1c, HDL cholesterol, and triglyceride were within the reference values.
Expression of SGLT2 on human tissues
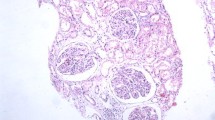
In all tissues, a positive signal was not detected in the negative control examination, which lacks a primary antibody for SGLT2. Kidney tissue was used as the positive control. The expression of SGLT2 was observed in the proximal renal tubule cells and the expression level was significantly higher than that in the negative control (Fig. 1A).


Expression of SGLT2 in human tissues evaluated by immunostaining. Blue indicates mitochondrial hematoxylin. Yellow indicates SGLT2. A Kidney (positive control), B liver, C heart, D spleen, E brain
In the liver, expression of SGLT2 was also observed. SGLT2 was found in the cytoplasm of hepatocytes. There was no lobular heterogeneity in SGLT2 expression. In addition, expression of SGLT2 was detected also at the apical membrane of bile duct cells (Fig. 1B). The expression levels in hepatocytes and bile duct cells were significantly higher than those in the negative controls (Fig. 1B).
In the heart tissue, expression of SGLT2 was found in the cytoplasm of the cardiomyocytes. In the spleen tissue, expression of SGLT2 also was found in the splenic cord. Weak expression of SGLT2 was observed in the brain tissue. The expression level of these tissues was significantly higher than the negative controls (Fig. 1C, D, E).
Comparison of the expression level of SGLT2 in tissues
In immunostaining, the expression of SGLT2 differed with the tissues. Besides proximal renal tubule cells used as a positive control, a higher expression level of SGLT2 was seen in the liver. Lower and very low expression levels of SGLT2 were seen in the spleen and the brain (Fig. 2).

The difference in SGLT2 expression levels in the kidney, liver, heart, spleen, and brain tissues of humans
Expression of SGLT2 in the lysate of human normal tissues
In immunoblotting, expression of SGLT2 was observed in the kidney, liver, heart, spleen, and brain. Similar to the results of immunohistochemistry, a higher expression level of SGLT2 was seen in the liver1 (Fig. 3).

The expression of SGLT2 in the kidney, liver, heart, spleen, and brain was evaluated by immunoblotting. Jurkat cell is positive control of SGLT2 expression
The hepatic expression level of SGLT2 in the stratification analyses according to the severity of age, sex, BMI, and hepatic fibrosis
We classified all patients into the younger and older groups, or low and high BMI groups according to the median age (median age, 69.5 [59.0 -79.3] years) and BMI (median 23.6, [20.9–24.9] years), respectively. There was no significant difference in hepatic expression of SGLT2 in the stratified analysis according to age, sex, and BMI (Fig. 4A, B, C). No significant difference was also observed in the expression levels of SGLT2 between patients with chronic hepatitis and liver cirrhosis (Fig. 4D).

Differences in hepatic expression levels of SGLT2 according to A sex, B age, C BMI, and D severity of liver disease
Associations between the hepatic expression level of SGLT2 and biochemical parameters
We classified all patients into high and low SGLT2 groups according to the median hepatic expression level of SGLT2. There was no significant difference in the serum levels of AST, ALT, and GGT between the High and Low SGLT2 groups (Figs. 5A, B, C). No significant differences were also observed in HbA1c, triglyceride, prothrombin activity, and NLR levels between the high and low SGLT2 groups (Figs. 5E, F, G).

Differences in biochemical parameters between the high and Low SGLT2 groups. A AST, B ALT, C GGT, D HbA1c, E triglycerides, F prothrombin activity, and G NLR. AST Aspartate aminotransferase, ALT alanine aminotransferase, GGT gamma-glutamyl transferase, HbA1c hemoglobin A1c, TG triglyceride; NLR: Neutrophil /lymphocyte ratio
Factors associated with the expression of SGLT2 analyzed by an undirected graphical model
We employed an undirected graphical model to reveal the direct interaction between SGLT2 and various factors. In the undirected graphical model, SGLT2 directly interacted with seven factors including sex, fatty change, triglyceride, HbA1c, NLR, creatinine, and albumin. Of these factors, SGLT2 showed the strongest interaction with sex (Fig. 6).

Undirected graphical model. The thick line, thin line, and dotted line indicate partial correlation coefficients of 0.6–1.0, 0.4–0.6, and 0.2–0.4, respectively. SGLT2 sodium–glucose cotransporter 2; Alb albumin, TG triglyceride, Cr creatinine, HbA1c hemoglobin A1c, T-Bil total bilirubin, BMI body mass index, GGT gamma-glutamyl transferase, BS blood sugar, CH/LC chronic hepatitis/liver cirrhosis, FIB-4 fibrosis-4 index, HDL high-density lipoprotein cholesterol
Discussion
In this study, we demonstrated that the expression of SGLT2 was observed in not only in the kidney, but also in the liver, heart, spleen, and brain. Of these organs, the hepatic expression of SGLT2 was the second -highest after the renal expression. In the undirected graphical model, SGLT2 directly interacted with various factors such as sex, fatty change, triglyceride, HbA1c, NLR, creatinine, and albumin.
The kidney was used as a positive control for immunostaining of SGLT2, and we reported that SGLT2 was found in the proximal renal tubule cells in this study. In rats, the SGLT2 gene is detected in the outer medulla of the kidney [19]. In addition, Rahmoune et al. reported the expression of SGLT2 in human proximal renal tubular cells isolated from the urine of patients with diabetes mellitus [3]. Moreover, Vrhovac et al. reported that the expression of SGLT2 is observed in the S1 and S2 segments of the human proximal tubule renal tissue [20]. Our results were in good agreement with previous reports. Thus, our immunostaining protocol is thought to be suitable to detect the protein expression of SGLT2 in human tissues.
In this study, we first demonstrated the expression of SGLT2 in the human liver. Expression of SGLT2 in the liver was second highest after that in the kidney. SGLT2 is thought to be found only in proximal renal tubule cells [19]. However, several recent studies have revealed the extra-renal expression of SGLT2. Bolla et al. reported that the expression of SGLT2 was observed in human duodenal mucosa in patients with diabetes mellitus [21]. Moreover, it is reported that various cancers have the expression of SGLT2 including pancreatic cancer, prostate cancer, and thyroid cancer [22, 23]. The expression of SGLT2 has also been observed in various human hepatoma cell lines [7, 8]. We also previously reported that SGLT2 is found in the mitochondria of Hep3B cells, regulating and regulating metabolic reprogramming and cell proliferation [8]. Furthermore, Zhao et al. reported that mRNA expression of SGLT2 was observed in the liver of cattle and the expression level was second highest after that in the kidney [9]. Moreover, Hasegawa et al. demonstrated that the accumulation level of luseogliflozin which binds to SGLT2 was second highest in the liver after that in the kidney in rats [24]. These previous studies support our findings that the expression of SGLT2 was found in human hepatocytes.
There was no significant difference in the hepatic expression of SGLT2 in terms of patient characteristics and biochemical parameters in our study. However, in the undirected graphical model, SGLT2 directly interacted with sex, fatty change, triglycerides, HbA1c, NLR, creatinine, and albumin. Of these factors, sex interacted the most with the expression level of SGLT2. An advantage of undirected graphical model analysis is bi-directional assessments and our results suggest the following two possibilities (1) sex was associated with SGLT2 expression or (2) SGLT2 expression was associated with sex. There was no significant difference in SGLT2 expression levels between men and women. This finding indicated that possibility 1 was denied and SGLT2 expression was associated with sex (possibility 2). To verify the possibility 2, we performed a sub-analysis and all subjects were classified into the lower or higher SGLT2 groups according to the cutoff value of 164.4, which was based on the receiver operating characteristic curve. In the higher SGLT2 group, the prevalence of males was significantly higher than in the lower SGLT2 groups (Supplementary figure).
Sabolic et al. reported that, in mice, the expression level of SGLT2 was male-dominant [25]. These sex differences may be related to testosterone levels. Sodium gradient-driven intravascular uptake of glucose was 2.5-fold higher in male mice than in female mice and testosterone treatment for females raised the levels to that in males [26]. Thus, these basic studies support our finding of an interaction between the hepatic expression of SGLT2 and sex in humans.
Moreover, Singh AK et al. performed a meta-analysis and reported the effect of SGLT2i was different in cardiovascular outcomes by gender [27, 28]. On the other hand, it is reported that SGLT2i adverse events including urinary tract/genital infection occur more frequently in women than men [29]. Thus, several previous studies reported gender differences in the effects of SGLT2 inhibitors.
We also reported that SGLT2 was found in the apical membrane of bile duct cells, cardiomyocytes, and splenic cells. Very low expression of SGLT2 was observed in the brain. We could not examine the detail of these expressions in this study. However, SGLT2 mRNA has been reported to be detected in the liver, cardiomyocytes, and spleen [9, 10]. Bile duct cells are known to reabsorb glucose from bile into the blood [30], suggesting the possible involvement of SGLT2. In addition, Ng et al. reported that the SGLT2 gene is observed in cardiomyocytes and is down-regulated by SGLT2 inhibitors, suggesting direct effects of SGLT2 inhibitors on cardiomyocytes [10]. Moreover, Hawley et al. reported that oral administration of SGLT2 inhibitor activates adenosine monophosphate-activated protein kinase in the spleen of mice [31]. Expression of SGLT2 has been reported to occur in patients with traumatic brain injury to increase glucose uptake increases in injured brain cells [11]. Thus, recent studies and our results suggest that SGLT2 may occur in various types of tissues.
This study has several limitations. First, this study was conducted in a single center with a small sample size. Second, all liver tissue samples were obtained from patients with chronic liver disease. Therefore, the expression of SGLT2 in normal human liver tissue remains unclear. Third, all samples were taken from Japanese individuals; therefore, racial differences in the expression of SGLT2 remain unclear. Accordingly, further study will be designed as an international multi-center study that includes a large number of samples with normal liver tissue specimens obtained by hepatic resection for metastatic liver tumors.
Conclusion
In this study, we demonstrated that SGLT2 occurred not only in the kidney but also in the liver, heart, spleen, and brain. Expression of SGLT2 in the liver was second highest after that in the kidney. The undirected graphical model revealed the complex interaction of hepatic expression levels of SGLT2 with various factors such as gender, inflammation, renal function, and lipid/glucose/protein metabolisms.
References
Poulsen SB, Fenton RA, Rieg T (2015) Sodium–glucose cotransport. Curr Opin Nephrol Hypertens 24:463–469
Novikov A, Vallon V (2016) Sodium glucose cotransporter 2 inhibition in the diabetic kidney: an update. Curr Opin Nephrol Hypertens 25:50–58
Rahmoune H, Thompson PW, Ward JM, Smith CD, Hong G, Brown J (2005) Glucose transporters in human renal proximal tubular cells isolated from the urine of patients with non-insulin-dependent diabetes. Diabetes 54:3427–3434
Kawarasaki S, Sawazaki H, Iijima H, Ng SP, Kwon J, Mohri S, Iwase M, Jheng HF, Takahashi H, Nomura W, Inoue K, Kawada T, Goto T (2020) comparative analysis of the preventive effects of canagliflozin, a sodium–glucose co-transporter-2 inhibitor, on body weight gain between oral gavage and dietary administration by focusing on fatty acid metabolism. Diabetes Metab Syndr Obes 13:4353–4359
Jones JG (2021) Non-invasive analysis of human liver metabolism by magnetic resonance spectroscopy. Metabolites 11(11):751
Auger C, Alhasawi A, Contavadoo M, Appanna VD (2015) Dysfunctional mitochondrial bioenergetics and the pathogenesis of hepatic disorders. Front Cell Dev Biol 3:40
Kaji K, Nishimura N, Seki K, Sato S, Saikawa S, Nakanishi K, Furukawa M, Kawaratani H, Kitade M, Moriya K, Namisaki T, Yoshiji H (2018) Sodium glucose cotransporter 2 inhibitor canagliflozin attenuates liver cancer cell growth and angiogenic activity by inhibiting glucose uptake. Int J Cancer 142:1712–1722
Nakano D, Kawaguchi T, Iwamoto H, Hayakawa M, Koga H, Torimura T (2020) Effects of canagliflozin on growth and metabolic reprograming in hepatocellular carcinoma cells: multi-omics analysis of metabolomics and absolute quantification proteomics (iMPAQT). PLoS ONE 15:e0232283
Zhao FQ, McFadden TB, Wall EH, Dong B, Zheng YC (2005) Cloning and expression of bovine sodium/glucose cotransporter SGLT2. J Dairy Sci 88:2738–2748
Ng KM, Lau YM, Dhandhania V, Cai ZJ, Lee YK, Lai WH, Tse HF, Siu CW (2018) Empagliflozin ammeliorates high glucose induced-cardiac dysfuntion in human ipsc-derived cardiomyocytes. Sci Rep 8:14872
Oerter S, Forster C, Bohnert M (2019) Validation of sodium/glucose cotransporter proteins in human brain as a potential marker for temporal narrowing of the trauma formation. Int J Legal Med 133:1107–1114
Verma S, McMurray JJV (2018) SGLT2 inhibitors and mechanisms of cardiovascular benefit: a state-of-the-art review. Diabetologia 61:2108–2117
Shaffner J, Chen B, Malhotra DK, Dworkin LD, Gong R (2021) Therapeutic targeting of SGLT2: A New Era in the treatment of diabetes and diabetic kidney disease. Front Endocrinol 12:749010
Fadini GP, Bonora BM, Zatti G, Vitturi N, Iori E, Marescotti MC, Albiero M, Avogaro A (2017) Effects of the SGLT2 inhibitor dapagliflozin on HDL cholesterol, particle size, and cholesterol efflux capacity in patients with type 2 diabetes: a randomized placebo-controlled trial. Cardiovasc Diabetol 16:42
Zheng PP, van der Weiden M, Kros JM (2014) Fast tracking of co-localization of multiple markers by using the nanozoomer slide scanner and NDPViewer. J Cell Physiol 229:967–973
Landini G, Martinelli G, Piccinini F (2021) Colour deconvolution: stain unmixing in histological imaging. Bioinformatics 37:1485–1487
Healy S, McMahon J, Owens P, Dockery P, FitzGerald U (2018) Threshold-based segmentation of fluorescent and chromogenic images of microglia, astrocytes and oligodendrocytes in FIJI. J Neurosci Methods 295:87–103
Choi H, Gim J, Won S, Kim YJ, Kwon S, Park C (2017) Network analysis for count data with excess zeros. BMC Genet 18:93
Wright EM (2001) Renal Na(+)-glucose cotransporters. Am J Physiol Renal Physiol 280:F10–F18
Vrhovac I, BalenEror D, Klessen D, Burger C, Breljak D, Kraus O, Radovic N, Jadrijevic S, Aleksic I, Walles T, Sauvant C, Sabolic I, Koepsell H (2015) Localizations of Na(+)-D-glucose cotransporters SGLT1 and SGLT2 in human kidney and of SGLT1 in human small intestine, liver, lung, and heart. Pflugers Arch 467:1881–1898
Bolla AM, Butera E, Pellegrini S, Caretto A, Bonfanti R, Zuppardo RA, Barera G, Cavestro GM, Sordi V, Bosi E (2020) Expression of glucose transporters in duodenal mucosa of patients with type 1 diabetes. Acta Diabetol 57:1367–1373
Scafoglio C, Hirayama BA, Kepe V, Liu J, Ghezzi C, Satyamurthy N, Moatamed NA, Huang J, Koepsell H, Barrio JR, Wright EM (2015) Functional expression of sodium–glucose transporters in cancer. Proc Natl Acad Sci U S A 112:E4111–E4119
Wang Y, Yang L, Mao L, Zhang L, Zhu Y, Xu Y, Cheng Y, Sun R, Zhang Y, Ke J, Zhao D (2022) SGLT2 inhibition restrains thyroid cancer growth via G1/S phase transition arrest and apoptosis mediated by DNA damage response signaling pathways. Cancer Cell Int 22:74
Hasegawa M, Chino Y, Horiuchi N, Hachiuma K, Ishida M, Fukasawa Y, Nakai Y, Yamaguchi J (2015) Preclinical metabolism and disposition of luseogliflozin, a novel antihyperglycemic agent. Xenobiotica 45:1105–1115
Sabolic I, Vrhovac I, Eror DB, Gerasimova M, Rose M, Breljak D, Ljubojevic M, Brzica H, Sebastiani A, Thal SC, Sauvant C, Kipp H, Vallon V, Koepsell H (2012) Expression of Na+-D-glucose cotransporter SGLT2 in rodents is kidney-specific and exhibits sex and species differences. Am J Physiol Cell Physiol 302:C1174–C1188
Mackovic M, Zimolo Z, Burckhardt G, Sabolic I (1986) Isolation of renal brush-border membrane vesicles by a low-speed centrifugation; effect of sex hormones on Na+-H+ exchange in rat and mouse kidney. Biochim Biophys Acta 862:141–152
Singh AK, Singh R (2020) Gender difference in cardiovascular outcomes with SGLT-2 inhibitors and GLP-1 receptor agonist in type 2 diabetes: a systematic review and meta-analysis of cardio-vascular outcome trials. Diabetes Metab Syndr 14:181–187
Singh AK, Singh R (2020) Reply to Sodium–glucose co-transporter-2 inhibitors, cardiovascular outcomes and the impact of gender: class effect or statistical play of chance? Diabetes Metab Syndr 14:335
Joung KI, Jung GW, Park HH, Lee H, Park SH, Shin JY (2020) Gender differences in adverse event reports associated with antidiabetic drugs. Sci Rep 10:17545
Masyuk AI, Masyuk TV, Tietz PS, Splinter PL, LaRusso NF (2002) Intrahepatic bile ducts transport water in response to absorbed glucose. Am J Physiol Cell Physiol 283:C785–C791
Hawley SA, Ford RJ, Smith BK, Gowans GJ, Mancini SJ, Pitt RD, Day EA, Salt IP, Steinberg GR, Hardie DG (2016) The Na+/glucose cotransporter inhibitor canagliflozin activates AMPK by Inhibiting mitochondrial function and increasing cellular AMP levels. Diabetes 65:2784–2794
Acknowledgements
We are grateful to Masako Hayakawa, Akihiko Kawahara, and Takato Kumagae for excellent technical assistance.
Funding
This work was supported by the Japan Society for the Promotion of Science (JSPS) Grant-in-Aid for Young Scientists (B) JP19K17446.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests
Takumi Kawaguchi received lecture fees from Janssen Pharmaceutical K.K., Mitsubishi Tanabe Pharma Corporation, and Otsuka Pharmaceutical Co. Ltd. The other authors have no conflicts of interest.
Ethical approval
This protocol conformed to the ethical guidelines of the 1975 Declaration of Helsinki, as reflected by the prior approval of the institutional review board of Kurume University School of Medicine (#21104). An opt-out approach was used to obtain informed consent from the patients, and personal information was protected during data collection.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nakano, D., Akiba, J., Tsutsumi, T. et al. Hepatic expression of sodium–glucose cotransporter 2 (SGLT2) in patients with chronic liver disease. Med Mol Morphol 55, 304–315 (2022). https://doi.org/10.1007/s00795-022-00334-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00795-022-00334-9